
Syntaxin 1A Gene Is Negatively Regulated in a Cell/Tissue Specific Manner by YY1 Transcription Factor, Which Binds to the −183 to −137 Promoter Region Together with Gene Silencing Factors Including Histone Deacetylase


Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Electrophoresis Mobility Shift Assays (EMSA) and Supershift Assay
2.3. Chromatin Immunoprecipitation (ChIP) Assay
2.4. Transient Cotransfection Assays and Luciferase Reporter Gene Assays
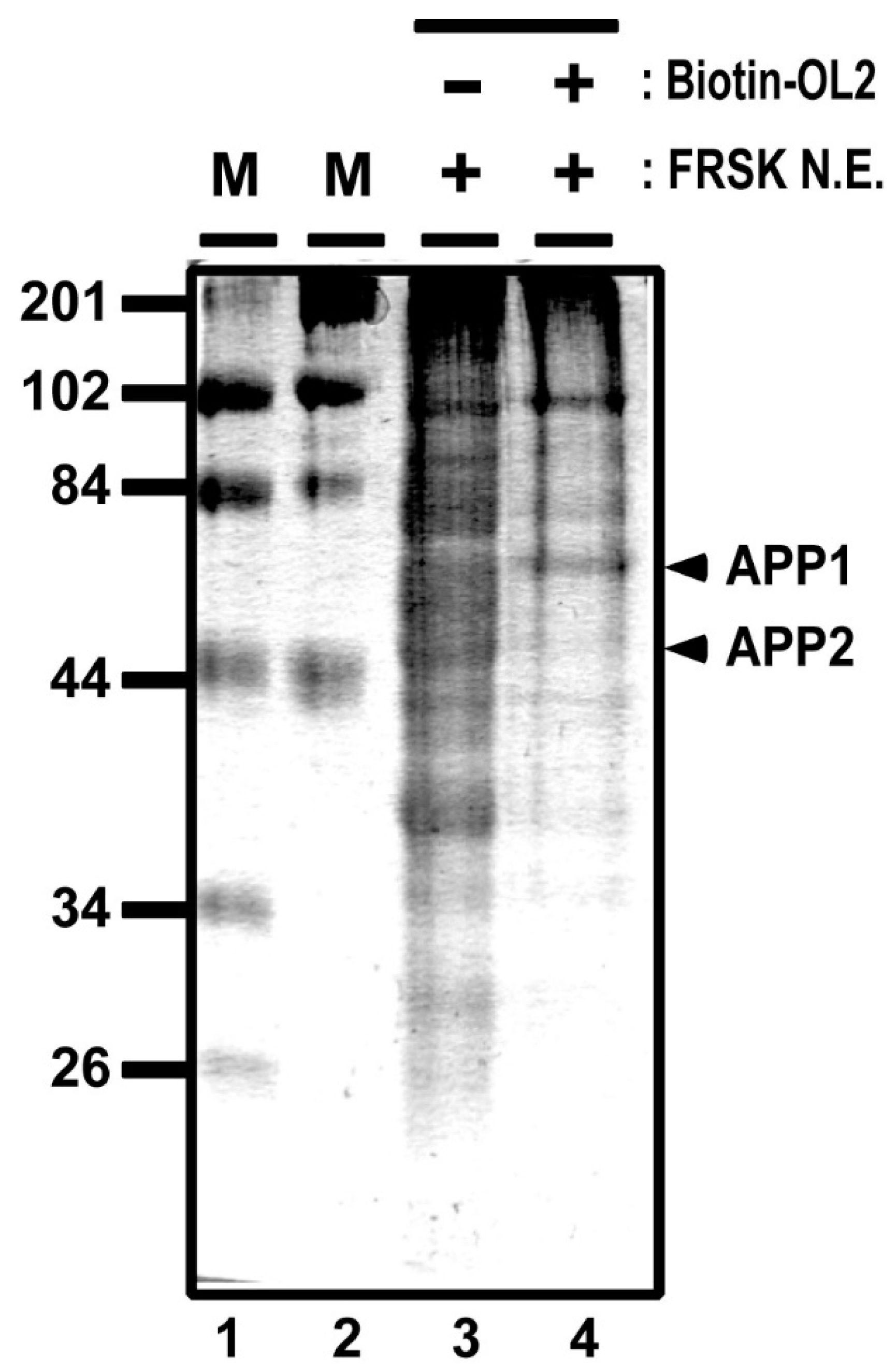
2.5. DNA Affinity Purification of Transcription Factor
2.6. Mass Spectrometry Analyses
2.7. Statistical Analysis
3. Results
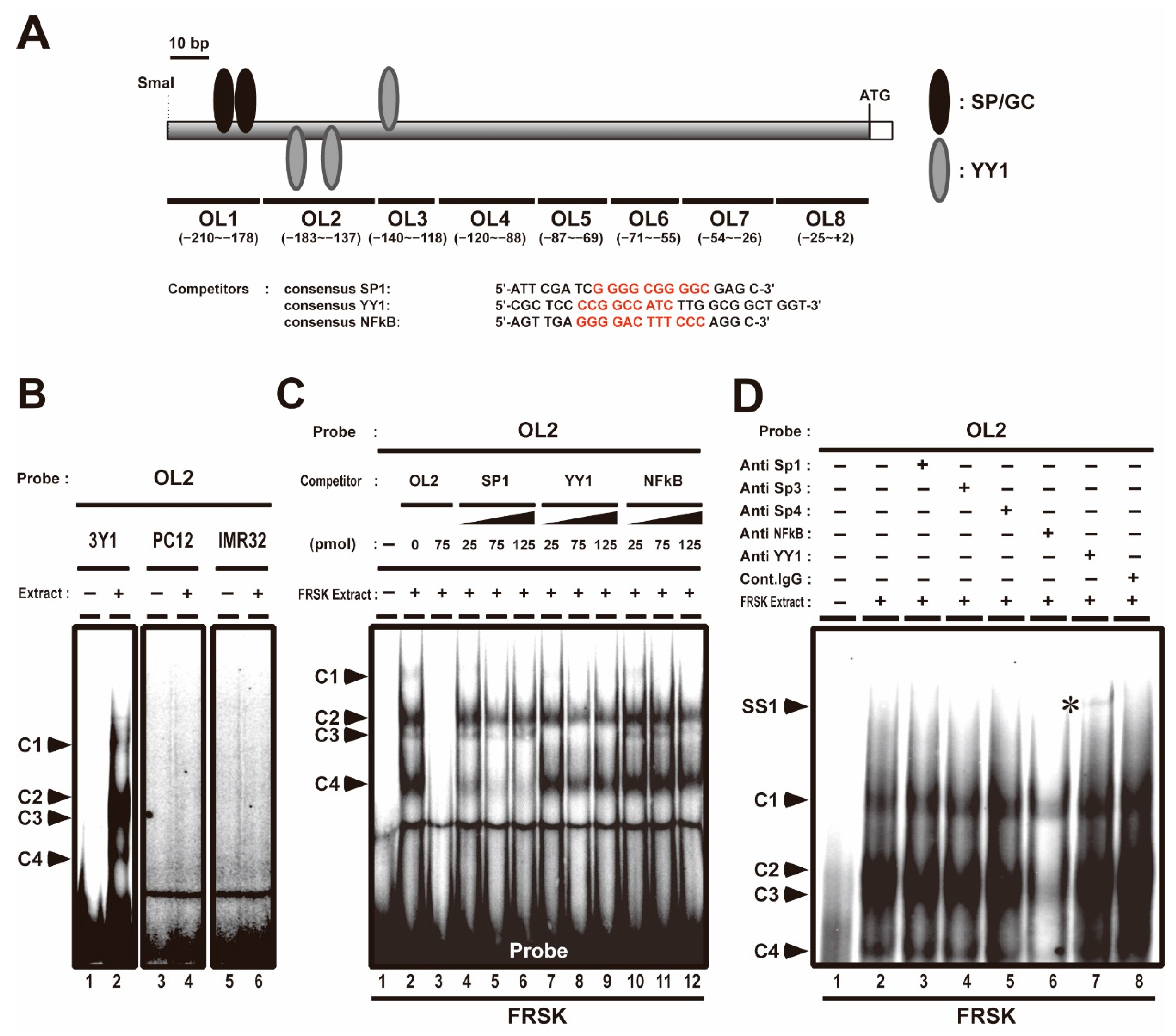
3.1. YY1 Transcription Factor Associates with the −183 to −137 OL2 Region of the Stx1a Core Promoter Region in Non-Neuronal Cells Which Do Not Express Stx1a
3.2. YY1 Associates with Stx1a–CPR in Cells and Tissues not Expressing Stx1a
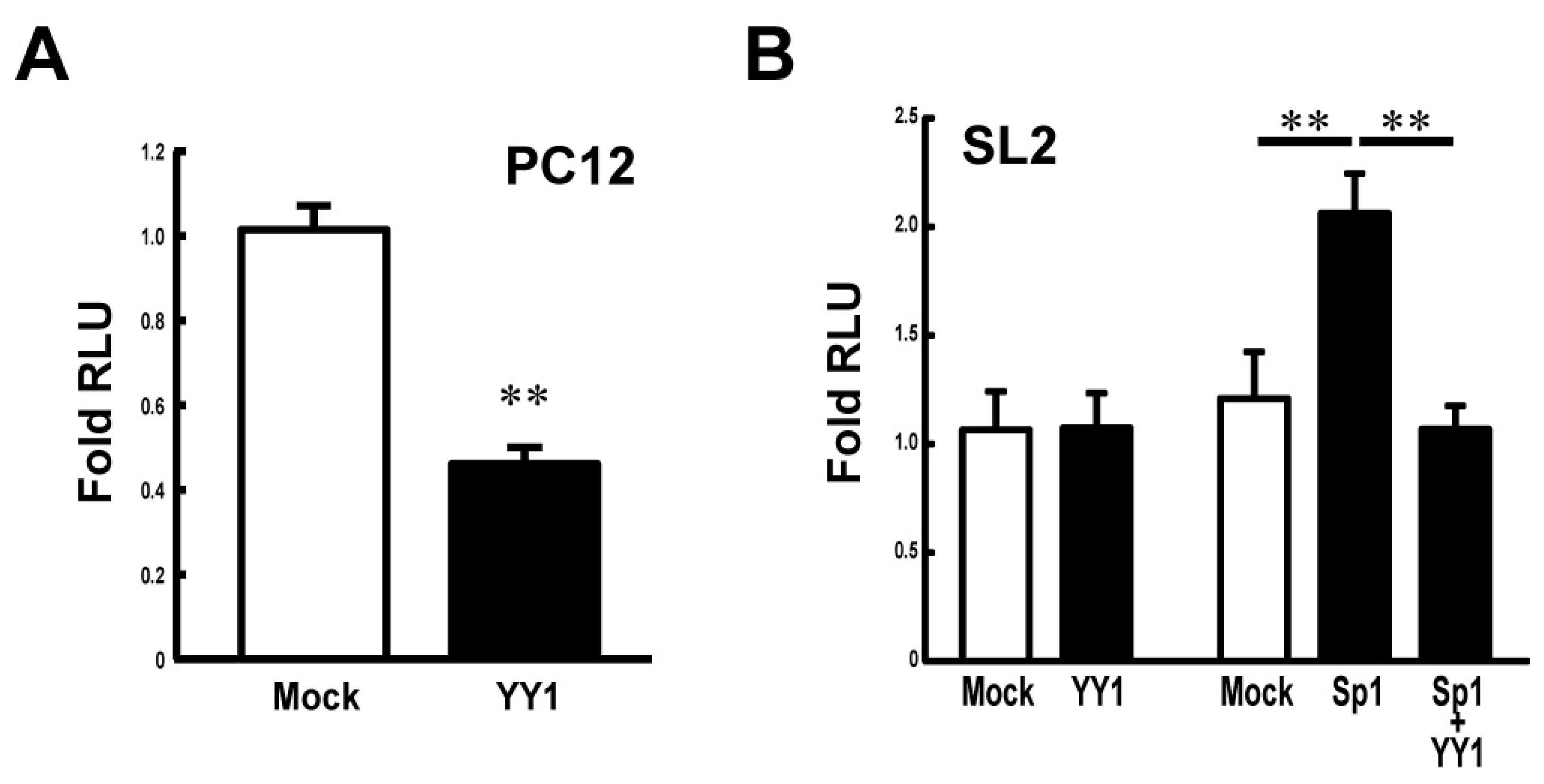
3.3. YY1 Negatively Regulates the Stx1a Transcription
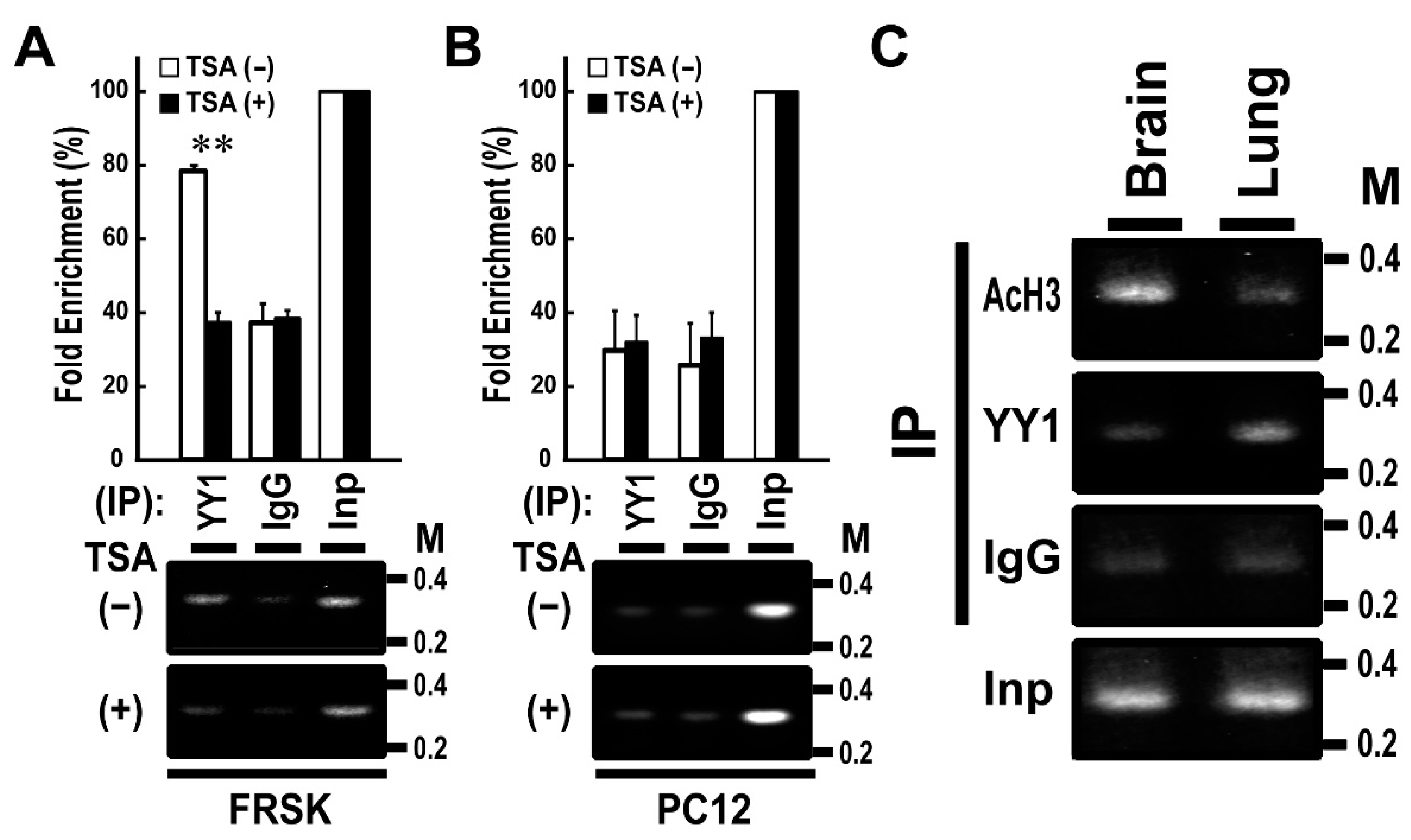
3.4. Gene Silencing Factors, Including Histone Deacetylase, Associates with the −183 to −137 OL2 Promoter Region
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inoue, A.; Obata, K.; Akagawa, K. Cloning and sequence analysis of cDNA for a neuronal cell membrane antigen, HPC-1. J. Biol. Chem. 1992, 267, 10613–10619. [Google Scholar] [CrossRef]
