Research Progress of a Potential Bioreactor: Duckweed

 ,
,
Abstract
1. Introduction
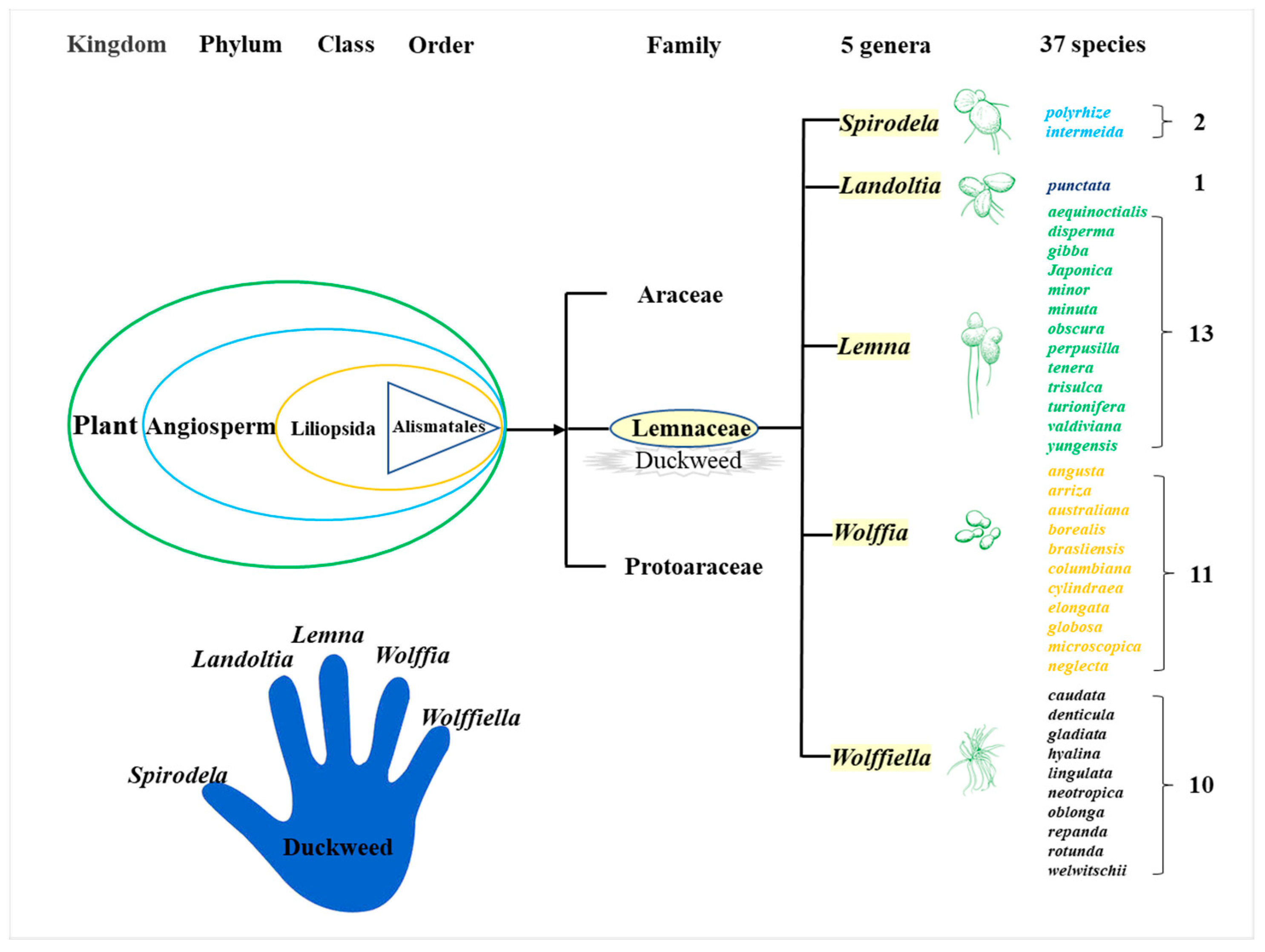
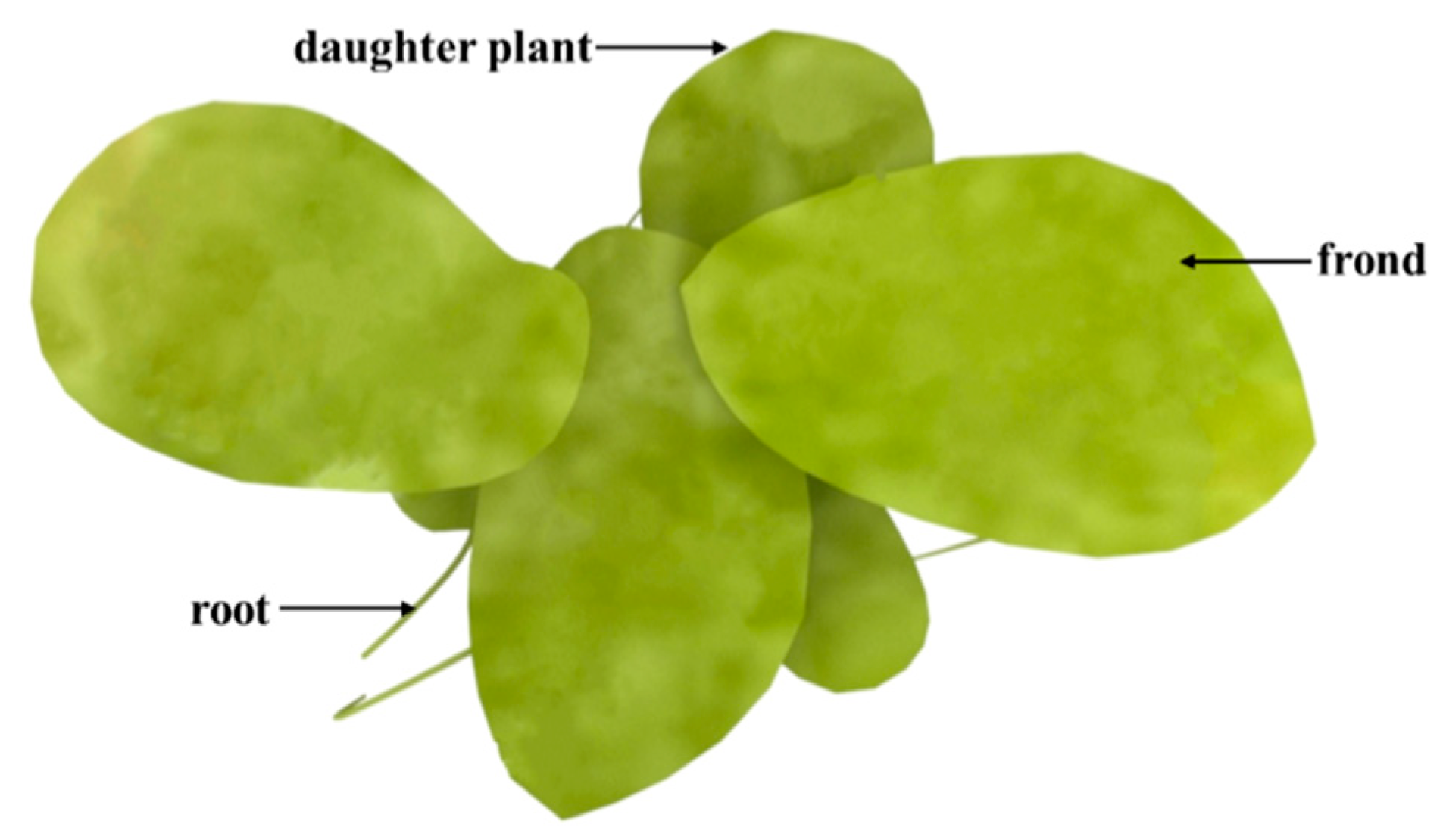
2. Biological Characteristics of Duckweed
2.1. Inherent Characteristics of Duckweed
2.2. Culture Conditions of Duckweed
3. Genetic Background of Duckweed
3.1. Chloroplast Genome of Duckweed
3.2. Mitochondrial Genome of Duckweed
3.3. Whole-Genome Sequencing of Duckweed
4. Advantages of the Duckweed Platform
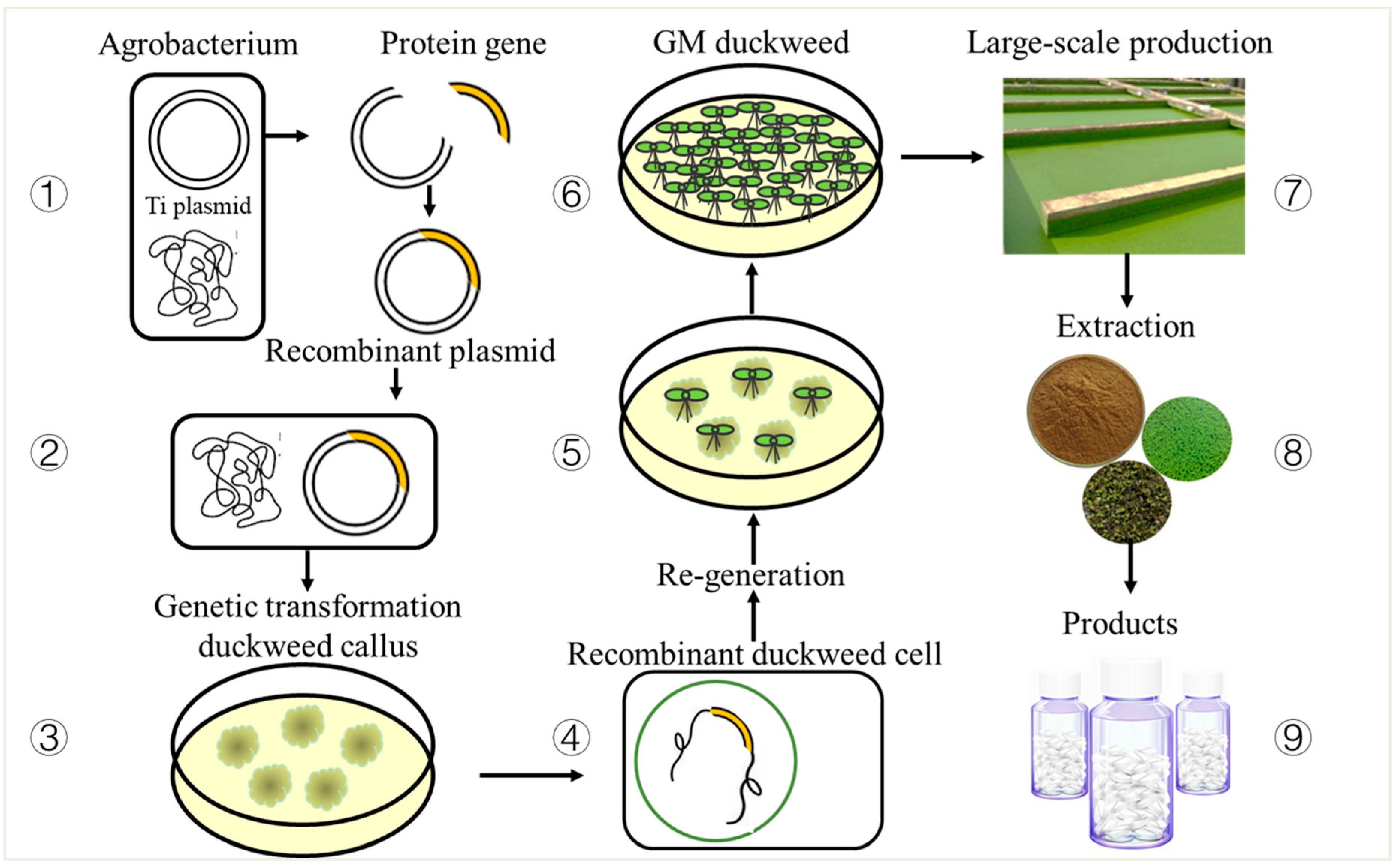
5. Genetic Transformation System for Duckweed
5.1. Duckweed Tissue Culture
5.2. Duckweed Transformation System
6. Synthetic Products of Duckweed
6.1. Vaccine Antigens
6.2. Therapeutic Products
6.3. Industrial Enzymes

| Species | Transformation System | Promoter | Product | Yield | Refs |
|---|---|---|---|---|---|
| Lemna minor | FTS | CaMV 35S | porcine epidemic diarrhea virus (PEDV) | can be detected by Western blotting | [67] |
| Lemna minor | CTS | CaMV 35S | M2e gene of avian influenza virus (H5N1) | 0.09–0.97 mg/g (FW) 0.12–1.96% (TSP) | [63] |
| Lemna minor | CTS | CaMV 35S | M2e fused to the ricin toxin B chain (MRTB–M130) | 0.25–2.5 µg/g (FW) 0.0006–0.01% (TSP) | [69] |
| Lemna minor | __ | __ | hemagglutinin (HA) | __ | [70] |
| Woffia globosa | CTS | CaMV 35S | LamB | can be detected by immunoblot | [60,71] |
| Lemna minor | CTS | mas | monoclonal antibody (mAb) | 2.1% (TSP) | [18,73] |
| Lemna minor | CTS | mas | human growth hormone | 609 mg/L (culture medium) | [72] |
| Lemna minor | CTS | mas | interferon-α2 | 30% (protein content of culture medium) | [72] |
| Lemna minor | CTS | mas | fragment of antigen binding (Fab) | 8.62 g/kg (DW) 4% TSP | [72] |
| Woffia arrhiza | CTS | CaMV 35S | human granulocyte colony-stimulating factor (G-CSF) | 35.5 mg/kg (FW) 0.194% (TSP) | [74] |
| Lemna minor | CTS | mas | plasminogen | 6.4% (TSP) | [75] |
| Lemna minor | CTS | mas | microplasminogen | 290 ng/mL (culture medium) | [75] |
| Spirodela oligorrhiza | CTS | CaMV 35S | aprotinin | 0.65 mg/L (culture medium) 3.7% (water soluble proteins) | [62] |
| Lemna minor | CTS | CaMV 35S | Endoglucanase E1 | Up to 0.24% (TSP) | [18,77] |
| Lemna minor | CTS | CaMV 35S | β-glucuronidase | 0.28 to 1.43% (TSP) | [65] |
| Lemna minor | CTS | CaMV 35S | hirudin | 0.02% (TSP) | [65] |
7. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, A.K.; Sharma, M.K. Plants as bioreactors: Recent developments and emerging opportunities. Biotechnol. Adv. 2009, 27, 811. [Google Scholar] [CrossRef]
- Rybicki, E.P. Plant-produced vaccines: Promise and reality. Drug Discov. Today 2009, 14, 16–24. [Google Scholar] [CrossRef]
- Goyal, R.; Sharma, R.; Lal, P.; Ramachandran, V.G. Edible vaccines: Current status and future. Indian J. Med. Microbiol. 2007, 25, 93–102. [Google Scholar] [CrossRef]
