
SIRT1 and SIRT2 Modulators: Potential Anti-Inflammatory Treatment for Depression?

Abstract
1. Background
1.1. Current Medication Treatments for Depression and Their Limitations
1.2. SIRT1 and SIRT2’s Role in Inflammation
2. Association of SIRT1, SIRT2 Polymorphisms and Depression
3. SIRT1 and SIRT2 Modulators in In Vitro Studies of Inflammation
4. SIRT1 and SIRT2 Modulators in In Vivo Studies of Depression
5. Outlook of the Treatment Strategy for Depression
Author Contributions
Funding
Conflicts of Interest
References
- Murray, C.J.L.; Lopez, A.D. A comprehensive assessment of mortality and disability from diseases, injuries, and risk factors in 1990 and projected to 2020. In The Global Burden of Disease; Harvard School of Public Health on behalf of the World Health Organization and the World Bank: Boston, MA, USA, 1996. [Google Scholar]
- World Health Organization. Depression Key Facts. Available online: https://www.who.int/news-room/fact-sheets/detail/depression (accessed on 20 November 2020).
- Roy, A.; Campbell, M.K. A unifying framework for depression: Bridging the major biological and psychosocial theories through stress. Clin. Investig. Med. 2013, 36, 170–E190. [Google Scholar] [CrossRef]
- Kohler, O.; Krogh, J.; Mors, O.; Benros, M.E. Inflammation in Depression and the Potential for Anti-Inflammatory Treatment. Curr. Neuropharmacol. 2016, 14, 732–742. [Google Scholar] [CrossRef]
- Loonen, A.J.M.; Ivanova, S.A. Circuits Regulating Pleasure and Happiness—Mechanisms of Depression. Front. Hum. Neurosci. 2016, 10, 571. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.; Koenen, K.C.; Aiello, A.E.; Wildman, D.E.; Santos, R.D.L.; Galea, S. Epigenetic and inflammatory marker profiles associated with depression in a community-based epidemiologic sample. Psychol. Med. 2010, 41, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Schiepers, O.J.; Wichers, M.C.; Maes, M. Cytokines and major depression. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2005, 29, 201–217. [Google Scholar] [CrossRef]
- Miller, A.H.; Raison, C.L. The role of inflammation in depression: From evolutionary imperative to modern treatment target. Nat. Rev. Immunol. 2016, 16, 22–34. [Google Scholar] [CrossRef]
- Penn, E.; Tracy, D.K. The drugs don’t work? antidepressants and the current and future pharmacological management of depression. Ther. Adv. Psychopharmacol. 2012, 2, 179–188. [Google Scholar] [CrossRef]
- National Institute of Mental Health. Sequenced Treatment Alternatives to Relieve Depression (STAR*D) Study; NIMH: Rockville, MD, USA, 2006.
- Trivedi, M.H.; Rush, A.J.; Wisniewski, S.R.; Nierenberg, A.A.; Warden, D.; Ritz, L.; Norquist, G.; Howland, R.H.; Lebowitz, B.; McGrath, P.J.; et al. Evaluation of Outcomes With Citalopram for Depression Using Measurement-Based Care in STAR*D: Implications for Clinical Practice. Am. J. Psychiatry 2006, 163, 28–40. [Google Scholar] [CrossRef]