- Bennett, M.K.; Calakos, N.; Scheller, R.H. Syntaxin: A synaptic protein implicated in docking of synaptic vesicles at presynaptic active zones. Science 1992, 257, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Jahn, R.; Scheller, R.H. SNAREs—engines for membrane fusion. Nat. Rev. Mol. Cell Biol. 2006, 7, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Sudhof, T.C.; Rothman, J.E. Membrane fusion: Grappling with SNARE and SM proteins. Science 2009, 323, 474–477. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Fujiwara, T.; Miyazawa, A.; Asakawa, S.; Shimizu, N.; Shimizu, Y.; Mikoshiba, K.; Akagawa, K. Mapping of the human HPC-1/syntaxin 1A gene (STX1A) to chromosome 7 band q11.2. Genomics 1997, 42, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Matsuoka, R.; Kimura, M.; Hirota, H.; Mikoshiba, K.; Shimizu, Y.; Shimizu, N.; Akagawa, K. Hemizygous deletion of the HPC-1/syntaxin 1A gene (STX1A) in patients with Williams syndrome. Cytogenet. Cell Genet. 1998, 82, 49–51. [Google Scholar] [CrossRef] [PubMed]
- Bellugi, U.; Lichtenberger, L.; Mills, D.; Galaburda, A.; Korenberg, J.R. Bridging cognition, the brain and molecular genetics: Evidence from Williams syndrome. Trends Neurosci. 1999, 22, 197–207. [Google Scholar] [CrossRef]
- Nakayama, T.; Mikoshiba, K.; Akagawa, K. The cell- and tissue-specific transcription mechanism of the TATA-less syntaxin 1A gene. FASEB J. 2016, 30, 525–543. [Google Scholar] [CrossRef]
