
Isolation and Characterization of Fungal Endophytes Isolated from Medicinal Plant Ephedra pachyclada as Plant Growth-Promoting

 , ,
, ,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Material Used
2.2. Plant Sampling and Study Area
2.3. Isolation of Fungal Endophytes
2.4. Identification of Fungal Endophytes
2.5. Investigation of the Plant Growth-Promoting Traits
2.5.1. Detection of Fungal Endophytes Extracellular Enzymatic Activity
2.5.2. Antimicrobial Activity of Fungal Endophytes
2.5.3. Phosphate Solubilization
2.5.4. Ammonia Production
2.5.5. Screening and Quantification of Indole-3-Acetic Acid (IAA)
2.6. Effect of the Selected Fungal Strains as Bio-Inoculant
2.6.1. Effect of Endophytic Fungal Inoculation on Root Growth
2.6.2. Greenhouse Experiment
Experimental Design
Culture Conditions
Plant Tissue Analyses
2.7. Statistical Analysis
3. Results and Discussion
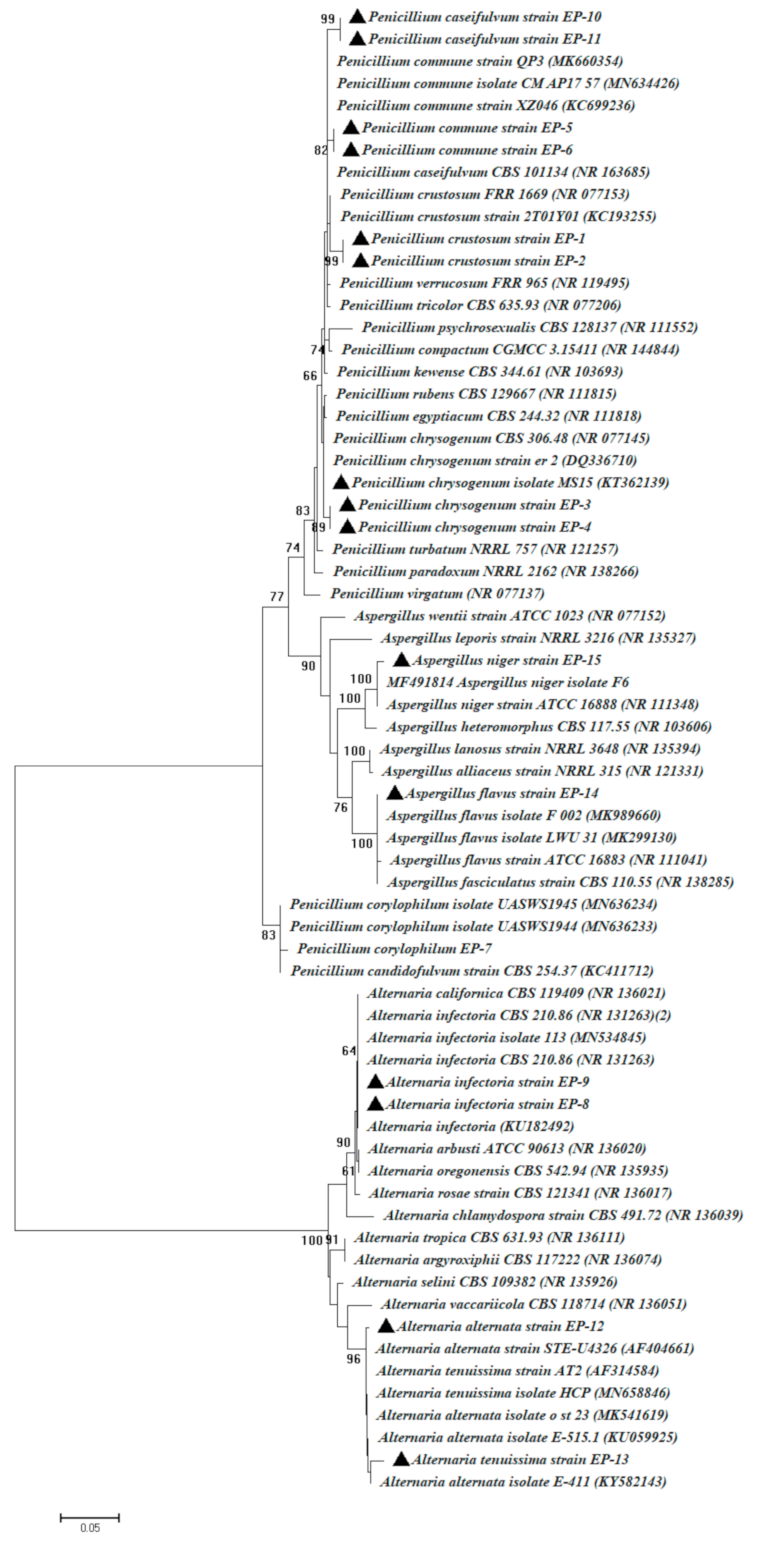
3.1. Isolation and Identification of Culturable Endophytic Fungi
3.2. Assessment of Endophytic Fungal Strains as Plant Growth-Promoting
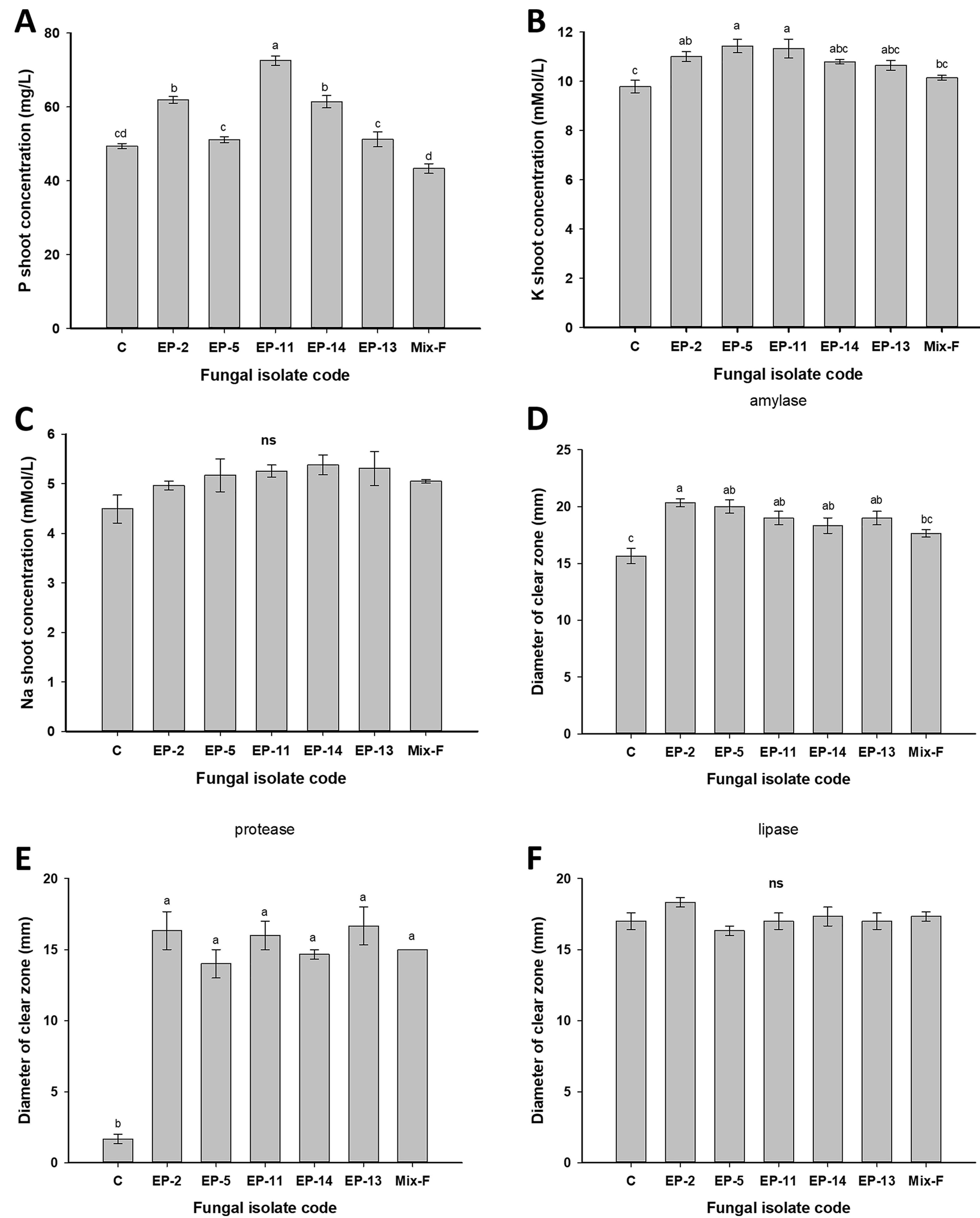
3.2.1. Extracellular Enzymatic Activities
3.2.2. Antimicrobial Activity
3.2.3. Phosphate Solubilization and Ammonia Production
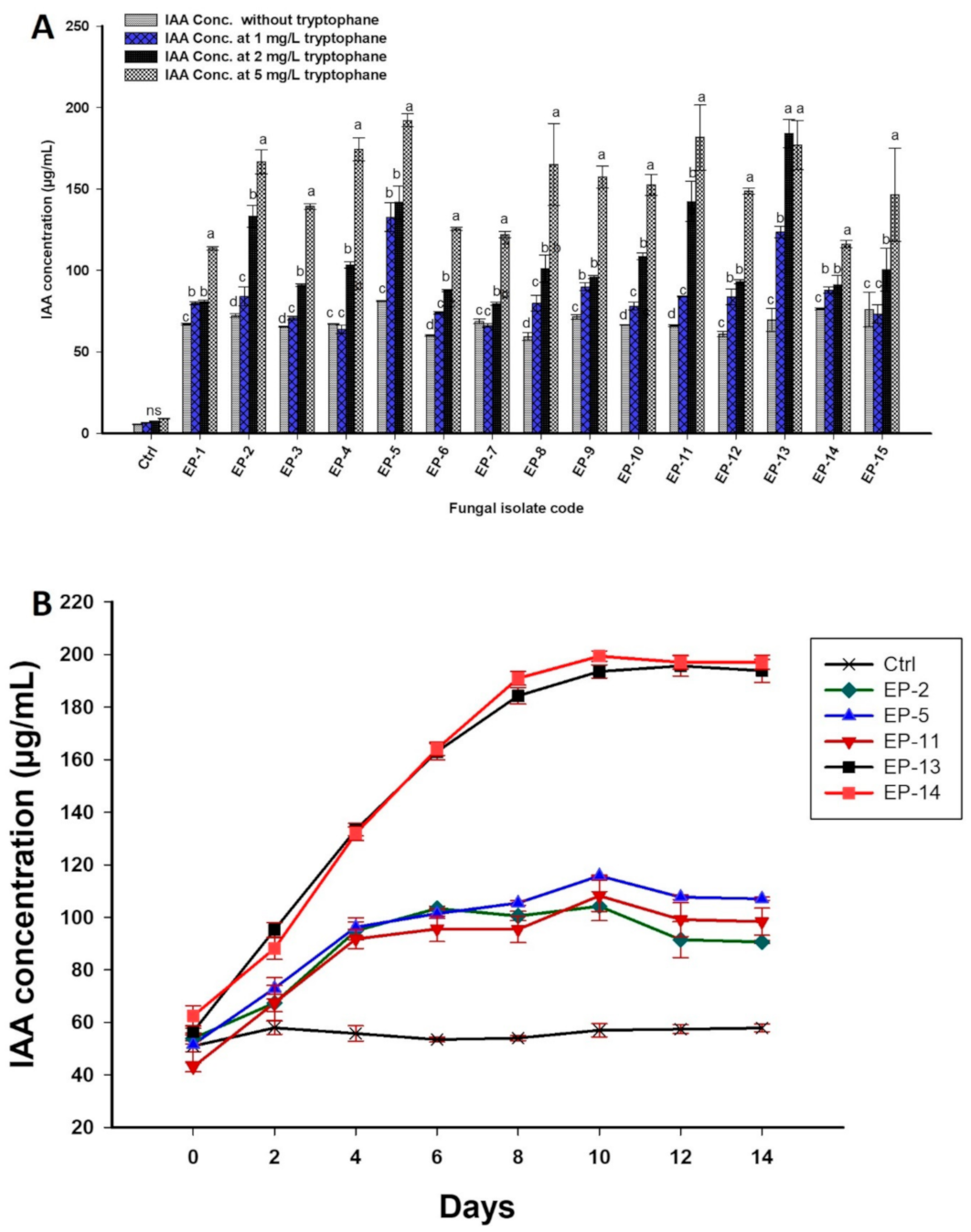
3.2.4. Qualitative and Quantitative IAA Productions
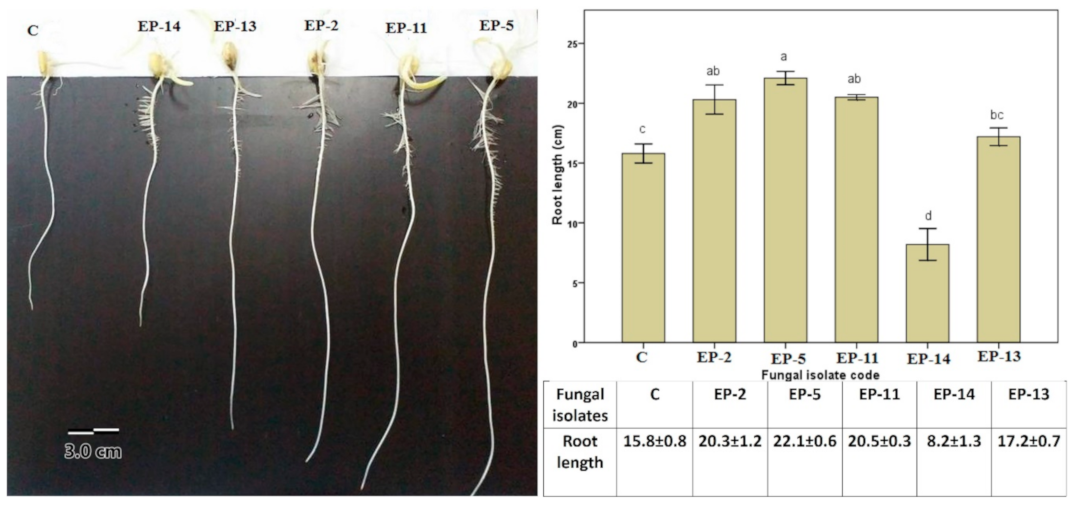
3.3. Effect of Endophytic Fungal Inoculation on Root Growth
3.4. Greenhouse Experiment
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Esawi, M.A.; Elkelish, A.; Soliman, M.; Elansary, H.O.; Zaid, A.; Wani, S.H. Serratia Marcescens BM1 Enhances Cadmium Stress Tolerance and Phytoremediation Potential of Soybean Through Modulation of Osmolytes, Leaf Gas Exchange, Antioxidant Machinery, and Stress-Responsive Genes Expression. Antioxidants 2020, 9, 43. [Google Scholar] [CrossRef]
- Elkelish, A.; Qari, S.H.; Mazrou, Y.S.A.; Abdelaal, K.A.A.; Hafez, Y.M.; Abu-Elsaoud, A.M.; Batiha, G.E.-S.; El-Esawi, M.A.; El Nahhas, N. Exogenous Ascorbic Acid Induced Chilling Tolerance in Tomato Plants Through Modulating Metabolism, Osmolytes, Antioxidants, and Transcriptional Regulation of Catalase and Heat Shock Proteins. Plants 2020, 9, 431. [Google Scholar] [CrossRef]
- Rafi, M.M.; Krishnaveni, M.; Charyulu, P. Phosphate-Solubilizing Microorganisms and Their Emerging Role in Sustainable Agriculture. In Recent Developments in Applied Microbiology and Biochemistry; Elsevier: Amsterdam, The Netherlands, 2019; pp. 223–233. [Google Scholar]
- El-Esawi, M.A.; Alayafi, A.A. Overexpression of Rice Rab7 Gene Improves Drought and Heat Tolerance and Increases Grain Yield in Rice (Oryza Sativa L.). Genes 2019, 10, 56. [Google Scholar] [CrossRef]
- Soliman, M.; Elkelish, A.; Souad, T.; Alhaithloul, H.; Farooq, M. Brassinosteroid Seed Priming with Nitrogen Supplementation Improves Salt Tolerance in Soybean. Physiol. Mol. Biol. Plants 2020, 26, 501–511. [Google Scholar] [CrossRef]
