
Treadmill Running Changes Endothelial Lipase Expression: Insights from Gene and Protein Analysis in Various Striated Muscle Tissues and Serum



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
- sedentary control (Ctrl),
- a group that underwent a moderately intense run on a treadmill set at +10° incline with the speed of 18 [m/min] for 30 [min] (M30),
- a group that underwent a fast run on a treadmill set at +10° incline with the speed of 28 [m/min] for 30 [min] (F30),
- a group that underwent a moderately intense run on a treadmill set at +10° incline with the speed of 18 [m/min] for 120 [min] (M120).
2.2. Western Blot Analysis
2.3. RNA Isolation and Gene Expression Analysis
- EL—forward primer (5′-3′) ACCAGAGTGGTGGGACGTAG; reverse primer (5′-3′) GGACAGCCTCCTGTTGATGT
- Cyclophilin A—forward primer (5′-3′) TGTCTCTTTTCGCCGCTTGCTG; reverse primer (5′-3′) CACCACCCTGGCACATGAATCC
2.4. Determination of EL, Cytokines, Total and HDL Cholesterol Levels in Serum
2.5. Statistics
3. Results
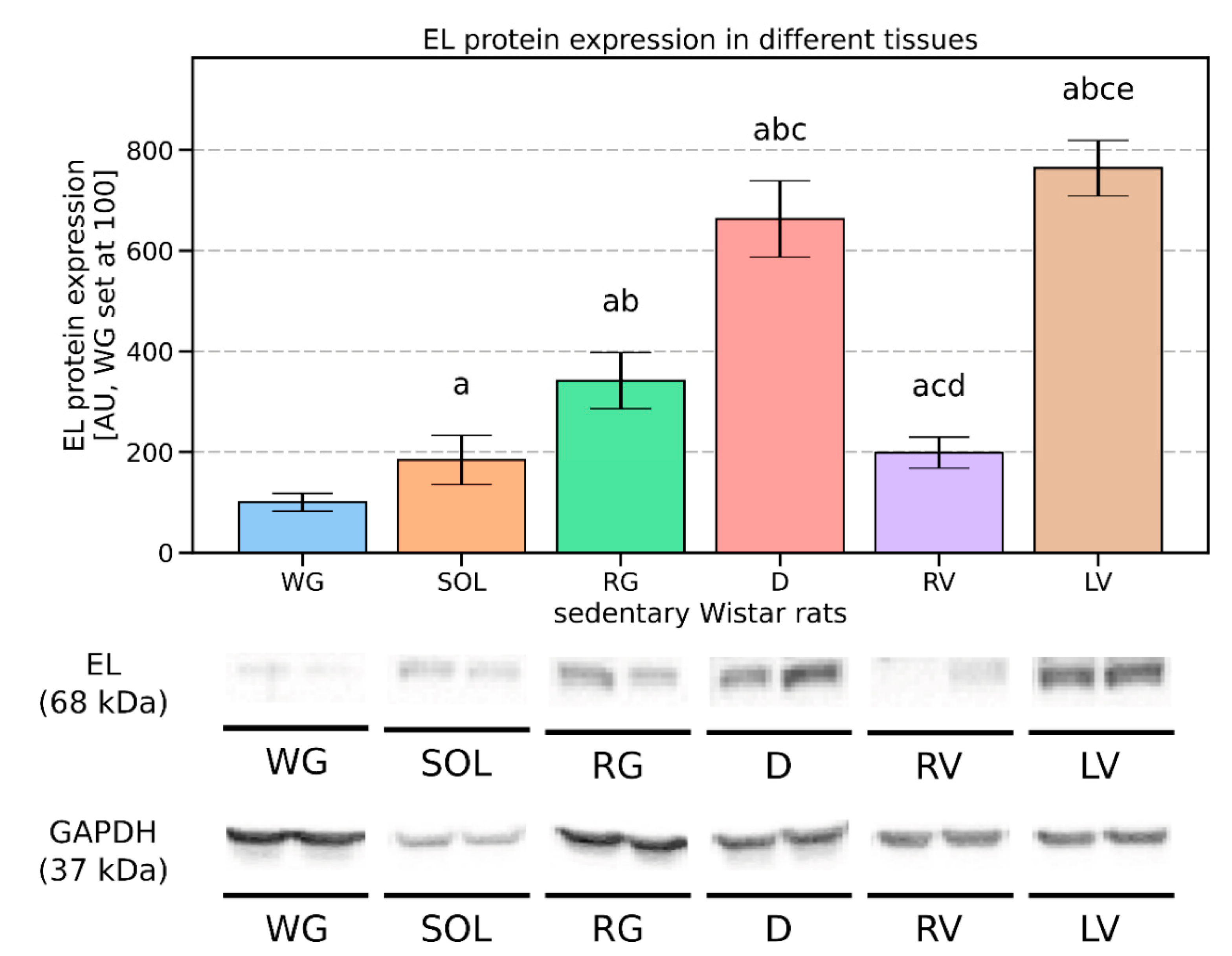
3.1. Endothelial Lipase’s Expression Pattern in the Vascular Beds of Rat Muscles
3.2. Treadmill Running Affects the Gene and Protein Expression of EL in Vascular Beds of Different Rat Muscles
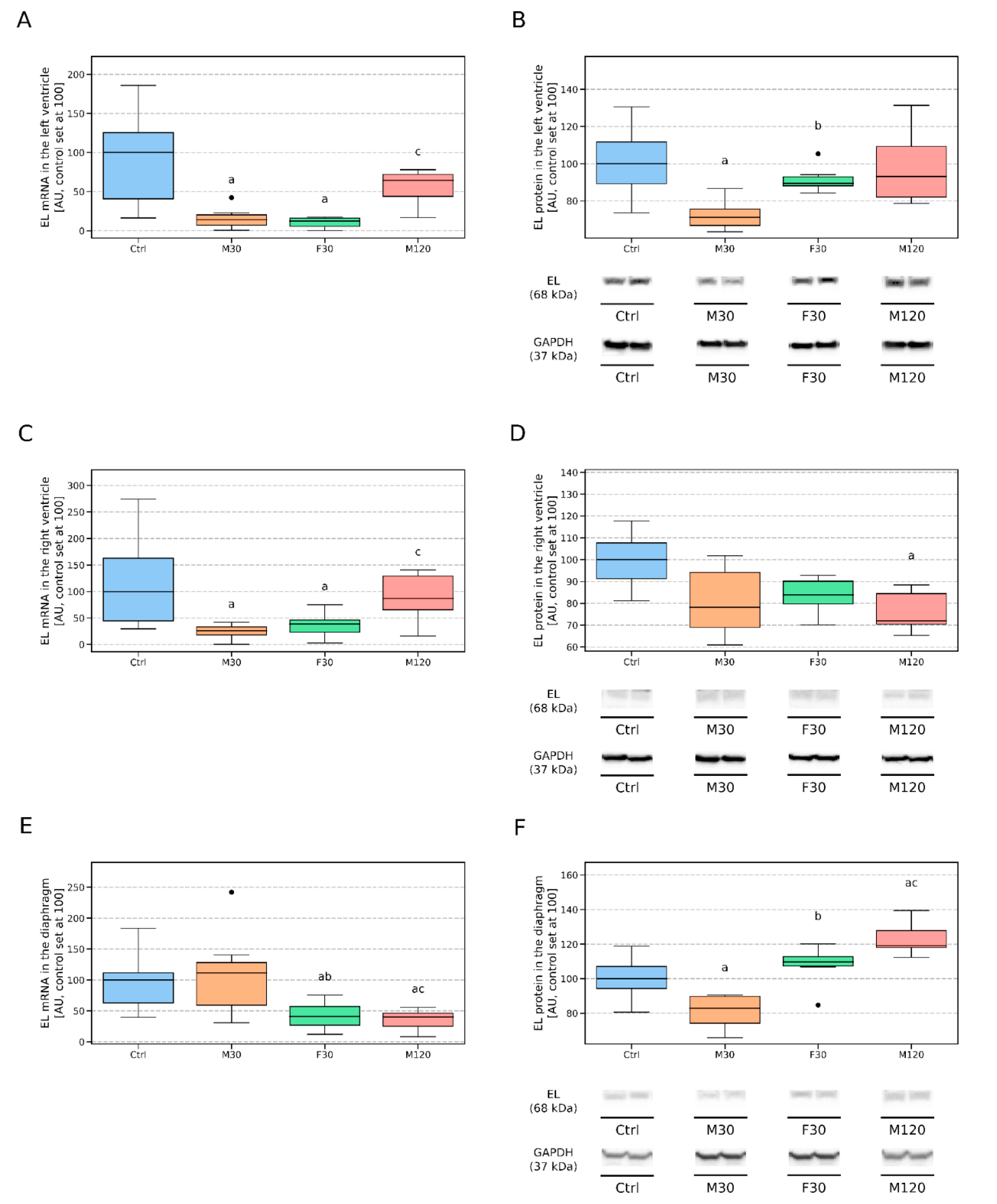
3.2.1. Left Ventricle (LV)
3.2.2. Right Ventricle (RV)
3.2.3. Diaphragm (D)
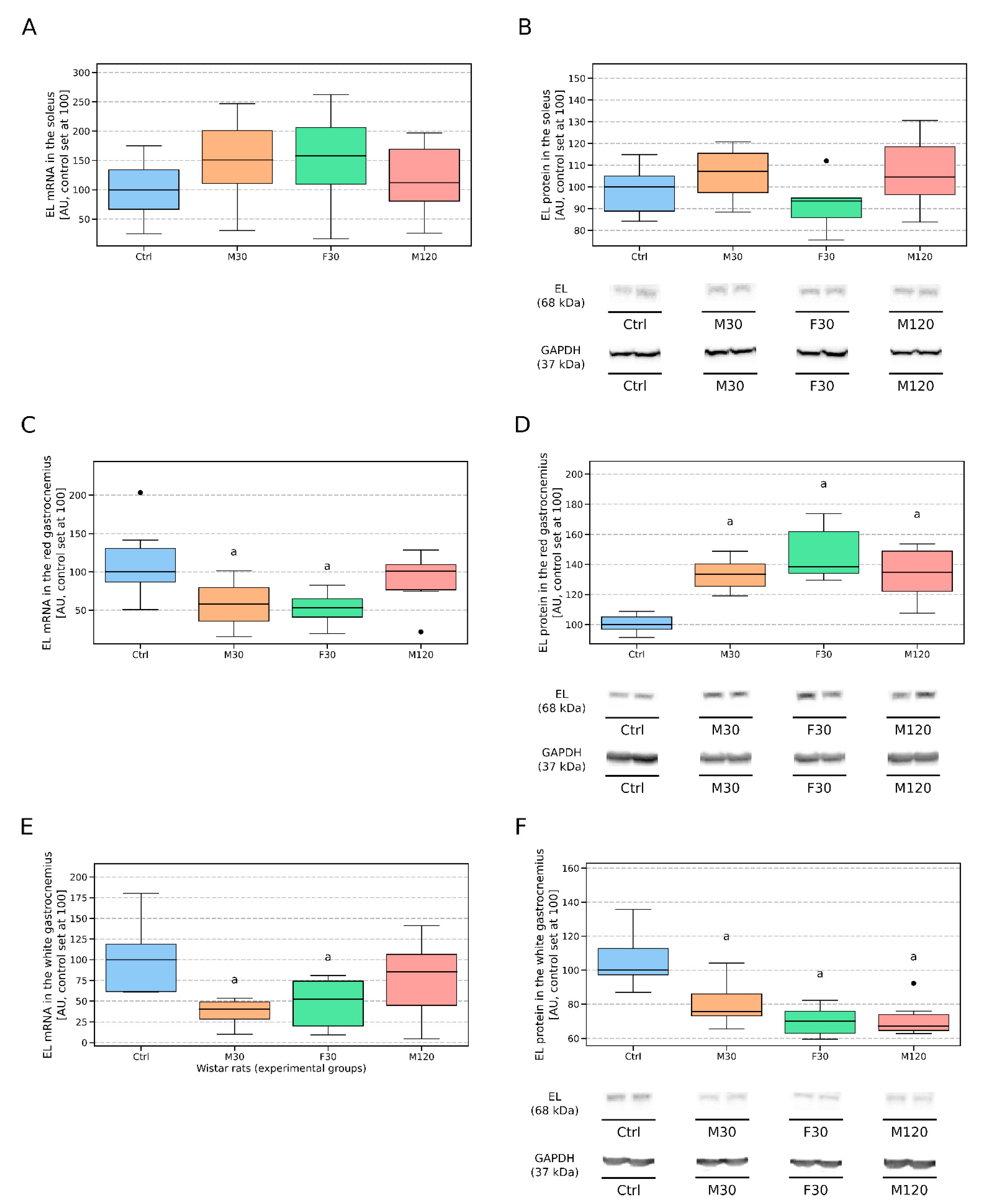
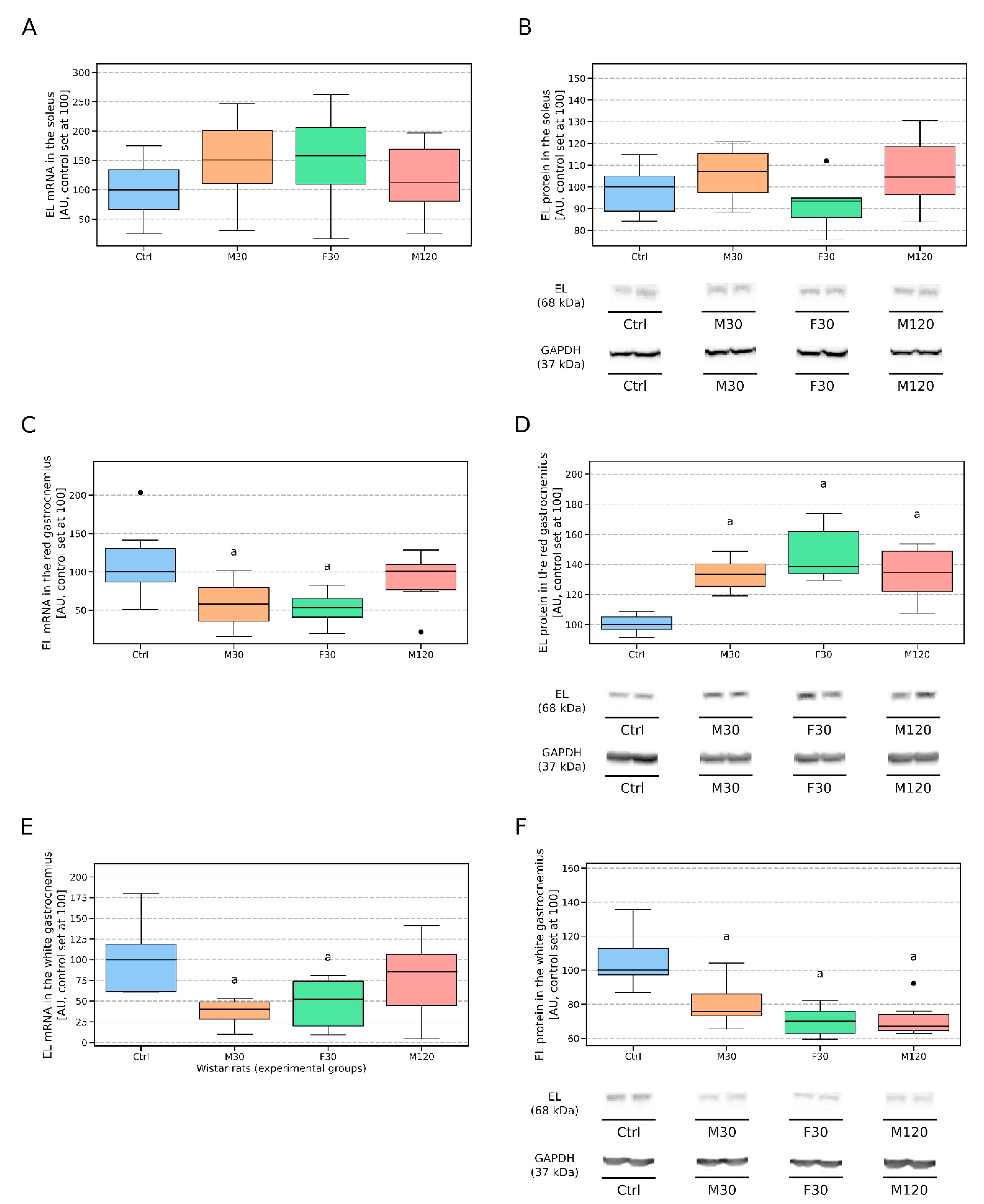
3.2.4. Soleus
3.2.5. Red Gastrocnemius
3.2.6. White Gastrocnemius
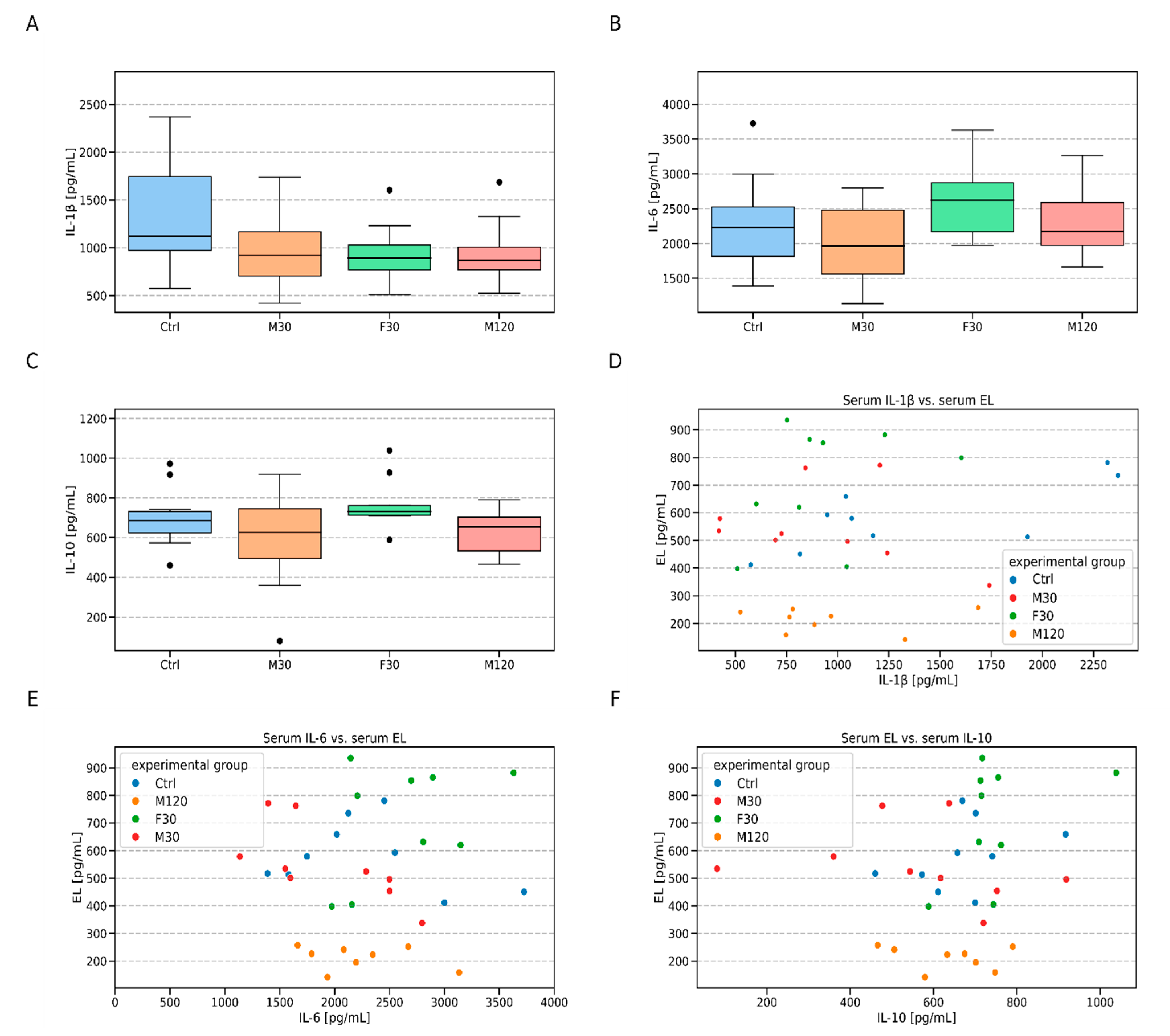
3.3. Serum Endothelial lipase (EL), Total- and HDL- Cholesterol and Cytokines (IL-1β, IL-6, and IL-10) Concentration
4. Discussion
4.1. Expression Profile of EL in Whole Muscle Tissue Homogenates
4.2. Changes in EL Expression Induced by a Single Bout of Exercise