- Gaynes, B.N.; Warden, D.; Trivedi, M.H.; Wisniewski, S.R.; Fava, M.; Rush, A.J. What did STAR*D teach us? Results from a large-scale, practical, clinical trial for patients with depression. Psychiatr. Serv. 2009, 60, 1439–1445. [Google Scholar] [CrossRef] [PubMed]
- Rush, A.J.; Trivedi, M.H.; Wisniewski, S.R.; Stewart, J.W.; Nierenberg, A.A.; Thase, M.E.; Ritz, L.; Biggs, M.M.; Warden, D.; Luther, J.F.; et al. Bupropion-SR, Sertraline, or Venlafaxine-XR after Failure of SSRIs for Depression. N. Engl. J. Med. 2006, 354, 1231–1242. [Google Scholar] [CrossRef]
- Trivedi, M.H.; Fava, M.; Wisniewski, S.R.; Thase, M.E.; Quitkin, F.; Warden, D.; Ritz, L.; Nierenberg, A.A.; Lebowitz, B.D.; Biggs, M.M.; et al. Medication Augmentation after the Failure of SSRIs for Depression. N. Engl. J. Med. 2006, 354, 1243–1252. [Google Scholar] [CrossRef]
- Thase, M.E.; Friedman, E.S.; Biggs, M.M.; Wisniewski, S.R.; Trivedi, M.H.; Luther, J.F.; Fava, M.; Nierenberg, A.A.; McGrath, P.J.; Warden, D.; et al. Cognitive therapy versus medication in augmentation and switch strategies as second-step treatments: A STAR*D report. Am. J. Psychiatry 2007, 164, 739–752. [Google Scholar] [CrossRef]
- Fava, M.; Rush, A.J.; Wisniewski, S.R.; Nierenberg, A.A.; Alpert, J.E.; McGrath, P.J.; Thase, M.E.; Warden, D.; Biggs, M.; Luther, J.F.; et al. A comparison of mirtazapine and nortriptyline following two consecutive failed medication treatments for depressed outpatients: A STAR*D report. Am. J. Psychiatry 2006, 163, 1161–1172. [Google Scholar] [CrossRef] [PubMed]
- Nierenberg, A.A.; Fava, M.; Trivedi, M.H.; Wisniewski, S.R.; Thase, M.E.; McGrath, P.J.; Alpert, J.E.; Warden, D.; Luther, J.F.; Niederehe, G.; et al. A Comparison of Lithium and T3Augmentation Following Two Failed Medication Treatments for Depression: A STAR*D Report. Am. J. Psychiatry 2006, 163, 1519–1530. [Google Scholar] [CrossRef] [PubMed]
- McGrath, P.J.; Stewart, J.W.; Fava, M.; Trivedi, M.H.; Wisniewski, S.R.; Nierenberg, A.A.; Thase, M.E.; Davis, L.; Biggs, M.M.; Shores-Wilson, K.; et al. Tranylcypromine Versus Venlafaxine Plus Mirtazapine Following Three Failed Antidepressant Medication Trials for Depression: A STAR*D Report. Am. J. Psychiatry 2006, 163, 1531–1541. [Google Scholar] [CrossRef]
- Huynh, N.N.; McIntyre, R.S. What are the implications of the STAR*D trial for primary care? a review and synthesis. Prim. Care Companion J. Clin. Psychiatry 2008, 10, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Landry, J.; Sutton, A.; Tafrov, S.T.; Heller, R.C.; Stebbins, J.; Pillus, L.; Sternglanz, R. The silencing protein SIR2 and its homologs are NAD-dependent protein deacetylases. Proc. Natl. Acad. Sci. USA 2000, 97, 5807–5811. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Liu, J.; Chen, D.; Yan, L.; Zheng, W. Sirtuin Inhibition: Strategies, Inhibitors, and Therapeutic Potential. Trends Pharmacol. Sci. 2017, 38, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Li, J.; Zhang, H.; Zhao, X.; Yan, L.-J.; Yang, X. Role and Possible Mechanisms of Sirt1 in Depression. Oxidative Med. Cell. Longev. 2018, 2018, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Brites, D.; Fernandes, A. Neuroinflammation and Depression: Microglia Activation, Extracellular Microvesicles and microRNA Dysregulation. Front. Cell. Neurosci. 2015, 9, 476. [Google Scholar] [CrossRef]
