
Identification and Biochemical Characterization of High Mobility Group Protein 20A as a Novel Ca2+/S100A6 Target

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
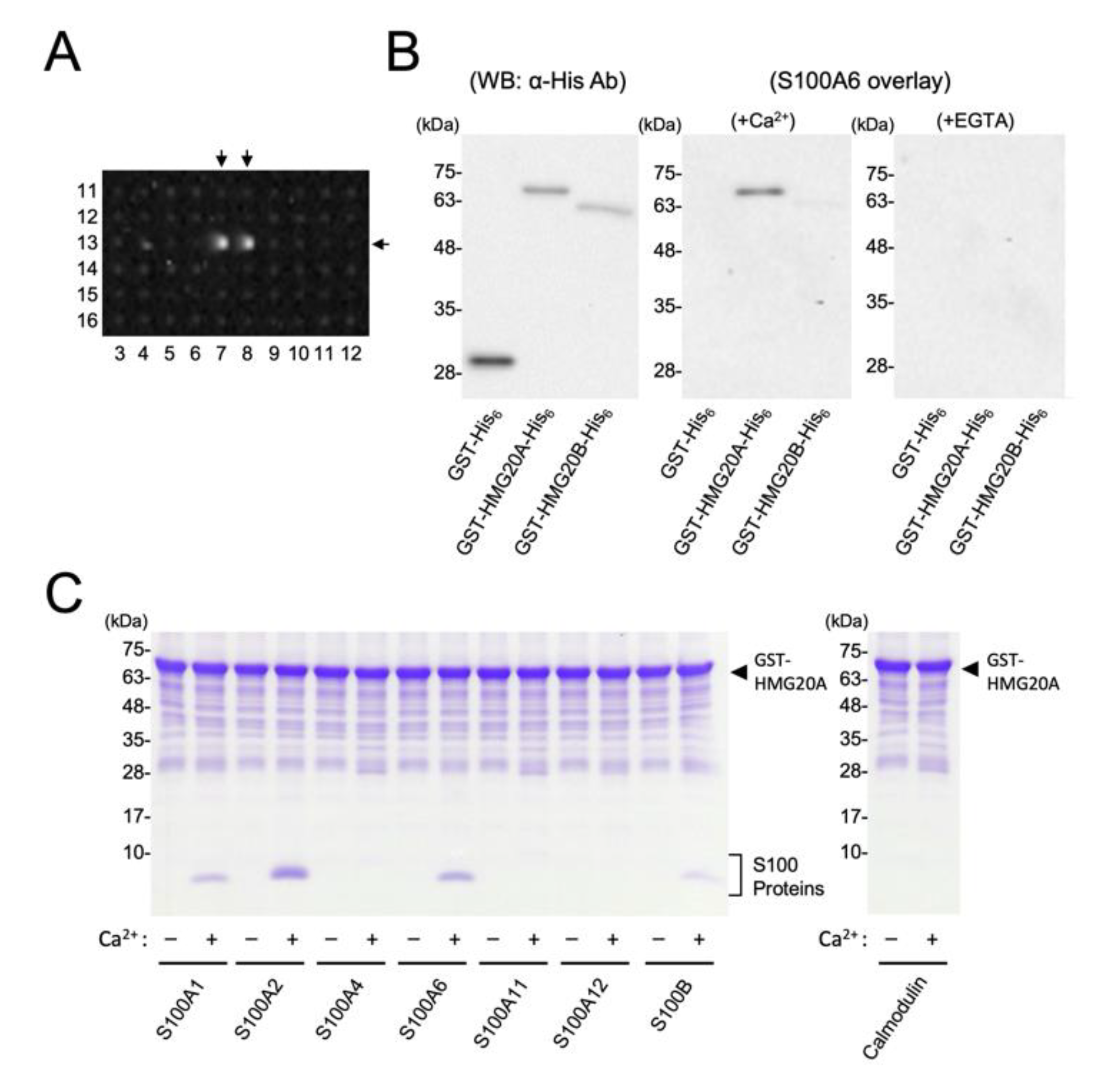
2.2. Screening of S100A6 Targets Using Human Protein Array
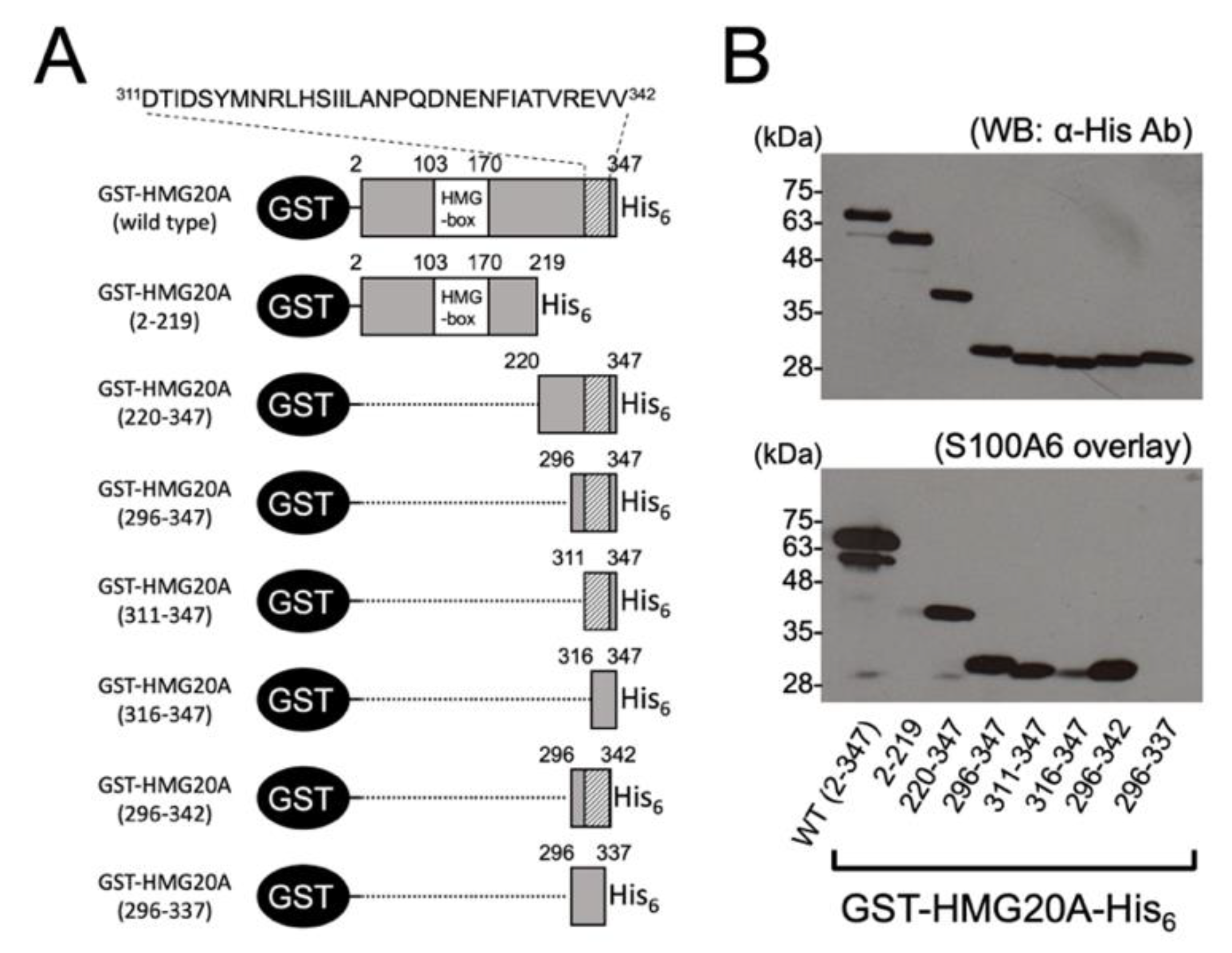
2.3. Construction, Expression, and Purification of GST-HMG20A and Mutants
- GST-HMG20A(2–219)-His6: 5′gggtctagaaaacttgatgactagctc3′/5′agggatgtcaaaaacagaccgttcc3′. GST-HMG20A(220–347)-His6: 5′gggtctagaaatatttacagaggaattcttg3′/5′acgatcgagtctgttcacaacttc3′. GST-HMG20A(296–347)-His6: 5′gggtctagaatttgccagcatgcccttgcc3′/5′acgatcgagtctgttcacaacttc3′. GST-HMG20A(311–347)-His6: 5′gggtctagaagacaccattgactc3′/5′acgatcgagtctgttcacaacttc3′. GST-HMG20A(316–347)-His6: 5′gggtctagaatatatgaacagactgcacag3′/5′acgatcgagtctgttcacaacttc3′. GST-HMG20A(296–342)-His6: 5′gggtctagaatttgccagcatgcccttgcc3′/5′cacaacttctcgaactgtagctatg3′. GST-HMG20A(296–337)-His6: 5′gggtctagaatttgccagcatgcccttgcc3′/5′tgtagctatgaagttttcattgtcttg3′.
