
In Vitro Preformed Biofilms of Bacillus safensis Inhibit the Adhesion and Subsequent Development of Listeria monocytogenes on Stainless-Steel Surfaces


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Evaluation of the Ability of B. safensis to Form Biofilms
2.1.1. Study Surface
2.1.2. Inoculum Preparation
2.1.3. Formation of Monospecies Biofilms In Vitro
2.1.4. Evaluation of Biofilm Formation through DEM
2.2. Evaluation of the Effect of Preformed B. safensis Biofilms on 22 Strains of L. monocytogenes
2.2.1. L. monocytogenes Strains
2.2.2. Inoculums Preparation
2.2.3. In Vitro Formation of Monospecies Biofilms and Subsequent Implantation of L. monocytogenes on the Structure
2.2.4. Quantification of Biofilms by ALOA, TEMPO, and qPCR
2.3. Evaluation of the Effect of Preformed Biofilms of Pseudomonas spp. and Candida zeylanoides on L. monocytogenes
2.4. Characterization of the Effect of Preformed B. safensis Biofilms on L. monocytogenes
2.5. Statistical Analysis
3. Results and Discussion
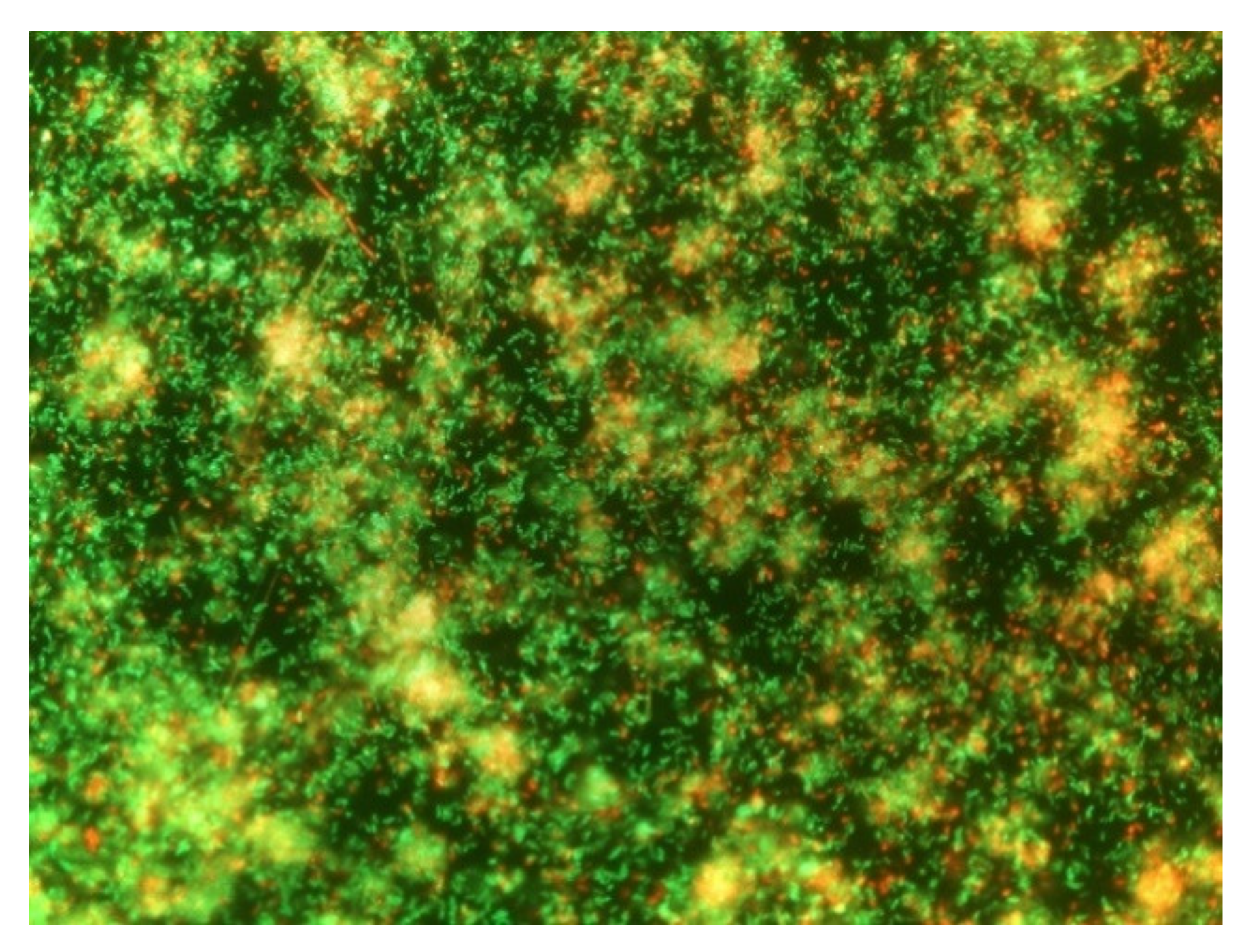
3.1. Evaluation of the Ability of B. safensis Biofilm Formation
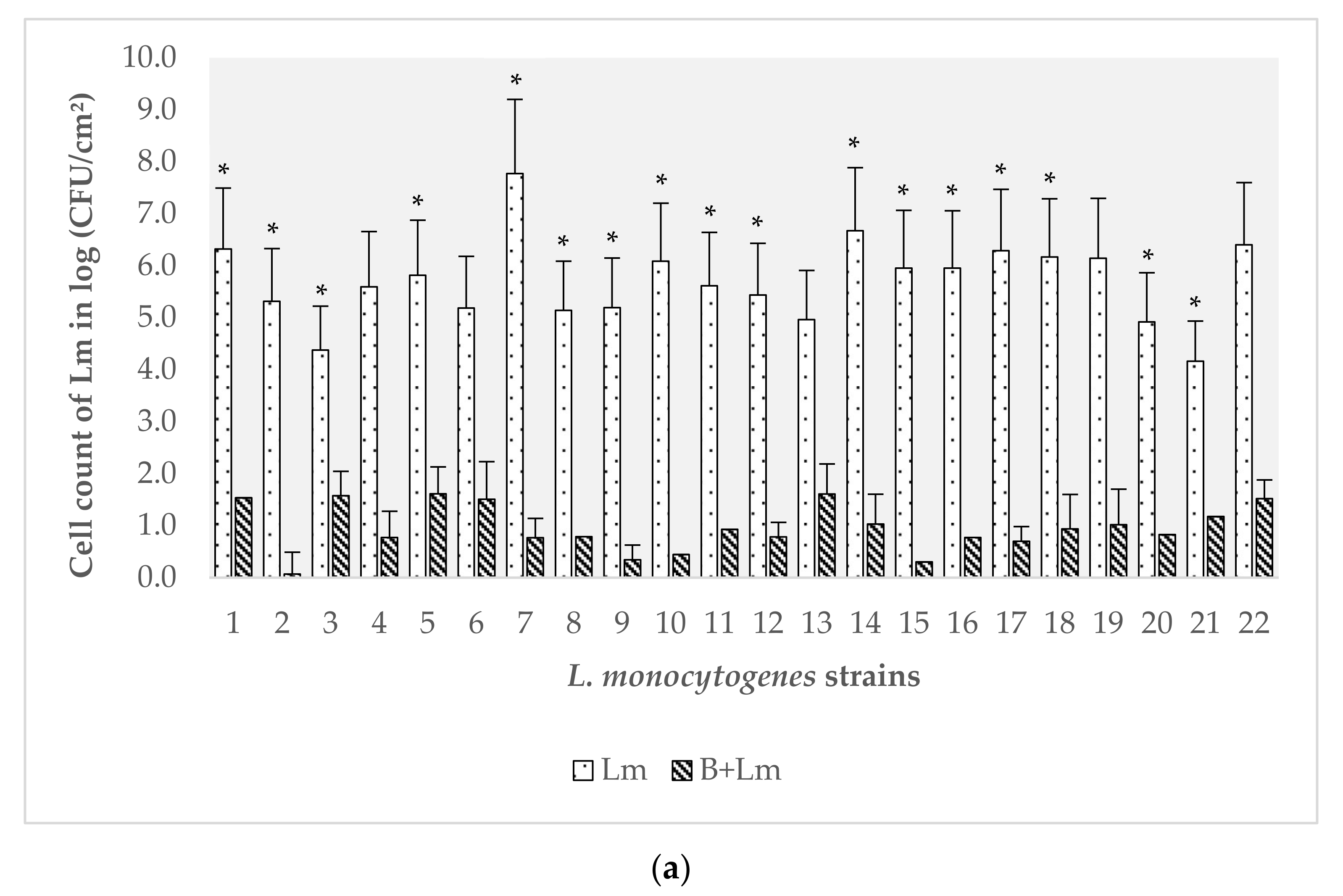
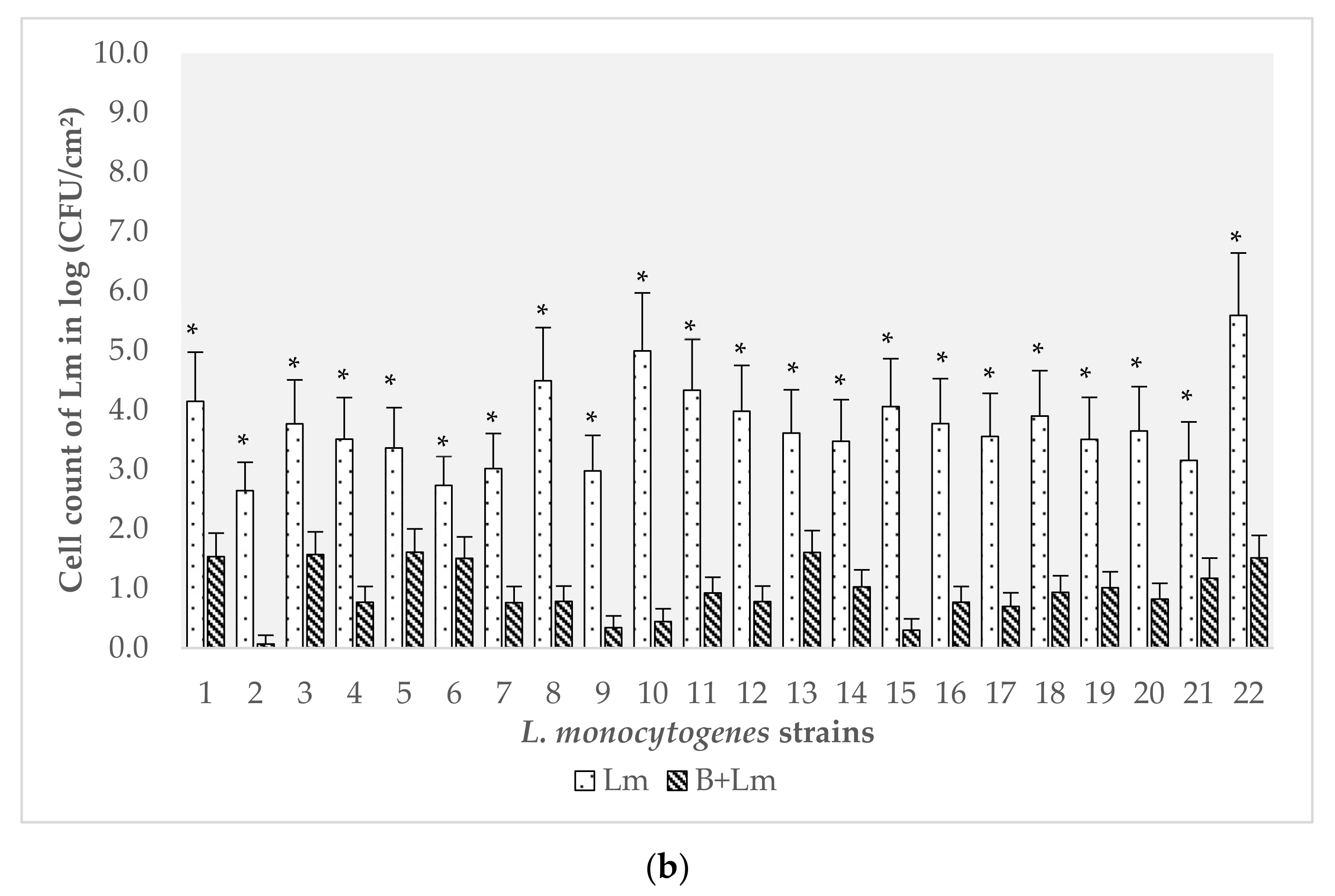
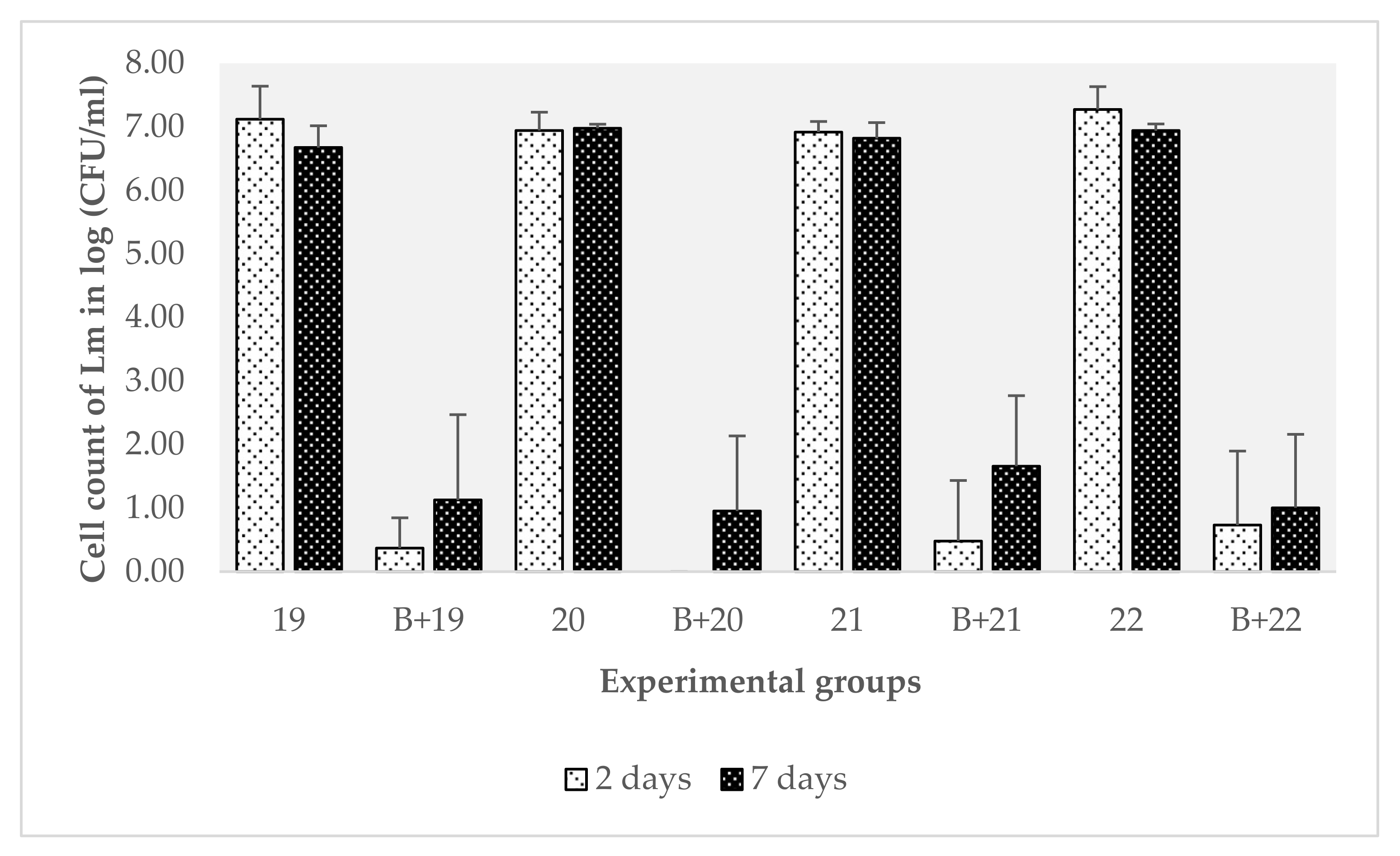
3.2. Evaluation of the Effect of B. safensis Preformed Biofilms on 22 Strains of L. monocytogenes
3.3. Evaluation of the Effect of Preformed Biofilms of Pseudomonas spp. and C. zeylanoides on L. monocytogenes
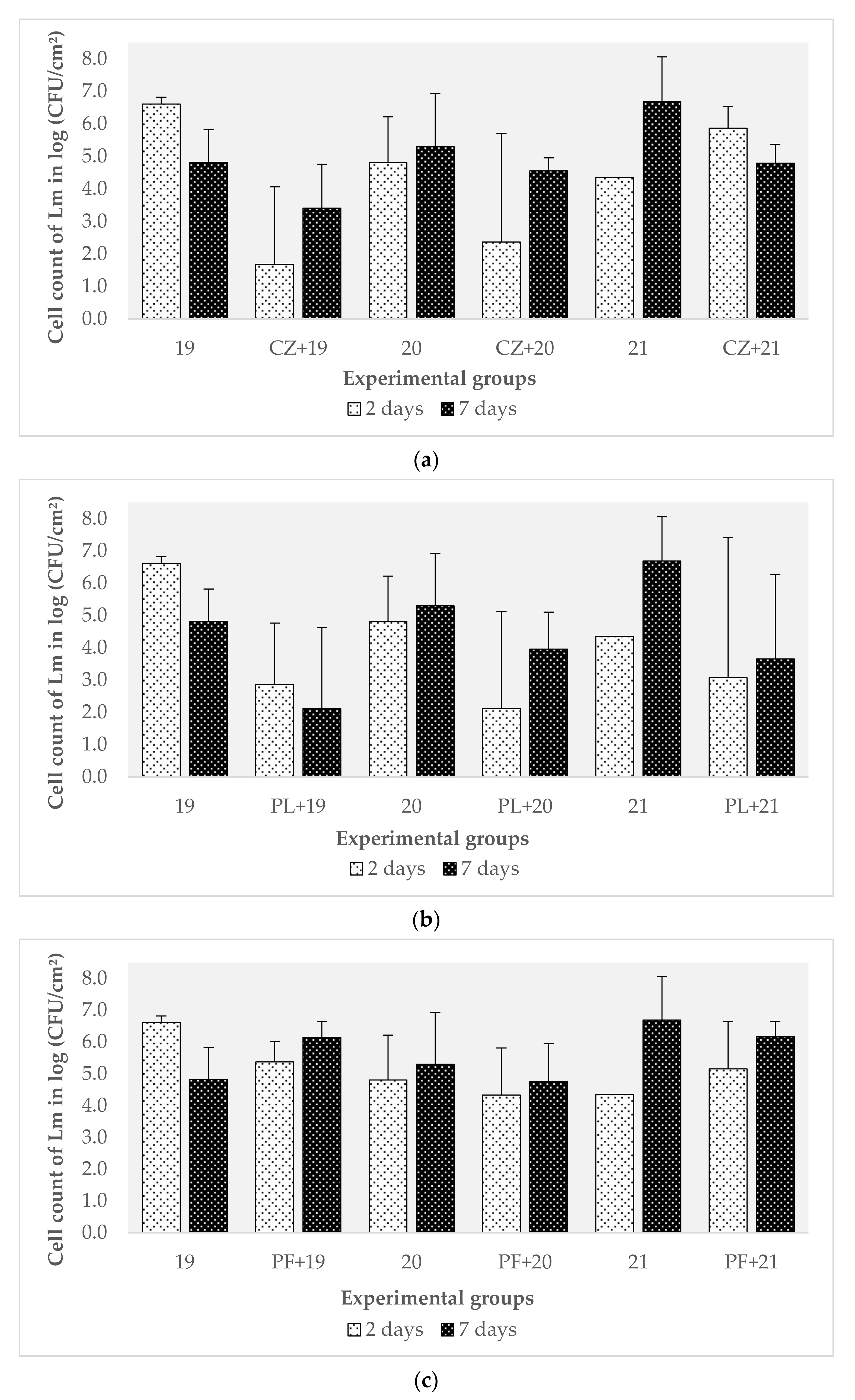
3.4. Characterization of the Effect of Preformed Biofilms of B. safensis on L. monocytogenes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carpentier, B.; Cerf, O. Review—Persistence of Listeria monocytogenes in food industry equipment and premises. Int. J. Food Microbiol. 2011, 145, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, M.; Chikindas, M.L. Listeria: A foodborne pathogen that knows how to survive. Int. J. Food Microbiol. 2007, 113, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Møretrø, T.; Langsrud, S.; Heir, E. Bacteria on meat abattoir process surfaces after sanitation: Characterisation of survival properties of Listeria monocytogenes and the commensal bacterial flora. Adv. Microbiol. 2013, 3, 255–264. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Listeriosis; ECDC: Stockholm, Sweden, 2020. [Google Scholar]