- He, Z.Q.; Du, X.; Yao, W.; Dai, J. Pharmaceutical proteins produced in plant bioreactor in recent years. Afr. J. Biotechnol. 2008, 7, 4917–4925. [Google Scholar]
- Tiwari, S.; Verma, P.C.; Singh, P.K.; Tuli, R. Plants as bioreactors for the production of vaccine antigens. Biotechnol. Adv. 2009, 27, 449–467. [Google Scholar] [CrossRef]
- Jani, D.; Meena, L.S.; Rizwan-Ul-Haq, Q.M.; Singh, Y.; Sharma, A.K.; Tyagi, A.K. Expression of cholera toxin B subunit in transgenic tomato plants. Transgenic Res. 2001, 11, 447–454. [Google Scholar] [CrossRef]
- Langridge, W.H.R. A plant-based multicomponent vaccine protects mice from enteric diseases. Nat. Biotechnol. 2001, 19, 548. [Google Scholar]
- Marquet-Blouin, E.; Bouche, F.; Steinmetz, A.; Muller, C. Neutralizing immunogenicity of transgenic carrot (Daucus carota L.)-derived measles virus hemagglutinin. Plant Mol. Biol. 2003, 51, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Giddings, G. Transgenic plants as factories for biopharmaceuticals. Nat. Biotechnol. 2000, 18, 1151. [Google Scholar] [CrossRef]
- Tremblay, R.; Wang, D.; Jevnikar, A.M.; Ma, S. Tobacco, a highly efficient green bioreactor for production of therapeutic proteins. Biotechnol. Adv. 2010, 28, 214–221. [Google Scholar] [CrossRef]
- Yano, M.; Hirai, T.; Kato, K.; Hiwasa-Tanase, K.; Fukuda, N.; Ezura, H. Tomato is a suitable material for producing recombinant miraculin protein in genetically stable manner. Plant Sci. 2010, 178, 469–473. [Google Scholar] [CrossRef]
- Kim, Y.M.; Lee, J.-Y.; Lee, T.; Lee, Y.-H.; Kim, S.-H.; Kang, S.-H.; Yoon, U.-H.; Ha, S.-H.; Lim, S.-H. The suppression of the glutelin storage protein gene in transgenic rice seeds results in a higher yield of recombinant protein. Plant Biotechnol. Rep. 2012, 6, 347–353. [Google Scholar] [CrossRef]
- Jane, B.; Lancet, B.J. Progress in potato-based vaccine for hepatitis B. Lancet 2000, 356, 1661. [Google Scholar]
- Nguyen, L.V.; Cox, K.M.; Ke, J.S.; Peele, C.G.; Dickey, L.F. Genetic engineering of a Lemna isoleucine auxotroph. Transgenic Res. 2012, 21, 1071–1083. [Google Scholar] [CrossRef]
- Ziegler, P.; Adelmann, K.; Zimmer, S.; Schmidt, C.; Appenroth, K.-J. Relative in vitro growth rates of duckweeds (Lemnaceae)–the most rapidly growing higher plants. Plant Biol. 2015, 17, 33–41. [Google Scholar] [CrossRef]
- Stomp, A.M. The duckweeds: A valuable plant for biomanufacturing. Biotechnol. Annu. Rev. 2005, 11, 69. [Google Scholar]
- Stomp, A.M.; Rajbhandari, N. Genetically Engineered Duckweed. U.S. Patent EP2283721(A3), 31 October 2012. [Google Scholar]
- Yamamoto, Y.T.; Rajbhandari, N.; Lin, X.; Bergmann, B.A.; Nishimura, Y.; Stomp, A.-M. Genetic transformation of duckweed Lemna gibba and Lemna minor. In Vitro Cell Dev. Biol. Plant 2001, 37, 349–353. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, H.; Stomp, A.-M.; Cheng, J.J. The production of duckweed as a source of biofuels. Biofuels 2012, 3, 589–601. [Google Scholar] [CrossRef]