- Nakayama, T.; Akagawa, K. Transcription regulation mechanism of the syntaxin 1A gene via protein kinase A. Biochem. J. 2017, 474, 2465–2473. [Google Scholar] [CrossRef]
- Lee, J.S.; Galvin, K.M.; See, R.H.; Eckner, R.; Livingston, D.; Moran, E.; Shi, Y. Relief of YY1 transcriptional repression by adenovirus E1A is mediated by E1A-associated protein p300. Genes. Dev. 1995, 9, 1188–1198. [Google Scholar] [CrossRef]
- Yao, Y.L.; Yang, W.M.; Seto, E. Regulation of transcription factor YY1 by acetylation and deacetylation. Mol. Cell. Biol. 2001, 21, 5979–5991. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.M.; Yao, Y.L.; Sun, J.M.; Davie, J.R.; Seto, E. Isolation and characterization of cDNAs corresponding to an additional member of the human histone deacetylase gene family. J. Biol. Chem. 1997, 272, 28001–28007. [Google Scholar] [CrossRef] [PubMed]
- Kalenik, J.L.; Chen, D.; Bradley, M.E.; Chen, S.J.; Lee, T.C. Yeast two-hybrid cloning of a novel zinc finger protein that interacts with the multifunctional transcription factor YY1. Nucleic Acids Res. 1997, 25, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Leblond-Francillard, M.; Dreyfus, M.; Rougeon, F. Isolation of DNA-protein complexes based on streptavidin and biotin interaction. Eur. J. Biochem. 1987, 166, 351–355. [Google Scholar] [CrossRef]
