Metabolism of Soy Isoflavones by Intestinal Bacteria: Genome Analysis of an Adlercreutzia equolifaciens Strain That Does Not Produce Equol




Abstract
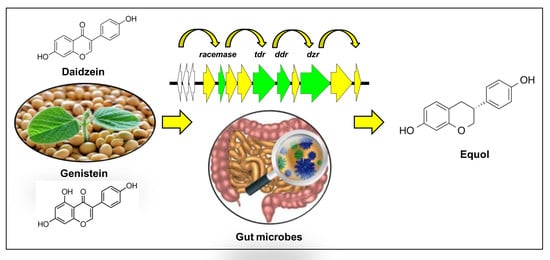
:
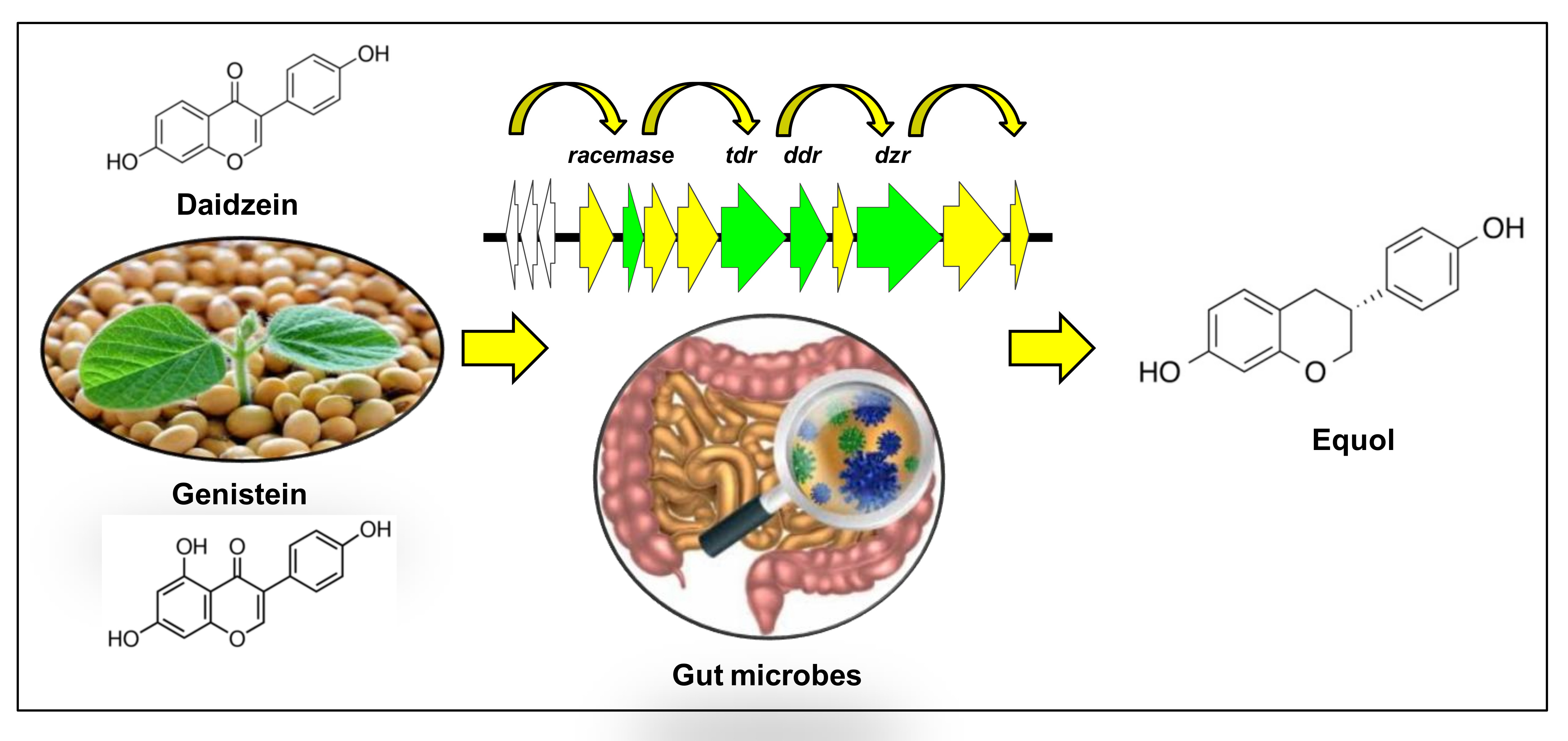
1. Introduction
2. Materials and Methods
2.1. Bacteria and Culture Conditions
2.2. Quantitative Real-Time PCR (qPCR)
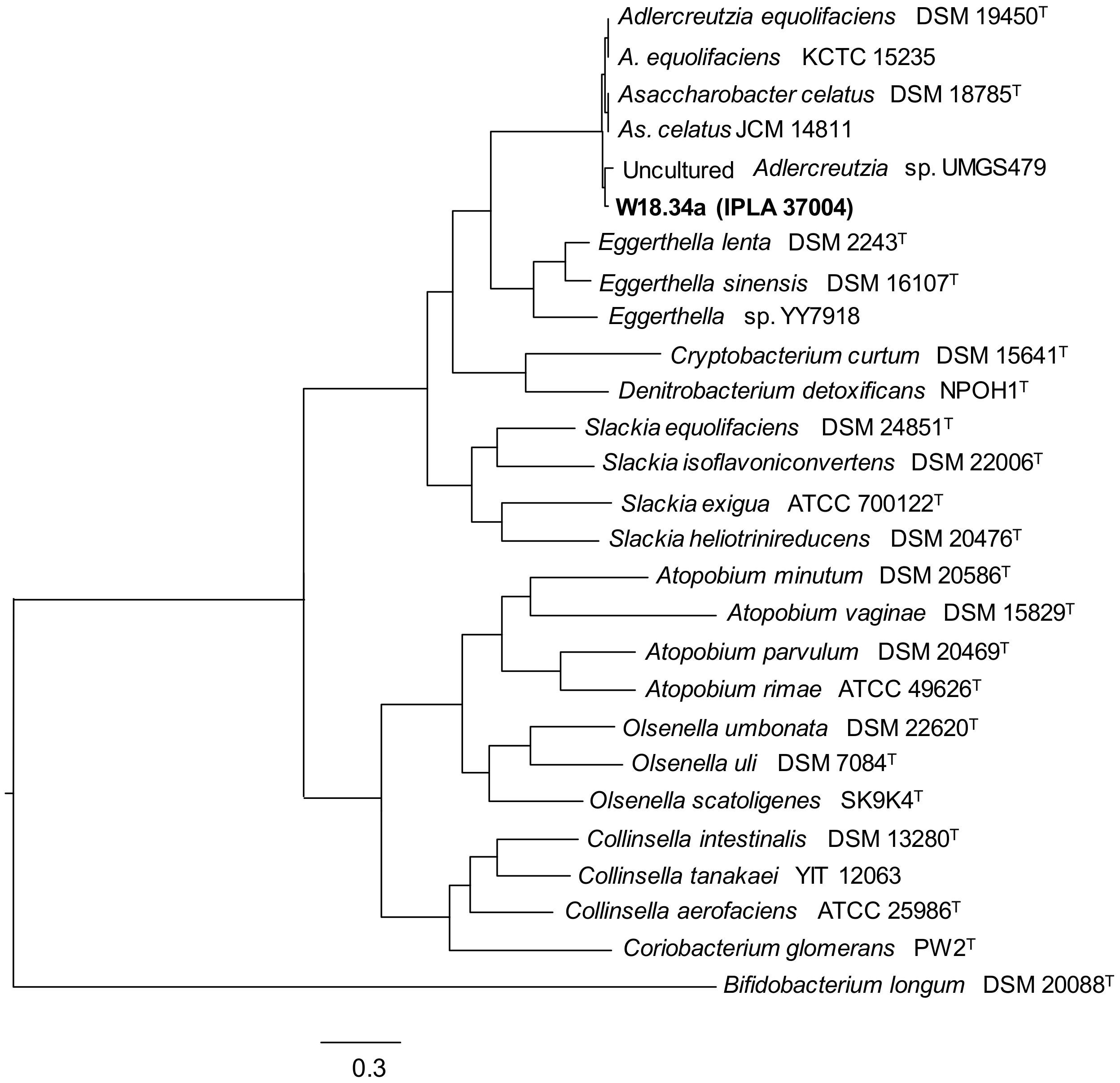
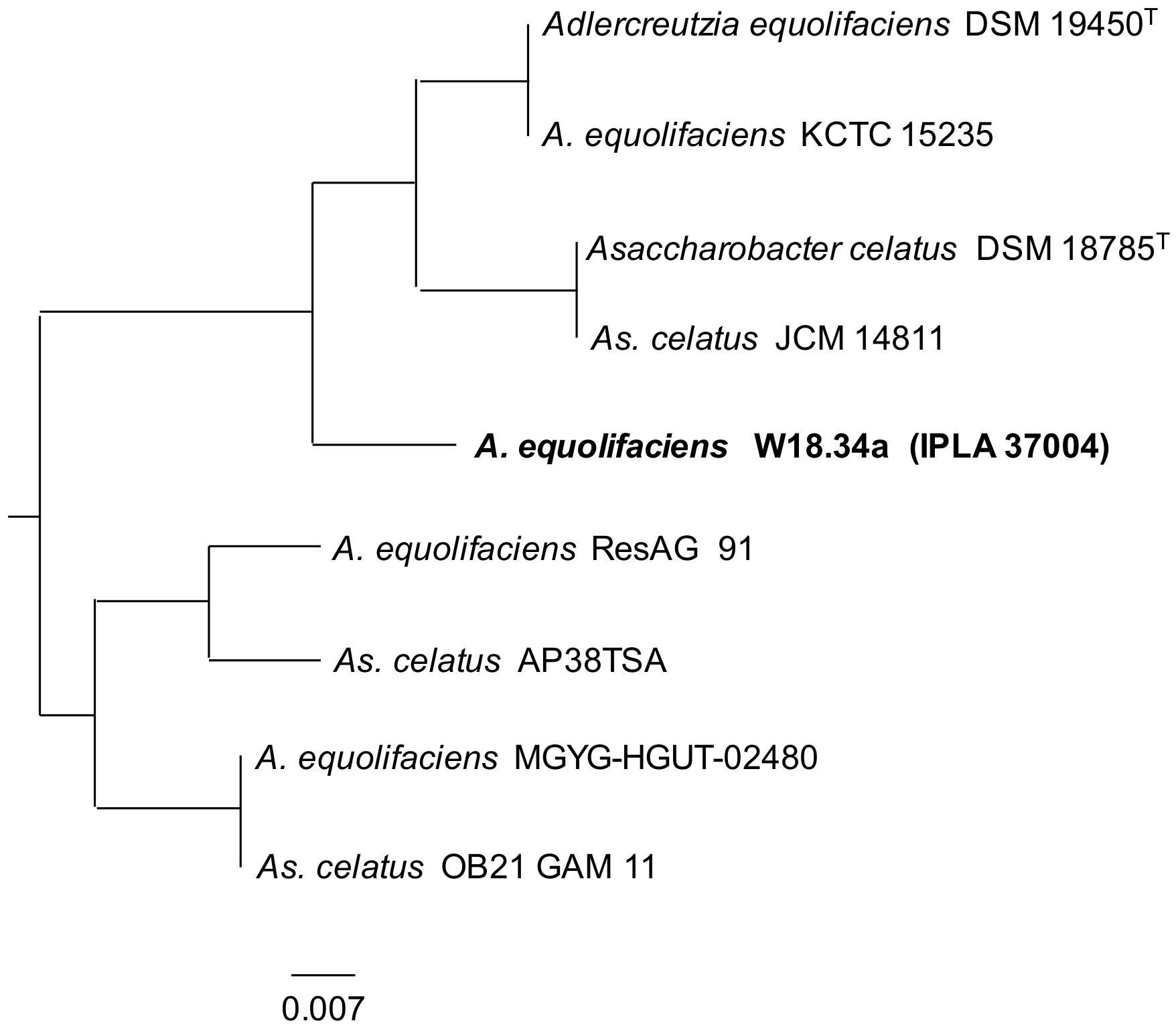
2.3. Identification of Bacteria
2.4. Typing of Isolates
2.5. Detection and Quantification of Isoflavones and Isoflavone Metabolites
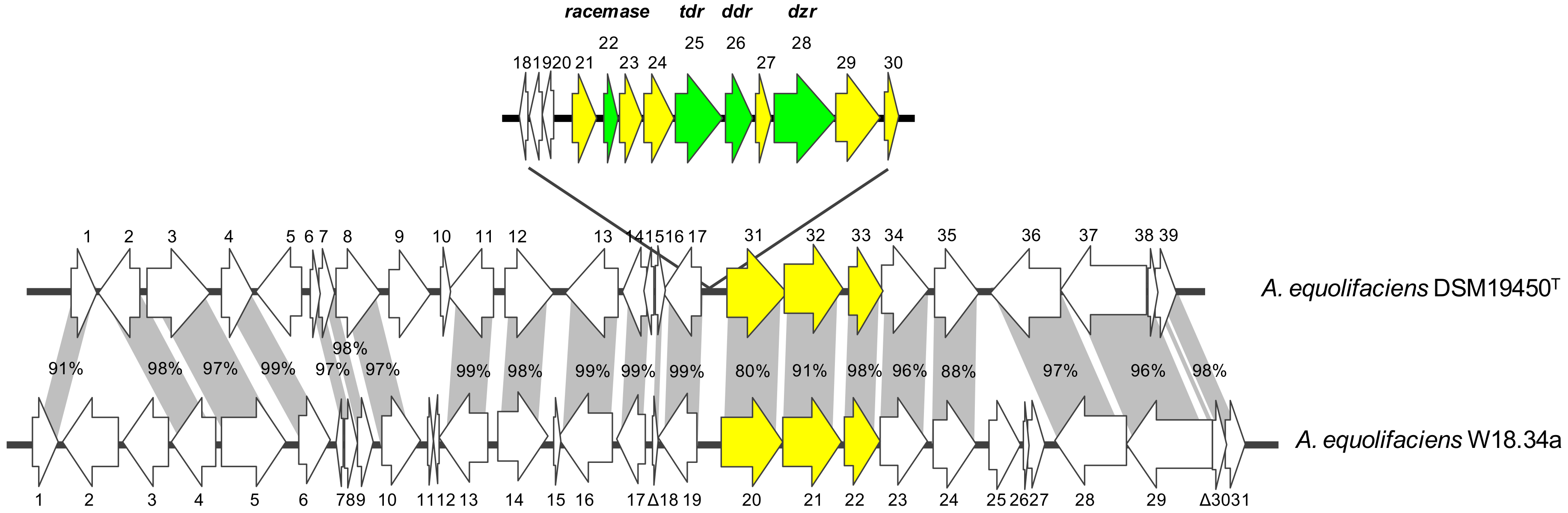
2.6. Genome Analysis of Adlercreutzia equolifaciens W18.34a
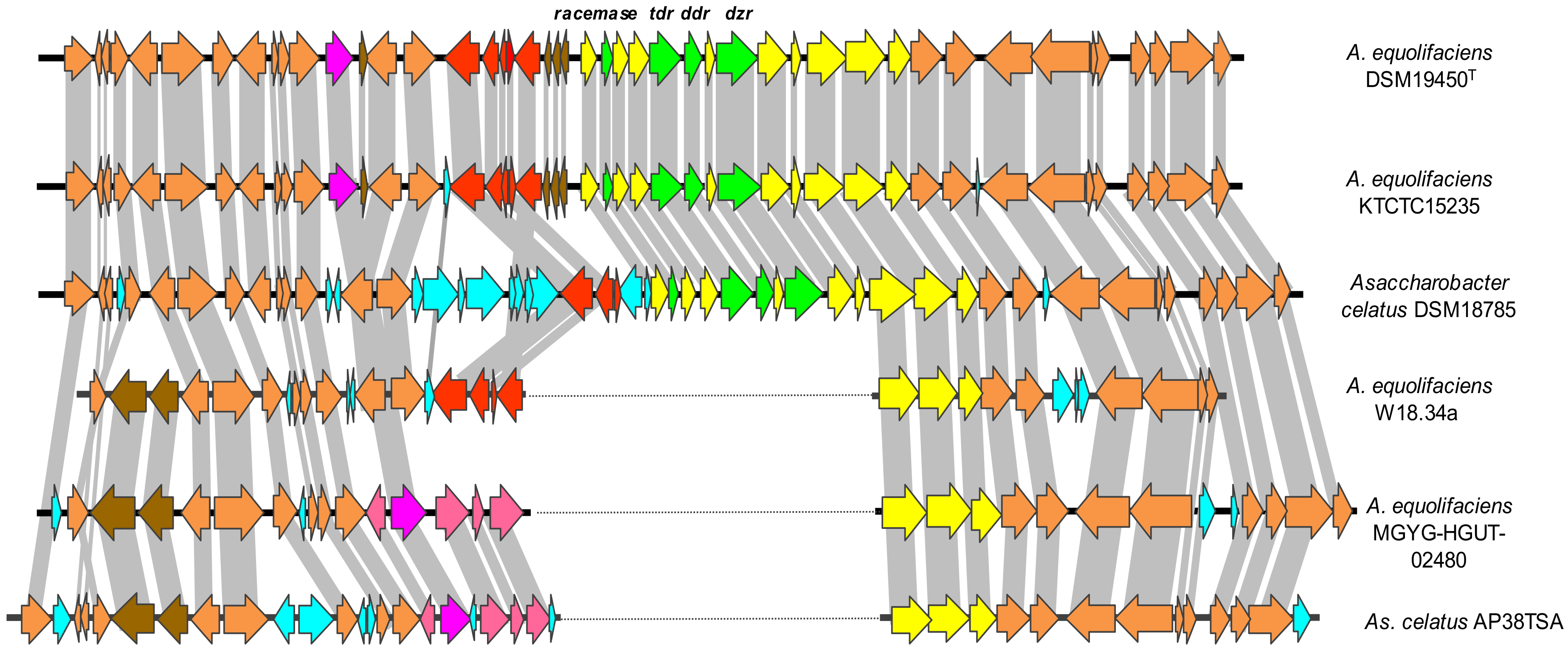
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zaheer, K.; Humayoun Akhtar, M. An updated review of dietary isoflavones: Nutrition, processing, bioavailability and impacts on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 1280–1293. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Badger, T.M.; Ronis, M.J.; Wu, X. Non-isoflavone phytochemicals in soy and their health effects. J. Agric. Food Chem. 2010, 58, 8119–8133. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Calvo, J.M.; Rodríguez-Iglesias, M.A.; Molinillo, J.M.; Macías, F.A. Soy isoflavones and their relationship with microflora: Beneficial effects on human health in equol producers. Phytochem. Rew. 2013, 12, 979–1000. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antiox. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.-P.; Wang, J.-H.; Liu, X. Metabolism of dietary soy isoflavones to equol by human intestinal microbiota. Implications for health. Mol. Nutr. Food Res. 2007, 51, 765–781. [Google Scholar] [CrossRef]
- Choi, E.J.; Kim, G.H. The antioxidant activity of daidzein metabolites, O-desmethylangolensin and equol, in HepG2 cells. Mol. Med. Rep. 2014, 9, 328–332. [Google Scholar] [CrossRef] [Green Version]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef]
- Islam, M.A.; Bekele, R.; Vanden Berg, J.H.; Kuswanti, Y.; Thapa, O.; Soltani, S.; van Leeuwen, F.X.; Rietjens, I.M.; Murk, A.J. Deconjugation of soy isoflavone glucuronides needed for estrogenic activity. Toxicol. In Vitro 2015, 29, 706–715. [Google Scholar] [CrossRef] [Green Version]
- Clavel, T.; Mapesa, J.O. Phenolics in human nutrition: Importance of the intestinal microbiome for isoflavone and lignan bioavailability. In Natural Products. Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Ramawat, K.G., Merillon, J.M., Eds.; Chapter 94; Springer: Berlin/Heidelberg, Germany, 2013; pp. 2433–2463. [Google Scholar]
- Setchell, K.D.; Cole, S.J. Method of defining equol-producer status and its frequency among vegetarians. J. Nutr. 2006, 136, 2188–2193. [Google Scholar] [CrossRef] [Green Version]
- Setchell, K.D.; Brown, N.M.; Summer, S.; King, E.C.; Heubi, J.E.; Cole, S.; Guy, T.; Hokin, B. Dietary factors influence production of the soy isoflavone metabolite S-(-)equol in healthy adults. J. Nutr. 2013, 143, 1950–1958. [Google Scholar] [CrossRef] [Green Version]
