
Activation of the α1β2γ2L GABAA Receptor by Physiological Agonists


Abstract
:1. Introduction
2. Results and Discussion
3. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Everington, E.A.; Gibbard, A.G.; Swinny, J.D.; Seifi, M. Molecular Characterization of GABA-A Receptor Subunit Diversity within Major Peripheral Organs and Their Plasticity in Response to Early Life Psychosocial Stress. Front. Mol. Neurosci. 2018, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.R.; Barnard, E.A. The benzodiazepine/GABA receptor complex: Molecular size in brain synaptic membranes and in solution. J. Neurochem. 1982, 39, 1507–1518. [Google Scholar] [CrossRef] [PubMed]
- Gladkevich, A.; Korf, J.; Hakobyan, V.P.; Melkonyan, K.V. The peripheral GABAergic system as a target in endocrine disorders. Auton. Neurosci. 2006, 124, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Barragan, A.; Weidner, J.M.; Jin, Z.; Korpi, E.R.; Birnir, B. GABAergic signalling in the immune system. Acta Physiol. 2015, 213, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Harrison, N.L.; Simmonds, M.A. Modulation of the GABA receptor complex by a steroid anaesthetic. Brain Res. 1984, 323, 287–292. [Google Scholar] [CrossRef]
- Kletke, O.; Gisselmann, G.; May, A.; Hatt, H.; Sergeeva, O.A. Partial agonism of taurine at gamma-containing native and recombinant GABAA receptors. PLoS ONE 2013, 8, e61733. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.S.; Gibbs, T.T.; Farb, D.H. Dual activation of GABAA and glycine receptors by beta-alanine: Inverse modulation by progesterone and 5 alpha-pregnan-3 alpha-ol-20-one. Eur. J. Pharmacol. 1993, 246, 239–246. [Google Scholar] [CrossRef]
- Scimemi, A. Plasticity of GABA transporters: An unconventional route to shape inhibitory synaptic transmission. Front. Cell. Neurosci. 2014, 8, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clements, J.D. Transmitter timecourse in the synaptic cleft: Its role in central synaptic function. Trends Neurosci. 1996, 19, 163–171. [Google Scholar] [CrossRef]
