
Ethylene-Cytokinin Interaction Determines Early Defense Response of Wheat against Stagonospora nodorum Berk.

 ,
,  , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant and Fungi Materials and Growth Conditions
2.2. Experimental Design
2.3. Isolation of RNA and Performing the Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.4. Biochemical Parameters
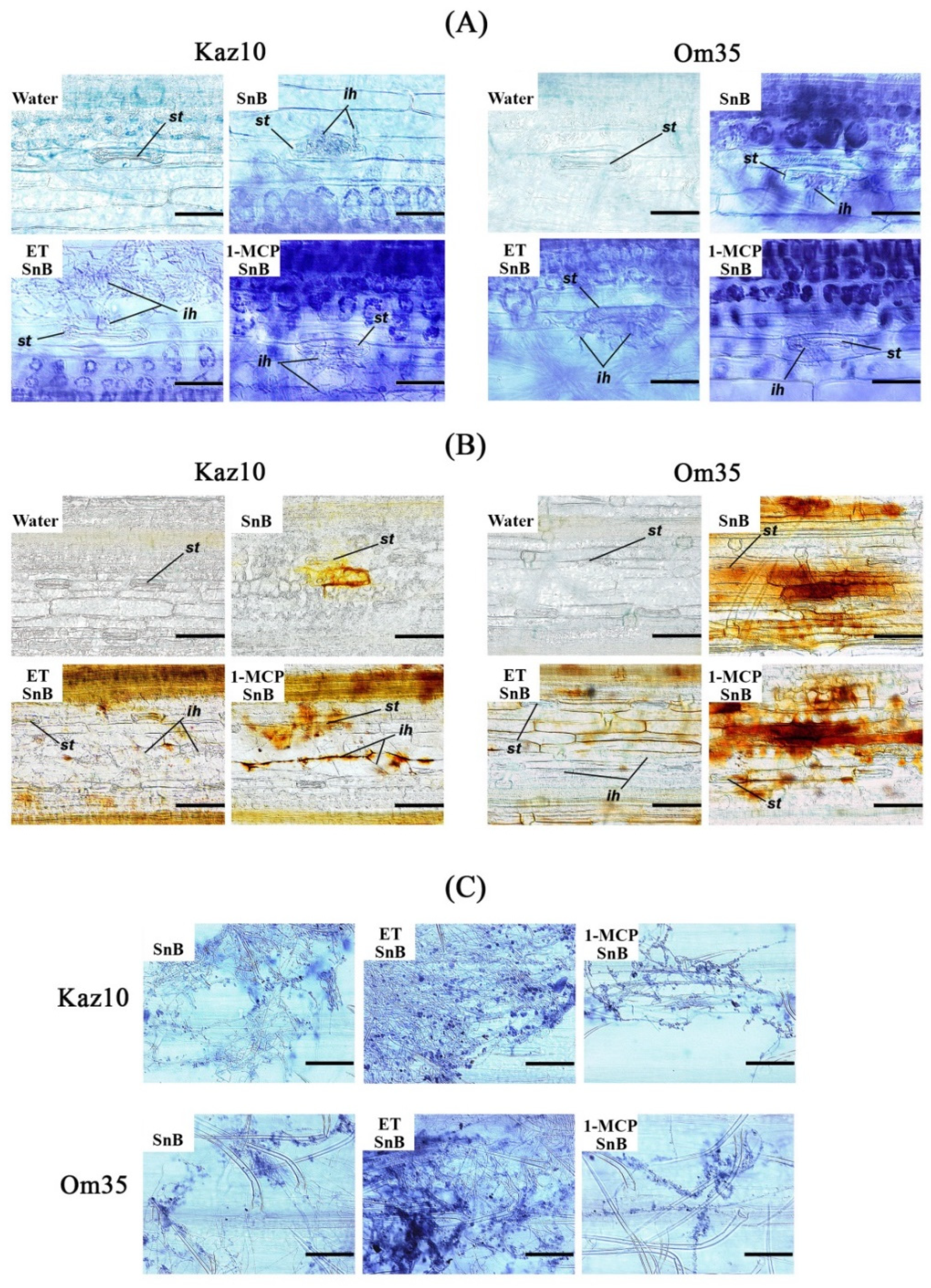
2.5. Visualization of H2O2, Superoxide Radical, and Fungal Mycelium in Wheat Tissues
2.6. Determination of Cytokinines
2.7. Statistics
3. Results
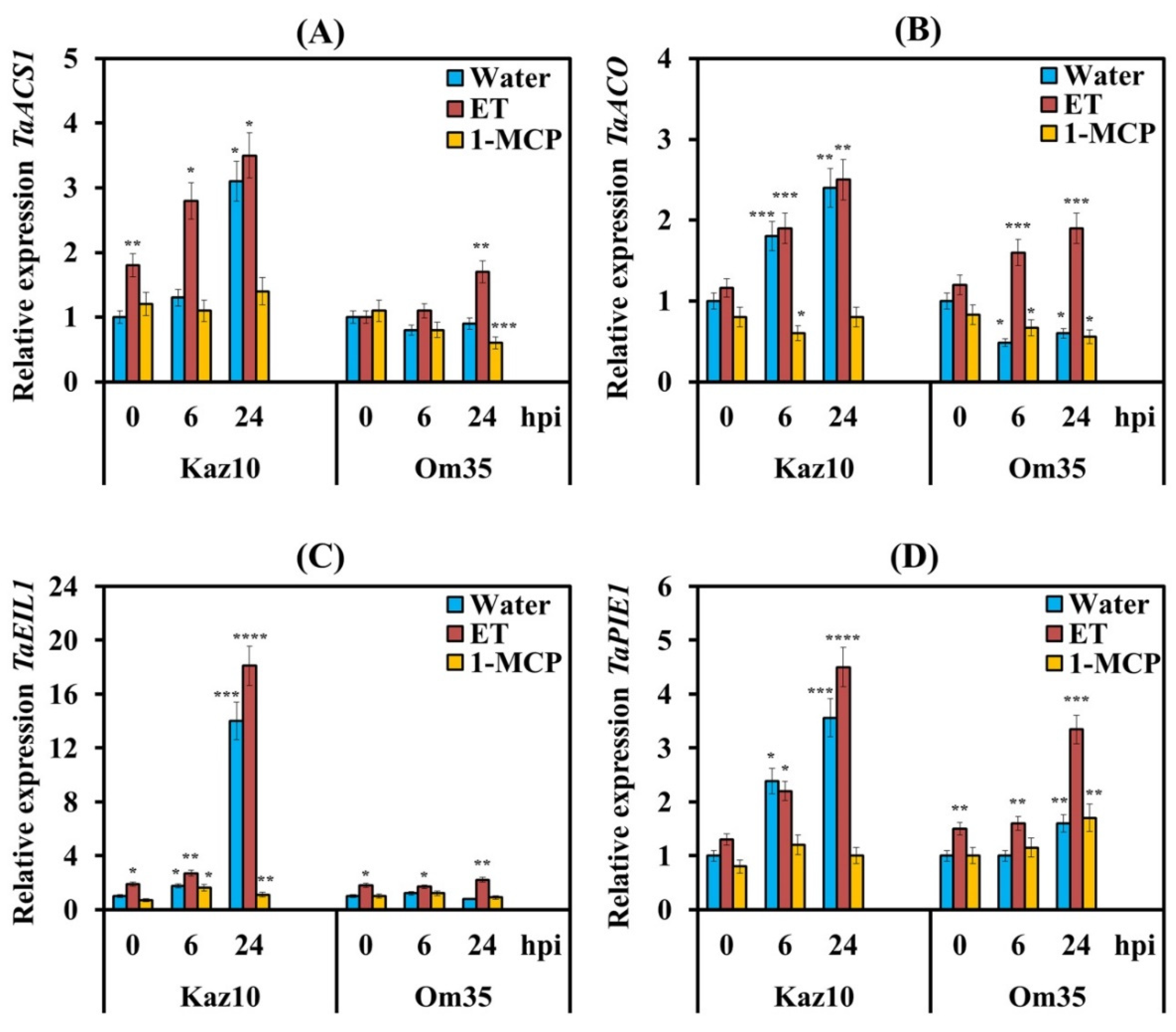
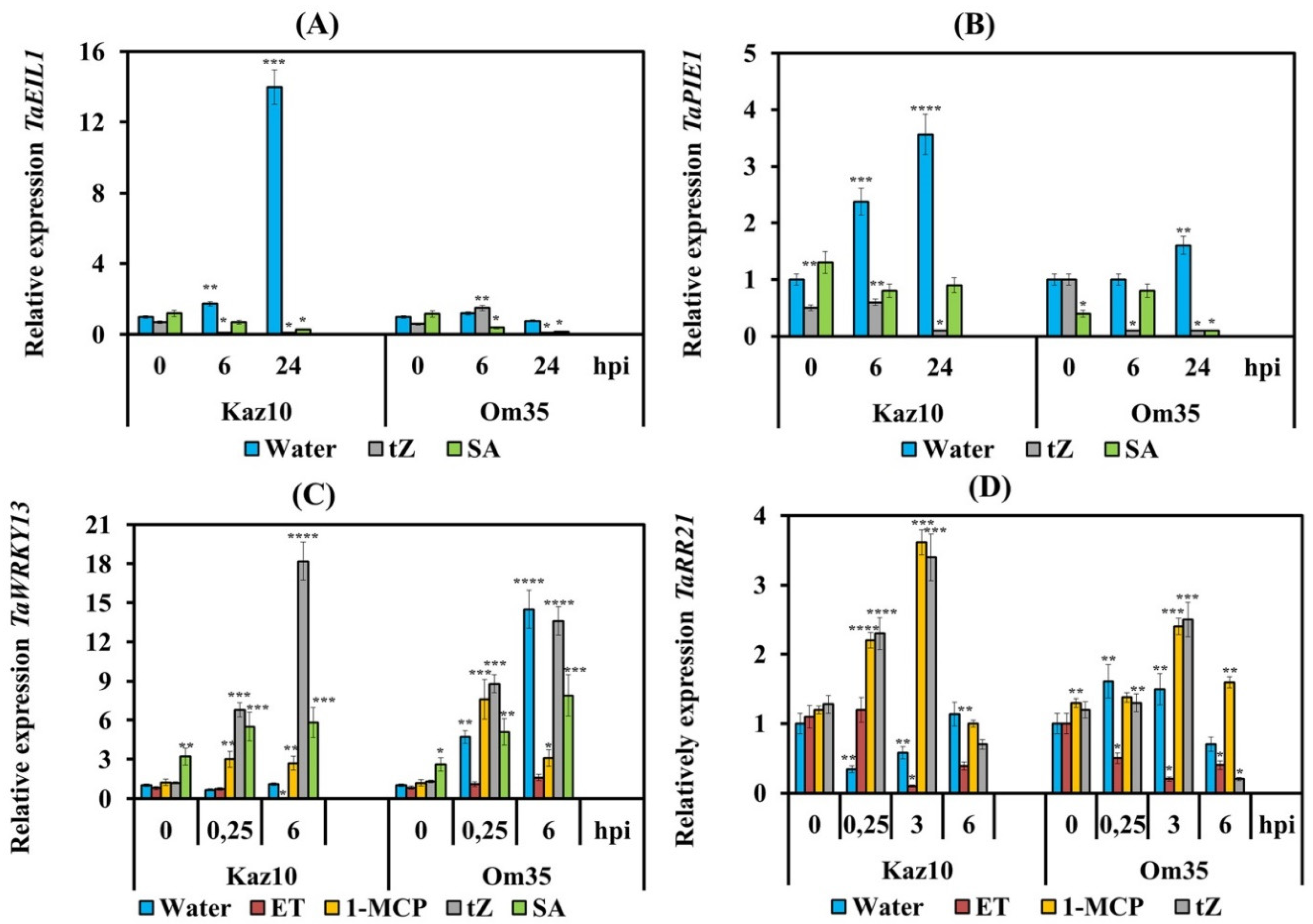
3.1. Biosynthesis and Signaling Pathway of Ethylene in Wheat Is Activated by the Effector Stagonospora nodorum Berk
3.2. SnTox3 Suppresses Oxidative Burst through the Ethylene Signaling Pathway by Regulating the Work of NADPH Oxidase and Provides the Growth of the Pathogen
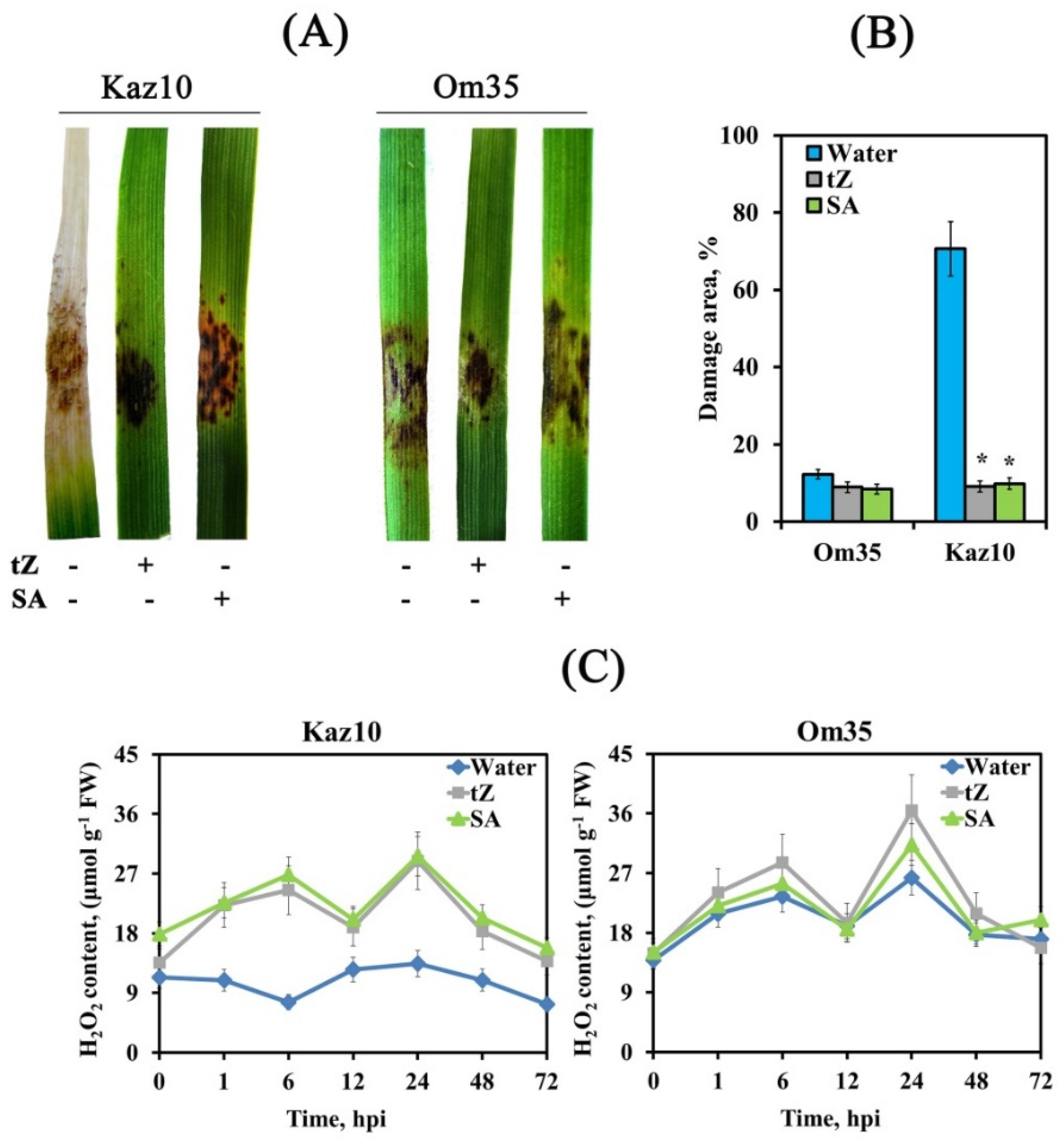
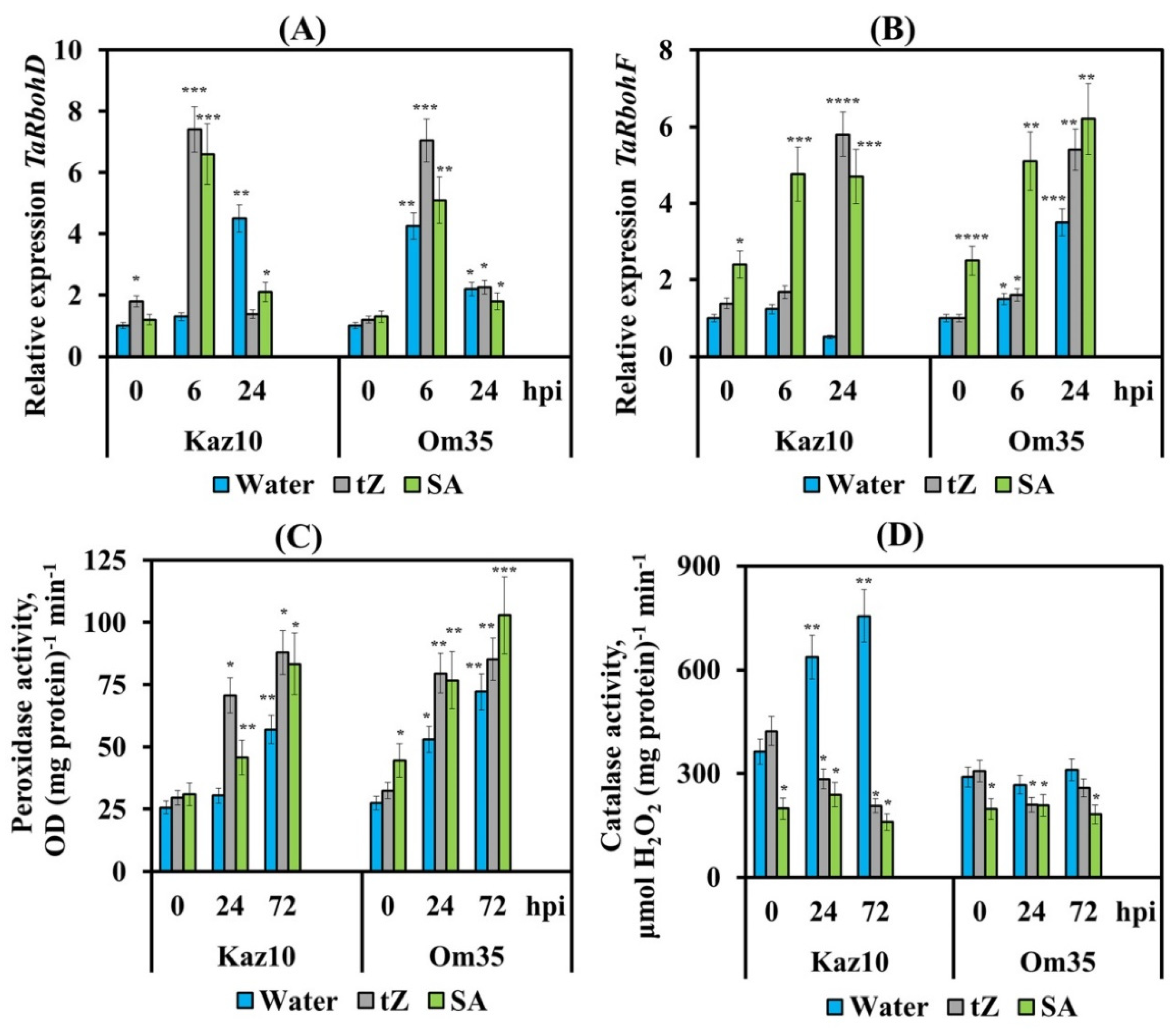
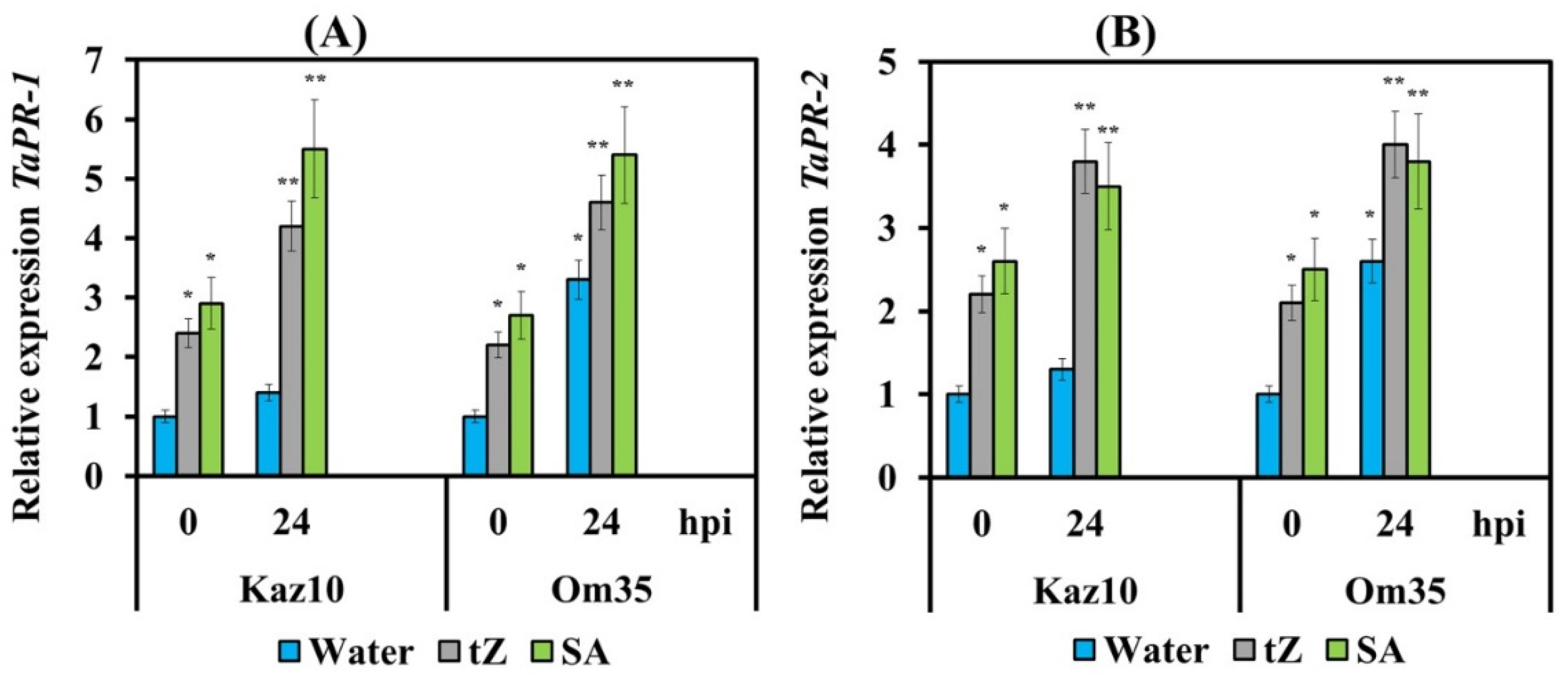
3.3. Cytokinins Enhance the Oxidative Burst, Limit the Growth of the Pathogen and Trigger the Salicylate Signaling Pathway
3.4. SnTox3 and Ethylene Regulate Biosynthesis and Metabolism of Cytokinins During Defense Response of Wheat against S. nodorum
3.5. Interaction of Signaling Pathways of Ethylene, CK, and SA
4. Discussion
4.1. Stagonospora nodorum NE SnTox3 Induces Biosynthesis and Signaling Pathway of Ethylene
4.2. Ethylene and Cytokinins Regulate ROS Production in Wheat Plants Infected with S. nodorum
4.2.1. Ethylene Promotes Penetration and Growth of the S. nodorum Fungus in Wheat Tissues
4.2.2. Ethylene Inhibited Biphasic H2O2 Production and Cytokinins Induced It at Early Stages of Infection with S. nodorum
4.2.3. SnTox3, Ethylene and Cytokinins Regulate the Work of Redox Enzymes in Wheat Plants Infected with S. nodorum
4.3. SnTox3 and Ethylene Inhibited Biosynthesis, Modulated Metabolism, and Activated Oxidative Degradation of Cytokinins
4.3.1. SnTox3 and Ethylene Regulated Cytokinin Biosynthesis in Wheat Plants Infected with S. nodorum
4.3.2. SnTox3 and Ethylene Activated Oxidative Degradation of Cytokinins in Wheat Plants Infected with S. nodorum
4.3.3. SnTox3 and Ethylene Modulated Metabolism of Cytokinins in Wheat Plants Infected with S. nodorum
4.4. Ethylene Induced by the Effector SnTox3 Suppresses the SA Signaling Pathway and Cytokinins Trigger it in Wheat Plants Infected with S. nodorum
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Robert-Seilaniantz, A.; Grantm, M.; Jones, J.D. Hormone crosstalk in plant disease and defense: More than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, S.S.; Mekureyaw, M.F.; Pandey, C.; Roitsch, T. Role of cytokinins for interactions of plants with microbial pathogens and pest insects. Front. Plant Sci. 2020, 10, 1777. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dang, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Oliver, R.P.; Solomon, P.S. New developments in pathogenicity and virulence of necrotrophs. Curr. Opin. Plant Biol. 2010, 13, 415–419. [Google Scholar] [CrossRef]
- Friesen, T.L.; Faris, J.D. Characterization of the wheat-Stagonospora nodorum disease system: What is the molecular basis of this quantitative necrotrophic disease interaction? Can. J. Plant Pathol. 2010, 32, 20–28. [Google Scholar] [CrossRef]
- Shi, G.; Zhang, Z.; Friesen, T.L.; Raats, D.; Fahima, T.; Brueggeman, R.S.; Lu, S.; Trick, H.N.; Liu, Z.; Chao, W.; et al. The hijacking of a receptor kinase–driven pathway by a wheat fungal pathogen leads to disease. Sci. Adv. 2016, 2, e1600822. [Google Scholar] [CrossRef]
- Phan, H.T.T.; Rybak, K.; Furuki, E.; Breen, S.; Solomon, P.S.; Oliver, R.P.; Tan, K.C. Differential effector gene expression underpins epistasis in a plant fungal disease. Plant J. 2016, 87, 343–354. [Google Scholar] [CrossRef]
- McDonald, M.C.; Solomon, P.S. Just the surface: Advances in the discovery and characterization of necrotrophic wheat effectors. Curr. Opin. Microbiol. 2018, 46, 14–18. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Z.; Faris, J.D.; Oliver, R.P.; Syme, R.; McDonald, M.C.; McDonald, B.A.; Solomon, P.S.; Lu, S.; Shelver, W.L.; et al. The cysteine rich necrotrophic effector SnTox1 produced by Stagonospora nodorum triggers susceptibility of wheat lines harboring Snn1. PLoS Pathog. 2012, 8, e1002467. [Google Scholar] [CrossRef]
