
Stop Codon Context-Specific Induction of Translational Readthrough


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. DNA Constructs and Cloning
2.2. Cell Culture
2.3. Flow Cytometry-Based Dual Reporter Assay and Translational Readthrough Calculation
2.4. Statistics
3. Results
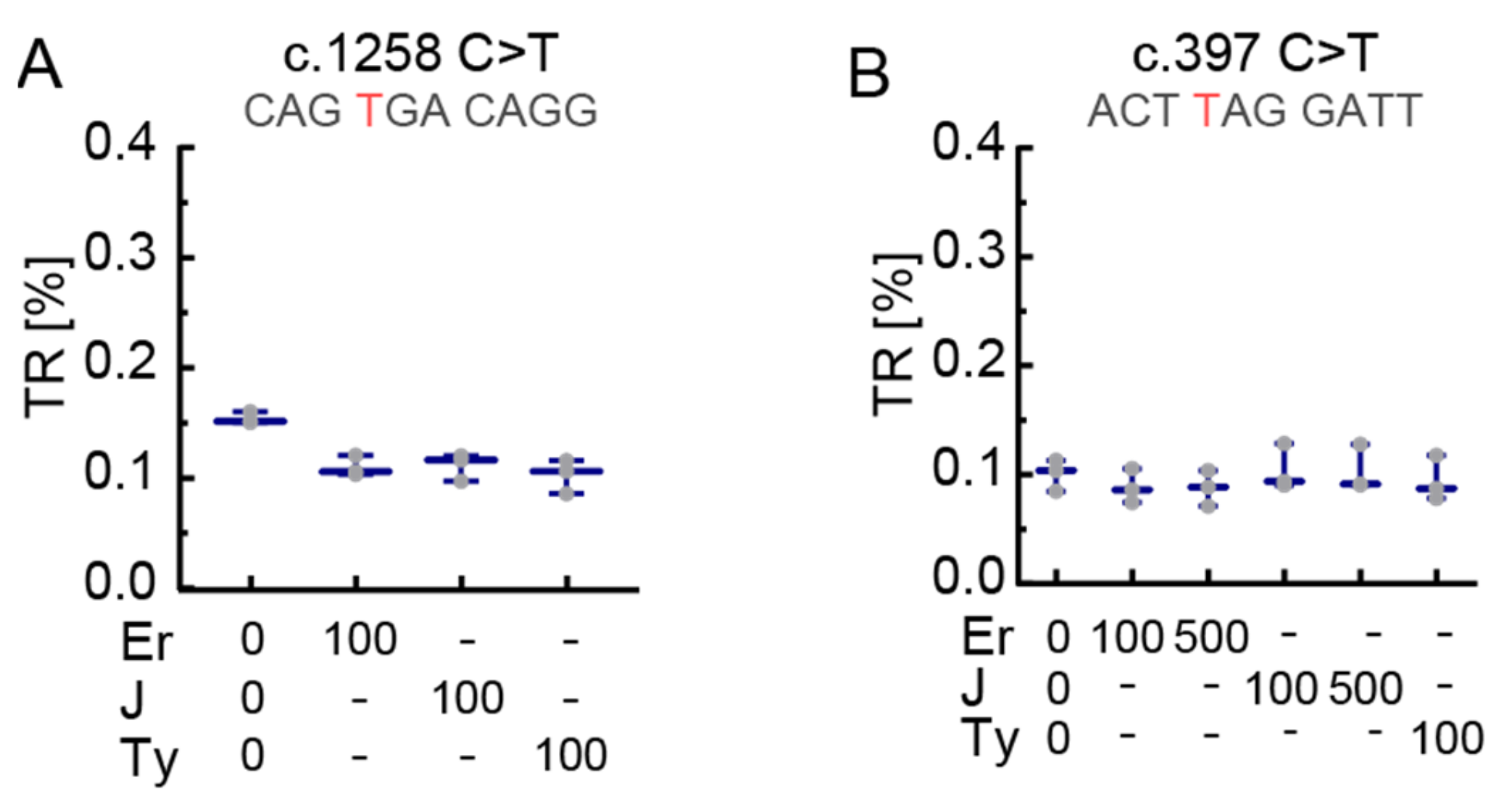
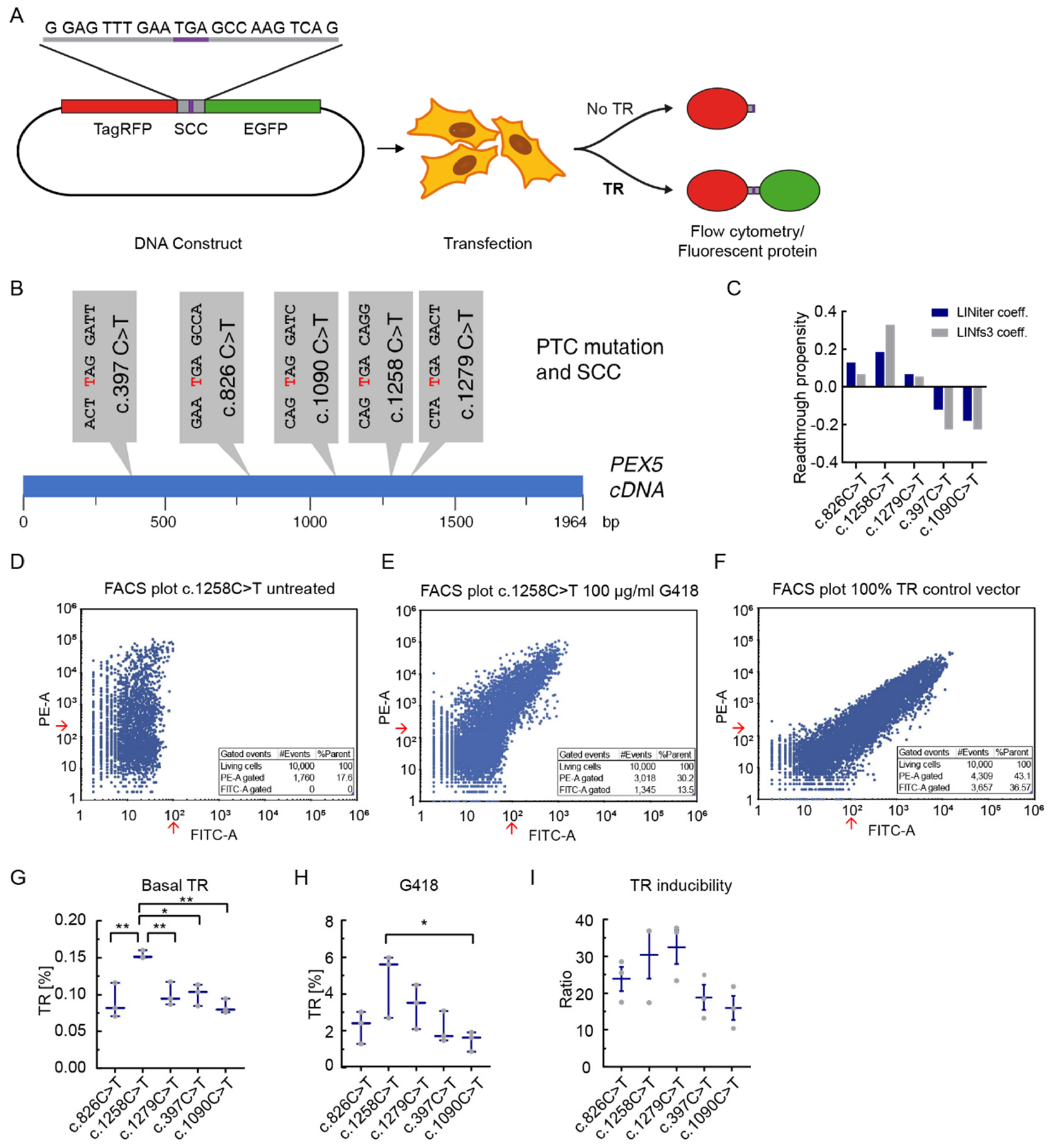
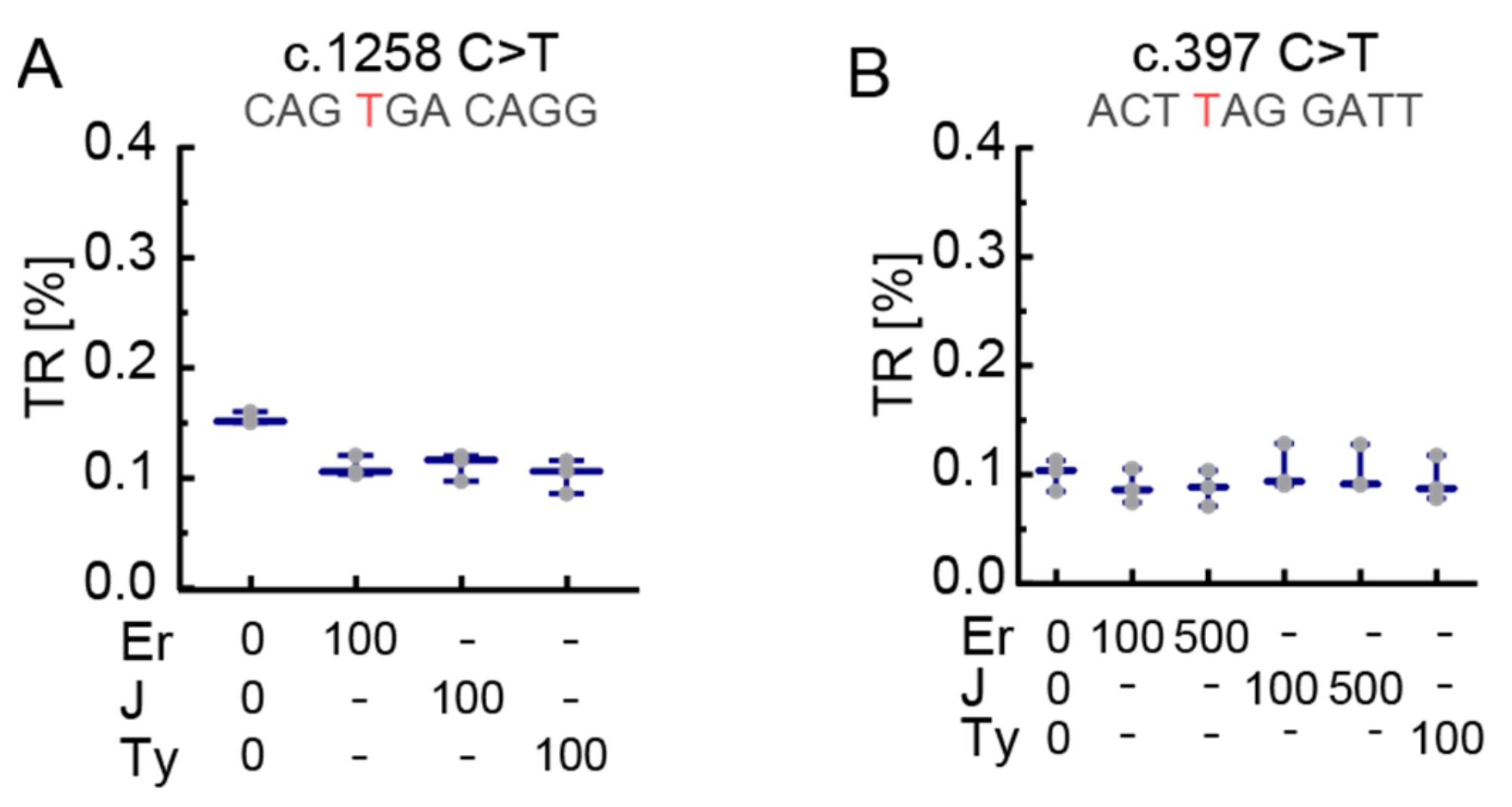
3.1. The Stop Codon Context Affects Basal Translational Readthrough and Induced Translational Readthrough in PEX5 Nonsense Mutations
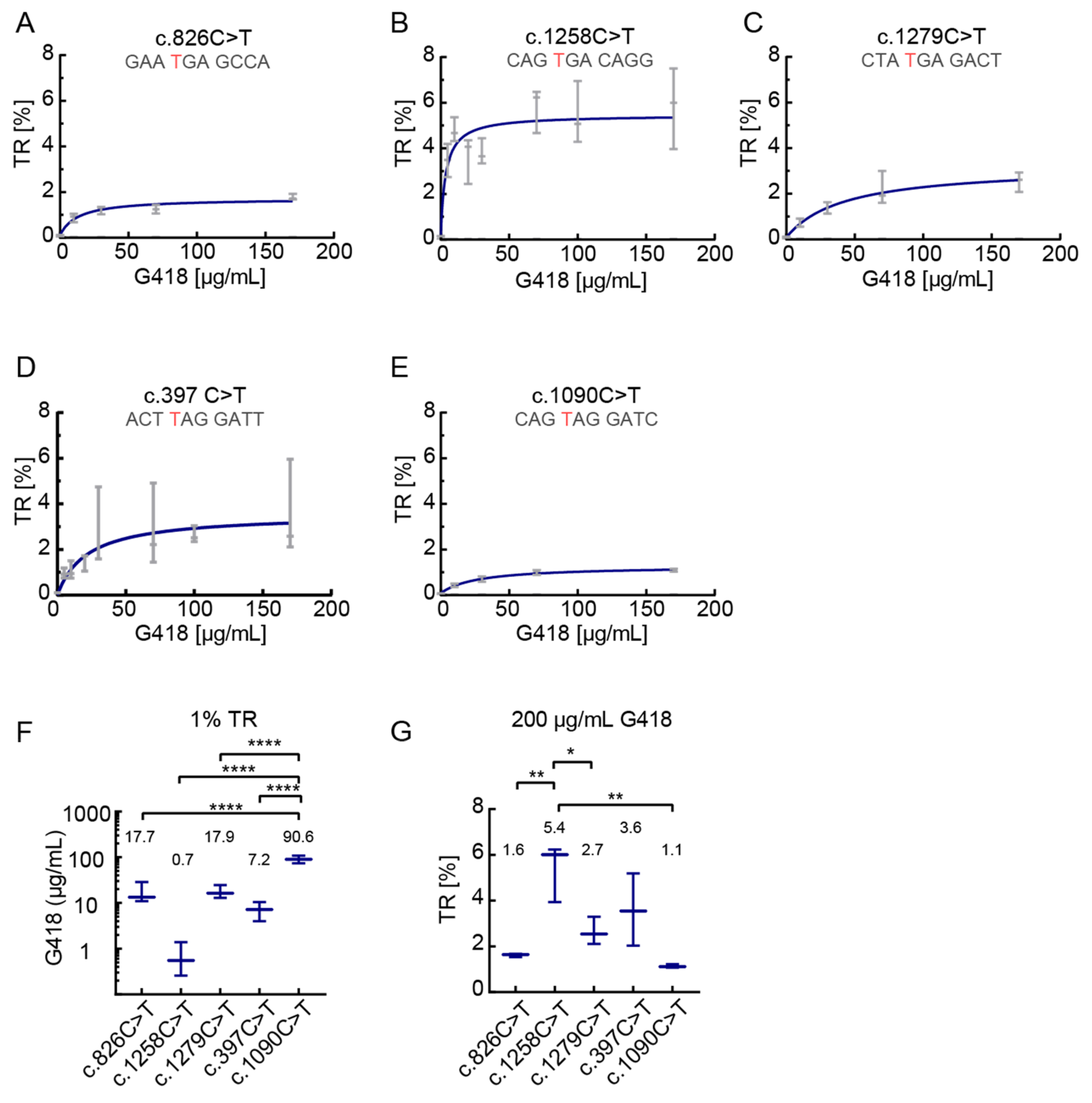
3.2. Dose-Dependent Induction of Translational Readthrough by G418
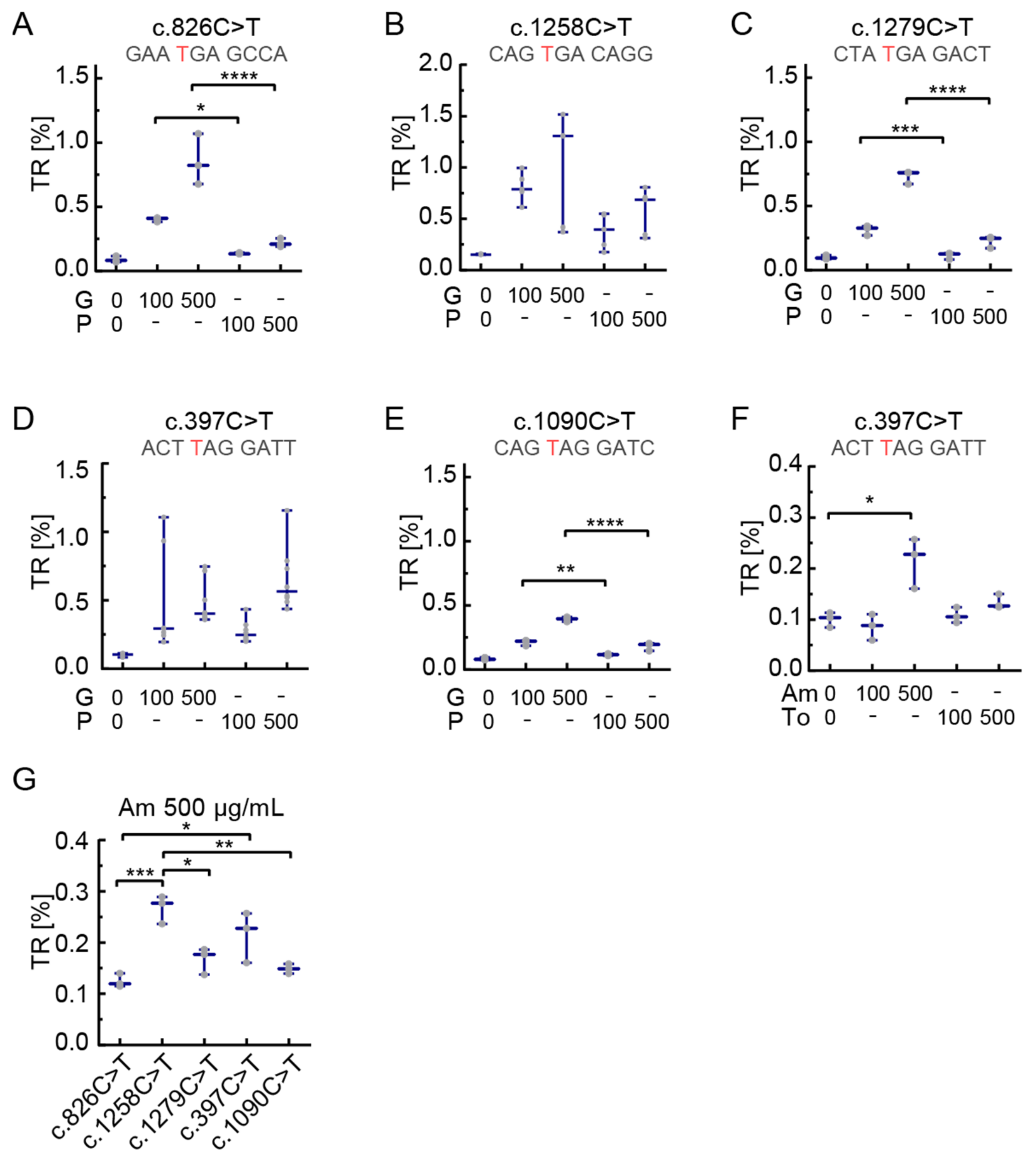
3.3. The Effect of Readthrough Induction Depends on the Stop Codon Context
4. Discussion
4.1. Stop Codon Context-Dependency of Readthrough Induction
4.2. Readthrough Stimulation as a Therapeutic Option for PBD
4.3. Readthrough Therapy as Personalized Medicine
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haendel, M.; Vasilevsky, N.; Unni, D.; Bologa, C.; Harris, N.; Rehm, H.; Hamosh, A.; Baynam, G.; Groza, T.; McMurry, J.; et al. How Many Rare Diseases Are There? Nat. Rev. Drug Discov. 2020, 19, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tambuyzer, E.; Vandendriessche, B.; Austin, C.P.; Brooks, P.J.; Larsson, K.; Miller Needleman, K.I.; Valentine, J.; Davies, K.; Groft, S.C.; Preti, R.; et al. Therapies for Rare Diseases: Therapeutic Modalities, Progress and Challenges Ahead. Nat. Rev. Drug Discov. 2020, 19, 93–111. [Google Scholar] [CrossRef] [PubMed]
- Mort, M.; Ivanov, D.; Cooper, D.N.; Chuzhanova, N.A. A Meta-Analysis of Nonsense Mutations Causing Human Genetic Disease. Hum. Mutat. 2008, 29, 1037–1047. [Google Scholar] [CrossRef] [PubMed]
- Keeling, K.M.; Xue, X.; Gunn, G.; Bedwell, D.M. Therapeutics Based on Stop Codon Readthrough. Annu. Rev. Genom. Hum. Genet. 2014, 15, 371–394. [Google Scholar] [CrossRef] [Green Version]
- Shimizu-Motohashi, Y.; Komaki, H.; Motohashi, N.; Takeda, S.; Yokota, T.; Aoki, Y. Restoring Dystrophin Expression in Duchenne Muscular Dystrophy: Current Status of Therapeutic Approaches. J. Pers. Med. 2019, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Salas-Marco, J.; Bedwell, D.M. Discrimination between Defects in Elongation Fidelity and Termination Efficiency Provides Mechanistic Insights into Translational Readthrough. J. Mol. Biol. 2005, 348, 801–815. [Google Scholar] [CrossRef]