4.3. Serum Endothelial Lipase (EL) and HDL-Cholesterol Levels
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ATGL | adipose triglyceride lipase |
| CGI-58 | 1-acylglycerol-3-phosphate O-acyltransferase (ABHD5) |
| EL | endothelial lipase |
| G0S2 | G0/G1 switch 2 protein |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| HDL | high density lipoproteins |
| HDL-C | high density lipoprotein cholesterol |
| HL | hepatic lipase |
| IL-10 | interleukin 10 |
| IL-1ra | interleukin 1 receptor antagonist |
| IL-1β | interleukin-1β |
| IL-6 | interleukin-6 |
| LDL | low density lipoproteins |
| LIPG | alternative abbreviation (aside from EL) used in case of endothelial lipase |
| LPL | lipoprotein lipase |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| SHR | spontaneously hypertensive rats |
| TG | triglyceride |
| TNFα | tumor necrosis factor α |
| VLDL | very low density lipoproteins |
| VO2max | maximal oxygen uptake |
| WKY | Wistar Kyoto rats |
References
- Ishida, T.; Choi, S.; Kundu, R.K.; Hirata, K.-I.; Rubin, E.M.; Cooper, A.D.; Quertermous, T. Endothelial lipase is a major determinant of HDL level. J. Clin. Investig. 2003, 111, 347–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, C.; Deng, R.; Wang, P.; Lu, X.; Zhao, X.; Wang, X.; Cai, R.; Lin, J. LIPG: An inflammation and cancer modulator. Cancer Gene Ther. 2021, 28, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, T.; Ishida, T.; Rader, D.J. Update on the Role of Endothelial Lipase in High-Density Lipoprotein Metabolism, Reverse Cholesterol Transport, and Atherosclerosis. Circ. J. 2010, 74, 2263–2270. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.E.; Han, S.-Y.; Wolfson, B.; Zhou, Q. The role of endothelial lipase in lipid metabolism, inflammation, and cancer. Histol. Histopathol. 2017, 33, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jaye, M.; Lynch, K.J.; A Krawiec, J.; Marchadier, D.; Maugeais, C.; Doan, K.; South, V.J.; Amin, D.V.; Perrone, M.H.; Rader, D.J. A novel endothelial-derived lipase that modulates HDL metabolism. Nat. Genet. 1999, 21, 424–428. [Google Scholar] [CrossRef]