- Wang, W.; Messing, J. Status of duckweed genomics and transcriptomics. Plant Biol. 2014, 17, 10–15. [Google Scholar] [CrossRef]
- An, D.; Li, C.; Zhou, Y.; Wu, Y.; Wang, W. Genomes and transcriptomes of duckweeds. Front. Chem. 2018, 6, 230. [Google Scholar] [CrossRef]
- Klaus, J.A.; Nikolai, B.; Eric, L. Telling duckweed apart: Genotyping technologies for the Lemnaceae. Chin. J. Appl. Environ. Biol. 2013, 19, 1–10. [Google Scholar] [CrossRef]
- Ekperusi, A.O.; Sikoki, F.D.; Nwachukwu, E.O. Application of common duckweed (Lemna minor) in phytoremediation of chemicals in the environment: State and future perspective. Chemosphere 2019, 223, 285–309. [Google Scholar] [CrossRef] [PubMed]
- Les, D.H.; Crawford, D.J.; Landolt, E.; Gabel, J.D.; Kimball, R.T. Phylogeny and systematics of Lemnaceae, the duckweed family. Syst. Bot. 2002, 27, 221–240. [Google Scholar]
- Xu, Y.L.; Ma, S.; Huang, M.; Peng, M.; Bog, M.; Sree, K.S.; Appenroth, K.-J.; Zhang, J. Species distribution, genetic diversity and barcoding in the duckweed family (Lemnaceae). Hydrobiologia 2015, 743, 75–87. [Google Scholar] [CrossRef]
- Les, D.H.; Crawford, D.J. Landoltia (Lemnaceae), a new genus of duckweeds. Novon 1999, 9, 530–533. [Google Scholar] [CrossRef]
- Hillman, W.S. The Lemnaceae or duckweeds. A review of the descriptive and experimental literature. Bot. Rev. 1961, 27, 221–287. [Google Scholar] [CrossRef]
- Thompson, B.G. The maximization of the productivity of aquatic plants for use in controlled ecological life support systems (CELSS). Acta Astronaut. 1989, 19, 269–273. [Google Scholar] [CrossRef]
- Frick, H. Callogenesis and carbohydrate utilization in Lemna minor 1. J. Plant Physiol. 1991, 137, 397–401. [Google Scholar] [CrossRef]
- Mardanov, A.V.; Ravin, N.V.; Kuznetsov, B.B.; Samigullin, T.H.; Antonov, A.S.; Kolganova, T.V.; Skyabin, K.G. Complete sequence of the duckweed (Lemna minor) chloroplast genome: Structural organization and phylogenetic relationships to other angiosperms. J. Mol. Evol. 2008, 66, 555–564. [Google Scholar] [CrossRef]
- Wang, W.Q.; Messing, J. High-throughput sequencing of three Lemnoideae (duckweeds) chloroplast genomes from total DNA. PLoS ONE 2011, 6, e24670. [Google Scholar] [CrossRef]
- Ding, Y.; Fang, Y.; Guo, L.; Li, Z.; He, K.; Zhao, J.; Zhao, H. Phylogenic study of Lemnoideae (duckweeds) through complete chloroplast genomes for eight accessions. PeerJ 2017, 5, e4186. [Google Scholar] [CrossRef]
- Zhang, Y.; An, D.; Li, C.; Zhao, Z.; Wang, W. The complete chloroplast genome of greater duckweed (Spirodela polyrhiza 7498) using PacBio long reads: Insights into the chloroplast evolution and transcription regulation. BMC Genom. 2020, 21, 76. [Google Scholar] [CrossRef]