- Nagore, L.I.; Nadeau, R.J.; Guo, Q.; Jadhav, Y.L.; Jarrett, H.W.; Haskins, W.E. Purification and characterization of transcription factors. Mass Spectrom. Rev. 2013, 32, 386–398. [Google Scholar] [CrossRef][Green Version]
- Rappsilber, J.; Mann, M.; Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2007, 2, 1896–1906. [Google Scholar] [CrossRef]
- Hagen, G.; Muller, S.; Beato, M.; Suske, G. Cloning by recognition site screening of two novel GT box binding proteins: A family of Sp1 related genes. Nucleic Acids Res. 1992, 20, 5519–5525. [Google Scholar] [CrossRef]
- Morchikh, M.; Cribier, A.; Raffel, R.; Amraoui, S.; Cau, J.; Severac, D.; Dubois, E.; Schwartz, O.; Bennasser, Y.; Benkirane, M. HEXIM1 and NEAT1 Long Non-coding RNA Form a Multi-subunit Complex that Regulates DNA-Mediated Innate Immune Response. Mol. Cell 2017, 67, 387–399.e385. [Google Scholar] [CrossRef]
- Huarte, M.; Guttman, M.; Feldser, D.; Garber, M.; Koziol, M.J.; Kenzelmann-Broz, D.; Khalil, A.M.; Zuk, O.; Amit, I.; Rabani, M.; et al. A large intergenic noncoding RNA induced by p53 mediates global gene repression in the p53 response. Cell 2010, 142, 409–419. [Google Scholar] [CrossRef]
- Chang, Y.L.; Peng, Y.H.; Pan, I.C.; Sun, D.S.; King, B.; Huang, D.H. Essential role of Drosophila Hdac1 in homeotic gene silencing. Proc. Natl. Acad. Sci. USA 2001, 98, 9730–9735. [Google Scholar] [CrossRef]
- Garfinkel, B.P.; Melamed-Book, N.; Anuka, E.; Bustin, M.; Orly, J. HP1BP3 is a novel histone H1 related protein with essential roles in viability and growth. Nucleic Acids Res. 2015, 43, 2074–2090. [Google Scholar] [CrossRef] [PubMed]