- Zilberberg, A.; Lahav, L.; Rosin-Arbesfeld, R. Restoration of APC Gene Function in Colorectal Cancer Cells by Aminoglycoside- and Macrolide-Induced Read-through of Premature Termination Codons. Gut 2010, 59, 496–507. [Google Scholar] [CrossRef]
- Friesen, W.J.; Johnson, B.; Sierra, J.; Zhuo, J.; Vazirani, P.; Xue, X.; Tomizawa, Y.; Baiazitov, R.; Morrill, C.; Ren, H.; et al. The Minor Gentamicin Complex Component, X2, Is a Potent Premature Stop Codon Readthrough Molecule with Therapeutic Potential. PLoS ONE 2018, 13, e0206158. [Google Scholar] [CrossRef] [Green Version]
- Qian, Y.; Guan, M.-X. Interaction of Aminoglycosides with Human Mitochondrial 12S RRNA Carrying the Deafness-Associated Mutation. Antimicrob. Agents Chemother. 2009, 53, 4612–4618. [Google Scholar] [CrossRef] [Green Version]
- Ali, B.H.; Al Za’abi, M.; Blunden, G.; Nemmar, A. Experimental Gentamicin Nephrotoxicity and Agents That Modify It: A Mini-Review of Recent Research. Basic Clin. Pharmacol. Toxicol. 2011, 109, 225–232. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.-X.; Korkmaz, A.; Fuentes-Broto, L. Drug-Mediated Ototoxicity and Tinnitus: Alleviation with Melatonin. J. Physiol. Pharmacol. 2011, 62, 151–157. [Google Scholar]
- Nudelman, I.; Glikin, D.; Smolkin, B.; Hainrichson, M.; Belakhov, V.; Baasov, T. Repairing Faulty Genes by Aminoglycosides: Development of New Derivatives of Geneticin (G418) with Enhanced Suppression of Diseases-Causing Nonsense Mutations. Bioorg. Med. Chem. 2010, 18, 3735–3746. [Google Scholar] [CrossRef]
- Goldmann, T.; Overlack, N.; Möller, F.; Belakhov, V.; van Wyk, M.; Baasov, T.; Wolfrum, U.; Nagel-Wolfrum, K. A Comparative Evaluation of NB30, NB54 and PTC124 in Translational Read-through Efficacy for Treatment of an USH1C Nonsense Mutation. EMBO Mol. Med. 2012, 4, 1186–1199. [Google Scholar] [CrossRef] [PubMed]
- Bidou, L.; Bugaud, O.; Belakhov, V.; Baasov, T.; Namy, O. Characterization of New-Generation Aminoglycoside Promoting Premature Termination Codon Readthrough in Cancer Cells. RNA Biol. 2017, 14, 378–388. [Google Scholar] [CrossRef] [Green Version]