- Korotkova, N.; Nauheimer, L.; Ter-Voskanyan, H.; Allgaier, M.; Borsch, T. Variability among the most rapidly evolving plastid genomic regions is lineage-specific: Implications of pairwise genome comparisons in Pyrus (Rosaceae) and other angiosperms for marker choice. PLoS ONE 2014, 9, e112998. [Google Scholar] [CrossRef]
- Jansen, R.K.; Raubeson, L.A.; Boore, J.L.; Depamphilis, C.W.; Chumley, T.W.; Haberle, R.C.; Wyman, S.K.; Alverson, A.J.; Peery, R.; Herman, S.J.; et al. Methods for obtaining and analyzing whole chloroplast genome sequences. Methods Enzymol. 2005, 395, 348. [Google Scholar]
- Wakasugi, T.; Tsudzuki, T.; Sugiura, M. The genomics of land plant chloroplasts: Gene content and alteration of genomic information by RNA editing. Photosynth Res. 2001, 70, 107–118. [Google Scholar] [CrossRef]
- Christensen, A.C. plant mitochondrial genome evolution can be explained by DNA repair mechanisms. Genome Biol. Evol. 2013, 5, 1079–1086. [Google Scholar] [CrossRef]
- Mackenzie, S.; McIntosh, L. Higher plant mitochondria. Plant Cell 1999, 11, 571–586. [Google Scholar]
- Wang, W.; Wu, Y.; Messing, J. The mitochondrial genome of an aquatic plant, Spirodela polyrhiza. PLoS ONE 2012, 7, e46747. [Google Scholar] [CrossRef]
- Wang, W.; Haberer, G.; Gundlach, H.; Gläßer, C.; Nussbaumer, T.; Luo, M.; Lomsadze, A.; Borodovsky, M.; A Kerstetter, R.; Shanklin, J.D.; et al. The Spirodela polyrhiza genome reveals insights into its neotenous reduction fast growth and aquatic lifestyle. Nat. Commun. 2014, 5, 3311. [Google Scholar] [CrossRef]
- Michael, T.P.; Bryant, D.; Gutierrez, R.; Borisjuk, N.; Chu, P.; Zhang, H.; Xia, J.; Zhou, J.; Peng, H.; El Baidouri, M.; et al. Comprehensive definition of genome features in Spirodela polyrhiza by high-depth physical mapping and short-read DNA sequencing strategies. Plant J. 2017, 89, 617–635. [Google Scholar] [CrossRef]
- Hoeck, A.V.; Horemans, N.; Monsieurs, P.; Cao, H.X.; Vandenhove, H.; Blust, R. The first draft genome of the aquatic model plant Lemna minoropens the route for future stress physiology research and biotechnological applications. Biotechnol. Biofuels 2015, 8, 188. [Google Scholar] [CrossRef] [PubMed]
- Ernst, E. Status of the Lemna gibba 7742a and Lemna minor 8627 genomes. ISCDRA 2016, 3, 9–10. [Google Scholar]
- Appenroth, K.J.; Sree, K.S.; Fakhoorian, T.; Lam, E. Resurgence of duckweed research and applications: Report from the third International Duckweed Conference. Plant Mol. Biol. 2015, 89, 647–654. [Google Scholar] [CrossRef]
- Hoang, P.T.N.; Schubert, V.; Meister, A.; Fuchs, J.; Schubert, I. Variation in genome size, cell and nucleus volume, chromosome number and rDNA loci among duckweeds. Sci. Rep. 2019, 9, 3234. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Kerstetter, R.A.; Michael, T.P. Evolution of genome size in duckweeds (Lemnaceae). J. Bot. 2011, 570319. [Google Scholar] [CrossRef]