- Abedinzadeh, M.; Etesami, H.; Alikhani, H.A. Characterization of rhizosphere and endophytic bacteria from roots of maize (Zea mays L.) plant irrigated with wastewater with biotechnological potential in agriculture. Biotechnol. Rep. 2019, 21, e00305. [Google Scholar]
- Hassan, S.E.-D. Plant growth-promoting activities for bacterial and fungal endophytes isolated from medicinal plant of Teucrium polium L. J. Adv. Res. 2017, 8, 687–695. [Google Scholar]
- Eid, A.M.; Salim, S.S.; Hassan, S.E.-D.; Ismail, M.A.; Fouda, A. Role of Endophytes in Plant Health and Abiotic Stress Management. In Microbiome in Plant Health and Disease: Challenges and Opportunities; Kumar, V., Prasad, R., Kumar, M., Choudhary, D.K., Eds.; Springer: Singapore, 2019; pp. 119–144. [Google Scholar]
- Hashim, A.M.; Alharbi, B.M.; Abdulmajeed, A.M.; Elkelish, A.; Hozzein, W.N.; Hassan, H.M. Oxidative Stress Responses of Some Endemic Plants to High Altitudes by Intensifying Antioxidants and Secondary Metabolites Content. Plants 2020, 9, 869. [Google Scholar] [CrossRef]
- Zamin, M.; Fahad, S.; Khattak, A.M.; Adnan, M.; Wahid, F.; Raza, A.; Wang, D.; Saud, S.; Noor, M.; Bakhat, H.F.; et al. Developing the First Halophytic Turfgrasses for the Urban Landscape from Native Arabian Desert Grass. Environ. Sci. Pollut. Res. 2020, 27, 39702–39716. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Ahmad, M. Overexpression of AtWRKY30 Transcription Factor Enhances Heat and Drought Stress Tolerance in Wheat (Triticum Aestivum L.). Genes 2019, 10, 163. [Google Scholar] [CrossRef]
- Fouda, A.H.; Hassan, S.E.-D.; Eid, A.M.; Ewais, E.E.-D. Biotechnological applications of fungal endophytes associated with medicinal plant Asclepias sinaica (Bioss.). Ann. Agric. Sci. 2015, 60, 95–104. [Google Scholar]
- Murali, M.; Mahendra, C.; Hema, P.; Rajashekar, N.; Nataraju, A.; Sudarshana, M.; Amruthesh, K. Molecular profiling and bioactive potential of an endophytic fungus Aspergillus sulphureus isolated from Sida acuta: A medicinal plant. Pharm. Biol. 2017, 55, 1623–1630. [Google Scholar] [CrossRef] [PubMed]
- Danin, A. Desert Vegetation of Israel and Sinai; Cana Publ. House: Jerusalem, Israel, 1983. [Google Scholar]
- Strobel, G.A. Endophytes as sources of bioactive products. Microbes Infect. 2003, 5, 535–544. [Google Scholar] [CrossRef]
- Rydin, C.; Pedersen, K.R.; Crane, P.R.; Friis, E.M. Former diversity of Ephedra (Gnetales): Evidence from early Cretaceous seeds from Portugal and North America. Ann. Bot. 2006, 98, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Selim, K.; El-Beih, A.; AbdEl-Rahman, T.; El-Diwany, A. Biodiversity and antimicrobial activity of endophytes associated with Egyptian medicinal plants. Mycosphere 2011, 2, 669–678. [Google Scholar] [CrossRef]