- Xue, X.; Mutyam, V.; Tang, L.; Biswas, S.; Du, M.; Jackson, L.A.; Dai, Y.; Belakhov, V.; Shalev, M.; Chen, F.; et al. Synthetic Aminoglycosides Efficiently Suppress Cystic Fibrosis Transmembrane Conductance Regulator Nonsense Mutations and Are Enhanced by Ivacaftor. Am. J. Respir. Cell Mol. Biol. 2014, 50, 805–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins-Dias, P.; Romão, L. Nonsense Suppression Therapies in Human Genetic Diseases. Cell Mol. Life Sci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Hofhuis, J.; Schueren, F.; Nötzel, C.; Lingner, T.; Gärtner, J.; Jahn, O.; Thoms, S. The Functional Readthrough Extension of Malate Dehydrogenase Reveals a Modification of the Genetic Code. Open Biol. 2016, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schueren, F.; Thoms, S. Functional Translational Readthrough: A Systems Biology Perspective. PLoS Genet. 2016, 12, e1006196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namy, O.; Hatin, I.; Rousset, J.P. Impact of the Six Nucleotides Downstream of the Stop Codon on Translation Termination. EMBO Rep. 2001, 2, 787–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schueren, F.; Lingner, T.; George, R.; Hofhuis, J.; Dickel, C.; Gärtner, J.; Thoms, S. Peroxisomal Lactate Dehydrogenase Is Generated by Translational Readthrough in Mammals. eLife 2014, 3, e03640. [Google Scholar] [CrossRef] [PubMed]
- Wangen, J.R.; Green, R. Stop Codon Context Influences Genome-Wide Stimulation of Termination Codon Readthrough by Aminoglycosides. eLife 2020, 9. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Rice, C.M. The Signal for Translational Readthrough of a UGA Codon in Sindbis Virus RNA Involves a Single Cytidine Residue Immediately Downstream of the Termination Codon. J. Virol. 1993, 67, 5062–5067. [Google Scholar] [CrossRef] [Green Version]
- McCaughan, K.K.; Brown, C.M.; Dalphin, M.E.; Berry, M.J.; Tate, W.P. Translational Termination Efficiency in Mammals Is Influenced by the Base Following the Stop Codon. Proc. Natl. Acad. Sci. USA 1995, 92, 5431–5435. [Google Scholar] [CrossRef] [Green Version]
- Hamburg, M.A.; Collins, F.S. The Path to Personalized Medicine. N. Engl. J. Med. 2010, 363, 301–304. [Google Scholar] [CrossRef]