- Jwa, N.-S.; Hwang, B.K. Convergent evolution of pathogen effectors toward reactive oxygen species signaling networks in plants. Front. Plant Sci. 2017, 8, 1687. [Google Scholar] [CrossRef]
- Shigenaga, A.M.; Berens, M.L.; Tsuda, K.; Argueso, C.T. Towards engineering of hormonal crosstalk in plant immunity. Curr. Opin. Plant Biol. 2017, 38, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Vleesschauwer, D.; Yang, Y.; Cruz, C.V.; Hofte, M. Abscisic acid-induced resistance against the brown spot pathogen Cochliobolus miyabeanus in rice involves MAP kinase-mediated repression of ethylene signaling. Physiol. Plant 2010, 152, 2036–2052. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K.; Lyons, R. Intervention of phytohormone pathways by pathogen effectors. Plant Cell 2014, 26, 2285–2309. [Google Scholar] [CrossRef] [PubMed]
- Broekgaarden, C.; Caarls, L.; Vos, I.A.; Pieterse, C.M.J.; van Wees, S.C.M. Ethylene: Traffic controller on hormonal crossroads to defense. Plant Physiol. 2015, 169, 2371–2379. [Google Scholar] [CrossRef] [PubMed]
- Spoel, S.H.; Johnson, J.S.; Dong, X. Regulation of tradeoffs between plant defenses against pathogens with different lifestyles. Proc. Natl. Acad. Sci. USA 2007, 104, 18842–18847. [Google Scholar] [CrossRef]
- Chen, X.; Steed, A.; Travella, S.; Keller, B.; Nicholson, P. Fusarium graminearum exploits ethylene signalling to colonize dicotyledonous and monocotyledonous plants. New Phytol. 2009, 182, 975–983. [Google Scholar] [CrossRef]
- Choi, J.; Huh, S.U.; Kojima, M.; Sakakibara, H.; Paek, K.H.; Hwang, I. The Cytokinin-activated transcription factor ARR2 promotes plant immunity via TGA3/NPR1-dependent salicylic acid signaling in Arabidopsis. Dev. Cell 2010, 19, 284–295. [Google Scholar] [CrossRef]
- Argueso, C.T.; Ferreira, F.J.; Epple, P.; To, J.P.C.; Hutchison, C.E.; Schaller, G.E.; Dangl, J.L.; Kieber, J.J. Two-component elements mediate interactions between cytokinin and salicylic acid in plant immunity. PLoS Genet. 2012, 8, e1002448. [Google Scholar] [CrossRef]
- Großkinsky, D.K.; Albacete, A.; Jammer, A.; Krbez, P.; van der Graaff, E.; Pfeifhofer, H.; Roitsch, T. A rapid phytohormone and phytoalexin screening method for physiological phenotyping. Mol. Plant 2014, 7, 1053–1056. [Google Scholar] [CrossRef]
- Shigenaga, A.; Argueso, C. No hormone to rule them all: Interactions of plant hormones during the responses of plants to pathogens. Semin. Cell Dev. Biol. 2016, 56, 174–189. [Google Scholar] [CrossRef]
- Sharipova, G.V.; Veselov, D.S.; Kudoyarova, G.R.; Timergalin, M.D.; Wilkinson, S. Effect of ethylene perception inhibitor on growth, water relations, and abscisic acid content in wheat plants under water deficit. Russ. J. Plant Physiol. 2012, 59, 573–580. [Google Scholar] [CrossRef]
- Smets, R.; Le, J.; Prinsen, E.; Verbelen, J.-P.; van Onckelen, H.A. Cytokinin-induced hypocotyl elongation in light-grown Arabidopsis plants with inhibited ethylene action or indole-3-acetic acid transport. Planta 2005, 221, 39–47. [Google Scholar] [CrossRef]
- O’Brien, J.A.; Benková, E. Cytokinin cross-talking during biotic and abiotic stress responses. Front. Plant Sci. 2013, 4, 451. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Kahmann, R. Manipulation of phytohormone pathways by effectors of filamentous plant pathogens. Front. Plant Sci. 2019, 10, 822. [Google Scholar] [CrossRef]
- Winterberg, B.; Du Fall, L.A.; Song, X.M.; Pascovici, D.; Care, N.; Molloy, M.; Ohms, S.; Solomon, P.S. The necrotrophic effector protein SnTox3 re-programs metabolism and elicits a strong defence response in susceptible wheat leaves. BMC Plant Biol. 2014, 14, 215. [Google Scholar] [CrossRef] [PubMed]
- Breen, S.; Williams, S.J.; Outram, M.; Kobe, B.; Solomon, P.S. Emerging insights into the functions of pathogenesis related protein. Trends Plant Sci. 2017, 22, 10. [Google Scholar] [CrossRef] [PubMed]
- Veselova, S.V.; Nuzhnaya, T.V.; Maksimov, I.V. The Effect of 1-methylcyclopropene on the components of pro and antioxidant systems of wheat and the development of defense reactions in fungal pathogenesis. Appl. Biochem. Microbiol. 2014, 50, 516–523. [Google Scholar] [CrossRef]