- An, D.; Zhou, Y.; Li, C.; Xiao, Q.; Wang, T.; Zhang, Y.; Wu, Y.; Li, Y.; Chao, D.-Y.; Messing, J.; et al. Plant evolution and environmental adaptation unveiled by long-read whole-genome sequencing of Spirodela. Proc. Natl. Acad. Sci. USA 2019, 116, 201910401. [Google Scholar] [CrossRef] [PubMed]
- Hull, A.K.; Criscuolo, C.; Mett, V.; Groen, H.; Steeman, W.; Westra, H.; Chapman, G.; Legutki, B.; Baillie, L.; Yusibov, V. Human-derived, plant-produced monoclonal antibody for the treatment of anthrax. Vaccine 2005, 23, 2082–2086. [Google Scholar] [CrossRef]
- Lam, E.; Appenroth, K.J.; Michael, T.; Mori, K.; Fakhoorian, T. Duckweed in bloom: The second International Conference on Duckweed Research and Applications heralds the return of a plant model for plant biology. Plant Mol. Biol. 2014, 84, 737–742. [Google Scholar] [CrossRef]
- Peng, Y.L.; Zhao, H.; Gao, X.F. A Method for Preserving Duckweed Living Materials. China Patent CN104025989B, 31 August 2016. [Google Scholar]
- Iqbal, S. Duckweed Aquaculture-Potentials, Possibilities and Limitations, for Combined Wastewater Treatment and Animal Feed Production in Developing Countries; SANDEC Report; EAWAG: Dübendorf, Switzerland, 1999; p. 99. [Google Scholar]
- Chang, W.C.; Chiu, P.L. Regeneration of Lemna-Gibba G-3 through callus-culture. Ztschrift Für Pflanzenphysiologie 1978, 89, 91–94. [Google Scholar] [CrossRef]
- Yu, L.; Wang, Y.; Xu, S.; Tang, X.; Zhao, J.; Yu, C.; He, G.; Xu, H.; Wang, S.; Tang, Y.; et al. Efficient genetic transformation and CRISPR/Cas9-mediated genome editing in Lemna aequinoctialis. Plant Biotechnol. J. 2019, 17, 2143–2152. [Google Scholar]
- Chhabra, G.; Chaudhary, D.; Sain00ger, M.; Jaiwal, P.K. Genetic transformation of Indian isolate of Lemna minor mediated by Agrobacterium tumefaciens and recovery of transgenic plants. Physiol. Mol. Biol. Plants 2011, 17, 129–136. [Google Scholar] [CrossRef]
- Yang, G.L.; Fang, Y.; Xu, Y.-L.; Tan, L.; Li, Q.; Liu, Y.; Lai, F.; Jin, Y.-L.; Du, A.-P.; He, K.; et al. Frond transformation system mediated by Agrobacterium tumefaciens for Lemna minor. Plant Mol. Biol. 2018, 98, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Han, Y.; Wu, D.; Yong, W.; Liu, M.; Wang, S.; Liu, W.; Lu, M.; Wei, Y.; Sun, J. Salt and cadmium stress tolerance caused by overexpression of the Glycine Max Na+/H+ Antiporter (GmNHX1) gene in duckweed (Lemna turionifera 5511). Aquat. Toxicol. 2017, 192, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Jain, M.; Vunsh, R.; Vishnevetsky, J.; Hanania, U.; Flaishman, M.; Perl, A.; Edelman, M. Callus induction and regeneration in Spirodela and Lemna. Plant Cell Rep. 2004, 22, 457–464. [Google Scholar] [CrossRef]
- Huang, M.; Fu, L.; Sun, X.; Di, R.; Zhang, J. Rapid and highly efficient callus induction and plant regeneration in the starch-rich duckweed strains of Landoltia punctata. Acta Physiol. Plant 2016, 38, 122. [Google Scholar] [CrossRef]