- Bielinski, S.J.; Olson, J.E.; Pathak, J.; Weinshilboum, R.M.; Wang, L.; Lyke, K.J.; Ryu, E.; Targonski, P.V.; Van Norstrand, M.D.; Hathcock, M.A.; et al. Preemptive Genotyping for Personalized Medicine: Design of the Right Drug, Right Dose, Right Time—Using Genomic Data to Individualize Treatment Protocol. Mayo Clin. Proc. 2014, 89, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebberink, M.S.; Mooijer, P.A.W.; Gootjes, J.; Koster, J.; Wanders, R.J.A.; Waterham, H.R. Genetic Classification and Mutational Spectrum of More than 600 Patients with a Zellweger Syndrome Spectrum Disorder. Hum. Mutat. 2011, 32, 59–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodt, G.; Braverman, N.; Wong, C.; Moser, A.; Moser, H.W.; Watkins, P.; Valle, D.; Gould, S.J. Mutations in the PTS1 Receptor Gene, PXR1, Define Complementation Group 2 of the Peroxisome Biogenesis Disorders. Nat. Genet. 1995, 9, 115–125. [Google Scholar] [CrossRef]
- Wiemer, E.A.; Nuttley, W.M.; Bertolaet, B.L.; Li, X.; Francke, U.; Wheelock, M.J.; Anné, U.K.; Johnson, K.R.; Subramani, S. Human Peroxisomal Targeting Signal-1 Receptor Restores Peroxisomal Protein Import in Cells from Patients with Fatal Peroxisomal Disorders. J. Cell Biol. 1995, 130, 51–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dranchak, P.K.; Di Pietro, E.; Snowden, A.; Oesch, N.; Braverman, N.E.; Steinberg, S.J.; Hacia, J.G. Nonsense Suppressor Therapies Rescue Peroxisome Lipid Metabolism and Assembly in Cells from Patients with Specific PEX Gene Mutations. J. Cell. Biochem. 2011, 112, 1250–1258. [Google Scholar] [CrossRef] [Green Version]
- Soliman, K.; Göttfert, F.; Rosewich, H.; Thoms, S.; Gärtner, J. Super-Resolution Imaging Reveals the Sub-Diffraction Phenotype of Zellweger Syndrome Ghosts and Wild-Type Peroxisomes. Sci. Rep. 2018, 8, 7809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofhuis, J.; Dieterle, S.; George, R.; Schueren, F.; Thoms, S. Dual Reporter Systems for the Analysis of Translational Readthrough in Mammals. In Peroxisomes: Methods and Protocols; Methods in Molecular Biology; Schrader, M., Ed.; Springer: New York, NY, USA, 2017; pp. 81–92. ISBN 978-1-4939-6937-1. [Google Scholar]
- Loughran, G.; Chou, M.-Y.; Ivanov, I.P.; Jungreis, I.; Kellis, M.; Kiran, A.M.; Baranov, P.V.; Atkins, J.F. Evidence of Efficient Stop Codon Readthrough in Four Mammalian Genes. Nucleic Acids Res. 2014, 42, 8928–8938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiebler, A.C.; Freitag, J.; Schink, K.O.; Stehlik, T.; Tillmann, B.A.M.; Ast, J.; Bölker, M. Ribosomal Readthrough at a Short UGA Stop Codon Context Triggers Dual Localization of Metabolic Enzymes in Fungi and Animals. PLoS Genet. 2014, 10, e1004685. [Google Scholar] [CrossRef] [PubMed]