- Satijn, D.P.; Hamer, K.M.; den Blaauwen, J.; Otte, A.P. The polycomb group protein EED interacts with YY1, and both proteins induce neural tissue in Xenopus embryos. Mol. Cell. Biol. 2001, 21, 1360–1369. [Google Scholar] [CrossRef] [PubMed]
- Gabriele, M.; Vulto-van Silfhout, A.T.; Germain, P.L.; Vitriolo, A.; Kumar, R.; Douglas, E.; Haan, E.; Kosaki, K.; Takenouchi, T.; Rauch, A.; et al. YY1 Haploinsufficiency Causes an Intellectual Disability Syndrome Featuring Transcriptional and Chromatin Dysfunction. Am. J. Hum. Genet. 2017, 100, 907–925. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Description | Score | Unique Peptides | PSMs |
|---|---|---|---|---|
| Q5FVM4 | Non-POU domain-containing octamer-binding protein | 7062.10 | 20 | 245 |
| P61980 | Heterogeneous nuclear ribonucleoprotein K | 6418.99 | 18 | 252 |
| Q4QQW4 | Histone deacetylase 1 | 152.09 | 3 | 5 |
| Q62733 | Lamina-associated polypeptide 2, isoform beta | 144.17 | 4 | 5 |
| P09414 | Nuclear factor 1A | 76.54 | 2 | 2 |
| Q6P747 | Heterochromatin protein 1 binding protein 3 | 38.16 | 2 | 2 |
| Accession | Description | Score | Unique Peptides | PSMs |
|---|---|---|---|---|
| 27545350 | Transcriptional repressor protein YY1 | 57.23 | 2 | 2 |