- Khvatkov, P.; Chernobrovkina, M.; Okuneva, A.; Shvedova, A.; Chaban, I.; Dolgov, S. Callus induction and regeneration in Wolffia arrhiza (L.) Horkel ex Wimm. Plant Cell Tissue Organ Cult. 2014, 120, 263–273. [Google Scholar] [CrossRef]
- Heenatigala, P.P.M.; Yang, J.; Bishopp, A.; Sun, Z.; Li, G.; Kumar, S.; Hu, S.; Wu, Z.; Lin, W.; Yao, L.; et al. Development of efficient protocols for stable and transient gene transformation for Wolffia globosa using Agrobacterium. Front. Chem. 2018, 6, 227. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Ma, X.; Zhong, Y.; Hu, Q.; Fu, M.; Han, Y. Callus induction and plant regeneration of Spirodela polyrhiza. Plant Cell Tissue Organ Cult. 2018, 135, 445–453. [Google Scholar] [CrossRef]
- Rival, S.; Wisniewski, J.-P.; Langlais, A.; Kaplan, H.; Freyssinet, G.; Vancanneyt, G.; Vunsh, R.; Perl, A.; Edelman, M. Spirodela (duckweed) as an alternative production system for pharmaceuticals: A case study, aprotinin. Transgenic Res. 2008, 17, 503–513. [Google Scholar] [CrossRef]
- Firsov, A.; Tarasenko, I.; Mitiouchkina, T.Y.; Ismailova, N.; Shaloiko, L.; Vainstein, A.; Dolgov, S. High-yield expression of M2e peptide of avian influenza virus H5N1 in transgenic duckweed plants. Mol. Biotechnol. 2015, 57, 653–661. [Google Scholar] [CrossRef]
- Cantó-Pastor, A.; Mollá-Morales, A.; Ernst, E.; Dahl, W.; Zhai, J.; Yan, Y.; Meyers, B.C.; Shanklin, J.; A Martienssen, R. Efficient transformation and artificial miRNA gene silencing in Lemna minor. Plant Biol. 2015, 17, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, O.N.; Mitiouchkina, T.Y.; Tarasenko, I.V.; Shaloiko, L.A.; Firsov, A.; Dolgov, S.V. Agrobacterium-mediated transformation of Lemna minor L. with Hirudin and β-glucuronidase genes. Appl. Biochem. Microbiol. 2019, 55, 805–815. [Google Scholar] [CrossRef]
- Kruse, C.; Boehm, R.; Voeste, D.; Barth, S.; Schnabl, H. Transient transformation of Wolffia columbiana by particle bombardment. Aquat. Bot. 2002, 72, 175–181. [Google Scholar] [CrossRef]
- Ko, S.M.; Sun, H.-J.; Oh, M.J.; Song, I.-J.; Kim, M.; Sin, H.-S.; Goh, C.-H.; Kim, Y.-W.; Lim, P.-O.; Lee, H.-Y.; et al. Expression of the protective antigen for PEDV in transgenic duckweed, Lemna minor. Hortic. Environ. Biotechnol. 2011, 52, 511. [Google Scholar] [CrossRef]
- Khvatkov, P.; Chernobrovkina, M.; Okuneva, A.; Pushin, A.; Dolgov, S. Transformation of Wolffia arrhiza (L.) Horkel ex Wimm. Plant Cell Tissue Organ Cult. 2015, 123, 299–307. [Google Scholar] [CrossRef]
- Firsov, A.; Tarasenko, I.; Mitiouchkina, T.; Shaloiko, L.; Kozlov, O.; Vinokurov, L.; Rasskazova, E.; Murashev, A.; Vainstein, A.; Dolgov, S. Expression and immunogenicity of M2e peptide of avian influenza virus H5N1 fused to ricin toxin B chain produced in duckweed plants. Front. Chem. 2018, 6, 22. [Google Scholar] [CrossRef]