- EFSA; ECDC. The European Union One Health 2018 zoonoses report. EFSA J. 2019, 17. [Google Scholar] [CrossRef]
- Giaouris, E.; Heir, E.; Desvaux, M.; Hébraud, M.; Møretrø, T.; Langsrud, S.; Doulgeraki, A.; Nychas, G.J.; Kacániová, M.; Czaczyk, K.; et al. Intra- and inter-species interactions within biofilms of important foodborne bacterial pathogens. Front. Microbiol. 2015, 6, 1–26. [Google Scholar] [CrossRef]
- Ortiz, S.; López, V.; Villatoro, D.; López, P.; Dávila, J.C.; Martínez-Suárez, J.V.; Lo, V.; Da, J.C. A 3-year surveillance of the genetic diversity and persistence of Listeria monocytogenes in an iberian pig slaughterhouse and processing plant. Foodborne Pathog. Dis. 2010, 7, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Poimenidou, S.V.; Chrysadakou, M.; Tzakoniati, A.; Bikouli, V.C.; Nychas, G.-J.; Skandamis, P.N. Variability of Listeria monocytogenes strains in biofilm formation on stainless steel and polystyrene materials and resistance to peracetic acid and quaternary ammonium compounds. Int. J. Food Microbiol. 2016, 237, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Bagge-Ravn, D.; Gardshodn, K.; Gram, L.; Vogel, B.F. Comparison of sodium hypochlorite—based foam and peroxyacetic acid - based fog sanitizing procedures in a salmon smokehouse: Survival of the general microflora and Listeria monocytogenes. J. Food Prot. 2003, 66, 592–598. [Google Scholar] [CrossRef]
- Bagge-Ravn, D.; Ng, Y.; Hjelm, M.; Christiansen, J.N.; Johansen, C.; Gram, L. The microbial ecology of processing equipment in different fish industries—analysis of the microflora during processing and following cleaning and disinfection. Int. J. Food Microbiol. 2003, 87, 239–250. [Google Scholar] [CrossRef]