- Song, K.B.; Atkinson, C.; Frankenfeld, C.L.; Jokela, T.; Wähälä, K.; Thomas, W.K.; Lampe, J.W. Prevalence of daidzein-metabolizing phenotypes differs between Caucasian and Korean American women and girls. J. Nutr. 2006, 136, 1347–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemperman, R.A.; Bolca, S.; Roger, L.C.; Vaughan, E.E. Novel approaches for analysing gut microbes and dietary polyphenols: Challenges and opportunities. Microbiology 2010, 156, 3224–3231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, K.M.; Reed, S.D.; Uchiyama, S.; Qu, C.; Ueno, T.; Iwashita, S.; Gunderson, G.; Fuller, S.; Lampe, J.W. A cross-sectional study of equol producer status and self-reported vasomotor symptoms. Menopause 2015, 22, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.; Kendall, C.W.; Marchie, A.; Liu, Z.; Vidgen, E.; Holmes, C.; Jackson, C.J.; Josse, R.G.; Pencharz, P.B.; Rao, A.V.; et al. Equol status and blood lipid profile in hyperlipidemia after consumption of diets containing soy foods. Am. J. Clin. Nutr. 2012, 95, 564–571. [Google Scholar] [CrossRef]
- Mayo, B.; Vázquez, L.; Flórez, A.B. Equol: A bacterial metabolite from the daidzein isoflavone and its presumed beneficial health effects. Nutrients 2019, 11, 2231. [Google Scholar] [CrossRef] [Green Version]
- Salam, N.; Jiao, J.Y.; Zhang, X.T.; Li, W.J. Updat0e on the classification of higher ranks in the phylum Actinobacteria. Int. J. Syst. Evol. Microbiol. 2020, 70, 1331–1355. [Google Scholar] [CrossRef]
- Maruo, T.; Sakamoto, M.; Ito, C.; Toda, T.; Benno, Y. Adlercreutzia equolifaciens gen. nov., sp. nov., an equol-producing bacterium isolated from human faeces, and emended description of the genus Eggerthella. Int. J. Syst. Evol. Microbiol. 2008, 58, 1221–1227. [Google Scholar] [CrossRef]
- Minamida, K.; Ota, K.; Nishimukai, M.; Tanaka, M.; Abe, A.; Sone, T.; Tomita, F.; Hara, H.; Asano, K. Asaccharobacter celatus gen. nov., sp. nov., isolated from rat caecum. Int. J. Syst. Evol. Microbiol. 2008, 58, 1238–1240. [Google Scholar] [CrossRef]
- Clavel, T.; Charrier, C.; Braune, A.; Wenning, M.; Blaut, M.; Haller, D. Isolation of bacteria from the ileal mucosa of TNFdeltaARE mice and description of Enterorhabdus mucosicola gen. nov., sp. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 1805–1812. [Google Scholar] [CrossRef] [Green Version]
- Matthies, A.; Blaut, M.; Braune, A. Isolation of a human intestinal bacterium capable of daidzein and genistein conversion. Appl. Environ. Microbiol. 2009, 75, 1740–1744. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.S.; Kitahara, M.; Sakamoto, M.; Hattori, M.; Benno, Y. Slackia equolifaciens sp. nov., a human intestinal bacterium capable of producing equol. Int. J. Syst. Evol. Microbiol. 2010, 60, 1721–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schröder, C.; Matthies, A.; Engst, W.; Blaut, M.; Braune, A. Identification and expression of genes involved in the conversion of daidzein and genistein by the equol-forming bacterium Slackia isoflavoniconvertens. Appl. Environ. Microbiol. 2013, 79, 3494–3502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, H.; Moriyama, K.; Nomoto, K.; Akaza, H. Identification of an enzyme system for daidzein-to-equol conversion in Slackia sp. strain NATTS. Appl. Environ. Microbiol. 2012, 78, 1228–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, Y.; Yasuda, S.; Takahashi, M.; Hayashi, T.; Miyazawa, N.; Sato, I.; Abiru, Y.; Uchiyama, S.; Hishigaki, H. Cloning and expression of a novel NADP(H)-dependent daidzein reductase, an enzyme involved in the metabolism of daidzein, from equol-producing Lactococcus strain 20–92. Appl. Environ. Microbiol. 2010, 76, 5892–5901. [Google Scholar] [CrossRef] [Green Version]