- Azumi, H.; Hirata, K.-I.; Ishida, T.; Kojima, Y.; Rikitake, Y.; Takeuchi, S.; Inoue, N.; Kawashima, S.; Hayashi, Y.; Itoh, H.; et al. Immunohistochemical localization of endothelial cell-derived lipase in atherosclerotic human coronary arteries. Cardiovasc. Res. 2003, 58, 647–654. [Google Scholar] [CrossRef] [Green Version]
- Hirata, K.-I.; Dichek, H.L.; Cioffi, J.A.; Choi, S.Y.; Leeper, N.J.; Quintana, L.; Kronmal, G.S.; Cooper, A.D.; Quertermous, T. Cloning of a Unique Lipase from Endothelial Cells Extends the Lipase Gene Family. J. Biol. Chem. 1999, 274, 14170–14175. [Google Scholar] [CrossRef] [Green Version]
- Broedl, U.C.; Jin, W.; Marchadier, D.; Secreto, A.; Rader, D.J. Tissue-specific expression pattern of human endothelial lipase in transgenic mice. Atheroscler. 2005, 181, 271–274. [Google Scholar] [CrossRef]
- Annema, W.; Tietge, U.J.F. Role of Hepatic Lipase and Endothelial Lipase in High-Density Lipoprotein—Mediated Reverse Cholesterol Transport. Curr. Atheroscler. Rep. 2011, 13, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Vigelsø, A.; Prats, C.; Ploug, T.; Dela, F.; Helge, J. Human skeletal muscle perilipin 2 and 3 expression varies with insulin sensitivity. J. Biomed. Sci. Eng. 2013, 6, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Shiu, S.W.; Tan, K.C.; Huang, Y.; Wong, Y. Type 2 diabetes mellitus and endothelial lipase. Atherosclerosis 2008, 198, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Potočnjak, I.; Trbušić, M.; Terešak, S.D.; Radulović, B.; Pregartner, G.; Berghold, A.; Tiran, B.; Marsche, G.; Degoricija, V.; Frank, S. Metabolic Syndrome Modulates Association between Endothelial Lipase and Lipid/Lipoprotein Plasma Levels in Acute Heart Failure Patients. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Ishida, T.; Miyashita, K.; Kinoshita, N.; Mori, K.; Yasuda, T.; Toh, R.; Nakajima, K.; Imamura, S.; Hirata, K.-I. Plasma Activity of Endothelial Lipase Impacts High-Density Lipoprotein Metabolism and Coronary Risk Factors in Humans. J. Atheroscler. Thromb. 2014, 21, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Qian, H.-Y.; Li, Z.-Z.; Zhang, J.-M.; Wang, S.; Tao, Y.; Gao, Y.-L.; Yin, C.-Q.; Que, B.; Sun, T.; et al. Role of endothelial lipase in atherosclerosis. Transl. Res. 2010, 156, 1–6. [Google Scholar] [CrossRef]
- Shimokawa, Y.; Hirata, K.-I.; Ishida, T.; Kojima, Y.; Inoue, N.; Quertermous, T.; Yokoyama, M. Increased expression of endothelial lipase in rat models of hypertension. Cardiovasc. Res. 2005, 66, 594–600. [Google Scholar] [CrossRef]
- Robert, J.; Lehner, M.; Frank, S.; Perisa, D.; Von Eckardstein, A.; Rohrer, L. Interleukin 6 Stimulates Endothelial Binding and Transport of High-Density Lipoprotein Through Induction of Endothelial Lipase. Arter. Thromb. Vasc. Biol. 2013, 33, 2699–2706. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.; Sun, G.-S.; Marchadier, D.; Octtaviani, E.; Glick, J.M.; Rader, D.J. Endothelial Cells Secrete Triglyceride Lipase and Phospholipase Activities in Response to Cytokines as a Result of Endothelial Lipase. Circ. Res. 2003, 92, 644–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broedl, U.C.; Jin, W.; Rader, D.J. Endothelial Lipase: A Modulator of Lipoprotein Metabolism Upregulated by Inflammation. Trends Cardiovasc. Med. 2004, 14, 202–206. [Google Scholar] [CrossRef] [PubMed]