- Selim, K.A.; Elkhateeb, W.A.; Tawila, A.M.; El-Beih, A.A.; Abdel-Rahman, T.M.; El-Diwany, A.I.; Ahmed, E.F. Antiviral and antioxidant potential of fungal endophytes of Egyptian medicinal plants. Fermentation 2018, 4, 49. [Google Scholar] [CrossRef]
- Peláez, F.; Collado, J.; Arenal, F.; Basilio, A.; Cabello, A.; Matas, M.D.; Garcia, J.; Del Val, A.G.; González, V.; Gorrochategui, J. Endophytic fungi from plants living on gypsum soils as a source of secondary metabolites with antimicrobial activity. Mycol. Res. 1998, 102, 755–761. [Google Scholar] [CrossRef]
- ALKahtani, M.D.; Fouda, A.; Attia, K.A.; Al-Otaibi, F.; Eid, A.M.; Ewais, E.E.-D.; Hijri, M.; St-Arnaud, M.; Hassan, S.E.-D.; Khan, N. Isolation and characterization of plant growth promoting endophytic bacteria from desert plants and their application as bioinoculants for sustainable agriculture. Agronomy 2020, 10, 1325. [Google Scholar] [CrossRef]
- Moubasher, A.; Moustafa, A. A survey of Egyptian soil fungi with special reference to Aspergillus, Pénicillium and Penicillium-related genera. Trans. Br. Mycol. Soc. 1970, 54, 35–44. [Google Scholar] [CrossRef]
- Woudenberg, J.; Groenewald, J.; Binder, M.; Crous, P. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef]
- Miller, D.; Bryant, J.; Madsen, E.; Ghiorse, W. Evaluation and optimization of DNA extraction and purification procedures for soil and sediment samples. Appl. Environ. Microbiol. 1999, 65, 4715–4724. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D., Shinsky, J., White, T., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Fouda, A.; Abdel-Maksoud, G.; Abdel-Rahman, M.A.; Salem, S.S.; Hassan, S.E.-D.; El-Sadany, M.A.-H. Eco-friendly approach utilizing green synthesized nanoparticles for paper conservation against microbes involved in biodeterioration of archaeological manuscript. Int. Biodeterior. Biodegrad. 2019, 142, 160–169. [Google Scholar] [CrossRef]
- Fouda, A.; Hassan, S.; Eid, A.M.; El-Din Ewais, E. The interaction between plants and bacterial endophytes under salinity stress. In Endophytes and Secondary Metabolites; Springer: Cham, Seitzerland, 2019; pp. 1–18. [Google Scholar]
- Kang, S.-M.; Bilal, S.; Shahzad, R.; Kim, Y.-N.; Park, C.-W.; Lee, K.-E.; Lee, J.-R.; Lee, I.-J. Effect of Ammonia and Indole-3-acetic Acid Producing Endophytic Klebsiella pneumoniae YNA12 as a Bio-Herbicide for Weed Inhibition: Special Reference with Evening Primroses. Plants 2020, 9, 761. [Google Scholar] [CrossRef] [PubMed]
- Fouda, A.; Hassan, S.E.-D.; Abdo, A.-M.; El-Gamal, M.-S. Antimicrobial, Antioxidant and Larvicidal activities of Spherical Silver Nanoparticles Synthesized by Endophytic Streptomyces spp. Biol. Trace Elem. Res. 2020, 195, 707–724. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Chauhan, N.S.; Chouhan, A.S.; Soni, V.; Omray, L. Antimicrobial screening of various extracts of Aphanmixis polystachya stems bark. Int. J. Adv. Pharm. Sci. 2010, 1, 147–150. [Google Scholar]