- Shimada, Y.; Takahashi, M.; Miyazawa, N.; Ohtani, T.; Abiru, Y.; Uchiyama, S.; Hishigaki, H. Identification of two novel reductases involved in equol biosynthesis in Lactococcus strain 20–92. J. Mol. Microbiol. Biotechnol. 2011, 2, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Matthies, A.; Loh, G.; Blaut, M.; Braune, A. Daidzein and genistein are converted to equol and 5-hydroxy-equol by human intestinal Slackia isoflavoniconvertens in gnotobiotic rats. J. Nutr. 2012, 142, 40–66. [Google Scholar] [CrossRef]
- Martinez, I.; Kim, J.; Duffy, P.R.; Schlegel, V.L.; Walter, J. Resistant starches types 2 and 4 have differential effects on the composition of the fecal microbiota in human subjects. PLoS ONE 2010, 5, e15046. [Google Scholar] [CrossRef] [Green Version]
- Jumpertz, R.; Le, D.S.; Turnbaugh, P.J.; Trinidad, C.; Bogardus, C.; Gordon, J.I.; Krakoff, J. Energy-balance studies reveal associations between gut microbes, caloric load, and nutrient absorption in humans. Am. J. Clin. Nutr. 2011, 94, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, L.; Guadamuro, L.; Giganto, F.; Mayo, B.; Flórez, A.B. Development and use of a real-time quantitative PCR method for detecting and quantifying equol-producing bacteria in human faecal samples and slurry cultures. Front. Microbiol. 2017, 8, 1155. [Google Scholar] [CrossRef]
- Ruiz-Barba, J.L.; Maldonado, A.; Jiménez-Díaz, R. Small-scale total DNA extraction from bacteria and yeast for PCR applications. Anal. Biochem. 2005, 347, 333–335. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: architecture and applications. BMC Bioinform. 2008, 10, 421. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 15 February 2020). [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. Available online: https://www.rdp.cme.msu.edu/ (accessed on 1 June 2019). [CrossRef] [PubMed] [Green Version]
- Rossetti, L.; Giraffa, G. Rapid identification of dairy lactic acid bacteria byM13-generated, RAPD-PCR fingerprint databases. J. Microbiol. Methods 2005, 63, 135–144. [Google Scholar] [CrossRef]
- Koeuth, T.; Versalovic, J.; Lupski, J.R. Differential subsequence conservation of interspersed repetitive Streptococcus pneumoniae BOX elements in diverse bacteria. Genome Res. 1995, 5, 408–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mättö, J.; Marinen, E.; Suihko, M.L.; Alander, M.; Palva, A.; Saarela, M. Genetic heterogeneity and functional properties of intestinal bifidobacteria. J. Appl. Microbiol. 2004, 97, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Redruello, B.; Guadamuro, L.; Cuesta, I.; Álvarez-Buylla, J.R.; Mayo, B.; Delgado, S. A novel UHPLC method for the rapid and simultaneous determination of daidzein, genistein and equol in human urine. J. Chromatogr. B Analyt. Technol. Biomed. Life 2015, 1005, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, L.; Flórez, A.B.; Mayo, B. Draft genome sequence of Adlercreutzia equolifaciens IPLA 37004, a human intestinal strain that does not produce equol from daidzein. Microbiol. Resour. Announc. 