2.4. GST Precipitation Assay
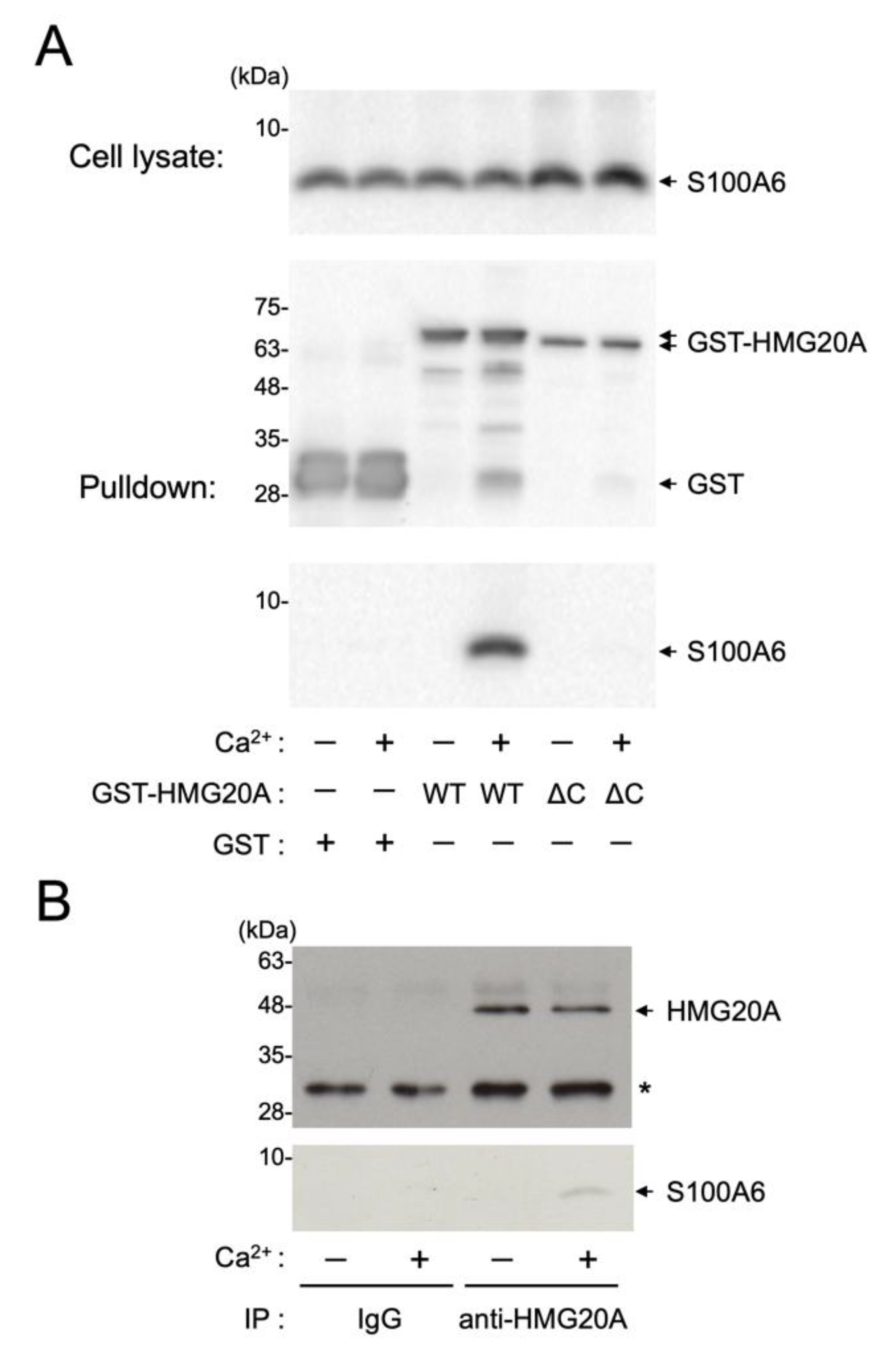
2.5. Cell Culture, Transfection, and GST Precipitation Assay
2.6. Immunoprecipitation
2.7. Other Methods
3. Results
3.1. Identification of HMG20A as an S100A6 Interacting Protein
3.2. Identification of S100A6-Binding Domain in HMG20A
3.3. Stoichiometric Binding of S100A6 with HMG20A
3.4. Interaction of HMG20A with Ca2+/S100A6 in Cultured Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marenholz, I.; Heizmann, C.W.; Fritz, G. S100 proteins in mouse and man: From evolution to function and pathology (including an update of the nomenclature). Biochem. Biophys. Res. Commun. 2004, 322, 1111–1122. [Google Scholar] [CrossRef] [PubMed]
- Donato, R. Intracellular and extracellular roles of S100 proteins. Microsc. Res. Tech. 2003, 60, 540–551. [Google Scholar] [CrossRef] [PubMed]
- Donato, R.; Sorci, G.; Giambanco, I. S100A6 protein: Functional roles. Cell. Mol. Life Sci. 2017, 74, 2749–2760. [Google Scholar] [CrossRef] [PubMed]
- Calabretta, B.; Battini, R.; Kaczmarek, L.; de Riel, J.K.; Baserga, R. Molecular cloning of the cDNA for a growth factor-inducible gene with strong homology to S-100, a calcium-binding protein. J. Biol. Chem. 1986, 261, 12628–12632. [Google Scholar] [CrossRef]
- Zeng, F.-Y.; Gerke, V.; Gabius, H.-J. Identification of annexin II, annexin VI and glyceraldehyde-3-phosphate dehydrogenase as calcyclin-binding proteins in bovine heart. Int. J. Biochem. 1993, 25, 1019–1027. [Google Scholar] [CrossRef]
- Tokumitsu, H.; Mizutani, A.; Minami, H.; Kobayashi, R.; Hidaka, H. A calcyclin-associated protein is a newly identified member of the Ca2+/phospholipid-binding proteins, annexin family. J. Biol. Chem. 1992, 267, 8919–8924. [Google Scholar] [CrossRef]
- Filipek, A.; Zasada, A.; Wojda, U.; Makuch, R.; Dąbrowska, R. Characterization of chicken gizzard calcyclin and examination of its interaction with caldesmon. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1996, 113, 745–752. [Google Scholar] [CrossRef]
- Golitsina, N.L.; Kordowska, J.; Wang, C.L.; Lehrer, S.S. Ca2+-dependent binding of calcyclin to muscle tropomyo-sin. Biochem. Biophys. Res. Commun. 1996, 220, 360–365. [Google Scholar] [CrossRef]
- Filipek, A.; Wojda, U. p30, a novel protein target of mouse calcyclin (S100A6). Biochem. J. 1996, 320, 585–587. [Google Scholar] [CrossRef]