- Badellino, K.O.; Wolfe, M.L.; Reilly, M.P.; Rader, D.J. Endothelial Lipase Is Increased In Vivo by Inflammation in Humans. Circulation 2008, 117, 678–685. [Google Scholar] [CrossRef] [Green Version]
- Paradis, M.-E.; Badellino, K.O.; Rader, D.J.; Deshaies, Y.; Couture, P.; Archer, W.R.; Bergeron, N.; Lamarche, B. Endothelial lipase is associated with inflammation in humans. J. Lipid Res. 2006, 47, 2808–2813. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.; Millar, J.S.; Broedl, U.; Glick, J.M.; Rader, D.J. Inhibition of endothelial lipase causes increased HDL cholesterol levels in vivo. J. Clin. Investig. 2003, 111, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Ichihirataa, K.; Ishidab, T.; Matsushitab, H.; Tsao, P.S.; Quertermous, T. Regulated Expression of Endothelial Cell-Derived Lipase. Biochem. Biophys. Res. Commun. 2000, 272, 90–93. [Google Scholar] [CrossRef]
- März, W.; Kleber, M.E.; Scharnagl, H.; Speer, T.; Zewinger, S.; Ritsch, A.; Parhofer, K.G.; Von Eckardstein, A.; Landmesser, U.; Laufs, U. HDL cholesterol: Reappraisal of its clinical relevance. Clin. Res. Cardiol. 2017, 106, 663–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Lu, J.; Li, J.; Guan, W.; Deng, S.; Deng, Q.; Ye, H.; Han, W.; Yu, Y.; Zhang, R. Serum Triglyceride Lipase Concentrations are Independent Risk Factors for Coronary Artery Disease and In-Stent Restenosis. J. Atheroscler. Thromb. 2019, 26, 762–774. [Google Scholar] [CrossRef] [PubMed]
- Vigelsø, A.; Prats, C.; Ploug, T.; Dela, F.; Helge, J. Higher Muscle Content of Perilipin 5 and Endothelial Lipase Protein in Trained Than Untrained Middle-Aged Men. Physiol. Res. 2016, 65, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, R.E.; Gollnick, P.D. Oxygen uptake of rats at different work intensities. Pflügers Arch. Eur. J. Physiol. 