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Srey, S.; Jahid, I.K.; Ha, S. Do Biofilm formation in food industries: A food safety concern. Food Control 2013, 31, 572–585. [Google Scholar] [CrossRef]
- Palmieri, G.; Balestrieri, M.; Capuano, F.; Proroga, Y.T.R.; Pomilio, F.; Centorame, P.; Riccio, A.; Marrone, R.; Anastasio, A. Bactericidal and antibiofilm activity of bactenecin-derivative peptides against the food-pathogen Listeria monocytogenes: New perspectives for food processing industry. Int. J. Food Microbiol. 2018, 279, 33–42. [Google Scholar] [CrossRef]
- Hascoët, A.S.; Ripolles-Avila, C.; Guerrero-Navarro, A.E.; Rodríguez-Jerez, J.J. Microbial ecology evaluation of an iberian pig processing plant through implementing SCH sensors and the influence of the resident microbiota on Listeria monocytogenes. Appl. Sci. 2019, 9, 4611. [Google Scholar] [CrossRef]
- Carpentier, B.; Chassaing, D. Interactions in biofilms between Listeria monocytogenes and resident microorganisms from food industry premises. Int. J. Food Microbiol. 2004, 97, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Ripolles-Avila, C.; Hascoët, A.S.; Martínez-Suárez, J.V.; Capita, R.; Rodríguez-Jerez, J.J. Evaluation of the microbiological contamination of food processing environments through implementing surface sensors in an iberian pork processing plant: An approach towards the control of Listeria monocytogenes. Food Control 2019, 99, 40–47. [Google Scholar] [CrossRef]
- Leroi, F.; Cornet, J.; Chevalier, F.; Cardinal, M.; Coeuret, G.; Chaillou, S.; Joffraud, J. Selection of bioprotective cultures for preventing cold-smoked salmon spoilage. Int. J. Food Microbiol. 2015, 213, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Dortu, C.; Huch, M.; Holzapfel, W.H.; Franz, C.M.A.P.; Thonart, P. Anti-listerial activity of bacteriocin-producing Lactobacillus curvatus CWBI-B28 and Lactobacillus sakei CWBI-B1365 on raw beef and poultry meat. Lett. Appl. Microbiol. 2008, 47, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, C.; Vanegas, C.; Klotz, B. Antagonistic effect of Lactobacillus strains against Escherichia coli and Listeria monocytogenes in milk. J. Dairy Res. 2011, 78, 136–143. [Google Scholar] [CrossRef]