- Xie, J.; Zhang, X.; Zhang, L. Negative regulation of inflammation by SIRT. Pharmacol. Res. 2013, 67, 60–67. [Google Scholar] [CrossRef]
- Paraíso, A.F.; Mendes, K.L.; Santos, S.H.S. Brain Activation of SIRT1: Role in Neuropathology. Mol. Neurobiol. 2013, 48, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Mucke, L. Paths of Convergence: Sirtuins in Aging and Neurodegeneration. Neuron 2008, 58, 10–14. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Chiang, H.-H.; Luo, H.; Zheng, Z.; Qiao, Q.; Wang, L.; Tan, M.; Ohkubo, R.; Mu, W.-C.; Zhao, S.; et al. An Acetylation Switch of the NLRP3 Inflammasome Regulates Aging-Associated Chronic Inflammation and Insulin Resistance. Cell Metab. 2020, 31, 580–591. [Google Scholar] [CrossRef]
- Buechler, N.; Wang, X.; Yoza, B.K.; McCall, C.E.; Vachharajani, V. Sirtuin 2 Regulates Microvascular Inflammation during Sepsis. J. Immunol. Res. 2017, 2017, 1–9. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, Y.; Cao, W.; Wei, X.; Chen, J.; Ying, W. SIRT2 plays significant roles in lpopolysaccharides-induced neuroinflammation and brain injury in mice. Neurochem. Res. 2016, 41, 2490–2500. [Google Scholar] [CrossRef]
- Lavu, S.; Boss, O.; Elliott, P.J.; Lambert, P.D. Sirtuins—Novel therapeutic targets to treat age-associated diseases. Nat. Rev. Drug Discov. 2008, 7, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Chen, S.; Liang, L.; Li, X.; Tang, P.; Rao, X.; Pan, M.; Xu, X.; Li, Y.; Yao, Y.; et al. SIRT1 inhibitors mitigate radiation-induced GI syndrome by enhancing intestinal-stem-cell survival. Cancer Lett. 2021, 501, 20–30. [Google Scholar] [CrossRef]
- Frazzi, R. SIRT1 in Secretory Organ Cancer. Front. Endocrinol. 2018, 9, 569. [Google Scholar] [CrossRef]
- Wang, T.-W.; Chern, E.; Hsu, C.-W.; Tseng, K.-C.; Chao, H.-M. SIRT1-Mediated Expression of CD24 and Epigenetic Suppression of Novel Tumor Suppressor miR-1185-1 Increases Colorectal Cancer Stemness. Cancer Res. 2020, 80, 5257–5269. [Google Scholar] [CrossRef]
- van der Meer, A.J.; Scicluna, B.P.; Moerland, P.D.; Lin, J.; Jacobson, E.W.; Vlasuk, G.P.; van der Poll, T. The Selective Sirtuin 1 Activator SRT2104 Reduces Endotoxin-Induced Cytokine Release and Coagulation Activation in Humans. Crit. Care Med. 2015, 43, e199–e202. [Google Scholar] [CrossRef]
- Krueger, J.G.; Suárez-Fariñas, M.; Cueto, I.; Khacherian, A.; Matheson, R.; Parish, L.C.; Leonardi, C.; Shortino, D.; Gupta, A.; Haddad, J.; et al. A Randomized, Placebo-Controlled Study of SRT2104, a SIRT1 Activator, in Patients with Moderate to Severe Psoriasis. PLoS ONE 2015, 10, e0142081. [Google Scholar] [CrossRef] [PubMed]