- Veselova, S.V.; Burkhanova, G.F.; Nuzhnaya, T.V.; Maksimov, I.V. Roles of ethylene and cytokinins in development of defense responses in Triticum aestivum plants infected with Septoria nodorum. Russ. J. Plant Physiol. 2016, 63, 609–619. [Google Scholar] [CrossRef]
- Veselova, S.V.; Burkhanova, G.F.; Nuzhnaya, T.V.; Rumyantsev, S.D.; Maksimov, I.V. Effect of the host-specific toxin SnTOX3 from Stagonospora nodorum on ethylene signaling pathway regulation and redox-state in common wheat. Vavilovskii Zhurnal Genet. Sel. Vavilov, J. Genet. Breed. 2019, 23, 856–864. [Google Scholar] [CrossRef]
- Vysotskaya, L.; Wilkinson, S.; Davies, W.J.; Arkhipova, T.; Kudoyarova, G. The effect of competition from neighbours on stomatal conductance in lettuce and tomato plants. Plant Cell Environ. 2011, 34, 729–737. [Google Scholar] [CrossRef]
- Warner, H.L.; Leopold, A.C. Ethylene evolution from 2-chloroethylphosphonic acid. Plant Physiol. 1969, 44, 156–158. [Google Scholar] [CrossRef] [PubMed]
- Babosha, A.V. Characteristics of dose-response dependence between zeatin and susceptibility of wheat seedlings to powdery mildew. Biol. Bull. 2012, 39, 534–541. [Google Scholar] [CrossRef]
- Ozeretskovskaya, O.L.; Vasyukova, N.I.; Chalenko, G.I.; Gerasimova, N.G.; Panina, Y.S.; Varlamov, V.P. Influence of systemic signal molecules on the rate of spread of the immunizing effect of elicitors over potato tissues. Appl. Biochem. Microbiol. 2004, 40, 213–216. [Google Scholar] [CrossRef]
- Troshina, N.B.; Yarullina, L.G.; Valeev, A.S.; Maksimov, I.V. Salicylic acid induces resistance to Septoria nodorum Berk. in Wheat. Izv. Akad. Nauk. Seriia Biol. Ross. Akad. Nauk. 2007, 34, 545–550. [Google Scholar] [CrossRef]
- Maximov, I.V.; Abizgildina, P.P.; Sorokan, A.V.; Burkhanova, G.F. Regulation of peroxidase activity under the influence of signaling molecules and Bacillus subtilis 26D in potato plants infected with Phytophthora infestans. Appl. Biochem. Microbiol. 2014, 50, 173–178. [Google Scholar] [CrossRef]
- Bindschedler, L.V.; Minibaeva, F.; Gardner, S.L.; Gerrish, C.; Davies, D.R.; Bolwell, G.P. Early signalling events in the apoplastic oxidative burst in suspension cultured French bean cells involve cAMP and Ca2+. New Phytol. 2001, 151, 185–194. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Zhangt, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, W.; Chan, Z.; Wu, Y. Endogenous cytokinin overproduction modulates ROS homeostasis and decreases salt stress resistance in Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 1004. [Google Scholar] [CrossRef]
- Plotnikova, L.Y.; Shtubey, T.Y. Influence of salicylic and succinic acids on the cytophysiological reactions of wheat infected by brown rust. Tsitologiia 2009, 51, 43–52. [Google Scholar]
- Veselov, S.Y.; Timergalina, L.N.; Akhiyarova, G.R.; Kudoyarova, G.R.; Korobova, A.V.; Ivanov, I.; Arkhipova, T.N.; Prinsen, E. Study of cytokinin transport from shoots to roots of wheat plants is informed by a novel method of differential localization of free cytokinin bases or their ribosylated forms by means of their specific fixation. Protoplasma 2018, 255, 1581–1594. [Google Scholar] [CrossRef]
- Arkhipova, T.N.; Veselov, S.U.; Melentiev, A.I.; Martynenko, E.V.; Kudoyarova, G.R. Ability of bacterium Bacillus subtilis to produce cytokinins and to influence the growth and endogenous hormone content of lettuce plants. Plant Soil 2005, 272, 201–209. [Google Scholar] [CrossRef]
- Kudoyarova, G.R.; Korobova, A.V.; Akhiyarova, G.R.; Arkhipova, T.N.; Zaytsev, D.Y.; Prinsen, E.; Egutkin, N.L.; Medvedev, S.S.; Veselov, S.Y. Accumulation of cytokinins in roots and their export to the shoots of durum wheat plants treated with the protonophore carbonyl cyanide m-chlorophenylhydrazone (CCCP). J. Exp. Bot. 2014. 65, 2287–2294. [CrossRef]
- Podgórska, A.; Burian, M.; Szal, B. Extra-cellular but extra-ordinarily important for cells: Apoplastic reactive oxygen species metabolism. Front. Plant Sci. 2017, 8, 1353. [Google Scholar] [CrossRef] [PubMed]
- Kadota, Y.; Shirasu, K.; Zipfe, C. Regulation of the NADPH oxidase RBOHD during plant immunity. Plant Cell Physiol. 2015, 56, 1472–1480. [Google Scholar] [CrossRef] [PubMed]
- Kunikowska, A.; Byczkowska, A.; Doniak, M.; Kaźmierczak, A. Cytokinins résumé: Their signaling and role in programmed cell death in plants. Plant Cell Rep. 2013, 32, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Romanov, G.A. How do cytokinins affect the cell? Russ. J. Plant Physiol. 2009, 56, 268–290. [Google Scholar] [CrossRef]