- Filipek, A.; Jastrzebska, B.; Nowotny, M.; Kuznicki, J. CacyBP/SIP, a Calcyclin and Siah-1-interacting Protein, Binds EF-hand Proteins of the S100 Family. J. Biol. Chem. 2002, 277, 28848–28852. [Google Scholar] [CrossRef] [PubMed]
- Shimamoto, S.; Takata, M.; Tokuda, M.; Oohira, F.; Tokumitsu, H.; Kobayashi, R. Interactions of S100A2 and S100A6 with the Tetratricopeptide Repeat Proteins, Hsp90/Hsp70-organizing Protein and Kinesin Light Chain. J. Biol. Chem. 2008, 283, 28246–28258. [Google Scholar] [CrossRef]
- Yamaguchi, F.; Umeda, Y.; Shimamoto, S.; Tsuchiya, M.; Tokumitsu, H.; Tokuda, M.; Kobayashi, R. S100 pro-teins modulate protein phosphatase 5 function: A link between Ca2+ signal transduction and protein dephosphorylation. J. Biol. Chem. 2012, 287, 13787–13798. [Google Scholar] [CrossRef] [PubMed]
- Shimamoto, S.; Tsuchiya, M.; Yamaguchi, F.; Kubota, Y.; Tokumitsu, H.; Kobayashi, R. Ca2+/S100 proteins inhibit the interaction of FKBP38 with Bcl-2 and Hsp90. Biochem. J. 2014, 458, 141–152. [Google Scholar] [CrossRef]
- Shimamoto, S.; Kubota, Y.; Yamaguchi, F.; Tokumitsu, H.; Kobayashi, R. Ca2+/S100 Proteins Act as Upstream Regulators of the Chaperone-associated Ubiquitin Ligase CHIP (C Terminus of Hsc70-interacting Protein). J. Biol. Chem. 2013, 288, 7158–7168. [Google Scholar] [CrossRef]
- Leclerc, E.; Fritz, G.; Weibel, M.; Heizmann, C.W.; Galichet, A. S100B and S100A6 Differentially Modulate Cell Survival by Interacting with Distinct RAGE (Receptor for Advanced Glycation End Products) Immunoglobulin Domains. J. Biol. Chem. 2007, 282, 31317–31331. [Google Scholar] [CrossRef]
- Filipek, A.; Leśniak, W. S100A6 and Its Brain Ligands in Neurodegenerative Disorders. Int. J. Mol. Sci. 2020, 21, 3979. [Google Scholar] [CrossRef]
- Furuya, Y.; Denda, M.; Sakane, K.; Ogusu, T.; Takahashi, S.; Magari, M.; Kanayama, N.; Morishita, R.; Tokumitsu, H. Identification of striated muscle activator of Rho signaling (STARS) as a novel calmodulin target by a newly developed genome-wide screen. Cell Calcium 2016, 60, 32–40. [Google Scholar] [CrossRef]
- Sakane, K.; Nishiguchi, M.; Denda, M.; Yamagchi, F.; Magari, M.; Kanayama, N.; Morishita, R.; Tokumitsu, H. Identification and characterization of a centrosomal protein, FOR20 as a novel S100A6 target. Biochem. Biophys. Res. Commun. 2017, 491, 980–985. [Google Scholar] [CrossRef] [PubMed]