- Nusser, Z.; Mody, I. Selective modulation of tonic and phasic inhibitions in dentate gyrus granule cells. J. Neurophysiol. 2002, 87, 2624–2628. [Google Scholar] [CrossRef]
- Shin, D.J.; Germann, A.L.; Covey, D.F.; Steinbach, J.H.; Akk, G. Analysis of GABAA receptor activation by combinations of agonists acting at the same or distinct binding sites. Mol. Pharmacol. 2019, 95, 70–81. [Google Scholar] [CrossRef] [Green Version]
- Germann, A.L.; Steinbach, J.H.; Akk, G. Application of the Co-Agonist Concerted Transition Model to Analysis of GABAA Receptor Properties. Curr. Neuropharmacol. 2019, 17, 843–851. [Google Scholar] [CrossRef]
- Steinbach, J.H.; Akk, G. Applying the Monod-Wyman-Changeux Allosteric Activation Model to Pseudo-Steady-State Responses from GABAA Receptors. Mol. Pharmacol. 2019, 95, 106–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forman, S.A. Monod-Wyman-Changeux allosteric mechanisms of action and the pharmacology of etomidate. Curr. Opin. Anaesthesiol. 2012, 25, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Germann, A.L.; Pierce, S.R.; Senneff, T.C.; Burbridge, A.B.; Steinbach, J.H.; Akk, G. Steady-state activation and modulation of the synaptic-type alpha1beta2gamma2L GABAA receptor by combinations of physiological and clinical ligands. Physiol. Rep. 2019, 7, e14230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, P.M.; Lloyd, K.G.; Chadwick, D.W. CSF gradients for amino acid neurotransmitters. J. Neurol. Neurosurg. Psychiatry 1988, 51, 1193–1200. [Google Scholar] [CrossRef] [Green Version]
- Ferraro, T.N.; Hare, T.A. Triple-column ion-exchange physiological amino acid analysis with fluorescent detection: Baseline characterization of human cerebrospinal fluid. Anal. Biochem. 1984, 143, 82–94. [Google Scholar] [CrossRef]