- Arnaud, D.; Lee, S.; Takebayashi, Y.; Choi, D.; Choi, J.; Sakakibara, H.; Hwanga, I. Cytokinin-mediated regulation of reactive oxygen species homeostasis modulates stomatal immunity in Arabidopsis. Plant Cell 2017, 29, 543–559. [Google Scholar] [CrossRef]
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef]
- Song, J.; Jiang, L.; Jameson, P.E. Co-ordinate regulation of cytokinin gene family members during flag leaf and reproductive development in wheat. BMC Plant Biol. 2012, 12, 78. [Google Scholar] [CrossRef]
- An, F.; Zhao, Q.; Ji, Y.; Li, W.; Jiang, Z.; Yu, X.; Zhang, C.; Han, Y.; He, W.; Liu, Y.; et al. Ethylene-Induced stabilization of ETHYLENE INSENSITIVE3 and EIN3-LIKE1 is mediated by proteasomal degradation of EIN3 Binding F-Box 1 and 2 That Requires EIN2 in Arabidopsis. Plant Cell 2010, 22, 2384–2401. [Google Scholar] [CrossRef]
- Chen, H.; Xue, L.; Chintamanani, S.; Germain, H.; Lin, H.; Cui, H.; Cai, R.; Zuo, J.; Tang, X.; Li, X.; et al. Ethylene INSENSITIVE3 and ETHYLENE INSENSITIVE3-LIKE1 repress SALICYLIC ACID INDUCTION DEFICIENT2 expression to negatively regulate plant innate immunity in Arabidopsis. Plant Cell 2009, 21, 2527–2540. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Sheen, J. Two-component circuitry in Arabidopsis cytokinin signal transduction. Nature 2001, 413, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Reddy, M.P.; Chikara, J. WRKY: Its structure, evolutionary relationship, DNA-Binding selectivity, role in stress tolerance and development of plants. Mol. Biol. Rep. 2010, 38, 3883–3896. [Google Scholar] [CrossRef] [PubMed]
- Faris, J.D.; Zhang, Z.; Lu, H.; Lu, S.; Reddy, L.; Cloutier, S.; Fellers, J.P.; Meinhardt, S.W.; Rasmussen, J.B.; Xu, S.S.; et al. A unique wheat disease resistance-like gene governs effector-triggered susceptibility to necrotrophic pathogens. Proc. Natl. Acad. Sci. USA 2010, 107, 13544–13549. [Google Scholar] [CrossRef] [PubMed]
- Pandelova, I.; Melania, B.F.; Manning, V.A.; Wilhelm, L.J.; Mockler, T.C.; Ciuffetti, L.M. Analysis of transcriptome changes induced by Ptr ToxA in wheat provides insights into the mechanisms of plant susceptibility. Mol. Plant 2009, 2, 1067–1083. [Google Scholar] [CrossRef]
- He, P.; Chintamanani, S.; Chen, Z.; Zhu, L.; Kunkel, B.N.; Alfano, J.R.; Tang, X.; Zhou, J.M. Activation of a COI1-dependent pathway in Arabidopsis by Pseudomonas syringae type III effectors and coronatine. Plant J. 2004, 37, 589–602. [Google Scholar] [CrossRef] [PubMed]
- Wi, S.J.; Ji, N.R.; Park, K.Y. Synergistic biosynthesis of biphasic ethylene and reactive oxygen species in response to hemibiotrophic Phytophthora parasitica in tobacco plants. Physiol. Plant 2012, 159, 251–265. [Google Scholar] [CrossRef]
- Yang, C.; Li, W.; Cao, J.; Meng, F.; Yu, Y.; Huang, J.; Jiang, L.; Liu, M.; Zhang, Z.; Chen, X.; et al. Activation of ethylene signaling pathways enhances disease resistance by regulating ROS and phytoalexin production in rice. Plant J. 2017, 89, 338–353. [Google Scholar] [CrossRef]
- Rodrigues, F.C.T.; Aucique-P’erez, C.E.; Fontes, B.A.; Ribeiro, D.M.; Rodrigues, F.A. Involvement of ethylene in wheat resistance to infection by Pyricularia oryzae. Physiol. Mol. Plant Pathol. 2020, 112, 101526. [Google Scholar] [CrossRef]
- Broekaert, W.F.; Delaure, S.L.; De Bolle, M.F.; Cammue, B.P. The role of ethylene in host-pathogen interactions. Annu. Rev. Phytopathol. 2006, 44, 393–416. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Geraats, B.P.; Linthorst, H.J. Ethylene as a modulator of disease resistance in plants. Trends Plant Sci. 2006, 11, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Bouchez, O.; Huard, C.; Lorrain, S.; Roby, D.; Balague, C. Ethylene is one of the key elements for cell death and defense response control in the Arabidopsis lesion mimic mutant vad1. Plant Physiol. 2007, 145, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Zermiani, M.; Zonin, E.; Nonis, A.; Begheldo, M.; Ceccato, L.; Vezzaro, A.; Baldan, B.; Trentin, A.; Masi, A.; Pegoraro, M.; et al. Ethylene negatively regulates transcript abundance of ROP-GAP rheostat-encoding genes and affects apoplastic reactive oxygen species homeostasis in epicarps of cold stored apple fruits. J. Exp. Bot. 2015, 8, 7255–7270. [Google Scholar] [CrossRef] [PubMed]