- Dean, J.; Keshavan, M. The neurobiology of depression: An integrated view. Asian J. Psychiatry 2017, 27, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Shadrina, M.; Bondarenko, E.A.; Slominsky, P.A. Genetics Factors in Major Depression Disease. Front. Psychiatry 2018, 9, 334. [Google Scholar] [CrossRef]
- Cai, N.; Bigdeli, T.B.; Kretzschmar, W.; Li, Y.; Liang, J.; Song, L. Sparse whole-genome sequencing identifies two loci for major depressive disorder. Nature 2015, 523, 588–591. [Google Scholar] [CrossRef]
- Liu, W.; Yan, H.; Zhou, D.; Cai, X.; Zhang, Y.; Li, S.; Zhou, D.-S.; Li, X.; Zhang, C.; Sun, Y.; et al. The depression GWAS risk allele predicts smaller cerebellar gray matter volume and reduced SIRT1 mRNA expression in Chinese population. Transl. Psychiatry 2019, 9, 333. [Google Scholar] [CrossRef]
- Hirata, T.; Otsuka, I.; Okazaki, S.; Mouri, K.; Horai, T.; Boku, S.; Takahashi, M.; Ueno, Y.; Sora, I.; Shirakawa, O.; et al. Major depressive disorder-associated SIRT1 locus affects the risk for suicide in women after middle age. Psychiatry Res. 2019, 278, 141–145. [Google Scholar] [CrossRef]
- Kishi, T.; Yoshimura, R.; Kitajima, T.; Okochi, T.; Okumura, T.; Tsunoka, T.; Yamanouchi, Y.; Kinoshita, Y.; Kawashima, K.; Fukuo, Y. SIRT1 gene is associated with major depressive disorder in the Japanese population. J. Affect. Disord. 2010, 126, 167–173. [Google Scholar] [CrossRef]
- Porcelli, S.; Salfi, R.; Politis, A.; Atti, A.R.; Albani, D.; Chierchia, A.; Polito, L.; Zisaki, A.; Piperi, C.; Liappas, I.; et al. Association between Sirtuin 2 gene rs10410544 polymorphism and depression in Alzheimer’s disease in two independent European samples. J. Neural Transm. 2013, 120, 1709–1715. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.-C.; Duan, K.-M.; Fang, C.; Li, D.-Y.; Zheng, S.-S.; Yang, S.-Q.; Yang, S.-T.; Yang, M.; Zhang, L.-B.; Wang, S.-Y. Correlations Between SIRT Genetic Polymorphisms and Postpartum Depressive Symptoms in Chinese Parturients Who Had Undergone Cesarean Section. Neuropsychiatr. Dis. Treat. 2020, 16, 3225–3238. [Google Scholar] [CrossRef]
- Himmerich, H.; Patsalos, O.; Lichtblau, N.; Ibrahim, M.A.A.; Dalton, B. Cytokine Research in Depression: Principles, Challenges, and Open Questions. Front. Psychiatry 2019, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, D.D.S.; Pacheco, M.T.; Frozza, R.L.; Bernardi, A. Anti-Inflammatory Effects of Resveratrol: Mechanistic Insights. Int. J. Mol. Sci. 2018, 19, 1812. [Google Scholar] [CrossRef]
- Hoffmann, E.; Wald, J.; Lavu, S.; Roberts, J.; Beaumont, C.; Haddad, J.; Elliott, P.; Westphal, C.; Jacobson, E. Pharmacokinetics and tolerability of SRT2104, a first-in-class small molecule activator of SIRT1, after single and repeated oral administration in man. Br. J. Clin. Pharmacol. 2012, 75, 186–196. [Google Scholar] [CrossRef]