- Sakane, K.; Yamaguchi, F.; Tsuchiya, M.; Kondo, R.; Kanayama, N.; Magari, M.; Hatano, N.; Kobayashi, R.; Tokumitsu, H. Interaction of S100A6 with Target Proteins In Vitro and in Living Cells. Adv. Struct. Saf. Stud. 2019, 1929, 367–377. [Google Scholar] [CrossRef]
- Hayashi, N.; Matsubara, M.; Takasaki, A.; Titani, K.; Taniguchi, H. An expression system of rat calmodulin us-ing T7 phage promoter in Escherichia coli. Protein Expr. Purif. 1998, 12, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, J.-C.; Chao, C.-C.; Young, M.-J.; Chang, Y.-T.; Cho, E.-C.; Chang, W. A Poxvirus Host Range Protein, CP77, Binds to a Cellular Protein, HMG20A, and Regulates Its Dissociation from the Vaccinia Virus Genome in CHO-K1 Cells. J. Virol. 2006, 80, 7714–7728. [Google Scholar] [CrossRef]
- Lee, M.; Daniels, M.J.; Garnett, M.J.; Venkitaraman, A.R. A mitotic function for the high-mobility group protein HMG20b regulated by its interaction with the BRC repeats of the BRCA2 tumor suppressor. Oncogene 2011, 30, 3360–3369. [Google Scholar] [CrossRef]
- Yurimoto, S.; Hatano, N.; Tsuchiya, M.; Kato, K.; Fujimoto, T.; Masaki, T.; Kobayashi, R.; Tokumitsu, H. Identification and Characterization of Wolframin, the Product of the Wolfram Syndrome Gene (WFS1), as a Novel Calmodulin-Binding Protein. Biochemistry 2009, 48, 3946–3955. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Tokumitsu, H.; Hatano, N.; Tsuchiya, M.; Yurimoto, S.; Fujimoto, T.; Ohara, N.; Kobayashi, R.; Sakagami, H. Identification and characterization of PRG-1 as a neuronal calmodulin-binding protein. Biochem. J. 2010, 431, 81–91. [Google Scholar] [CrossRef]
- Sumoy, L.; Carim, L.; Escarceller, M.; Nadal, M.; Gratacòs, M.; Pujana, M.; Estivill, X.; Peral, B. HMG20A and HMG20B map to human chromosomes 15q24 and 19p13.3 and constitute a distinct class of HMG-box genes with ubiquitous expression. Cytogenet. Cell Genet. 2000, 88, 62–67. [Google Scholar] [CrossRef]
- Bianchi, M.E.; Agresti, A. HMG proteins: Dynamic players in gene regulation and differentiation. Curr. Opin. Genet. Dev. 2005, 15, 496–506. [Google Scholar] [CrossRef]