- Jasim, B.; Jimtha, C.J.; Jyothis, M.; Radhakrishnan, E. Plant growth promoting potential of endophytic bacteria isolated from Piper nigrum. Plant Growth Regul. 2013, 71, 1–11. [Google Scholar] [CrossRef]
- Marques, A.P.; Pires, C.; Moreira, H.; Rangel, A.O.; Castro, P.M. Assessment of the plant growth promotion abilities of six bacterial isolates using Zea mays as indicator plant. Soil Biol. Biochem. 2010, 42, 1229–1235. [Google Scholar] [CrossRef]
- Fouda, A.; Eid, A.M.; Elsaied, A.; El-Belely, E.F.; Barghoth, M.G.; Azab, E.; Gobouri, A.A.; Hassan, S.E. Plant Growth-Promoting Endophytic Bacterial Community Inhabiting the Leaves of Pulicaria incisa (Lam.) DC Inherent to Arid Regions. Plants 2021, 10, 76. [Google Scholar] [CrossRef]
- Ahmad, F.; Ahmad, I.; Khan, M.S. Indole acetic acid production by the indigenous isolates of Azotobacter and fluorescent Pseudomonas in the presence and absence of tryptophan. Turk. J. Biol. 2005, 29, 29–34. [Google Scholar]
- Tepe, B.; Daferera, D.; Sokmen, A.; Sokmen, M.; Polissiou, M. Antimicrobial and antioxidant activities of the essential oil and various extracts of Salvia tomentosa Miller (Lamiaceae). Food Chem. 2005, 90, 333–340. [Google Scholar] [CrossRef]
- Vermelho, A.B.; Meirelles, M.N.L.; Lopes, A.; Petinate, S.D.G.; Chaia, A.A.; Branquinha, M.H. Detection of extracellular proteases from microorganisms on agar plates. Mem. Do Inst. Oswaldo Cruz 1996, 91, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Wolf, B. A comprehensive system of leaf analyses and its use for diagnosing crop nutrient status. Commun. Soil Sci. Plant Anal. 1982, 13, 1035–1059. [Google Scholar] [CrossRef]
- Cottenie, A.; Verloo, M.; Kiekens, L.; Velghe, G.; Camerlynck, R. Chemical analysis of plant and soil laboratory of analytical and agrochemistry. State Univ. GentBelg. 1982, 42, 280–284. [Google Scholar]
- Hassan, S.E.-D.; Salem, S.S.; Fouda, A.; Awad, M.A.; El-Gamal, M.S.; Abdo, A.M. New approach for antimicrobial activity and bio-control of various pathogens by biosynthesized copper nanoparticles using endophytic actinomycetes. J. Radiat. Res. Appl. Sci. 2018, 11, 262–270. [Google Scholar] [CrossRef]
- Hassan, S.E.-D.; Fouda, A.; Radwan, A.A.; Salem, S.S.; Barghoth, M.G.; Awad, M.A.; Abdo, A.M.; El-Gamal, M.S. Endophytic actinomycetes Streptomyces spp mediated biosynthesis of copper oxide nanoparticles as a promising tool for biotechnological applications. Jbic J. Biol. Inorg. Chem. 2019, 24, 377–393. [Google Scholar] [CrossRef]
- Yang, L.; Li, W.J.; Long, J.; Yang, A.M.; Yang, Z.D.; Liu, X.F.; Hua, D.; Wang, W.J.; Ma, J.H. Isolation of Endophytic Fungi from Ephedra Intermedia and Research Antibacterial Activity of Secondary Metabolite Produced by the Fungi; Advanced Materials Research; Trans Tech Publ.: Freinbach, Switzerland, 2014; pp. 488–492. [Google Scholar]