- Castellano, P.; Pérez Ibarreche, M.; Blanco Massani, M.; Fontana, C.; Vignolo, G. Strategies for pathogen biocontrol using lactic acid bacteria and their metabolites: A focus on meat ecosystems and industrial environments. Microorganisms 2017, 5, 38. [Google Scholar] [CrossRef]
- Chacon, R.; Martinez, R.; Dini Staliano, C.; Diogo, A.; Vieira, S.; Lissete, M.; Villarreal, M.; Todorov, S.D.; Marta, S.; Saad, I.; et al. Bacteriocin production and inhibition of Listeria monocytogenes by Lactobacillus sakei subsp. sakei 2a in a potentially synbiotic cheese spread. Food Microbiol. 2015, 48, 143–152. [Google Scholar] [CrossRef]
- Ortuño, R. Alertas y contaminación: La percepción del consumidor sobre la seguridad alimentaria. Available online: https://www.ainia.es/tecnoalimentalia/consumidor/percepcion-consumidor-seguridad-alimentaria/ (accessed on 21 January 2021).
- Loussaïf, L.; Moigno, E. La perception de l’offre responsable par le consommateur français. Une étude exploratoire dans la grande distribution alimentaire. Ethics Econ. 2012, 9, 162–179. [Google Scholar]
- Niewczas, M. Consumers’ reactions to food scares. Int. J. Consum. Stud. 2014, 38, 251–257. [Google Scholar] [CrossRef]
- Lateef, A.; Adelere, I.A.; Gueguim-Kana, E.B. The biology and potential biotechnological applications of Bacillus safensis. Biologia 2015, 70, 411–419. [Google Scholar] [CrossRef]
- Mayer, F.L.; Kronstad, J.W. Disarming fungal pathogens: Bacillus safensis inhibits virulence factor production and biofilm formation by Cryptococcus neoformans and Candida albicans. MBio 2017, 8, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Abdelli, F.; Jardak, M.; Elloumi, J.; Stien, D. Antibacterial, anti-adherent and cytotoxic activities of surfactin (s) from a lipolytic strain Bacillus safensis F4. Biodegradation 2019, 30, 287–300. [Google Scholar] [CrossRef] [PubMed]