2020, 9, e01537-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2017, 45, D535–D542. Available online: https://www.patricbrc.org (accessed on 20 April 2019). [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Cock, P.J.; Antao, T.; Chang, J.T.; Chapman, B.A.; Cox, C.J.; Dalke, A.; Friedberg, I.; Hamelryck, T.; Kauff, F.; Wilczynski, B. Biopython: Freely available Python tools for computational molecular biology and bioinformatics. Bioinformatics 2009, 25, 1422–1423. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.C.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusova, T.; Ciufo, S.; Federhen, S.; Fedorov, B.; McVeigh, R.; O’Neill, K.; Tolstoy, I.; Zaslavsky, L. Update on RefSeq microbial genomes resources. Nucleic Acids Res. 2015, 43, D599–D605. Available online: https://www.ncbi.nlm.nih.gov/genome/microbes/ (accessed on 15 July 2015). [CrossRef] [PubMed]
- Flórez, A.B.; Vázquez, L.; Rodríguez, J.; Redruello, B.; Mayo, B. Transcriptional regulation of the equol biosynthesis gene cluster in Adlercreutzia equolifaciens DSM19450T. Nutrients 2019, 11, 993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chadha, R.; Bhalla, Y.; Jain, A.; Chadha, K.; Karan, M. Dietary soy isoflavone: A mechanistic insight. Nat. Prod. Commun. 2017, 12, 627–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Gallegos, R.; Matano, L.M.; Butler, N.L.; Hantman, N.; Kaili, M.; Coyne, M.J.; Comstock, L.E.; Malamy, M.H.; Barquera, B. Genetic and biochemical analysis of anaerobic respiration in Bacteroides fragilis and its importance in vivo. mBio 2020, 11, e03238-19. [Google Scholar] [CrossRef] [Green Version]
- Decroos, K.; Vanhemmens, S.; Cattoir, S.; Boon, N.; Verstraete, W. Isolation and characterisation of an equol-producing mixed microbial culture from a human faecal sample and its activity under gastrointestinal conditions. Arch. Microbiol. 2005, 183, 45–55. [Google Scholar] [CrossRef]
- Wang, X.L.; Kim, K.T.; Lee, J.H.; Hur, H.G.; Kim, S.I. C-ring cleavage of isoflavones daidzein and genistein by a newly-isolated human intestinal bacterium Eubacterium ramulus Julong 601. J. Microbiol. Biotech. 2004, 14, 766–771. [Google Scholar]
- Obara, A.; Kinoshita, M.; Hosoda, K.; Yokokawa, A.; Shibasaki, H.; Ishii, K. Identification of equol-7-glucuronide-4’-sulfate, monoglucuronides and monosulfates in human plasma of 2 equol producers after administration of kinako by LC-ESI-MS. Pharmacol. Res. Perspect. 2019, 7, e00478. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.G.; Kim, J.; Kim, E.J.; Lee, S.H.; Choi, K.Y.; Kazlauskas, R.J.; Kim, B.G. Biosynthesis of (-)-5-hydroxy-equol and 5-hydroxy-dehydroequol from soy isoflavone genistein using microbial whole cell bioconversion. ACS Chem. Biol. 2017, 12, 2883–28890. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M.; Arqués, J.; Medina, M.; Gaya, P.; de Las Rivas, B.; Muñoz, R. Bioactivation of phytoestrogens: Intestinal bacteria and health. Crit. Rev. Food Sci. Nutr. 2016, 56, 1826–1843. [Google Scholar] [CrossRef] [PubMed]
- Franke, A.A.; Lai, J.F.; Halm, B.M. Absortion, distribution, metabolism, and excretion of isoflavonoids after soy intake. Arch. Biochem. Biophys. 2014, 59, 24–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tho, H.; Oshima, K.; Suzuki, T.; Hattori, M.; Morita, H. Complete genome sequence of the equol-producing bacterium Adlercreutzia equolifaciens DSM 19450. Genome Announc. 2013, 1, e00742. [Google Scholar]