| 149065165 | Nuclear respiratory factor 1 (predicted) | 51.96 | 2 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakayama, T.; Fukutomi, T.; Terao, Y.; Akagawa, K. Syntaxin 1A Gene Is Negatively Regulated in a Cell/Tissue Specific Manner by YY1 Transcription Factor, Which Binds to the −183 to −137 Promoter Region Together with Gene Silencing Factors Including Histone Deacetylase. Biomolecules 2021, 11, 146. https://doi.org/10.3390/biom11020146
Nakayama T, Fukutomi T, Terao Y, Akagawa K. Syntaxin 1A Gene Is Negatively Regulated in a Cell/Tissue Specific Manner by YY1 Transcription Factor, Which Binds to the −183 to −137 Promoter Region Together with Gene Silencing Factors Including Histone Deacetylase. Biomolecules. 2021; 11(2):146. https://doi.org/10.3390/biom11020146
Chicago/Turabian StyleNakayama, Takahiro, Toshiyuki Fukutomi, Yasuo Terao, and Kimio Akagawa. 2021. "Syntaxin 1A Gene Is Negatively Regulated in a Cell/Tissue Specific Manner by YY1 Transcription Factor, Which Binds to the −183 to −137 Promoter Region Together with Gene Silencing Factors Including Histone Deacetylase" Biomolecules 11, no. 2: 146. https://doi.org/10.3390/biom11020146
APA StyleNakayama, T., Fukutomi, T., Terao, Y., & Akagawa, K. (2021). Syntaxin 1A Gene Is Negatively Regulated in a Cell/Tissue Specific Manner by YY1 Transcription Factor, Which Binds to the −183 to −137 Promoter Region Together with Gene Silencing Factors Including Histone Deacetylase. Biomolecules, 11(2), 146. https://doi.org/10.3390/biom11020146