- Kumar, A.; Kumar, A.; Devi, S.; Patil, S.; Payal, C.; Negi, S. Isolation, screening and characterization of bacteria from rhizospheric soils for different plant growth promotion (PGP) activities: An in vitro study. Recent Res. Sci. Technol. 2012, 4, 1–5. [Google Scholar]
- Il Kim, P.; Chung, K.-C. Production of an antifungal protein for control of Colletotrichum lagenarium by Bacillus amyloliquefaciens MET0908. Fems Microbiol. Lett. 2004, 234, 177–183. [Google Scholar] [CrossRef]
- Hallmann, J.; Quadt-Hallmann, A.; Mahaffee, W.; Kloepper, J. Bacterial endophytes in agricultural crops. Can. J. Microbiol. 1997, 43, 895–914. [Google Scholar] [CrossRef]
- Kavamura, V.N.; Santos, S.N.; Da Silva, J.L.; Parma, M.M.; Ávila, L.A.; Visconti, A.; Zucchi, T.D.; Taketani, R.G.; Andreote, F.D.; De Melo, I.S. Screening of Brazilian cacti rhizobacteria for plant growth promotion under drought. Microbiol. Res. 2013, 168, 183–191. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Giovanardi, D.; Stefani, E. Plant growth promoting and biocontrol activity of Streptomyces spp. as endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef]
- Hugo, W.B.; Russell, A.D. Pharmaceutical Microbiology; Blackwell Science: New Work, NY, USA, 1998. [Google Scholar]
- Wang, F.; Jiao, R.; Cheng, A.; Tan, S.; Song, Y. Antimicrobial potentials of endophytic fungi residing in Quercus variabilis and brefeldin A obtained from Cladosporium sp. World J. Microbiol. Biotechnol. 2007, 23, 79–83. [Google Scholar] [CrossRef]
- Manganyi, M.C.; Tchatchouang, C.-D.K.; Regnier, T.; Bezuidenhout, C.C.; Ateba, C.N. Bioactive Compound Produced by Endophytic Fungi Isolated From Pelargonium sidoides Against Selected Bacteria of Clinical Importance. Mycobiology 2019, 47, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Eze, P.M.; Nnanna, J.C.; Okezie, U.; Buzugbe, H.S.; Abba, C.C.; Chukwunwejim, C.R.; Okoye, F.B.; Esimone, C.O. Screening of metabolites from endophytic fungi of some Nigerian medicinal plants for antimicrobial activities. EuroBiotech J. 2019, 3, 10–18. [Google Scholar] [CrossRef]
- Li, X.; Geng, X.; Xie, R.; Fu, L.; Jiang, J.; Gao, L.; Sun, J. The endophytic bacteria isolated from elephant grass (Pennisetum purpureum Schumach) promote plant growth and enhance salt tolerance of hybrid Pennisetum. Biotechnol. Biofuels 2016, 9, 190. [Google Scholar] [CrossRef] [PubMed]
- Patle, P.; Navnage, N.; Ramteke, P. Endophytes in plant system: Roles in growth promotion, mechanism and their potentiality in achieving agriculture sustainability. Int. J. Chem. Stud. 2018, 6, 270–274. [Google Scholar]
- Liu, F.-P.; Liu, H.-Q.; Zhou, H.-L.; Dong, Z.-G.; Bai, X.-H.; Bai, P.; Qiao, J.-J. Isolation and characterization of phosphate-solubilizing bacteria from betel nut (Areca catechu) and their effects on plant growth and phosphorus mobilization in tropical soils. Biol. Fertil. Soils 2014, 50, 927–937. [Google Scholar] [CrossRef]