- Shimada, Y.; Takahashi, M.; Miyazawa, N.; Abiru, Y.; Uchiyama, S.; Hishigaki, H. Identification of a novel dihydrodaidzein racemase essential for biosynthesis of equol from daidzein in Lactococcus sp. strain 20–92. Appl. Environ. Microbiol. 2012, 78, 4902–4907. [Google Scholar] [CrossRef] [Green Version]
- Thawornkuno, C.; Tanaka, M.; Sone, T.; Asano, K. Biotransformation of daidzein to equol by crude enzyme from Asaccharobacter celatus AHU1763 required an anaerobic environment. Biosci. Biotechnol. Biochem. 2009, 73, 1435–1438. [Google Scholar] [CrossRef]
- Bolca, S.; Possemiers, S.; Herregat, A.; Huybrechts, I.; Heyerick, A.; De Vriese, S.; Verbruggen, M.; Depypere, H.; De Keukeleire, D.; Bracke, M.; et al. Microbial and dietary factors are associated with the equol producer phenotype in healthy postmenopausal women. J. Nutr. 2007, 137, 2242–2246. [Google Scholar] [CrossRef] [Green Version]
- Schwen, R.J.; Nguyen, L.; Charles, R.L. Elucidation of the metabolic pathway of S-equol in rat, monkey and man. Food Chem. Toxicol. 2012, 50, 2074–2083. [Google Scholar] [CrossRef]
- Nakatsu, C.H.; Arsmstrong, A.; Cavijo, A.P.; Martin, B.R.; Barnes, S.; Weaver, C.M. Fecal bacterial community changes associated with isoflavone metabolites in postmenopausal women after soy bar consumption. PLoS ONE 2014, 9, e108924. [Google Scholar] [CrossRef]
- Sugiyama, Y.; Nagata, Y.; Fukuta, F.; Takayanagi, A.; Masumori, N.; Tsukamoto, T.; Akasaka, H.; Ohnishi, H.; Saito, S.; Miura, T.; et al. Counts of Slackia sp. strain NATTS in intestinal flora are correlated to serum concentrations of equol both in prostate cancer cases and controls in Japanese men. Asian Pac. J. Cancer Prev. 2014, 15, 2693–2697. [Google Scholar] [CrossRef] [Green Version]
- Cho, G.-S.; Ritzmann, F.; Eckstein, M.; Huch, M.; Briviba, K.; Behsnilian, D.; Neve, H.; Franz, C.M.A.P. Quantification of Slackia and Eggerthella spp. in human feces and adhesion of representatives strains to Caco-2 Cells. Front. Microbiol. 2016, 7, 658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guadamuro, L.; Delgado, S.; Redruello, B.; Flórez, A.B.; Suárez, A.; Martínez-Camblor, P.; Mayo, B. Equol status and changes in fecal microbiota in menopausal women receiving long-term treatment for menopause symptoms with a soy-isoflavone concentrate. Front. Microbiol. 2015, 6, 777. [Google Scholar] [CrossRef] [PubMed]
- Braune, A.; Blaut, M. Evaluation of inter-individual differences in gut bacterial isoflavone bioactivation in humans by PCR-based targeting of genes involved in equol formation. J. Appl. Microbiol. 2018, 124, 220–231. [Google Scholar] [CrossRef] [PubMed]

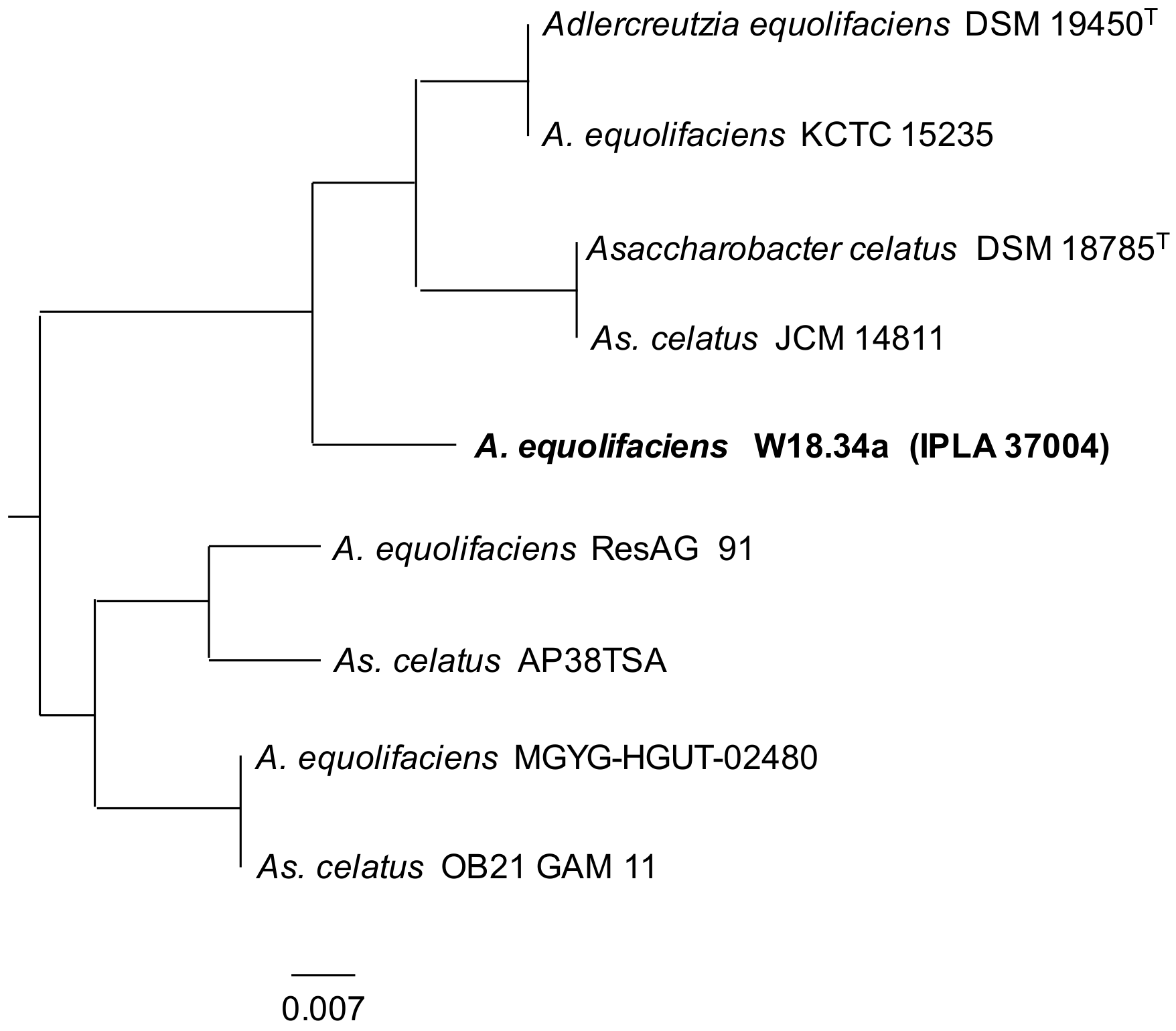
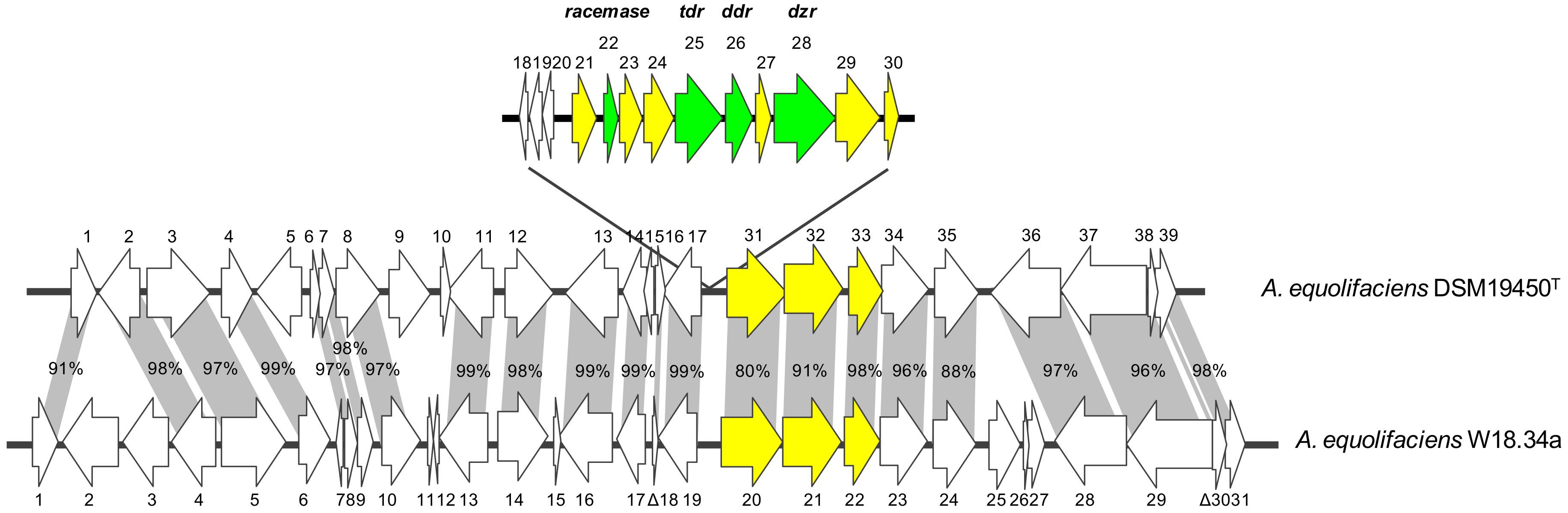
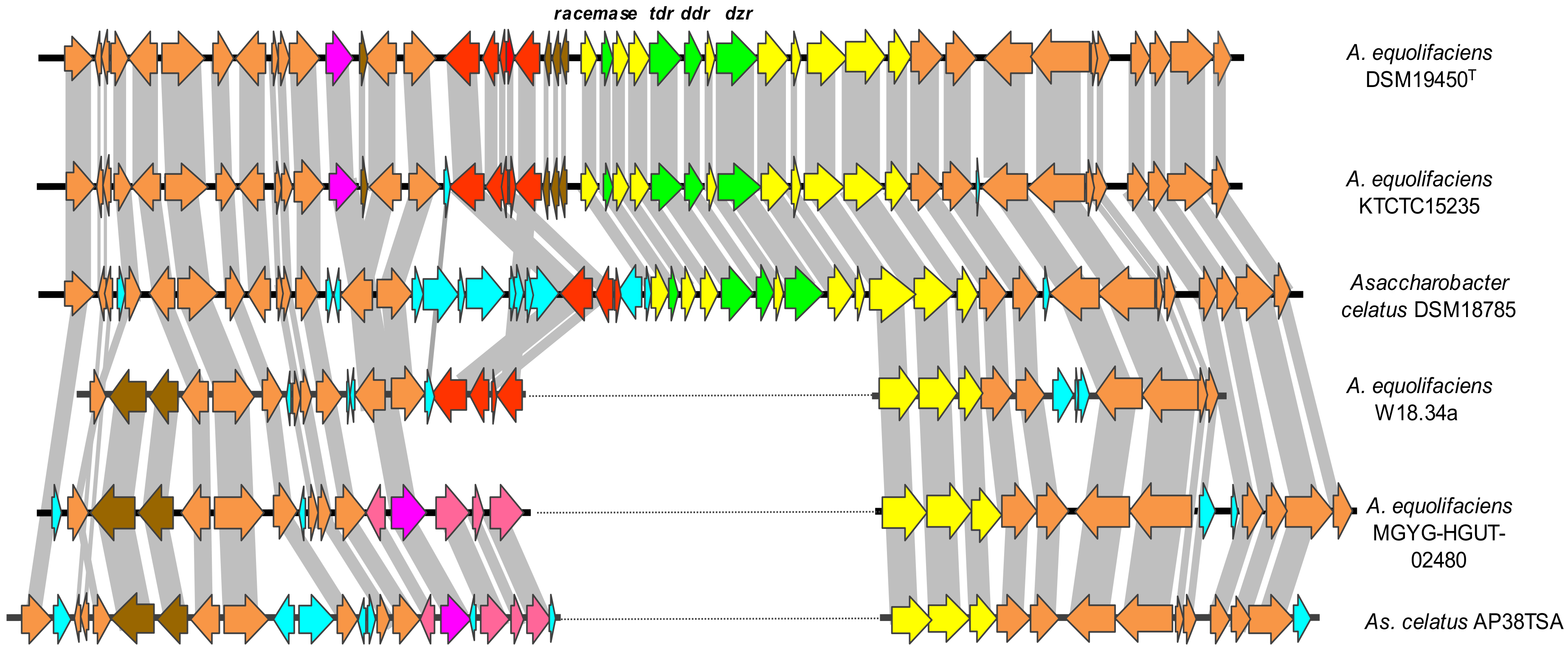

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Class | Family | Species | Isolate a | % Identity | Homology to Sequence |
|---|---|---|---|---|---|---|
| Proteobacteria | γ-Proteobacteria | Enterobacteriaceae | Escherichia coli | W3.16 | 98.76 | DQ857002.1 |
| E. coli | W3.21 | 99.00 | CP026491.1 | |||
| E. coli | W8.4b | 100.00 | KR265473.1 | |||
| E. coli | W8.5 | 99.00 | MF067516.1 | |||