- De Araujo, L.L.G.C.; Sodré, L.G.P.; Brasil, L.R.; Domingos, D.F.; de Oliveira, V.M.; da Cruz, G.F. Microbial enhanced oil recovery using a biosurfactant produced by Bacillus safensis isolated from mangrove microbiota—Part I biosurfactant characterization and oil displacement test. J. Pet. Sci. Eng. 2019, 180, 950–957. [Google Scholar] [CrossRef]
- AENOR ISO 13697. Antisépticos y Desinfectantes Químicos; Ensayo Cuantitativo de Superficie no Porosa Para la Evaluación de la Actividad Bactericida y/o Fungicida de los Desinfectantes Químicos Utilizados en Productos Alimenticios, en la Industria, en el Hogar y en Colectividad; AENOR: Madrid, Spain, 2015; p. 35. [Google Scholar]
- Fuster-Valls, N. Importancia del Control Higiénico de las Superficies Alimentarias Mediante Técnicas Rápidas y Tradicionales Para Evitar y/o Minimizar las Contaminaciones Cruzadas; Universidad Autónoma de Barcelona: Bellaterra, Spain, 2006. [Google Scholar]
- Ripolles-Avila, C.; Hascoët, A.S.; Guerrero-Navarro, A.E.; Rodríguez-Jerez, J.J. Establishment of incubation conditions to optimize the in vitro formation of mature Listeria monocytogenes biofilms on food-contact surfaces. Food Control 2018, 92, 240–248. [Google Scholar] [CrossRef]
- Ortiz, S.; López-alonso, V.; Rodríguez, P. The connection between persistent, disinfectant-resistant Listeria monocytogenes strains from two geographically separate Iberian pork processing plants: Evidence from comparative genome analysis. Appl. Environ. Microbiol. 2016, 82, 308–317. [Google Scholar] [CrossRef]
- López, V.; Ortiz, S.; Corujo, A.; López, P.; Poza, D.; Navas, J.; Moreno, R.; Martínez-Suárez, J.V. Different contamination patterns of lineage I and II strains of Listeria monocytogenes in a Spanish broiler abattoir. Poult. Sci. 2008, 87, 1874–1882. [Google Scholar] [CrossRef]
- López, V.; Ortiz, S.; Corujo, A.; López, P.; Navas, J.; Moreno, R.; Martínez-Suárez, J.V. Traceback identification of an ingredient (pork dewlap) as the possible source of Listeria monocytogenes serotype 4b contamination in raw chicken products. J. Food Prot. 2007, 70, 1513–1517. [Google Scholar] [CrossRef]