- Belozerskaya, T.A.; Gessler, N.N. Oxidative stress and differentiation in Neurospora crassa. Microbiology 2006, 75, 427–431. [Google Scholar] [CrossRef]
- Barna, B.; Fodor, J.; Harrach, B.D.; Pogány, M.; Király, Z. The Janus face of reactive oxygen species in resistance and susceptibility of plants to necrotrophic and biotrophic pathogens. Plant Physiol. Biochem. 2012, 59, 37–43. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; van Breusegem, F. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef]
- Yoshioka, H.; Sugie, K.; Park, H.J.; Maeda, H.; Tsuda, N.; Kawakita, K.; Doke, N. Induction of plant gp91 phox homolog by fungal cell wall, arachidonic acid, and salicylic acid in potato. Mol. Plant Microbe Interact. 2001, 14, 725–736. [Google Scholar] [CrossRef]
- Xie, Y.J.; Xu, S.; Han, B.; Wu, M.Z.; Yuan, X.X.; Han, Y.; Gu, Q.; Xu, D.K.; Yang, Q.; Shen, W.B. Evidence of Arabidopsis salt acclimation induced by upregulation of HY1 and the regulatory role of RbohD-derived reactive oxygen species synthesis. Plant J. 2011, 66, 280–292. [Google Scholar] [CrossRef]
- Mur, L.A.; Laarhoven, L.J.; Harren, F.J.; Hall, M.A.; Smith, A.R. Nitric oxide interacts with salicylate to regulate biphasic ethylene production during the hypersensitive response. Plant Physiol. 2008, 148, 1537–1546. [Google Scholar] [CrossRef]
- Ding, L.; Xu, H.; Yi, H.; Yang, L.; Kong, Z.; Zhang, L.; Xue, S.; Jia, H.; Ma, Z. Resistance to hemi-biotrophic F. graminearum infection is associated with coordinated and ordered expression of diverse defense signaling pathways. PLoS ONE 2011, 6, e19008. [Google Scholar] [CrossRef]
- Xia, X.-J.; Zhou, Y.-H.; Shi, K.; Zhou, J.; Foyer, C.H.; Yu, J.-Q. Interplay between reactive oxygen species and hormones in the control of plant development and stress tolerance. J. Exp. Bot. 2015, 66, 2839–2856. [Google Scholar] [CrossRef] [PubMed]
- Mlejnek, P.; Doležel, P.; Procházka, S. Intracellular phosphorylation of benzyladenosine is related to apoptosis induction in tobacco BY-2 cells. Plant Cell Environ. 2003, 26, 1723–1735. [Google Scholar] [CrossRef]
- Daudi, A.; Cheng, Z.; O’Brien, J.A.; Mammarella, N.; Khan, S.; Ausubel, F.M.; Bolwell, P. The apoplastic oxidative burst peroxidase in Arabidopsis is a major component of pattern-triggered immunity. Plant Cell 2012, 24, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Bolwell, P.P.; Page, A.; Pi’slewska, M.; Wojtaszek, P. Pathogenic infection and the oxidative defences in plant apoplast. Protoplasma 2001, 217, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Dmochowska-Boguta, M.; Nadolska-Orczyk, A.; Orczyk, W. Roles of peroxidases and NADPH oxidases in the oxidative response of wheat (Triticum aestivum) to brown rust (Puccinia triticina) infection. Plant Pathol. 2013, 62, 993–1002. [Google Scholar] [CrossRef]
- Torres, M.A.; Dangl, J.L.; Jones, J.D. Arabidopsis gp91phox homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response. Proc. Natl. Acad. Sci. USA 2002, 99, 517–522. [Google Scholar] [CrossRef]
- Chaouch, S.; Queval, G.; Noctor, G. AtRbohF is a crucial modulator of defence-associated metabolism and a key factor in the interplay between intracellular oxidative stress and pathogenesis responses in Arabidopsis. Plant J. 2012, 69, 613–627. [Google Scholar] [CrossRef]
- Torres, M.A.; Jones, J.D.G.; Dangl, J.L. Pathogen-induced, NADPHoxidase derived reactive oxygen intermediates suppress spread of celldeath in Arabidopsis thaliana. Nat. Genet. 2005, 37, 1130–1134. [Google Scholar] [CrossRef]
- Mersmann, S.; Bourdais, G.; Rietz, S.; Robatzek, S. Ethylene signaling regulates accumulation of the FLS2 receptor and is required for the oxidative burst contributing to plant immunity. Plant Physiol. 2010, 154, 391–400. [Google Scholar] [CrossRef]
- Almagro, L.; Gomez Ros, L.V.; Belchi-Navarro, S.; Bru, R.; Ros Barcello, A.; Pedreno, M.A. Class III peroxidases in plant defence reactions. J. Exp. Bot. 2009, 60, 377–390. [Google Scholar] [CrossRef]
- Golemiec, E.; Tokarz, K.; Wielanek, M.; Niewiadomska, E. A dissection of the effects of ethylene, H2O2 and high irradiance on antioxidants and several genes associated with stress and senescence in tobacco leaves. J. Plant Physiol. 2014, 171, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Yang, M.; Wang, J.; Jiang, C.-Z.; Wang, Q. Application of exogenous ethylene inhibits postharvest peel browning of ‘Huangguan’ pear. Front. Plant Sci. 2017, 7, e2029. [Google Scholar] [CrossRef] [PubMed]