- Donmez, G.; Outeiro, T.F. SIRT1 and SIRT2: Emerging targets in neurodegeneration. EMBO Mol. Med. 2013, 5, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Xu, Z.M.; Lu, L.Y.; Nie, H.; Ding, J.; Ying, W.H.; Tian, H.L. SIRT2 inhibition exacerbates neuroinflammation and blood-brain barrier disruption in experimental traumatic brain injury by enhancing NF-κB p65 acetylation and activation. J. Neurochem. 2016, 136, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Romeo-Guitart, D.; Leiva-Rodríguez, T.; Espinosa-Alcantud, M.; Sima, N.; Vaquero, A.; Martín, H.D.-; Ruano, D.; Casas, C. SIRT1 activation with neuroheal is neuroprotective but SIRT2 inhibition with AK7 is detrimental for disconnected motoneurons. Cell Death Dis. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Lee, Y.G.; Reader, B.F.; Herman, D.; Streicher, A.; Englert, J.A.; Ziegler, M.; Chung, S.; Karpurapu, M.; Park, G.Y.; Christman, J.W.; et al. Sirtuin 2 enhances allergic asthmatic inflammation. JCI Insight 2019, 4. [Google Scholar] [CrossRef]
- Orecchia, A.; Scarponi, C.; Di Felice, F.; Cesarini, E.; Avitabile, S.; Mai, A.; Mauro, M.L.; Sirri, V.; Zambruno, G.; Albanesi, C.; et al. Sirtinol treatment reduces inflammation in human dermal microvascular endothelial cells. PLoS ONE 2011, 6, e24307. [Google Scholar] [CrossRef]
- Dai, H.; Sinclair, D.A.; Ellis, J.L.; Steegborn, C. Sirtuin activators and inhibitors: Promises, achievements, and challenges. Pharmacol. Ther. 2018, 188, 140–154. [Google Scholar] [CrossRef]
- Abe-Higuchi, N.; Uchida, S.; Yamagata, H.; Higuchi, F.; Hobara, T.; Hara, K.; Kobayashi, A.; Watanabe, Y. Hippocampal Sirtuin 1 Signaling Mediates Depression-like Behavior. Biol. Psychiatry 2016, 80, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Borges, G.; Berrocoso, E.; Mico, J.A.; Neto, F. ERK1/2: Function, signaling and implication in pain and pain-related anxio-depressive disorders. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2015, 60, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, Q.; Cai, Y.; Sun, D.; He, X.; Wang, L.; Yu, D.; Li, X.; Xiong, X.; Xu, H.; et al. Resveratrol counteracts lipopolysaccharide-induced depressive-like behaviors via enhanced hippocampal neurogenesis. Oncotarget 2016, 7, 56045–56059. [Google Scholar] [CrossRef]
- Kim, H.-D.; Hesterman, J.; Call, T.; Magazu, S.; Keeley, E.; Armenta, K.; Kronman, H.; Neve, R.L.; Nestler, E.J.; Ferguson, D. SIRT1 Mediates Depression-Like Behaviors in the Nucleus Accumbens. J. Neurosci. 2016, 36, 8441–8452. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.-M.; Zhang, J.-R.; Wan, T.-F.; Wang, Y.; Chen, H.-S.; Liu, L. SRT2104 attenuates chronic unpredictable mild stress-induced depressive-like behaviors and imbalance between microglial M1 and M2 phenotypes in the mice. Behav. Brain Res. 2020, 378, 112296. [Google Scholar] [CrossRef]