- Nain, L.; Yadav, R.; Saxena, J. Characterization of multifaceted Bacillus sp. RM-2 for its use as plant growth promoting bioinoculant for crops grown in semi arid deserts. Appl. Soil Ecol. 2012, 59, 124–135. [Google Scholar]
- Banik, A.; Dash, G.K.; Swain, P.; Kumar, U.; Mukhopadhyay, S.K.; Dangar, T.K. Application of rice (Oryza sativa L.) root endophytic diazotrophic Azotobacter sp. strain Avi2 (MCC 3432) can increase rice yield under green house and field condition. Microbiol. Res. 2019, 219, 56–65. [Google Scholar] [CrossRef]
- Ribeiro, C.M.; Cardoso, E.J.B.N. Isolation, selection and characterization of root-associated growth promoting bacteria in Brazil Pine (Araucaria angustifolia). Microbiol. Res. 2012, 167, 69–78. [Google Scholar] [CrossRef]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.-L.; Touraine, B.; Moënne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef]
- Lin, L.; Li, Z.; Hu, C.; Zhang, X.; Chang, S.; Yang, L.; Li, Y.; An, Q. Plant growth-promoting nitrogen-fixing enterobacteria are in association with sugarcane plants growing in Guangxi, China. Microbes Environ. 2009, 27, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Patten, C.L.; Glick, B.R. Role of Pseudomonas putida indole acetic acid in development of the host plant root system. Appl. Environ. Microbiol. 2002, 68, 3795–3801. [Google Scholar] [CrossRef] [PubMed]
- Manulis, S.; Haviv-Chesner, A.; Brandl, M.T.; Lindow, S.E.; Barash, I. Differential involvement of indole-3-acetic acid biosynthetic pathways in pathogenicity and epiphytic fitness of Erwinia herbicola pv. gypsophilae. Mol. Plant-Microbe Interact. 1998, 11, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Del Carmen Orozco-Mosqueda, M.; Del Carmen Rocha-Granados, M.; Glick, B.R.; Santoyo, G. Microbiome engineering to improve biocontrol and plant growth-promoting mechanisms. Microbiol. Res. 2018, 208, 25–31. [Google Scholar] [CrossRef]
- Hijri, M. Analysis of a large dataset of mycorrhiza inoculation field trials on potato shows highly significant increases in yield. Mycorrhiza 2016, 26, 209–214. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Strain Code | GenBank Accession Number | Homolog Sequences | Sequence Identity % | Closest Accession Number |
|---|---|---|---|---|
| EP-1 | MN954764 | Penicillium crustosum | 98.9% | NR077153 |
| 98.5% | NR163685 | |||
| EP-2 | MN954765 | Penicillium crustosum | 98.7% | NR077163 |
| EP-3 | MN954766 | Penicillium chrysogenum | 99.3% | NR077145 |
| EP-4 | MN954767 | Penicillium chrysogenum | 99.3% | NR077145 |
| EP-5 | MN954768 | Penicillium commune | 99.6% | MN634426 |
| 99.6% | MN634422 | |||
| EP-6 | MN954769 | Penicillium commune | 99.6% | MN634426 |
| 99.6% | MN634422 | |||