1976, 362, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Ariano, M.A.; Edgerton, V.R.; Armstrong, R.B. Hindlimb muscle fiber populations of five mammals. J. Histochem. Cytochem. 1973, 21, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, T.E.; Armstrong, R.B. Rat locomotory muscle fiber activity during trotting and galloping. J. Appl. Physiol. 1978, 44, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Miklosz, A.; Lukaszuk, B.; Chabowski, A.; Rogowski, F.; Kurek, K.; Żendzian-Piotrowska, M. Hyperthyroidism Evokes Myocardial Ceramide Accumulation. Cell. Physiol. Biochem. 2015, 35, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Choromanska, B.; Mysliwiec, P.; Hady, H.R.; Dadan, J.; Mysliwiec, H.; Bonda, T.; Chabowski, A.; Miklosz, A. The implication of adipocyte ATP-binding cassette A1 and G1 transporters in metabolic complications of obesity. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2019, 70, 143–152. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Miklosz, A.; Baranowski, M.; Lukaszuk, B.; Zabielski, P.; Chabowski, A.; Gorski, J. Effect of acute exercise on mRNA and protein expression of main components of the lipolytic complex in different skeletal muscle types in the rat. J. Physiol. Pharmacol 2019, 70, 425–433. [Google Scholar]
- Murakami, S.; Fujino, H.; Takeda, I.; Momota, R.; Kumagishi, K.; Ohtsuka, A. Comparison of capillary architecture between slow and fast muscles in rats using a confocal laser scanning microscope. Acta Med. Okayama 2010, 64, 11–18. [Google Scholar]
- Roseguini, B.T.; Laughlin, M.H. Muscle Blood Flow and Vascularization in Response to Exercise and Training; Zoladz, J., Ed.; Academic Press: London, UK, 2019; pp. 379–389. [Google Scholar]
- Rehman, S.; Khan, A.; Rehman, A. Physiology, Coronary Circulation. 2020. Available online: https://www.ncbi.nlm.nih.gov/books/NBK482413/ (accessed on 30 January 2021).