| E. coli | W8.22b | 99.00 | CP019455.1 | |||
| E. coli | W8.24b | 99.00 | CP026491.1 | |||
| E. coli | W8.35b | 99.00 | KR265473.1 | |||
| E. coli | W8.36b | 99.31 | MH782081.1 | |||
| E. coli | W8.39a | 99.68 | KY524291.1 | |||
| E. coli | W8.40 | 99.79 | KY524291.1 | |||
| E. coli | W8.47 | 99.00 | CP019455.1 | |||
| E. coli | W8.52b | 99.00 | KR265473.1 | |||
| E. coli | W8.53b | 99.00 | KP789326.1 | |||
| E. coli | W8.56c | 98.41 | KP244257.1 | |||
| E. coli | W18.17b | 99.88 | MH111527.1 | |||
| E. coli | W18.34b | 99.00 | CP019455.1 | |||
| E. coli | W18.19b | 99.00 | LC270191.1 | |||
| Firmicutes | Bacilli | Enterococcaceae | Enterococcus durans | W8.29 | 99.21 | KY962884.1 |
| Enterococcus lactis | W8.56b | 99.68 | MH431810.1 | |||
| Clostridia | Anaerococcus spp. | W3.10b | 96.69 | NR_114314.1 | ||
| Anaerococcus spp. | W8.44a | 95.15 | NR_114314.1 | |||
| Anaerococcus vaginalis | W3.18 | 98.43 | NR_114314.1 | |||
| A. vaginalis | W8.44b | 98.33 | NR_041937.1 | |||
| Ruminococcaceae | Bittarella massiliensis | W18.24a | 99.46 | LN881596.1 | ||
| B. massiliensis | W18.24b | 99.55 | LN881596.1 | |||
| Finegoldia magna | W8.4a | 99.80 | KC311751.1 | |||
| F. magna | W8.35a | 99.00 | KR232883.1 | |||
| Peptoniphilus gorbachii | W8.22a | 99.00 | NR_115885.1 | |||
| P. gorbachii | W8.33 | 99.00 | NR_115885.1 | |||
| Streptococcaceae | Streptococcus anginosus | W8.21 | 99.00 | JN787165.1 | ||
| Bacteroidetes | Bacteroidia | Bacteroidaceae | Bacteroides vulgatus | W8.28 | 99.00 | CP013020.1 |
| Porphyromonadaceae | Parabacteroides spp. | W8.41 | 95.56 | CP022754.1 | ||
| Actinobacteria | Actinomycetales | Actinomycetaceae | Actinomyces neuii | W8.24a | 99.56 | NR_042429.1 |
| Bifidobacteriales | Bifidobacteriaceae | Bifidobacterium adolescentis | W8.34 | 99.30 | KC174855.1 | |
| B. adolescentis | W8.36a | 99.00 | KC174855.1 | |||
| B. adolescentis | W8.39b | 99.00 | LT223639.1 | |||
| B. adolescentis | W8.45 | 99.00 | LT223639.1 | |||
| B. adolescentis | W8.54b | 96.31 | HM009032.1 | |||
| Bifidobacterium animalis | W3.3 | 99.31 | MK561779.1 | |||
| Bifidobacterium longum | W8.17 | 99.00 | KC174855.1 | |||
| B. longum | W8.49a | 98.00 | HM009032.1 | |||
| Coriobacteriia | Coriobacteriaceae | Collinsella aerofaciens | W8.23 | 99.00 | KP233323.1 | |
| C. aerofaciens | W8.49b | 99.00 | KR232866.1 | |||
| Collinsella massiliensis | W18.17a | 100.00 | NR_144579.1 | |||
| C. massiliensis | W18.9 | 99.00 | NR_144579.1 | |||
| C. massiliensis | W18.21a | 99.00 | NR_144579.1 | |||
| C. massiliensis | W18.21c | 99.00 | NR_144579.1 | |||
| C. massiliensis | W18.21d | 99.00 | NR_144579.1 | |||
| C. massiliensis | W18.25 | 99.00 | NR_144579.1 | |||
| C. massiliensis | W18.29 | 99.00 | NR_144579.1 | |||
| Collinsella stercoris | W18.1 | 97.80 | KP233278.1 | |||
| Eggerthellaceae | Adlercreutzia equolifaciens | W18.34a | 100.00 | AB693938.1 | ||
| Gordonibacter urolithinfaciens | W18.23 | 99.00 | NR_148261.1 | |||
| G. urolithinfaciens | W18.26 | 100.00 | LT900217.1 | |||
| Eggerthella lenta | W3.2 | 99.00 | KX683977.1 | |||
| E. lenta | W3.10a | 99.00 | KX683993.1 | |||
| E. lenta | W3.11 | 99.00 | KX683977.1 | |||
| E. lenta | W3.12 | 99.00 | KX683977.1 | |||
| E. lenta | W3.13 | 100.00 | KX683977.1 | |||
| E. lenta | W3.14 | 99.00 | KX683977.1 | |||