- Dabrowski, M.; Bukowy-Bieryllo, Z.; Zietkiewicz, E. Translational Readthrough Potential of Natural Termination Codons in Eucaryotes—The Impact of RNA Sequence. RNA Biol. 2015, 12, 950–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manuvakhova, M.; Keeling, K.; Bedwell, D.M. Aminoglycoside Antibiotics Mediate Context-Dependent Suppression of Termination Codons in a Mammalian Translation System. RNA 2000, 6, 1044–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunn, G.; Dai, Y.; Du, M.; Belakhov, V.; Kandasamy, J.; Schoeb, T.R.; Baasov, T.; Bedwell, D.M.; Keeling, K.M. Long-Term Nonsense Suppression Therapy Moderates MPS I-H Disease Progression. Mol. Genet. Metab. 2014, 111, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Floquet, C.; Hatin, I.; Rousset, J.-P.; Bidou, L. Statistical Analysis of Readthrough Levels for Nonsense Mutations in Mammalian Cells Reveals a Major Determinant of Response to Gentamicin. PLoS Genet. 2012, 8, e1002608. [Google Scholar] [CrossRef]
- Sangkuhl, K.; Schulz, A.; Römpler, H.; Yun, J.; Wess, J.; Schöneberg, T. Aminoglycoside-Mediated Rescue of a Disease-Causing Nonsense Mutation in the V2 Vasopressin Receptor Gene in Vitro and in Vivo. Hum. Mol. Genet. 2004, 13, 893–903. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Belakhov, V.; Kandasamy, J.; Baasov, T.; Li, S.-C.; Li, Y.-T.; Bedwell, D.M.; Keeling, K.M. The Designer Aminoglycoside NB84 Significantly Reduces Glycosaminoglycan Accumulation Associated with MPS I-H in the Idua-W392X Mouse. Mol. Genet. Metab. 2012, 105, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Khajavi, M.; Inoue, K.; Lupski, J.R. Nonsense-Mediated MRNA Decay Modulates Clinical Outcome of Genetic Disease. Eur. J. Hum. Genet. 2006, 14, 1074–1081. [Google Scholar] [CrossRef] [Green Version]
- Linde, L.; Boelz, S.; Nissim-Rafinia, M.; Oren, Y.S.; Wilschanski, M.; Yaacov, Y.; Virgilis, D.; Neu-Yilik, G.; Kulozik, A.E.; Kerem, E.; et al. Nonsense-Mediated MRNA Decay Affects Nonsense Transcript Levels and Governs Response of Cystic Fibrosis Patients to Gentamicin. J. Clin. Invest. 2007, 117, 683–692. [Google Scholar] [CrossRef] [Green Version]