- Argandona, V.H.; Chaman, M.; Cardemil, L.; Munoz, O.; Zúniga, G.E.; Corcuera, L.J. Ethylene production and peroxidase activity in aphid-infested barley. J. Chem. Ecol. 2001, 27, 53–68. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Yue, X.; Wang, R.; Zhang, Y. Ethylene mediates UV-B-induced stomatal closure via peroxidase-dependent hydrogen peroxide synthesis in Vicia faba (L.). J. Exp. Bot. 2011, 62, 2657–2666. [Google Scholar] [CrossRef]
- Maksimov, I.V.; Yarullina, L.G.; Burkhanova, G.F.; Zaikina, E.A. Relationship between the aggressiveness and catalase activity of Septoria nodorum Berk. in wheat. Biol. Bull. 2013, 40, 441–446. [Google Scholar] [CrossRef]
- Sewelam, N.; Kazan, K.; Thomas-Hall, S.R.; Kidd, B.N.; Manners, J.M.; Schenk, P.M. Ethylene response factor 6 is a regulator of reactive oxygen species signaling in Arabidopsis. PLoS ONE 2013, 8, e70289. [Google Scholar] [CrossRef]
- Metraux, J.P. Systemic acquired resistance and salicylic acid: Current state of knowledge. Eur. J. Plant Pathol. 2001, 107, 13–18. [Google Scholar] [CrossRef]
- Jiang, C.J.; Shimono, M.; Sugano, S.; Kojima, M.; Liu, X.; Inoue, H.; Sakakibara, H.; Takatsuji, H. Cytokinins act synergistically with salicylic acid to activate defense gene expression in rice. Mol. Plant Microbe Interact. 2013, 3, 287–296. [Google Scholar] [CrossRef]
- Naseem, M.; Kaltdorf, M.; Dandekar, T. The nexus between growth and defence signalling: Auxin and cytokinin modulate plant immune response pathways. J. Exp. Bot. 2015, 66, 4885–4896. [Google Scholar] [CrossRef]
- Bielach, A.; Hrtyan, M.; Tognetti, V.B. Plants under Stress: Involvement of auxin and cytokinin. Int. J. Mol. Sci. 2017, 18, 1427. [Google Scholar] [CrossRef]
- Wybouw, B.; De Rybe, B. Cytokinin—A developing story. Trends Plant Sci. 2019, 24, 2. [Google Scholar] [CrossRef]
- Walters, D.R.; McRoberts, N. Plants and biotrophs: A pivotal role for cytokinins? Trends Plant Sci. 2006, 11, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Igari, K.; Endo, S.; Hibara, K.I.; Aida, M.; Sakakibara, H.; Kawasaki, T.; Tasaka, M. Constitutive activation of a CC-NB-LRR protein alters Growth-defense nexus: Auxin and cytokinin crosstalk morphogenesis through the cytokinin pathway in Arabidopsis. Plant J. 2008, 55, 14–27. [Google Scholar] [CrossRef]
- Babosha, A.V. Regulation of resistance and susceptibility in wheatpowdery mildew pathosystem with exogenous cytokinins. J. Plant Physiol. 2009, 166, 1892–1903. [Google Scholar] [CrossRef] [PubMed]
- Grosskinsky, D.K.; Naseem, M.; Abdelmohsen, U.R.; Plickert, N.; Engelke, T.; Griebel, T.; Zeier, J.; Novak, O.; Strnad, M.; Pfeifhofer, H.; et al. Cytokinins mediate resistance against Pseudomonas syringae in tobacco through increased antimicrobial phytoalexin synthesis independent of salicylic acid signalling. Plant Physiol. 2011, 157, 815–830. [Google Scholar] [CrossRef] [PubMed]
- Marmath, K.K.; Giri, P.; Taj, G.; Pandey, D.; Kumar, A. Effect of zeatin on the infection process and expression of MAPK-4 during pathogenesis of Alternaria brassicae in non-host and host Brassica plants. Afr. J. Biotechnol. 2013, 12, 2164–2174. [Google Scholar] [CrossRef]
- Reusche, M.; Klásková, J.; Thole, K.; Truskina, J.; Novák, O.; Janz, D.; Strnad, M.; Spíchal, L.; Lipka, V.; Teichman, T. Stabilization of cytokinin levels enhances arabidopsis resistance against Verticillium longisporum. Mol. Plant Microbe Interact. 2013, 26, 850–860. [Google Scholar] [CrossRef]
- Spoustová, P.; Hýsková, V.; Müller, K.; Schnablová, R.; Ryšlavá, H.; Čeřovská, N.; Malbeck, J.; Cvikrová, M.; Synková, H. Tobacco susceptibility to Potato virus Y(NTN) infection is affected by grafting and endogenous cytokinin content. Plant Sci. 2015, 235, 25–36. [Google Scholar] [CrossRef]
- Chang, H.; Jones, M.L.; Banowetz, G.M.; Clark, D.G. Overproduction of cytokinins in petunia flwers transformed with PSAG12-IPT delays corolla senescence and decreases sensitivity to ethylene. Plant Physiol. 2003, 132, 2174–2183. [Google Scholar] [CrossRef]