| E. lenta | W3.15 | 100.00 | KX683977.1 | |||
| E. lenta | W8.15 | 98.00 | KX683885.1 | |||
| E. lenta | W8.16 | 99.00 | KX683992.1 | |||
| E. lenta | W8.27 | 99.00 | KP944189.1 | |||
| E. lenta | W8.31 | 99.00 | KP944189.1 | |||
| E. lenta | W8.42 | 99.00 | JX104026.1 | |||
| E. lenta | W8.43 | 99.00 | KX683992.1 | |||
| E. lenta | W8.52a | 99.00 | KP944190.1 | |||
| E. lenta | W8.53a | 99.00 | KX683992.1 | |||
| E. lenta | W8.54a | 99.00 | KP944189.1 | |||
| E. lenta | W8.56a | 98.00 | JX104026.1 | |||
| E. lenta | W18.18 | 99.00 | KX683993.1 | |||
| E. lenta | W18.21b | 100.00 | KX683993.1 | |||
| Equol-Producing Strain | Daidzein Supplementation (µM) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 12.5 | 25 | 50 | 100 | |||||||||||||
| Daidzein-Derived Metabolite (µM) | ||||||||||||||||
| Daidzein | Equol | Daidzein | Equol | Daidzein | Equol | Daidzein | Equol | |||||||||
| 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | |
| A. equolifaciens DSM19450T | - | - | 16.6 | 7.7 | - | - | 33.6 | 13.5 | - | - | 60.9 | 25.8 | 2.3 * | - | 126.2 | 53.4 |
| As. celatus DSM18785T | - | - | 3.1 | 3.3 | nd | - | nd | 6.3 | - | - | 14.6 | 12.0 | - | - | 27.7 | 27.8 |
| E. mucosicola DSM19490T | 2.1 * | 1.4 * | 4.5 | 2.0 * | - | - | 17.9 | 6.7 | 5.1 * | 1.4 * | 26.2 | 10.5 | 60.3 | 36.2 | nd | nd |
| S. equolifaciens DSM24851T | - | - | 5.8 | 7.4 | - | - | 14.4 | 13.3 | 1.1 * | - | 39.2 | 23.3 | 1.8 * | - | 110.0 | 47.8 |
| S. isoflavoniconvertens DSM22006T | - | - | 18.2 | 7.9 | - | - | 37.0 | 14.3 | - | - | 67.6 | 24.6 | - | - | 149.0 | 55.0 |
| Isolate | DZEN a (100 μM) | GTEN (100 μM) | |||||
|---|---|---|---|---|---|---|---|
| DZEN | D-DZEN | O-DMA | EQUOL | GTEN | D-GTEN | ||
| Escherichia coli | W3.16 | 61.2 | - | - | - | 64.9 | 4.8 * |
| E. coli | W3.21 | 61.9 | - | - | - | 60.0 | 4.3 * |
| E. coli | W8.4b | 65.8 | - | - | - | 60.6 | 4.2 * |
| E. coli | W8.24b | 85.1 | - | 3.3 * | - | 65.9 | 4.2 * |
| E. coli | W8.35b | 76.3 | - | 2.8 * | - | 60.4 | 5.5 * |
| E. coli | W8.36b | 73.2 | 1.5 * | 6.8 | - | 68.6 | - |
| E. coli | W8.40 | 73.2 | 3.0 * | 3.0 * | - | 66.6 | - |
| E. coli | W8.47 | 83.8 | - | 5.5 * | - | 62.8 | - |
| E. coli | W8.53b | 82.5 | - | 10.3 | - | 72.1 | - |
| E. coli | W18.17b | 87.5 | - | 11.2 | - | 72.0 | - |
| Enterococcus durans | W8.29 | 85.1 | - | 3.7 * | - | 117.9 | - |
| Enterococcus lactis | W8.56b | 87.0 | - | 10.8 | - | 71.7 | - |
| Anaerococcus spp. | W3.10b | 93.6 | - | 10.9 | - | 75.5 | - |
| Anaerococcus spp. | W8.44a | 82.1 | 3.4 | 2.9 * | - | 62.9 | - |
| Anaerococcus vaginalis | W3.18 | 62.0 | - | 2.3 * | - | 78.9 | - |
| Anaerococcus vaginalis | W8.44b | 79.8 | - | - | - | 48.2 | - |
| Bittarella massiliensis | W18.24a | 84.5 | - | 10.2 | - | 78.2 | - |
| Finegoldia magna | W8.4a | 61.2 | - | - | - | 55.7 | 2.7 * |
| F. magna | W8.35a | 68.0 | 1.1 * | 5.2 * | - | 62.6 | - |
| Peptoniphilus gorbachii | W8.22a | 69.5 | 3.9 | 10.0 | - | 40.3 | 4.3 * |
| P. gorbachii | W8.33 | 59.6 | 2.9 * | 2.5 * | - | 40.3 | - |
| Streptococcus anginosus | W8.21 | 99.6 | - | 2.9 * | - | 53.7 | - |
| Bacteroides vulgatus | W8.28 | 73.1 | - | 3.7 * | - | 92.3 | 6.3 |
| Parabacteroides spp. | W8.41 | 72.3 | - | - | - | 68.6 | - |
| Actinomyces neuii | W8.24a | 53.5 | 2.8 * | 5.1 * | - | 83.2 | 7.7 |