- Curtis, D.R.; Watkins, J.C. The excitation and depression of spinal neurones by structurally related amino acids. J. Neurochem. 1960, 6, 117–141. [Google Scholar] [CrossRef]
- Jia, F.; Yue, M.; Chandra, D.; Keramidas, A.; Goldstein, P.A.; Homanics, G.E.; Harrison, N.L. Taurine is a potent activator of extrasynaptic GABA(A) receptors in the thalamus. J. Neurosci. 2008, 28, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierce, S.R.; Senneff, T.C.; Germann, A.L.; Akk, G. Steady-state activation of the high-affinity isoform of the alpha4beta2delta GABAA receptor. Sci. Rep. 2019, 9, 15997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akk, G.; Shin, D.J.; Germann, A.L.; Steinbach, J.H. GABA Type A Receptor Activation in the Allosteric Coagonist Model Framework: Relationship between EC50 and Basal Activity. Mol. Pharmacol. 2018, 93, 90–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusch, D.; Zhong, H.; Forman, S.A. Gating allosterism at a single class of etomidate sites on a1b2g2L GABAA receptors accounts for both direct activation and agonist modulation. J. Biol. Chem. 2004, 279, 20982–20992. [Google Scholar] [CrossRef] [Green Version]
- Concas, A.; Mostallino, M.C.; Porcu, P.; Follesa, P.; Barbaccia, M.L.; Trabucchi, M.; Purdy, R.H.; Grisenti, P.; Biggio, G. Role of brain allopregnanolone in the plasticity of gamma-aminobutyric acid type A receptor in rat brain during pregnancy and after delivery. Proc. Natl. Acad. Sci. USA 1998, 95, 13284–13289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, J.J.; Peters, J.A.; Sturgess, N.C.; Hales, T.G. Steroid modulation of the GABAA receptor complex: Electrophysiological studies. Ciba. Found Symp. 1990, 153, 56–71. [Google Scholar] [CrossRef] [PubMed]
- Puia, G.; Santi, M.R.; Vicini, S.; Pritchett, D.B.; Purdy, R.H.; Paul, S.M.; Seeburg, P.H.; Costa, E. Neurosteroids act on recombinant human GABAA receptors. Neuron 1990, 4, 759–765. [Google Scholar] [CrossRef]