- Zdarska, M.; Dobisová, T.; Gelová, Z.; Pernisová, M.; Dabravolski, S.; Hejátko, J. Illuminating light, cytokinin, and ethylene signalling crosstalk in plant development. J. Exp. Bot. 2015, 66, 4913–4931. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Korobova, A.V.; Vysotskaya, L.B.; Vasinskaya, A.N.; Kudoyarova, G.R.; Kuluev, B.R.; Veselov, S.Y. Dependence of root biomass accumulation on the content and metabolism of cytokinins in ethylene-insensitive plants. Russ. J. Plant Physiol. 2016, 63, 597–603. [Google Scholar] [CrossRef]
- Naseem, M.; Dandekar, T. The role of auxin–cytokinin antagonism in plant–pathogen interactions. PLoS Pathog. 2012, 8, e1003026. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Koeck, M.; Williams, S.J.; Ashton, A.R.; Lawrence, G.J.; Sakakibara, H.; Kojima, M.; Böttcher, C.; Ericsson, D.J.; Hardham, A.R.; et al. Structural and functional insights into the modulation of the activity of a flax cytokinin oxidase by flax rust effector AvrL567-A. Mol. Plant Pathol. 2019, 20, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Avalbaev, A.M.; Somov, K.A.; Yuldashev, R.A.; Shakirova, F.M. Cytokinin oxidase is key enzyme of cytokinin degradation. Biochemistry 2012, 77, 1354–1361. [Google Scholar] [CrossRef]
- Xu, J.; Chen, L.; Sun, H.; Wusiman, N.; Sun, W.; Li, B.; Gao, Y.; Kong, J.; Zhang, D.; Zhang, X.; et al. Crosstalk between cytokinin and ethylene signaling pathways regulates leaf abscission in cotton in response to chemical defoliants. J. Exp. Bot. 2019, 70, 1525–1538. [Google Scholar] [CrossRef]
- Hai, N.N.; Chuong, N.N.; Tu, N.H.C.; Kisiala, A.; Hoang, X.L.T.; Thao, N.P. Role and regulation of cytokinins in plant response to drought stress. Plants 2020, 9, 422. [Google Scholar] [CrossRef]
- Taverner, E.; Lethama, D.S.; Wanga, J.; Cornishb, E.; Willcocks, D.A. Influence of ethylene on cytokinin metabolism in relation to Petunia corolla senescence. Phytochemistry 1999, 51, 341–347. [Google Scholar] [CrossRef]
- Hass, C.; Lohrmann, J.; Albrecht, V.; Sweere, U.; Humme, F.; Yoo, S.D.; Hwang, I.; Zhu, T.; Schafer, E.; Kudla, J.; et al. The response regulator 2 mediates ethylene signalling and hormone signal integration in Arabidopsis. EMBO J. 2004, 23, 3290–3302. [Google Scholar] [CrossRef]
- Shi, Y.; Tian, S.; Hou, L.; Huang, X.; Zhang, X.; Guo, H.; Yang, S. Ethylene signaling negatively regulates freezing tolerance by repressing expression of CBF and Type-A ARR genes in Arabidopsis. Plant Cell 2012, 24, 2578–2595. [Google Scholar] [CrossRef]
- Zdarska, M.; Cuyacot, A.R.; Tarr, P.T.; Yamoune, A.; Szmitkowska, A.; Hrdinova, V.; Gelova, Z.; Meyerowitz, E.M.; Hejatko, J. ETR1 Integrates response to ethylene and cytokinins into a single multistep phosphorelay pathway to control root growth. Mol. Plant 2019, 12, 1338–1352. [Google Scholar] [CrossRef]
- Huo, R.; Liu, Z.; Yu, X.; Li, Z. The interaction network and signaling specificity of two-component system in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 4898. [Google Scholar] [CrossRef] [PubMed]
- Zubo, Y.O.G.; Schaller, E. Role of the cytokinin-activated Type-B response regulators in hormone crosstalk. Plants 2020, 9, 166. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Choi, D.; Lee, S.; Ryu, C.M.; Hwang, I. Cytokinins and plant immunity: Old foes or new friends? Trends Plant Sci. 2011, 7, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Naseem, M.; Wölflng, M.; Dandekar, T. Cytokinins for immunity beyond growth, galls and green islands. Trends Plant Sci. 2014, 19, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Rashotte, A.M.; Carson, S.D.B.; To, J.P.C.; Kieber, J.J. Expression profiling of cytokinin action in arabidopsis. Plant Physiol. 2003, 132, 1998–2011. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.J.; Park, J.-Y.; Ku, S.-J.; Ha, Y.-M.; Kim, S.; Kim, M.D.; Oh, M.-H.; Kim, J. Genome-wide expression profiling of ARABIDOPSIS RESPONSE REGULATOR 7 (ARR7) overexpression in cytokinin response. Mol. Genet. Genom. 2007, 277, 115–137. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Xiao, J.; Ding, X.; Xiong, M.; Cai, M.; Cao, Y.; Li, X.; Xu, C.; Wang, S. OsWRKY13 mediates rice disease resistance by regulating defense-related genes in salicylate- and jasmonate-dependent signaling. Mol. Plant Microbe Interact. 2007, 20, 492–499. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Treatments | The Most Biologically Active Forms and Inactive Forms of CKs | ||||||