- Bertran, K. Expression of H5 hemagglutinin vaccine antigen in common duckweed (Lemna minor) protects against H5N1 high pathogenicity avian influenza virus challenge in immunized chickens. Vaccine 2015, 33, 3456–3462. [Google Scholar] [CrossRef]
- Heenatigala, P.P.M.; Sun, Z.; Yang, J.; Zhao, X.; Hou, H. Expression of LamB vaccine antigen in Wolffia globosa (duck weed) against fish vibriosis. Front. Immunol. 2020, 11, 9. [Google Scholar] [CrossRef]
- Dickey, L.; Gasdaska, J.; Cox, K. Expression of Biologically Active Polypeptides in Duckweed. U.S. Patent 6,815,184, 11 September 2004. [Google Scholar]
- Cox, K.M.; Sterling, J.D.; Regan, J.T.; Gasdaska, J.R.; Frantz, K.K.; Peele, C.G.; Black, A.; Passmore, D.; Moldovan-Loomis, C.; Srinivasan, M.; et al. Glycan optimization of a human monoclonal antibody in the aquatic plant Lemna minor. Nat. Biotechnol. 2006, 24, 1591–1597. [Google Scholar] [CrossRef]
- Khvatkow, P.; Firsov, A.; Shvedova, A.; Shaloiko, L.; Kozlov, O.; Chernobrovkina, M.; Pushin, A.; Tarasenko, I.; Chaban, I.; Dolgov, S. Development of Wolffia arrhiza as a producer for recombinant human granulocyte colony-stimulating factor. Front. Chem. 2018, 6, 304. [Google Scholar] [CrossRef]
- Spencer, D.; Dickey, L.F.; Gasdaska, J.R.; Wang, X.; Cox, K.M.; Peele, C.G. Expression of Plasminogen and Microplasminogen in Duckweed. U.S. Patent AU2005212431(B2), 15 July 2010. [Google Scholar]
- Sun, Y.; Cheng, J.J.; Himmel, M.E.; Skory, C.D.; Adney, W.S.; Thomas, S.R.; Tisserat, B.; Nishimura, Y.; Yamamoto, Y.T. Expression and characterization of Acidothermus cellulolyticus E1 endoglucanase in transgenic duckweed Lemna minor 8627. Bioresour. Technol. 2007, 98, 2866–2872. [Google Scholar] [CrossRef] [PubMed]
- Hobom, B. Gene surgery: On the threshold of synthetic biology. Med. Klin. 1980, 75, 834. [Google Scholar] [PubMed]
- Fischer, R.; Emans, N. Molecular farming of pharmaceutical proteins. Transgenic Res. 2000, 9, 279–299. [Google Scholar] [CrossRef] [PubMed]
- Daniell, H.; Streatfield, S.J.; Wycoff, K. Medical molecular farming: Production of antibodies, biopharmaceuticals and edible vaccines in plants. Trends Plant Sci. 2001, 6, 219–226. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}


| Genera | Species | CGS (bp) | IRs (bp) | GN | References |
|---|---|---|---|---|---|
| Spirodela | Spirodela polyrhiza 7498 | 168,956 | 31,844 | MN419335 | [33] |
| Landoltia | Landoltia punctata ZH0202 | 171,013 | 31,899 | KY993962 | [32] |
| Lemna | Lemna minor | 165,955 | 31,223 | DQ400350 | [30] |
| Woffia | Woffia australiana 7733 | 168,704 | 31,930 | JN160605 | [31] |
| Woffiella | Woffiella lingulata 7289 | 169,337 | 31,683 | JN160604 | [31] |
| Species | Genome Size (Mb) | Platform | Sequencing Coverage | Protein Coding Gene | Scaffold N50 | Contig N50 (Kb) | Repeat (%) | References |
|---|---|---|---|---|---|---|---|---|
| Spirodela polyriza 7498 | 158 | 454 and Sanger | 21 | 19,623 | 3.8 Mb | 18 | 17 | [40] |