- Brogna, S.; McLeod, T.; Petric, M. The Meaning of NMD: Translate or Perish. Trends Genet. 2016, 32, 395–407. [Google Scholar] [CrossRef] [Green Version]
- Dyle, M.C.; Kolakada, D.; Cortazar, M.A.; Jagannathan, S. How to Get Away with Nonsense: Mechanisms and Consequences of Escape from Nonsense-Mediated RNA Decay. Wiley Interdiscip. Rev. RNA 2020, 11, e1560. [Google Scholar] [CrossRef] [PubMed]
- Baradaran-Heravi, A.; Balgi, A.D.; Hosseini-Farahabadi, S.; Choi, K.; Has, C.; Roberge, M. Effect of Small Molecule ERF3 Degraders on Premature Termination Codon Readthrough. Nucleic Acids Res. 2021, 49, 3692–3708. [Google Scholar] [CrossRef] [PubMed]
- Hosseini-Farahabadi, S.; Baradaran-Heravi, A.; Zimmerman, C.; Choi, K.; Flibotte, S.; Roberge, M. Small Molecule Y-320 Stimulates Ribosome Biogenesis, Protein Synthesis, and Aminoglycoside-Induced Premature Termination Codon Readthrough. PLoS Biol. 2021, 19, e3001221. [Google Scholar] [CrossRef] [PubMed]
- Beznosková, P.; Bidou, L.; Namy, O.; Valášek, L.S. Increased Expression of Tryptophan and Tyrosine TRNAs Elevates Stop Codon Readthrough of Reporter Systems in Human Cell Lines. Nucleic Acids Res. 2021, 49, 5202–5215. [Google Scholar] [CrossRef] [PubMed]
- Thada, V.; Miller, J.N.; Kovács, A.D.; Pearce, D.A. Tissue-Specific Variation in Nonsense Mutant Transcript Level and Drug-Induced Read-through Efficiency in the Cln1(R151X) Mouse Model of INCL. J. Cell Mol. Med. 2016, 20, 381–385. [Google Scholar] [CrossRef]
- Khwaja, A.; Bjorkholm, M.; Gale, R.E.; Levine, R.L.; Jordan, C.T.; Ehninger, G.; Bloomfield, C.D.; Estey, E.; Burnett, A.; Cornelissen, J.J.; et al. Acute Myeloid Leukaemia. Nature Rev. Dis. Primers 2016, 2, 1–22. [Google Scholar] [CrossRef]
- Forrest, S.J.; Geoerger, B.; Janeway, K.A. Precision Medicine in Pediatric Oncology. Curr. Opin. Pediatr. 2018, 30, 17–24. [Google Scholar] [CrossRef]
- Nakagawa, H.; Fujita, M. Whole Genome Sequencing Analysis for Cancer Genomics and Precision Medicine. Cancer Sci. 2018, 109, 513–522. [Google Scholar] [CrossRef] [Green Version]
- Schiffelers, R.; Storm, G.; Bakker-Woudenberg, I. Liposome-Encapsulated Aminoglycosides in Pre-Clinical and Clinical Studies. J. Antimicrob. Chemother. 2001, 48, 333–344. [Google Scholar] [CrossRef] [Green Version]