- Wynder, C.; Hakimi, M.-A.; Epstein, J.A.; Shilatifard, A.; Shiekhattar, R. Recruitment of MLL by HMG-domain protein iBRAF promotes neural differentiation. Nat. Cell Biol. 2005, 7, 1113–1117. [Google Scholar] [CrossRef] [PubMed]
- Ceballos-Chávez, M.; Rivero, S.; García-Gutiérrez, P.; Rodríguez-Paredes, M.; García-Domínguez, M.; Bhattacharya, S.S.; Reyes, J.C. Control of neuronal differentiation by sumoylation of BRAF35, a subunit of the LSD1-CoREST histone demethylase complex. Proc. Natl. Acad. Sci. USA 2012, 109, 8085–8090. [Google Scholar] [CrossRef] [PubMed]
- Rivero, S.; Ceballos-Chávez, M.; Bhattacharya, S.S.; Reyes, J.C. HMG20A is required for SNAI1-mediated epithelial to mesenchymal transition. Oncogene 2015, 34, 5264–5276. [Google Scholar] [CrossRef] [PubMed]
- Cross, S.S.; Hamdy, F.C.; Deloulme, J.C.; Rehman, I. Expression of S100 proteins in normal human tissues and common cancers using tissue microarrays: S100A6, S100A8, S100A9 and S100A11 are all overexpressed in common cancers. Histopathology 2005, 46, 256–269. [Google Scholar] [CrossRef]
- Tomas, A.; Moss, S.E. Calcium- and Cell Cycle-dependent Association of Annexin 11 with the Nuclear Envelope. J. Biol. Chem. 2003, 278, 20210–20216. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-T.; Dimitrova, Y.N.; Schneider, G.; Ridenour, W.B.; Bhattacharya, S.; Soss, S.E.; Caprioli, R.M.; Filipek, A.; Chazin, W.J. Structure of the S100A6 Complex with a Fragment from the C-Terminal Domain of Siah-1 Interacting Protein: A Novel Mode for S100 Protein Target Recognition. Biochemistry 2008, 47, 10921–10932. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sudo, T.; Hidaka, H. Regulation of Calcyclin (S100A6) Binding by Alternative Splicing in the N-terminal Regulatory Domain of Annexin XI Isoforms. J. Biol. Chem. 1998, 273, 6351–6357. [Google Scholar] [CrossRef] [PubMed]
- Nowotny, M.; Spiechowicz, M.; Jastrzebska, B.; Filipek, A.; Kitagawa, K.; Kuznicki, J. Calcium-regulated interac-tion of Sgt1 with S100A6 (calcyclin) and other S100 proteins. J. Biol. Chem. 2003, 278, 26923–26928. [Google Scholar] [CrossRef]
- Yatime, L.; Betzer, C.; Jensen, R.K.; Mortensen, S.; Jensen, P.H.; Andersen, G.R. The Structure of the RAGE:S100A6 Complex Reveals a Unique Mode of Homodimerization for S100 Proteins. Structure 2016, 24, 2043–2052. [Google Scholar] [CrossRef]