| Spirodela polyriza 9509 | 160 | Illumina and BioNano | 95 | 18,507 | 7.6 Mb | 19 | 23.8 | [41] |
| Lemna minor 5500 | 481 | Illumina | 120 | 22,382 | 23.6 Kb | 20.9 | 61.5 | [42] |
| Lemna minor 8627 | 800 | Illumina and PacBio | __ | __ | __ | 222 | __ | [43] |
| Lemna gibba 7742a | 450 | Illumina | __ | 21,830 | 520 Kb | 53 | __ | [43] |
| Plant | Duckweed | Soybean | Peanut | Alfalfa |
|---|---|---|---|---|
| Dry biomass (t/ha/year) | 17.6 | 1.59 | 1.6–3.12 | 4.37–15.69 |
| Crude protein content (%) | 37 | 42 | 23.6 | 15.9–17 |
| Genera | Species | Callus Induction | Plant Regeneration | Refs | ||
|---|---|---|---|---|---|---|
| BM | Supplements (mg/L) | BM | Supplements (mg/L) | |||
| Spirodela | Spirodela oligorrhiza SP | WP + 2% So + 1% Ma | PCA (5) + 2IP (2) | WP + 0.5% Su | TDZ (1) | [57] |
| Spirodela polyrhiza 5543 | 1/2 MS + 1% So | 2,4-D (5) + 6-BA (2) | 1/2 MS + 1% So | 2,4-D (5) + 6-BA (2) | [61] | |
| Landoltia | Landoltia punctata 8717 | 1/2 MS + 1% So | 2,4-D (3.5) + Di (15) + 2IP (2) | WP + 0.5% Su + 1% So | 2IP (1) | [57] |
| Landoltia punctata 5502, 8721, and 9264 | MS + 1% So | 2, 4-D (15) + 6-BA (2) | MS + 0.5% Su | 6-BA (1) | [58] | |
| Lemna | Lemna gibba G3 | MS + 3% Su | 2, 4-D (10) + 2IP (1) | MS + 3% Su | IAA (4) + KT (1) | [52] |
| Lemna gibba var. Hurfeish | B5 + 1% Su | Di (50) + 6-BA (15) | B5 + 1% Su | TDZ (1) | [57] | |
| Lemna aequinotialis 6002 | MS + 3% Su | 2, 4-D (1) + TDZ (0.1) | B5 + 1% Su | IAA (0.45) + KT (1) | [53] | |
| Lemna minor | B5 + 1% Su | 2, 4-D (11) + TDZ (1.1) | B5 + 1% Su | IAA (4.4) + KT (1.1) | [54] | |
| Lemna minor ZH0055 | MS + 3% Su | 2, 4-D (10) + TDZ (5) | MS + 3% Su | IAA (4.4) + KT (1.1) | [55] | |
| Lemna turionifera 5511 | B5 + 1.5% Su | 2,4-D (1) + Di (15) + 6-BA (1) | B5 + 1.5% Su | L-serine (105) | [56] | |
| Woffia | Woffia arrhiza (L.) Horkel ex Wimm | SH + 2% Su | 2, 4-D (5) + 6-BA (0.5) | SH + 2% Su | regulator-free | [59] |
| Woffia globosa | SH + 2% Su | 2, 4-D (5) + 6-BA (0.5) | SH + 2% Su | regulator-free | [59,60] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.-L.; Feng, D.; Liu, Y.-T.; Lv, S.-M.; Zheng, M.-M.; Tan, A.-J. Research Progress of a Potential Bioreactor: Duckweed. Biomolecules 2021, 11, 93. https://doi.org/10.3390/biom11010093
Yang G-L, Feng D, Liu Y-T, Lv S-M, Zheng M-M, Tan A-J. Research Progress of a Potential Bioreactor: Duckweed. Biomolecules. 2021; 11(1):93. https://doi.org/10.3390/biom11010093
Chicago/Turabian StyleYang, Gui-Li, Dan Feng, Yu-Ting Liu, Shi-Ming Lv, Meng-Meng Zheng, and Ai-Juan Tan. 2021. "Research Progress of a Potential Bioreactor: Duckweed" Biomolecules 11, no. 1: 93. https://doi.org/10.3390/biom11010093
APA StyleYang, G.-L., Feng, D., Liu, Y.-T., Lv, S.-M., Zheng, M.-M., & Tan, A.-J. (2021). Research Progress of a Potential Bioreactor: Duckweed. Biomolecules, 11(1), 93. https://doi.org/10.3390/biom11010093