- Campbell, K.C.M.; Meech, R.P.; Klemens, J.J.; Gerberi, M.T.; Dyrstad, S.S.W.; Larsen, D.L.; Mitchell, D.L.; El-Azizi, M.; Verhulst, S.J.; Hughes, L.F. Prevention of Noise- and Drug-Induced Hearing Loss with D-Methionine. Heart Res. 2007, 226, 92–103. [Google Scholar] [CrossRef]
- Du, M.; Keeling, K.M.; Fan, L.; Liu, X.; Bedwell, D.M. Poly-L-Aspartic Acid Enhances and Prolongs Gentamicin-Mediated Suppression of the CFTR-G542X Mutation in a Cystic Fibrosis Mouse Model. J. Biol. Chem. 2009, 284, 6885–6892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nudelman, I.; Rebibo-Sabbah, A.; Shallom-Shezifi, D.; Hainrichson, M.; Stahl, I.; Ben-Yosef, T.; Baasov, T. Redesign of Aminoglycosides for Treatment of Human Genetic Diseases Caused by Premature Stop Mutations. Bioorg. Med. Chem. Lett. 2006, 16, 6310–6315. [Google Scholar] [CrossRef] [PubMed]
- Goldmann, T.; Rebibo-Sabbah, A.; Overlack, N.; Nudelman, I.; Belakhov, V.; Baasov, T.; Ben-Yosef, T.; Wolfrum, U.; Nagel-Wolfrum, K. Beneficial Read-through of a USH1C Nonsense Mutation by Designed Aminoglycoside NB30 in the Retina. Invest. Ophthalmol. Vis. Sci. 2010, 51, 6671–6680. [Google Scholar] [CrossRef] [PubMed]
- Rowe, S.M.; Sloane, P.; Tang, L.P.; Backer, K.; Mazur, M.; Buckley-Lanier, J.; Nudelman, I.; Belakhov, V.; Bebok, Z.; Schwiebert, E.; et al. Suppression of CFTR Premature Termination Codons and Rescue of CFTR Protein and Function by the Synthetic Aminoglycoside NB54. J. Mol. Med. 2011, 89, 1149–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandasamy, J.; Atia-Glikin, D.; Shulman, E.; Shapira, K.; Shavit, M.; Belakhov, V.; Baasov, T. Increased Selectivity toward Cytoplasmic versus Mitochondrial Ribosome Confers Improved Efficiency of Synthetic Aminoglycosides in Fixing Damaged Genes: A Strategy for Treatment of Genetic Diseases Caused by Nonsense Mutations. J. Med. Chem. 2012, 55, 10630–10643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, D.K.; Mullenders, J.; Pott, J.; Boj, S.F.; Landskroner-Eiger, S.; Goddeeris, M.M. Targeting G542X CFTR Nonsense Alleles with ELX-02 Restores CFTR Function in Human-Derived Intestinal Organoids. J. Cyst. Fibros. 2021. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, N.; Carr, A.-J.; Lane, A.; Moeller, F.; Chen, L.L.; Aguilà, M.; Nommiste, B.; Muthiah, M.N.; Kanuga, N.; Wolfrum, U.; et al. Translational Read-through of the RP2 Arg120stop Mutation in Patient IPSC-Derived Retinal Pigment Epithelium Cells. Hum. Mol. Genet. 2015, 24, 972–986. [Google Scholar] [CrossRef] [Green Version]