- Ferland, C.L.; Hawley, W.R.; Puckett, R.E.; Wineberg, K.; Lubin, F.D.; Dohanich, G.P.; Schrader, L.A. Sirtuin Activity in Dentate Gyrus Contributes to Chronic Stress-Induced Behavior and Extracellular Signal-Regulated Protein Kinases 1 and 2 Cascade Changes in the Hippocampus. Biol. Psychiatry 2013, 74, 927–935. [Google Scholar] [CrossRef]
- Erburu, M.; Muñoz-Cobo, I.; Diaz-Perdigon, T.; Mellini, P.; Suzuki, T.; Puerta, E.; Tordera, R.M. SIRT2 inhibition modulate glutamate and serotonin systems in the prefrontal cortex and induces antidepressant-like action. Neuropharmacology 2017, 117, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Erburu, M.; Muñoz-Cobo, I.; Domínguez-Andrés, J.; Beltran, E.; Suzuki, T.; Mai, A.; Valente, S.; Puerta, E.; Tordera, R. Chronic stress and antidepressant induced changes in Hdac5 and Sirt2 affect synaptic plasticity. Eur. Neuropsychopharmacol. 2015, 25, 2036–2048. [Google Scholar] [CrossRef] [PubMed]
- Palanza, P. Animal models of anxiety and depression: How are females different? Neurosci. Biobehav. Rev. 2001, 25, 219–233. [Google Scholar] [CrossRef]
- Eid, R.S.; Gobinath, A.R.; Galea, L.A. Sex differences in depression: Insights from clinical and preclinical studies. Prog. Neurobiol. 2019, 176, 86–102. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Wang, J.; Wang, D.; Li, C.; Liu, B.; Fang, X.; You, J.; Guo, M.; Lu, X.-Y. SIRT1 in forebrain excitatory neurons produces sexually dimorphic effects on depression-related behaviors and modulates neuronal excitability and synaptic transmission in the medial prefrontal cortex. Mol. Psychiatry 2020, 25, 1094–1111. [Google Scholar] [CrossRef]
- Eyre, H.; Stuart, M.; Baune, B. A phase-specific neuroimmune model of clinical depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 54, 265–274. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Single-Nucleotide Polymorphism (SNP) | Population | Study Type | Subject Number | Impact |
|---|---|---|---|---|
| rs1245800 (SIRT1) | Chinese women with major depressive disorder (MDD) | Genome-wide association study (GWAS) | 9000 | SNP rs1245800 contributes to the risk of MDD [38]. |
| rs1245800 | Chinese Han population | GWAS | 4855 | SNP rs1245800 contributes to reduced SIRT1 expression and cerebellar grey matter volume [39]. |
| rs1245800 and rs4746720 (SIRT1) | Japanese women | GWAS | 1538 | SNP rs1245800 and rs4746720 associate with suicide. No association with SIRT1 expression in the dorsolateral prefrontal cortex [40]. |
| rs10997875 (SIRT1) | Japanese population | Case-control study | 1221 | SNP rs10997875 associates with MDD [41]. |
| rs10410544 (SIRT2) | Alzheimer’s disease (AD) patients in European populations | Case-control study | 1172 | T/T SNP genotype might exert protection against depressive symptoms in AD [42]. |
| rs2873703, rs4801933 (SIRT2) | Chinese women undergoing caesarean section | Case-control study | 368 | T/T genotype SNPs are correlated with the onset of postpartum depressive symptoms [43]. |
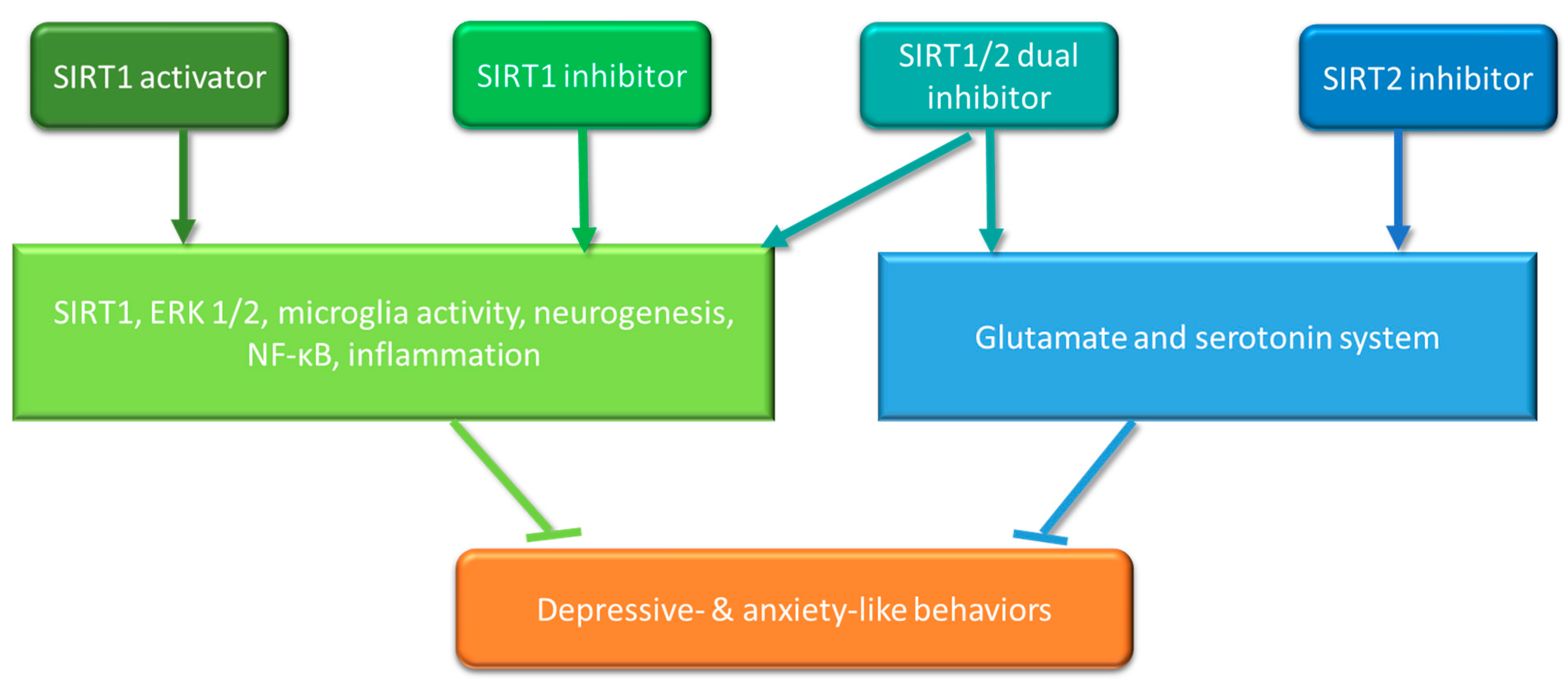