- Luisi, S.; Petraglia, F.; Benedetto, C.; Nappi, R.E.; Bernardi, F.; Fadalti, M.; Reis, F.M.; Luisi, M.; Genazzani, A.R. Serum allopregnanolone levels in pregnant women: Changes during pregnancy, at delivery, and in hypertensive patients. J. Clin. Endocrinol. Metab. 2000, 85, 2429–2433. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, R.M.; Marini, H.; Kim, W.J.; Rogawski, M.A. Anticonvulsant activity of androsterone and etiocholanolone. Epilepsia 2005, 46, 819–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
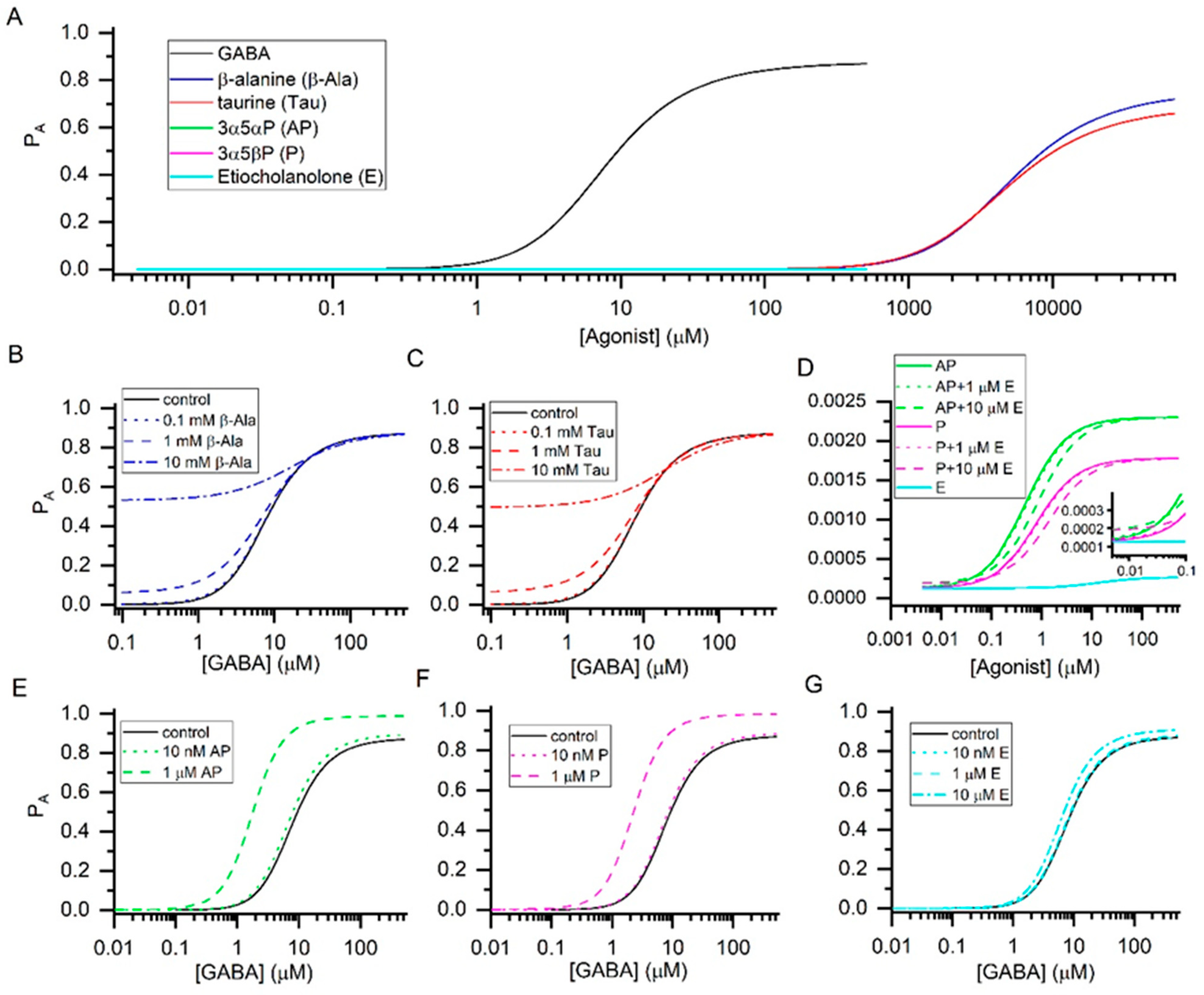

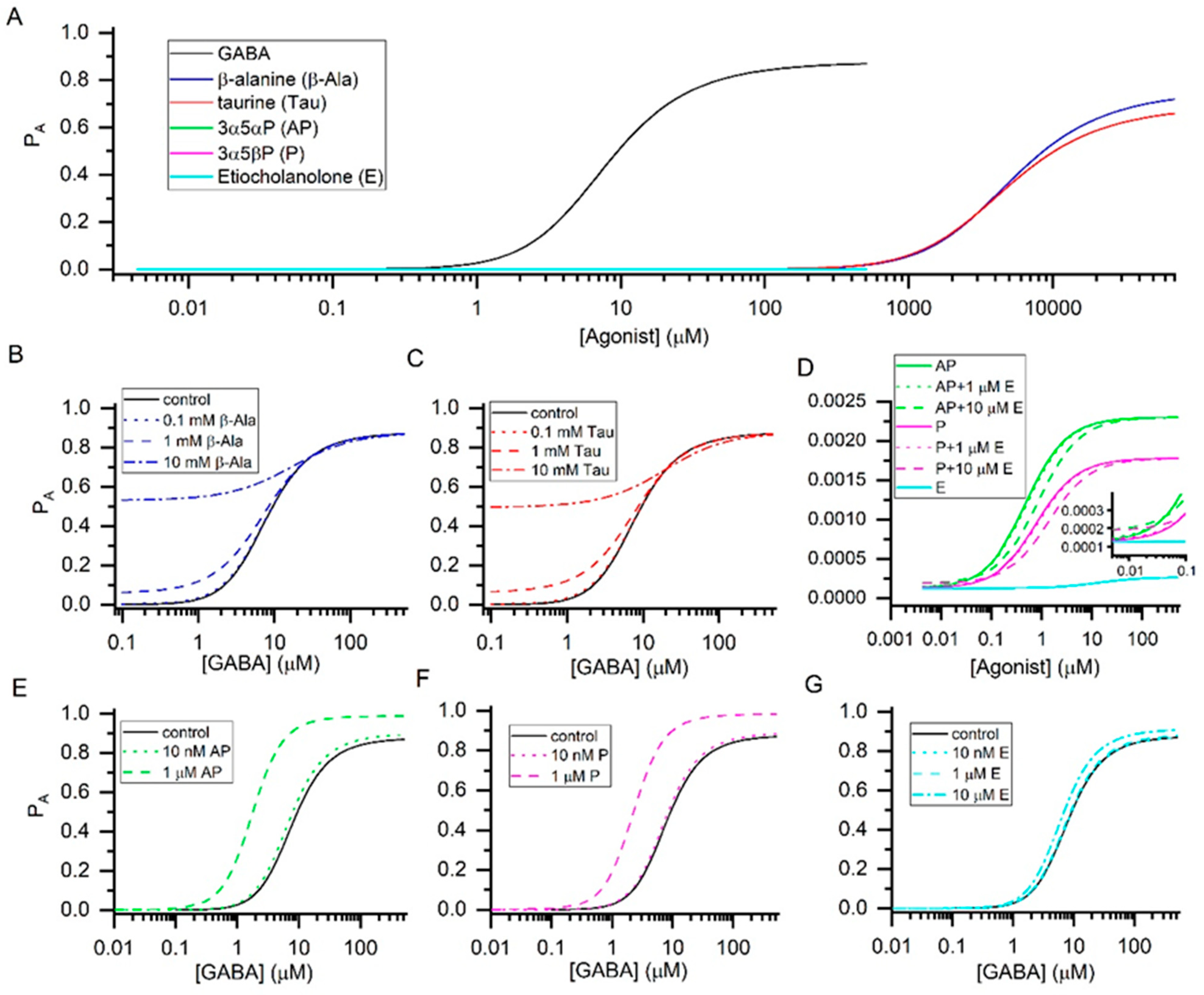{kind=link}
| Agonist | KR (µM) | c | Maximal PA |
|---|---|---|---|
| GABA | 16 ± 3 | 0.0042 ± 0.0003 | 0.88 ± 0.06 |
| β-Alanine | 6554 ± 1026 | 0.0064 ± 0.0010 | 0.75 ± 0.12 |
| Taurine | 5100 ± 1200 | 0.0075 ± 0.0006 | 0.69 ± 0.06 |
| 3α5αP | 0.27 ± 0.07 | 0.233 ± 0.018 | 0.0023 ± 0.0002 |
| 3α5βP | 0.45 ± 0.06 | 0.265 ± 0.010 | 0.0018 ± 0.0001 |
| Etiocholanolone | 11.1 ± 1.5 | 0.685 ± 0.009 | 0.0003 ± 0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pierce, S.R.; Germann, A.L.; Akk, G. Activation of the α1β2γ2L GABAA Receptor by Physiological Agonists. Biomolecules 2021, 11, 1864. https://doi.org/10.3390/biom11121864
Pierce SR, Germann AL, Akk G. Activation of the α1β2γ2L GABAA Receptor by Physiological Agonists. Biomolecules. 2021; 11(12):1864. https://doi.org/10.3390/biom11121864
Chicago/Turabian StylePierce, Spencer R., Allison L. Germann, and Gustav Akk. 2021. "Activation of the α1β2γ2L GABAA Receptor by Physiological Agonists" Biomolecules 11, no. 12: 1864. https://doi.org/10.3390/biom11121864
APA StylePierce, S. R., Germann, A. L., & Akk, G. (2021). Activation of the α1β2γ2L GABAA Receptor by Physiological Agonists. Biomolecules, 11(12), 1864. https://doi.org/10.3390/biom11121864