- López, V.; Navas, J.; Martínez-Suárez, J.V. Low potential virulence associated with mutations in the inlA and prfA Genes in Listeria monocytogenes isolated from raw retail poultry meat. J. Food Prot. 2013, 76, 129–132. [Google Scholar] [CrossRef]
- Ortiz, S.; López, V.; Martínez-Suárez, J.V. The influence of subminimal inhibitory concentrations of benzalkonium chloride on biofilm formation by Listeria monocytogenes. Int. J. Food Microbiol. 2014, 189, 106–112. [Google Scholar] [CrossRef]
- Ripolles-Avila, C. Supervivencia de Listeria monocytogenes Sobre Superficies de Contacto con Alimentos: Un Abordaje Multidisciplinar de un Problema Complejo; Universitat Autònoma de Barcelona: Bellaterra, Spain, 2018. [Google Scholar]
- Langsrud, S.; Moen, B.; Møretrø, T.; Løype, M.; Heir, E. Microbial dynamics in mixed culture biofilms of bacteria surviving sanitation of conveyor belts in salmon-processing plants. J. Appl. Microbiol. 2016, 120, 366–378. [Google Scholar] [CrossRef]
- Leriche, V.; Carpentier, B. Limitation of adhesion and growth of Listeria monocytogenes on stainless steel surfaces by Staphylococcus sciuri biofilms. J. Appl. Microbiol. 2000, 88, 594–605. [Google Scholar] [CrossRef]
- Phillips, C.A. Bacterial biofilms in food processing environments: A review of recent developments in chemical and biological control. Int. J. Food Sci. Technol. 2016, 51, 1731–1743. [Google Scholar] [CrossRef]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as complex differentiated communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef]
- Coughlan, L.M.; Cotter, P.D.; Hill, C.; Alvarez-Ordóñez, A. New weapons to fight old enemies: Novel strategies for the (Bio)control of bacterial biofilms in the food industry. Front. Microbiol. 2016, 7, 1–21. [Google Scholar] [CrossRef]
- Winkelströter, L.K.; Gomes, B.C.; Thomaz, M.R.S.; Souza, V.M.; Martinis, E.C.P. De Lactobacillus sakei 1 and its bacteriocin influence adhesion of Listeria monocytogenes on stainless steel surface. Food Control 2011, 22, 1404–1407. [Google Scholar] [CrossRef]