|---|---|---|---|---|---|---|---|---|
| Z | ZR | iP | iPR | Z-9G | Z-OG | Amount of CK Active Forms (Z, ZR, iP, iPR) | ||
| Kaz10 | Water | 40.5 ± 3.2 | 11.7 ± 0.9 | 5.1 ± 0.4 | 24.9 ± 1.9 | 40.9 ± 3.2 | 11.4 ± 0.9 | 82.2 ± 6.5 |
| SnB | 30.3 ± 2.4 * | 11.2 ± 0.9 | 14.1 ± 1.1 * | 12.4 ± 0.9 * | 63.1 ± 5.0 * | 23.3 ± 1.8 * | 67.8 ± 5.4 * | |
| ET | 36.5 ± 2.9 | 15.7 ± 1.3 | 8.3 ± 0.7 | 18.5 ± 1.4 | 63.1 ± 5.0 * | 39.9 ± 3.1 ** | 78.8 ± 6.3 * | |
| ET + SnB | 32.3 ± 2.6 * | 16.1 ± 1.3 | 17.1 ± 1.4 * | 14.2 ± 1.1 * | 68.4 ± 5.4 * | 36.2 ± 2.9 ** | 79.7 ± 6.3 * | |
| 1-MCP | 38.9 ± 3.1 | 14.1 ± 1.1 | 7.1 ± 0.5 | 21.3 ± 1.7 | 39.7 ± 3.1 ** | 8.9 ± 0.7 | 81.5 ± 6.5 | |
| 1-MCP + SnB | 60.6 ± 4.8 ** | 32.9 ± 2.6 * | 33.6 ± 2.6 ** | 36.9 ± 2.9 ** | 34.1 ± 2.7 ** | 6.6 ± 0.5 *** | 164.0 ± 13.1 ** | |
| Om35 | Water | 28.9 ± 2.3 | 13.7 ± 1.1 | 8.2 ± 0.6 | 22.4 ± 1.7 | 54.7 ± 4.3 | 3.7 ± 0.2 | 73.3 ± 5.8 |
| SnB | 82.6 ± 6.1 * | 38.6 ± 3.0 * | 37.7 ± 3.0 * | 49.8 ± 3.9 * | 43.8 ± 3.5 * | 0.8 ± 0.6 * | 208.8 ± 16.7 * | |
| ET | 36.4 ± 2.9 ** | 13.1 ± 1.0 | 9.2 ± 0.7 | 23.0 ± 1.8 | 55.5 ± 4.4 | 13.5 ± 1.0 ** | 81.8 ± 6.5 ** | |
| ET + SnB | 30.4 ± 2.4 | 17.1 ± 1.3 | 33.6 ± 2.6 * | 34.8 ± 2.7 ** | 66.8 ± 5.3 ** | 45.1 ± 3.6 *** | 116.0 ± 9.2 *** | |
| 1-MCP | 33.1 ± 2.6 | 13.4 ± 1.0 | 10.9 ± 0.8 | 28.4 ± 2.2 | 39.2 ± 3.1 * | 9.9 ± 0.7 ** | 85.9 ± 6.8 ** | |
| 1-MCP + SnB | 85.8 ± 6.9 * | 40.4 ± 3.2 * | 31.9 ± 2.5 * | 56.8 ± 4.5 * | 26.2 ± 2.0 *** | 0.9 ± 0.1 * | 215.0 ± 17.2 * | |
| Variety | Treatments | The Most Biologically Active Forms and Inactive Forms of CKs | ||||||
|---|---|---|---|---|---|---|---|---|
| Z | ZR | iP | iPR | Z-9G | Z-OG | Amount of CK Active Forms (Z, ZR, iP, iPR) | ||
| Kaz10 | Water | 39.8 ± 3.1 | 11.6 ± 0.9 | 4.9 ± 0.4 | 22.5 ± 1.8 | 34.8 ± 2.7 | 8.8 ± 0.7 | 79.0 ± 6.3 |
| Sn4VD | 70.3 ± 5.6 * | 19.3 ± 1.5 * | 27.3 ± 2.1 * | 47.7 ± 3.8 * | 42.3 ± 3.3 * | 0.6 ± 0.1 * | 164.8 ± 13.1 * | |
| ET | 49.5 ± 3.9 | 15.2 ± 1.2 | 8.6 ± 0.6 ** | 16.7 ± 1. | 43.3 ± 3.4 * | 16.7 ± 1.3 ** | 90.2 ± 7.2 | |
| ET + Sn4VD | 63.5 ± 5.0 * | 18.1 ± 1.4 * | 25.2 ± 2.0 * | 34.6 ± 2.7 ** | 47.3 ± 3.7 * | 14.0 ± 1.1 ** | 141.5 ± 11.3 * | |
| 1-MCP | 72.7 ± 5.8 * | 11.7 ± 0.9 | 5.1 ± 0.4 | 26.0 ± 2.0 | 43.7 ± 3.5 * | 1.0 ± 0.1 * | 115.6 ± 9.2 ** | |
| 1-MCP + Sn4VD | 90.0 ± 7.2 ** | 17.6 ± 1.4 * | 32.3 ± 2.5 * | 52.2 ± 4.1 * | 29.1 ± 2.3 ** | 1.4 ± 0.1 * | 192.2 ± 15.3 *** | |
| Om35 | Water | 39.7 ± 3.1 | 11.9 ± 0.9 | 9.7 ± 0.7 | 20.9 ± 1.6 | 40.9 ± 3.2 | 7.6 ± 0.6 | 82.4 ± 6.6 |
| Sn4VD | 70.2 ± 5.6 * | 24.5 ± 1.9 * | 26.4 ± 2.1 * | 41.5 ± 3.3 * | 37.2 ± 2.9 | 1.0 ± 0.1* | 162.8 ± 13.0 * | |
| ET | 39.9 ± 3.2 | 14.8 ± 1.1 | 11.8 ± 0.9 | 20.6 ± 1.6 | 49.9 ± 4.0 | 17.7 ± 1.4 ** | 87.3 ± 6.9 | |
| ET + Sn4VD | 46.8 ± 3.7 ** | 23.5 ± 1.8 * | 30.6 ± 2.4 * | 42.8 ± 3.4 * | 41.5 ± 3.3 | 9.5 ± 0.7 | 143.7 ± 11.5 * | |
| 1-MCP | 35.0 ± 2.8 | 16.5 ± 1.3 | 8.0 ± 0.6 | 17.3 ± 1.3 | 40.7 ± 3.2 | 9.9 ± 0.8 | 76.9 ± 6.1 | |
| 1-MCP + Sn4VD | 88.3 ± 7.0 *** | 28.0 ± 3.7 * | 46.8 ± 3.7 ** | 43.7 ± 3.5 * | 37.7 ± 3.0 | 1.4 ± 0.1 * | 207.0 ± 16.5 ** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veselova, S.V.; Nuzhnaya, T.V.; Burkhanova, G.F.; Rumyantsev, S.D.; Khusnutdinova, E.K.; Maksimov, I.V. Ethylene-Cytokinin Interaction Determines Early Defense Response of Wheat against Stagonospora nodorum Berk. Biomolecules 2021, 11, 174. https://doi.org/10.3390/biom11020174
Veselova SV, Nuzhnaya TV, Burkhanova GF, Rumyantsev SD, Khusnutdinova EK, Maksimov IV. Ethylene-Cytokinin Interaction Determines Early Defense Response of Wheat against Stagonospora nodorum Berk. Biomolecules. 2021; 11(2):174. https://doi.org/10.3390/biom11020174
Chicago/Turabian StyleVeselova, Svetlana V., Tatyana V. Nuzhnaya, Guzel F. Burkhanova, Sergey D. Rumyantsev, Elza K. Khusnutdinova, and Igor V. Maksimov. 2021. "Ethylene-Cytokinin Interaction Determines Early Defense Response of Wheat against Stagonospora nodorum Berk." Biomolecules 11, no. 2: 174. https://doi.org/10.3390/biom11020174
APA StyleVeselova, S. V., Nuzhnaya, T. V., Burkhanova, G. F., Rumyantsev, S. D., Khusnutdinova, E. K., & Maksimov, I. V. (2021). Ethylene-Cytokinin Interaction Determines Early Defense Response of Wheat against Stagonospora nodorum Berk. Biomolecules, 11(2), 174. https://doi.org/10.3390/biom11020174