- Khazaei, M.; Fallahzadeh, A.R.; Sharifi, M.R.; Afsharmoghaddam, N.; Javanmard, S.H.; Salehi, E. Effects of diabetes on myocardial capillary density and serum angiogenesis biomarkers in male rats. Clinical 2011, 66, 1419–1424. [Google Scholar] [CrossRef] [Green Version]
- Maier, T.; Güell, M.; Serrano, L. Correlation of mRNA and protein in complex biological samples. FEBS Lett. 2009, 583, 3966–3973. [Google Scholar] [CrossRef] [Green Version]
- Lukaszuk, B.; Miklosz, A.; Zendzian-Piotrowska, M.; Wojcik, B.; Gorski, J.; Chabowski, A. Changes in the Diaphragm Lipid Content after Administration of Streptozotocin and High-Fat Diet Regime. J. Diabetes Res. 2017, 2017, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mikłosz, A.; Łukaszuk, B.; Baranowski, M.; Chabowski, A.; Górski, J. Assessment of the Main Compounds of the Lipolytic System in Treadmill Running Rats: Different Response Patterns between the Right and Left Ventricle. Int. J. Mol. Sci. 2019, 20, 2556. [Google Scholar] [CrossRef] [Green Version]
- Zong, P.; Tune, J.D.; Downey, H.F. Mechanisms of Oxygen Demand/Supply Balance in the Right Ventricle. Exp. Biol. Med. 2005, 230, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Olianti, C.; Costantini, I.; Giardini, F.; Lazzeri, E.; Crocini, C.; Ferrantini, C.; Pavone, F.S.; Camici, P.G.; Sacconi, L. 3D imaging and morphometry of the heart capillary system in spontaneously hypertensive rats and normotensive controls. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Chabowski, A.; Żendzian-Piotrowska, M.; Miklosz, A.; Lukaszuk, B.; Kurek, K.; Górski, J. Fiber Specific Changes in Sphingolipid Metabolism in Skeletal Muscles of Hyperthyroid Rats. Lipids 2013, 48, 697–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, P.; Kroese, A.J. Capillary supply in soleus and gastrocnemius muscles of man. Pflügers Archiv. Eur. J. Physiol. 1978, 375, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, N. Effect of endurance training on muscle fiber type composition and capillary supply in rat diaphragm. Graefe’s Arch. Clin. Exp. Ophthalmol. 1987, 56, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Keens, T.G.; Chen, V.; Patel, P.; O’Brien, P.; Levison, H.; Ianuzzo, C.D. Cellular adaptations of the ventilatory muscles to a chronic increased respiratory load. J. Appl. Physiol. 1978, 44, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, G.F.; Padykula, H.A. Cytological Studies of Fiber Types in Skeletal Muscle. A comparative study of the mammalian diaphragm. J. Cell Biol. 1966, 28, 333–354. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.B.; Laughlin, M.H. Exercise blood flow patterns within and among rat muscles after training. Am. J. Physiol. Circ. Physiol. 1984, 246, 59–68. [Google Scholar] [CrossRef]
- Armstrong, R.B.; Laughlin, M.H. Metabolic indicators of fibre recruitment in mammalian muscles during locomotion. J. Exp. Biol. 1985, 115, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Lawler, J.M.; Powers, S.K.; Hammeren, J.; Martin, A.D. Oxygen cost of treadmill running in 24-month-old Fischer-344 rats. Med. Sci. Sports Exerc. 1993, 25, 1259–1264. [Google Scholar] [CrossRef]
- Swain, D.P. Moderate or Vigorous Intensity Exercise: Which Is Better for Improving Aerobic Fitness? Prev. Cardiol. 2005, 8, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.A.; Kersten, S.; Qi, L. Lipoprotein Lipase and Its Regulators: An Unfolding Story. Trends Endocrinol. Metab. 2021, 32, 48–61. [Google Scholar] [CrossRef]
- Bay, M.L.; Pedersen, B.K. Muscle-Organ Crosstalk: Focus on Immunometabolism. Front. Physiol. 2020, 11, 1–8. [Google Scholar] [CrossRef]
- Pedersen, B.K. Physical activity and muscle–brain crosstalk. Nat. Rev. Endocrinol. 2019, 15, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Legård, G.E.; Pedersen, B.K. Muscle as an Endocrine Organ. In Muscle and Exercise Physiology; Zoladz, J., Ed.; Academic Press: London, UK, 2019; pp. 285–307. [Google Scholar]
- Philip, C.; Fischer, P. Interleukin-6 in Acute Exercise and Training: What is the Biological Relevance; Department of Infectious Diseases Rigshospitalet, University Hospital Copenhagen: Copenhagen, Denmark, 2016; pp. 6–33. [Google Scholar]
- Pedersen, B.K. The anti-inflammatory effect of exercise: Its role in diabetes and cardiovascular disease control. Essays Biochem. 2006, 42, 105–117. [Google Scholar] [CrossRef]
- Piccirillo, R. Exercise-Induced Myokines with Therapeutic Potential for Muscle Wasting. Front. Physiol. 2019, 10, 287–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, M.T.; Etienne, J.; McClure, W.C.; Pavey, B.S.; Holloway, A.K. Role of local contractile activity and muscle fiber type on LPL regulation during exercise. Am. J. Physiol. Metab. 1998, 275, 1016–1022. [Google Scholar] [CrossRef]