- Pérez-Ibarreche, M.; Castellano, P.; Leclercq, A.; Vignolo, G. Control of Listeria monocytogenes biofilms on industrial surfaces by the bacteriocin-producing Lactobacillus sakei CRL1862. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef]
- Dunn, K.A.; Bielawski, J.P.; Ward, T.J.; Urquhart, C.; Gu, H. Reconciling ecological and genomic divergence among lineages of Listeria under an “extended mosaic genome concept”. Mol. Biol. Evol. 2009, 26, 2605–2615. [Google Scholar] [CrossRef]
- Ghoul, M.; Mitri, S. The ecology and evolution of microbial competition. Trends Microbiol. 2016, 24, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, U.T.; Burrows, L.L. DNase I and proteinase K impair Listeria monocytogenes biofilm formation and induce dispersal of pre-existing biofilms. Int. J. Food Microbiol. 2014, 187, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Mellefont, L.A.; McMeekin, T.A.; Ross, T. Effect of relative inoculum concentration on Listeria monocytogenes growth in co-culture. Int. J. Food Microbiol. 2008, 121, 157–168. [Google Scholar] [CrossRef]
- Lourenço, A.; Machado, H.; Brito, L. Biofilms of Listeria monocytogenes produced at 12 °C either in pure culture or in co-culture with Pseudomonas aeruginosa showed reduced susceptibility to sanitizers. J. Food Sci. 2011, 76, 142–148. [Google Scholar] [CrossRef]
- Puga, C.H.; Dahdouh, E.; SanJose, C.; Orgaz, B. Listeria monocytogenes colonizes Pseudomonas fluorescens biofilms and induces matrix over-production. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Puga, C.H.; Orgaz, B.; SanJose, C. Listeria monocytogenes impact on mature or old Pseudomonas fluorescens biofilms during growth at 4 and 20 °C. Front. Microbiol. 2016, 7, 1–9. [Google Scholar] [CrossRef]
- Heir, E.; Møretrø, T.; Simensen, A.; Langsrud, S. Listeria monocytogenes strains show large variations in competitive growth in mixed culture biofilms and suspensions with bacteria from food processing environments. Int. J. Food Microbiol. 2018, 275, 46–55. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Strain | Serotype | Origin |