| EP-7 | MN954770 | Penicillium corylophilum | 99.4% | MN636234 |
| 99.4% | MN636233 | |||
| EP-8 | MN954771 | Alternaria infectoria | 99.8% | KU182492 |
| EP-9 | MN954772 | Alternaria infectoria | 99.8% | MN534845 |
| 99.8% | MH205934 | |||
| 99.8% | MK801346 | |||
| EP-10 | MN954773 | Penicillium caseifulvum | 99.04% | NR163685 |
| EP-11 | MN954774 | Penicillium caseifulvum | 99.04% | NR163685 |
| EP-12 | MN954775 | Alternaria alternata | 99.6% | KY582143 |
| 99.6% | KX622121 | |||
| 99.6% | KU059951 | |||
| EP-13 | MN954776 | Alternaria tenuissima | 99.4% | AF314584 |
| EP-14 | MN954777 | Aspergillus flavus | 99.6% | NR111041 |
| EP-15 | MN954778 | Aspergillus niger | 99.4% | LC195003 |
| Fungal Isolate Code | The Diameter of Clear Zones (mm) | |||||
|---|---|---|---|---|---|---|
| Amylase | CMCase | Gelatinase | Pectinase | Xylanase | Catalase | |
| Control | 0 | 0 | 0 | 0 | 0 | 0 |
| EP-1 | 20.7 ± 0.7 | 36.3 ± 0.7 | 30.0 ± 2.9 | 37.0 ± 1.0 | 23.7 ± 0.7 | + |
| EP-2 | 32.7 ± 1.5 | 37.0 ± 1.0 | 0 | 35.3 ± 0.3 | 26.3 ± 0.7 | + |
| EP-3 | 8.7 ± 0.7 | 40.7 ± 0.7 | 0 | 34.0 ± 3.1 | 0 | + |
| EP-4 | 0 | 37.0 ± 1.0 | 26.3 ± 0.9 | 35.3 ± 1.5 | 28.3 ± 0.7 | + |
| EP-5 | 0 | 28.7 ± 0.7 | 37.0 ± 1.0 | 27.3 ± 0.7 | 25.7 ± 1.2 | + |
| EP-6 | 34.3 ± 2.3 | 39.3 ± 1.3 | 0 | 32.7 ± 1.5 | 40.7 ± 0.7 | + |
| EP-7 | 38.7 ± 0.7 | 39.3 ± 0.3 | 22.0 ± 1.2 | 38.3 ± 0.9 | 0 | + |
| EP-8 | 31.7 ± 1.7 | 38.7 ± 0.7 | 36.7 ± 0.7 | 37.7 ± 1.5 | 41.7 ± 0.9 | + |
| EP-9 | 23.3 ± 1.7 | 39.3 ± 0.7 | 28.3 ± 1.7 | 40.0 ± 1.2 | 32.7 ± 0.7 | + |
| EP-10 | 29.3 ± 2.3 | 41.3 ± 0.7 | 39.03 ± 0.7 | 40.7 ± 0.7 | 32.7 ± 0.7 | + |
| EP-11 | 32.3 ± 1.5 | 41.7 ± 0.9 | 33.7 ± 1.9 | 41.0 ± 2.1 | 39.3 ± 0.7 | + |
| EP-12 | 39.3 ± 0.7 | 37.3 ± 1.8 | 39.3 ± 0.7 | 41.0 ± 1.0 | 42.7 ± 1.3 | + |
| EP-13 | 32.7 ± 1.5 | 36.0 ± 1.0 | 38.0 ± 1.2 | 41.7 ± 1.7 | 43.7 ± 0.7 | + |
| EP-14 | 27.7 ± 1.5 | 38.7 ± 0.7 | 33.7 ± 1.9 | 35.3 ± 0.3 | 25.7 ± 1.3 | + |
| EP-15 | 37.7 ± 1.5 | 39.3 ± 1.8 | 33.3 ± 1.7 | 39.3 ± 0.7 | 43.7 ± 0.9 | + |
| Fungal Isolate Code | Diameter of Clear Zones (mm) | |||||
|---|---|---|---|---|---|---|
| P. aeruginosa | S. typhimurium | E. coli | S. aureus | B. subtilis | C. albicans | |
| Control | 0 | 0 | 0 | 0 | 0 | 0 |
| EP-1 | 16.0 ± 0.8 | 14.2 ± 0.7 | 14.3 ± 0.9 | 13.7 ± 0.7 | 15.0 ± 0.6 | 0 |
| EP-2 | 0 | 0 | 0 | 20.3 ± 0.8 | 15.0 ± 0.7 | 0 |
| EP-3 | 15.0 ± 0.6 | 0 | 12.3 ± 0.7 | 14.7 ± 1.03 | 13.0 ± 0.9 | 0 |
| EP-4 | 18.0 ± 0.7 | 0 | 12.3 ± 0.8 | 0 | 14.3 ± 0.6 | 0 |
| EP-5 | 13.0 ± 0.9 | 15.3 ± 0.9 | 0 | 0 | 30.0 ± 1.7 | 0 |
| EP-6 | 16.0 ± 0.4 | 12 ± 0.6 | 20.3 ± 0.6 | 16.3 ± 0.7 | 16.7 ± 0.9 | 25.3 ± 1.3 |
| EP-7 | 13.0 ± 1.3 | 14.3 ± 0.5 | 15.3 ± 0.8 | 20.3 ± 0.9 | 14.7 ± 0.9 | 21.7 ± 1.7 |
| EP-8 | 14.0 ± 0.9 | 0 | 15 ± 0.9 | 0 | 0 | 0 |
| EP-9 | 18.0 ± 0.7 | 11.7 ± 0.7 | 15.3 ± 0.7 | 0 | 0 | 23.3 ± 3.3 |
| EP-10 | 15.0 ± 0.8 | 13.7 ± 0.6 | 14.3 ± 0.6 | 0 | 0 | 0 |