| SIRT1/SIRT2 Modulators | Depressive-Like In Vivo Models | Administration Route | Results |
|---|---|---|---|
| Resveratrol (SIRT1 activator) | Adult male BALB mice + chronic ultra-mild stress | Intraperitoneal (i.p.) injection | Depression-like and anxiety-like behaviours ↓, SIRT1 activation ↑, phosphorylation of ERK1/2 ↑ [53]. * |
| Adult male C57/BL6 mice + LPS challenge | i.p. | Depressive-like behaviours ↓, microglia activation ↓, neurogenesis ↑, SIRT1 ↑, NF-κB ↓ [55]. | |
| Adult male C57/BL6 mice + chronic social defeat stress | Bilateral infusion into nucleus accumbens | Depressive- and anxiety-like behaviours ↑ [56]. | |
| SRT2104 (SIRT1 activator) | Adult male C57BL/6 mice + chronic unpredictable mild stress | Bilateral infusion into hippocampus | Depressive-like behaviour ↓, IL-6, IL-1β and iNOS ↓, IL-10, TNF-β and Abrignase1 ↑, microglia polarization ↓ via GSK3β/PTEN signalling pathway [57]. |
| Adult male BALB mice + repeated restraint stress | Bilateral injection into dentate gyrus | Social interaction ↑, sucrose preference ↑, phosphorylation of ERK1/2 ↑ [53]. | |
| EX527 (SIRT1 inhibitor) | Adult male C57/BL6 mice + chronic social defeat stress | Bilateral infusion into nucleus accumbens | Depressive- and anxiety-like behaviours ↓ [56]. |
| Adult male BALB mice + chronic ultra-mild stress | Bilateral injection into hippocampus | Social interaction ↓, latency to feed ↑ [53]. | |
| Sirtinol (SIRT1/2 dual inhibitor) | Adult male BALB mice + chronic ultra-mild stress | Bilateral injection into dentate gyrus | Social interaction ↓, latency to feed ↑, immobility time ↑ [53]. |
| Adult male Wistar rats + chronic variable stress | Infusion to dentate gyrus | Development of anhedonia ↓, stress-induced deficits in spatial memory ↓, ERK1/2 activity, Bcl-2 expression and histone acetylation ↑ [58]. | |
| 33i (SIRT2 inhibitor) | Adult male C57BL/6J mice + chronic mild stress | i.p. | Stress-induced anhedonia and social avoidance ↓ via regulating glutamate and serotonin system in the prefrontal cortex [59]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Anoopkumar-Dukie, S.; Davey, A.K. SIRT1 and SIRT2 Modulators: Potential Anti-Inflammatory Treatment for Depression? Biomolecules 2021, 11, 353. https://doi.org/10.3390/biom11030353
Zhang Y, Anoopkumar-Dukie S, Davey AK. SIRT1 and SIRT2 Modulators: Potential Anti-Inflammatory Treatment for Depression? Biomolecules. 2021; 11(3):353. https://doi.org/10.3390/biom11030353
Chicago/Turabian StyleZhang, Yuqing, Shailendra Anoopkumar-Dukie, and Andrew Keith Davey. 2021. "SIRT1 and SIRT2 Modulators: Potential Anti-Inflammatory Treatment for Depression?" Biomolecules 11, no. 3: 353. https://doi.org/10.3390/biom11030353
APA StyleZhang, Y., Anoopkumar-Dukie, S., & Davey, A. K. (2021). SIRT1 and SIRT2 Modulators: Potential Anti-Inflammatory Treatment for Depression? Biomolecules, 11(3), 353. https://doi.org/10.3390/biom11030353