|---|---|---|---|
| 1 | 4423 | 1/2a | [32] |
| 2 | 5873 | 1/2a | CECT |
| 3 | A7 | 1/2a | [33] |
| 4 | CDL69 | 1/2a | [32] |
| 5 | EGD-e | 1/2a | [32] |
| 6 | P12 | 1/2a | [34] |
| 7 | R6 | 1/2a | [35] |
| 8 | S1(R) | 1/2a | [36] |
| 9 | S1(S) | 1/2a | [36] |
| 10 | S2-1 | 1/2a | [36] |
| 11 | S2-2 | 1/2a | [32] |
| 12 | S2bac | 1/2a | [36] |
| 13 | 5366 | 4b | CECT |
| 14 | 5672 | 4b | CECT |
| 15 | 935 | 4b | CECT |
| 16 | S10-1 | 2a | [32] |
| 17 | 911 | 1/2c | CECT |
| 18 | Lm 1 | 4b | Isolated from an industrial meat processing environment in 2017 [16] |
| 19 | Lm 2 | 4b | |
| 20 | Lm 3 | 1/2a | |
| 21 | Lm 4 | 1/2a | |
| 22 | Lm 5 | 1/2b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hascoët, A.-S.; Ripolles-Avila, C.; Cervantes-Huamán, B.R.H.; Rodríguez-Jerez, J.J. In Vitro Preformed Biofilms of Bacillus safensis Inhibit the Adhesion and Subsequent Development of Listeria monocytogenes on Stainless-Steel Surfaces. Biomolecules 2021, 11, 475. https://doi.org/10.3390/biom11030475
Hascoët A-S, Ripolles-Avila C, Cervantes-Huamán BRH, Rodríguez-Jerez JJ. In Vitro Preformed Biofilms of Bacillus safensis Inhibit the Adhesion and Subsequent Development of Listeria monocytogenes on Stainless-Steel Surfaces. Biomolecules. 2021; 11(3):475. https://doi.org/10.3390/biom11030475
Chicago/Turabian StyleHascoët, Anne-Sophie, Carolina Ripolles-Avila, Brayan R. H. Cervantes-Huamán, and José Juan Rodríguez-Jerez. 2021. "In Vitro Preformed Biofilms of Bacillus safensis Inhibit the Adhesion and Subsequent Development of Listeria monocytogenes on Stainless-Steel Surfaces" Biomolecules 11, no. 3: 475. https://doi.org/10.3390/biom11030475
APA StyleHascoët, A.-S., Ripolles-Avila, C., Cervantes-Huamán, B. R. H., & Rodríguez-Jerez, J. J. (2021). In Vitro Preformed Biofilms of Bacillus safensis Inhibit the Adhesion and Subsequent Development of Listeria monocytogenes on Stainless-Steel Surfaces. Biomolecules, 11(3), 475. https://doi.org/10.3390/biom11030475