| EP-11 | 20.3 ± 0.6 | 14.0 ± 0.9 | 0 | 0 | 0 | 21.3 ± 0.7 |
| EP-12 | 15.0 ± 0.9 | 15.6 ± 1.8 | 0 | 15.7 ± 0.7 | 12.0 ± 0.5 | 0 |
| EP-13 | 14.0 ± 0.3 | 12 ± 0.6 | 13.3 ± 0.8 | 16 ± 0.8 | 0 | 0 |
| EP-14 | 22.6 ± 0.8 | 25.4 ± 1.4 | 20 ± 0.8 | 0 | 20.3 ± 0.9 | 0 |
| EP-15 | 16.0 ± 1.1 | 16.3 ± 0.7 | 0 | 0 | 0 | 0 |
| Fungal Isolate Code | Phosphate Solubilizing Activity (Diameter of A Clear Zone (mm) | Ammonia Production |
|---|---|---|
| Control | 0 a | 0 |
| EP-1 | 0 a | +++ |
| EP-2 | 11.3 ± 0.7 b | +++ |
| EP-3 | 10.3 ± 0.3 bc | +++ |
| EP-4 | 0 a | +++ |
| EP-5 | 0 a | +++ |
| EP-6 | 0 a | +++ |
| EP-7 | 0 a | ++ |
| EP-8 | 0 a | +++ |
| EP-9 | 0 a | +++ |
| EP-10 | 0 a | ++ |
| EP-11 | 0 a | ++ |
| EP-12 | 0 a | +++ |
| EP-13 | 0 a | +++ |
| EP-14 | 13.0 ± 0.6 d | +++ |
| EP-15 | 0 a | ++ |
| Fungal Isolate Code | Fresh Weight (mg) | Dry Weight (mg) | ||
|---|---|---|---|---|
| Shoot | Root | Shoot | Root | |
| C | 597 ± 21.9 b | 986.6 ± 37.5 ab | 84.4 ± 5.02 ab | 309.3 ± 16.9 a |
| EP-2 | 793 ± 68.4 a | 1112.2 ± 52.7 ab | 88.7 ± 5.3 ab | 333 ± 22.1 a |
| EP-5 | 637.2 ± 34.1 ab | 1104.6 ± 82.4 ab | 97.9 ± 6.9 ab | 278.6 ± 19.3 a |
| EP-11 | 694.6 ± 37.2 ab | 794.2 ± 56.8 b | 100.04 ± 7.2 b | 328.2 ± 14.7 a |
| EP-14 | 659.2 ± 27 ab | 1103.8 ± 124.7 ab | 88.9 ± 4.1 ab | 298.9 ± 22.3 a |
| EP-13 | 614.4 ± 37.1 b | 1038.2 ± 67.9 ab | 88.2 ± 2.02 ab | 317.2 ± 20.2 a |
| Mix_F | 676.6 ± 25.6 ab | 1177.8 ± 87.6 a | 99.9 ± 11.1 a | 293.9 ± 25.1 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalil, A.M.A.; Hassan, S.E.-D.; Alsharif, S.M.; Eid, A.M.; Ewais, E.E.-D.; Azab, E.; Gobouri, A.A.; Elkelish, A.; Fouda, A. Isolation and Characterization of Fungal Endophytes Isolated from Medicinal Plant Ephedra pachyclada as Plant Growth-Promoting. Biomolecules 2021, 11, 140. https://doi.org/10.3390/biom11020140
Khalil AMA, Hassan SE-D, Alsharif SM, Eid AM, Ewais EE-D, Azab E, Gobouri AA, Elkelish A, Fouda A. Isolation and Characterization of Fungal Endophytes Isolated from Medicinal Plant Ephedra pachyclada as Plant Growth-Promoting. Biomolecules. 2021; 11(2):140. https://doi.org/10.3390/biom11020140
Chicago/Turabian StyleKhalil, Ahmed Mohamed Aly, Saad El-Din Hassan, Sultan M. Alsharif, Ahmed M. Eid, Emad El-Din Ewais, Ehab Azab, Adil A. Gobouri, Amr Elkelish, and Amr Fouda. 2021. "Isolation and Characterization of Fungal Endophytes Isolated from Medicinal Plant Ephedra pachyclada as Plant Growth-Promoting" Biomolecules 11, no. 2: 140. https://doi.org/10.3390/biom11020140
APA StyleKhalil, A. M. A., Hassan, S. E.-D., Alsharif, S. M., Eid, A. M., Ewais, E. E.-D., Azab, E., Gobouri, A. A., Elkelish, A., & Fouda, A. (2021). Isolation and Characterization of Fungal Endophytes Isolated from Medicinal Plant Ephedra pachyclada as Plant Growth-Promoting. Biomolecules, 11(2), 140. https://doi.org/10.3390/biom11020140