- Badellino, K.O.; Wolfe, M.L.; Reilly, M.P.; Rader, D.J. Endothelial Lipase Concentrations Are Increased in Metabolic Syndrome and Associated with Coronary Atherosclerosis. PLoS Med. 2005, 3, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miksztowicz, V.; Schreier, L.; McCoy, M.; Lucero, D.; Fassio, E.; Billheimer, J.; Rader, D.J.; Berg, G. Role of SN1 Lipases on Plasma Lipids in Metabolic Syndrome and Obesity. Arter. Thromb. Vasc. Biol. 2014, 34, 669–675. [Google Scholar] [CrossRef] [Green Version]
- Paradis, M.-E.; Badellino, K.O.; Rader, D.J.; Tchernof, A.; Richard, C.; Luu-The, V.; Deshaies, Y.; Bergeron, J.; Archer, W.R.; Couture, P.; et al. Visceral Adiposity and Endothelial Lipase. J. Clin. Endocrinol. Metab. 2006, 91, 3538–3543. [Google Scholar] [CrossRef] [Green Version]
- Assmann, G.; Gotto, A.M., Jr. HDL Cholesterol and Protective Factors in Atherosclerosis. Circulation 2004, 109, III-8–III-14. [Google Scholar] [CrossRef] [Green Version]
- Szostak, J.; Laurant, P. The forgotten face of regular physical exercise: A “natural” anti-atherogenic activity. Clin. Sci. 2011, 121, 91–106. [Google Scholar] [CrossRef] [Green Version]
- Muscella, A.; Stefàno, E.; Lunetti, P.; Capobianco, L.; Marsigliante, S. The Regulation of Fat Metabolism During Aerobic Exercise. Biomolecules 2020, 10, 1699. [Google Scholar] [CrossRef]
- Hargreaves, M.; Spriet, L.L. Exercise Metabolism: Fuels for the Fire. Cold Spring Harb. Perspect. Med. 2018, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Hirata, K.-I.; Ishida, T.; Quertermous, T.; Cooper, A.D. Endothelial lipasee: A new lipase on the block. J. Lipid Res. 2002, 43, 1763–1769. [Google Scholar] [CrossRef] [Green Version]
- Bartels, E.D.; Nielsen, J.E.; Lindegaard, M.L.; Hulten, L.M.; Schroeder, T.V.; Nielsen, L.B. Endothelial lipase is highly expressed in macrophages in advanced human atherosclerotic lesions. Atherosclerosis 2007, 195, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.D.; Crouse, S.F.; Goodpaster, B.; Kelley, D.; Moyna, N.; Pescatello, L. The acute versus the chronic response to exercise. Med. Sci. Sports Exerc. 2001, 33, 438–445. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Analysis | r (Pearson Correlation Coefficient) | p-Value Unadjusted | p-Value Adjusted (BH) |
|---|---|---|---|
| Within Ctrl | 0.910 | 0.001 | 0.005 |
| Within M30 | 0.450 | 0.229 | 0.382 |
| Within F30 | −0.290 | 0.450 | 0.450 |
| Within M120 | 0.350 | 0.402 | 0.450 |
| All groups together | 0.340 | 0.044 | 0.110 |
| Type of Analysis | r (Pearson Correlation Coefficient) | p-Value Unadjusted | p-Value Adjusted (BH) |
|---|---|---|---|
| Within Ctrl | 0.74 | 0.022 | 0.110 |
| Within M30 | −0.33 | 0.392 | 0.490 |
| Within F30 | 0.35 | 0.352 | 0.490 |
| Within M120 | −0.02 | 0.955 | 0.955 |
| All groups together | 0.18 | 0.299 | 0.490 |
| Type of Analysis | r (Pearson Correlation Coefficient) | p-Value Unadjusted | p-Value Adjusted (BH) |
|---|---|---|---|
| Within Ctrl | −0.29 | 0.455 | 0.583 |
| Within M30 | −0.70 | 0.037 | 0.185 |
| Within F30 | 0.40 | 0.283 | 0.582 |
| Within M120 | −0.30 | 0.466 | 0.582 |
| All groups together | 0.06 | 0.737 | 0.737 |
| Type of Analysis | r (Pearson Correlation Coefficient) | p-Value Unadjusted | p-Value Adjusted (BH) |
|---|---|---|---|
| Within Ctrl | 0.37 | 0.326 | 0.543 |
| Within M30 | −0.27 | 0.486 | 0.586 |
| Within F30 | 0.45 | 0.221 | 0.543 |
| Within M120 | −0.23 | 0.586 | 0.586 |
| All groups together | 0.19 | 0.283 | 0.543 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikłosz, A.; Łukaszuk, B.; Chabowski, A.; Górski, J. Treadmill Running Changes Endothelial Lipase Expression: Insights from Gene and Protein Analysis in Various Striated Muscle Tissues and Serum. Biomolecules 2021, 11, 906. https://doi.org/10.3390/biom11060906
Mikłosz A, Łukaszuk B, Chabowski A, Górski J. Treadmill Running Changes Endothelial Lipase Expression: Insights from Gene and Protein Analysis in Various Striated Muscle Tissues and Serum. Biomolecules. 2021; 11(6):906. https://doi.org/10.3390/biom11060906
Chicago/Turabian StyleMikłosz, Agnieszka, Bartłomiej Łukaszuk, Adrian Chabowski, and Jan Górski. 2021. "Treadmill Running Changes Endothelial Lipase Expression: Insights from Gene and Protein Analysis in Various Striated Muscle Tissues and Serum" Biomolecules 11, no. 6: 906. https://doi.org/10.3390/biom11060906
APA StyleMikłosz, A., Łukaszuk, B., Chabowski, A., & Górski, J. (2021). Treadmill Running Changes Endothelial Lipase Expression: Insights from Gene and Protein Analysis in Various Striated Muscle Tissues and Serum. Biomolecules, 11(6), 906. https://doi.org/10.3390/biom11060906