- Matalonga, L.; Arias, Á.; Tort, F.; Ferrer-Cortés, X.; Garcia-Villoria, J.; Coll, M.J.; Gort, L.; Ribes, A. Effect of Readthrough Treatment in Fibroblasts of Patients Affected by Lysosomal Diseases Caused by Premature Termination Codons. Neurotherapeutics 2015, 12, 874–886. [Google Scholar] [CrossRef] [Green Version]
- Bolze, F.; Mocek, S.; Zimmermann, A.; Klingenspor, M. Aminoglycosides, but Not PTC124 (Ataluren), Rescue Nonsense Mutations in the Leptin Receptor and in Luciferase Reporter Genes. Sci. Rep. 2017, 7, 1020. [Google Scholar] [CrossRef] [Green Version]
- Torriano, S.; Erkilic, N.; Baux, D.; Cereso, N.; De Luca, V.; Meunier, I.; Moosajee, M.; Roux, A.-F.; Hamel, C.P.; Kalatzis, V. The Effect of PTC124 on Choroideremia Fibroblasts and IPSC-Derived RPE Raises Considerations for Therapy. Sci. Rep. 2018, 8, 8234. [Google Scholar] [CrossRef] [PubMed]
- Kosmidis, G.; Veerman, C.C.; Casini, S.; Verkerk, A.O.; van de Pas, S.; Bellin, M.; Wilde, A.A.M.; Mummery, C.L.; Bezzina, C.R. Readthrough-Promoting Drugs Gentamicin and PTC124 Fail to Rescue Nav1.5 Function of Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes Carrying Nonsense Mutations in the Sodium Channel Gene SCN5A. Circ. Arrhythm. Electrophysiol. 2016, 9. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schilff, M.; Sargsyan, Y.; Hofhuis, J.; Thoms, S. Stop Codon Context-Specific Induction of Translational Readthrough. Biomolecules 2021, 11, 1006. https://doi.org/10.3390/biom11071006
Schilff M, Sargsyan Y, Hofhuis J, Thoms S. Stop Codon Context-Specific Induction of Translational Readthrough. Biomolecules. 2021; 11(7):1006. https://doi.org/10.3390/biom11071006
Chicago/Turabian StyleSchilff, Mirco, Yelena Sargsyan, Julia Hofhuis, and Sven Thoms. 2021. "Stop Codon Context-Specific Induction of Translational Readthrough" Biomolecules 11, no. 7: 1006. https://doi.org/10.3390/biom11071006
APA StyleSchilff, M., Sargsyan, Y., Hofhuis, J., & Thoms, S. (2021). Stop Codon Context-Specific Induction of Translational Readthrough. Biomolecules, 11(7), 1006. https://doi.org/10.3390/biom11071006