- Artegiani, B.; Labbaye, C.; Sferra, A.; Quaranta, M.T.; Torreri, P.; Macchia, G.; Ceccarini, M.; Petrucci, T.C.; Mac-ioce, P. The interaction with HMG20a/b proteins suggests a potential role for β-dystrobrevin in neuronal differentiation. J. Biol. Chem. 2010, 285, 24740–24750. [Google Scholar] [CrossRef] [PubMed]
- Yamada, J.; Jinno, S. S100A6 (calcyclin) is a novel marker of neural stem cells and astrocyte precursors in the subgranular zone of the adult mouse hippocampus. Hippocampus 2013, 24, 89–101. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S100A6 Targets | S100A6-Binding Sequence | Reference | |
|---|---|---|---|
| human HMG20A | 311 | DTIDSYMNRLHSIILANPQDNENFIATVREVV 342 | this study |
| mouse SIP | 189 | SEGLMNVLKKIYEDGDDDMKRTINKAWVESR 219 | [33] |
| human FOR20 | 1 | MATVAELKAVLKDTLEKKGVLGHLKARIRA 30 | [18] |
| rabbit Annexin XI-A | 49 | QDYLSGMAANMSGT 62 | [34] |
| human Sgt1 | 263 | KEEEKNEKLEGDAALNRLFQQIYSDGSDEVKRAM | [35] |
| NKSFMESGGTVLSTNWSDVGKRKVEINPPDDMEW | |||
| KKY 333 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamamoto, M.; Kondo, R.; Hozumi, H.; Doi, S.; Denda, M.; Magari, M.; Kanayama, N.; Hatano, N.; Morishita, R.; Tokumitsu, H. Identification and Biochemical Characterization of High Mobility Group Protein 20A as a Novel Ca2+/S100A6 Target. Biomolecules 2021, 11, 510. https://doi.org/10.3390/biom11040510
Yamamoto M, Kondo R, Hozumi H, Doi S, Denda M, Magari M, Kanayama N, Hatano N, Morishita R, Tokumitsu H. Identification and Biochemical Characterization of High Mobility Group Protein 20A as a Novel Ca2+/S100A6 Target. Biomolecules. 2021; 11(4):510. https://doi.org/10.3390/biom11040510
Chicago/Turabian StyleYamamoto, Maho, Rina Kondo, Haruka Hozumi, Seita Doi, Miwako Denda, Masaki Magari, Naoki Kanayama, Naoya Hatano, Ryo Morishita, and Hiroshi Tokumitsu. 2021. "Identification and Biochemical Characterization of High Mobility Group Protein 20A as a Novel Ca2+/S100A6 Target" Biomolecules 11, no. 4: 510. https://doi.org/10.3390/biom11040510
APA StyleYamamoto, M., Kondo, R., Hozumi, H., Doi, S., Denda, M., Magari, M., Kanayama, N., Hatano, N., Morishita, R., & Tokumitsu, H. (2021). Identification and Biochemical Characterization of High Mobility Group Protein 20A as a Novel Ca2+/S100A6 Target. Biomolecules, 11(4), 510. https://doi.org/10.3390/biom11040510