| Bifidobacterium adolescentis | W8.34 | 96.4 | - | - | - | 79.8 | 16.0 |
| B. adolescentis | W8.36a | 110.5 | - | 3.4 * | - | 109.8 | 8.3 |
| B. adolescentis | W8.45 | 86.6 | - | 5.3 | - | 123.1 | 10.5 |
| Bifidobacterium animalis | W3.3 | 85.2 | - | 10.4 | - | 88.8 | - |
| Bifidobacterium longum | W8.17 | 62.2 | 3.8 | 10.3 | - | 82.2 | 3.9 * |
| B. longum | W8.49a | 136.1 | - | 2.3 * | - | 124.4 | - |
| Collinsella aerofaciens | W8.23 | 109.1 | 3.7 | 11.7 | - | 84.6 | 6.1 * |
| C. aerofaciens | W8.49b | 70.9 | - | - | - | 63.2 | - |
| Collinsella massiliensis | W18.9 | 62.3 | - | 12.0 | - | 52.3 | - |
| C. massiliensis | W18.17a | 85.7 | - | 11.2 | - | 69.5 | - |
| C. massiliensis | W18.21a | 91.8 | - | 8.3 | - | 81.7 | - |
| C. massiliensis | W18.21c | 89.2 | - | 8.8 | - | 70.9 | - |
| C. massiliensis | W18.21d | 85.5 | - | 10.4 | - | 79.9 | - |
| C. massiliensis | W18.25 | 84.6 | - | 12.0 | - | 76.5 | - |
| C. massiliensis | W18.29 | 78.4 | - | - | - | 93.6 | - |
| Collinsella stercoris | W18.1 | 87.8 | - | 9.4 | - | 81.2 | - |
| Adlercreutzia equolifaciens | W18.34a | 87.4 | - | 11.6 | - | 71.5 | - |
| Gordonibacter urolithinfaciens | W18.23 | 76.9 | - | 10.8 | - | 73.3 | - |
| Eggerthella lenta | W3.2 | 90.7 | - | 12.5 | - | 95.1 | - |
| E. lenta | W3.10a | 86.4 | - | 11.3 | - | 78.0 | - |
| E. lenta | W3.11 | 89.1 | - | 11.6 | - | 80.4 | - |
| E. lenta | W3.12 | 86.4 | - | 8.7 | - | 79.1 | - |
| E. lenta | W3.13 | 88.3 | - | 10.7 | - | 81.9 | - |
| E. lenta | W3.14 | 89.7 | - | 11.6 | - | 79.3 | - |
| E. lenta | W8.15 | 66.3 | 3.9 | 9.8 | - | 58.6 | 6.4 |
| E. lenta | W8.16 | 65.7 | 3.3 | 8.7 | - | 58.4 | 3.4 * |
| E. lenta | W8.27 | 94.3 | - | 3.7 * | - | 82.1 | 4.4 * |
| E. lenta | W8.31 | 117.2 | - | - | - | 90.0 | 6.8 |
| E. lenta | W8.42 | 78.1 | 3.5 | 2.9 * | - | 62.2 | - |
| E. lenta | W8.52a | 97.9 | - | 8.9 | - | 63.5 | - |
| E. lenta | W8.53a | 87.3 | - | 9.0 | - | 73.0 | - |
| E. lenta | W18.18 | 74.1 | - | 12.3 | - | 66.2 | - |
| Uninoculated medium | - | 98.1 ± 5.8 | 1.4 ± 0.2 | - | - | 95.6 ± 0.4 | 1.6 ± 0.6 |
| S. isoflavoniconvertens | DSM 22006T | - | 1.6 ± 0.2 | - | 164.3 ± 13.4 | 55.3 | 19.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez, L.; Flórez, A.B.; Redruello, B.; Mayo, B. Metabolism of Soy Isoflavones by Intestinal Bacteria: Genome Analysis of an Adlercreutzia equolifaciens Strain That Does Not Produce Equol. Biomolecules 2020, 10, 950. https://doi.org/10.3390/biom10060950
Vázquez L, Flórez AB, Redruello B, Mayo B. Metabolism of Soy Isoflavones by Intestinal Bacteria: Genome Analysis of an Adlercreutzia equolifaciens Strain That Does Not Produce Equol. Biomolecules. 2020; 10(6):950. https://doi.org/10.3390/biom10060950
Chicago/Turabian StyleVázquez, Lucía, Ana Belén Flórez, Begoña Redruello, and Baltasar Mayo. 2020. "Metabolism of Soy Isoflavones by Intestinal Bacteria: Genome Analysis of an Adlercreutzia equolifaciens Strain That Does Not Produce Equol" Biomolecules 10, no. 6: 950. https://doi.org/10.3390/biom10060950
APA StyleVázquez, L., Flórez, A. B., Redruello, B., & Mayo, B. (2020). Metabolism of Soy Isoflavones by Intestinal Bacteria: Genome Analysis of an Adlercreutzia equolifaciens Strain That Does Not Produce Equol. Biomolecules, 10(6), 950. https://doi.org/10.3390/biom10060950