Agricultural Use of Copper and Its Link to Alzheimer’s Disease

 ,
,  , ,
, , 
Abstract
1. Alzheimer’s Disease
2. Alzheimer’s Disease and Copper
2.1. Copper Connection with Alzheimer’s Disease: Biochemistry Literature
2.2. Copper Connection with Alzheimer’s Disease: Medicine Literature
2.3. Copper Connection with Alzheimer’s Disease: Neuroscience
3. Use of Copper in Agriculture
3.1. Copper Used in Plant Disease Management
3.2. Copper Accumulation in Soil and Water
4. Agricultural Use to Copper and Its Link to Alzheimer’s Disease
5. Human Health Risk Assessment Related to Copper Use in Agriculture
6. Agroecological Strategies to Reduce the Use of Copper in Agriculture
7. Conclusions
Supplementary Materials
Supplementary File 1Funding
Conflicts of Interest
References
- WHO | Disease Burden and Mortality Estimates. Available online: http://origin.who.int/healthinfo/global_burden_disease/estimates/en/ (accessed on 6 May 2020).
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef]
- Masters, C.L.; Selkoe, D.J. Biochemistry of amyloid beta-protein and amyloid deposits in Alzheimer disease. Cold Spring Harb. Perspect. Med. 2012, 2, a006262. [Google Scholar] [CrossRef] [PubMed]
- Goedert, M.; Spillantini, M.G. A century of Alzheimer’s disease. Science 2006, 314, 777–781. [Google Scholar] [CrossRef]
- Alzheimer’s Association, Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2019, 15, 321–387.
- Schneider, L.S.; Mangialasche, F.; Andreasen, N.; Feldman, H.; Giacobini, E.; Jones, R.; Mantua, V.; Mecocci, P.; Pani, L.; Winblad, B.; et al. Clinical trials and late-stage drug development for Alzheimer’s disease: An appraisal from 1984 to 2014. J. Intern. Med. 2014, 275, 251–283. [Google Scholar] [CrossRef] [PubMed]
- Exposure, Exposure, Exposure? At CTAD, Aducanumab Scientists Make a Case | ALZFORUM. Available online: https://www.alzforum.org/news/conference-coverage/exposure-exposure-exposure-ctad-aducanumab-scientists-make-case (accessed on 4 May 2020).
- Zegers, I.; Beetham, R.; Keller, T.; Sheldon, J.; Bullock, D.; MacKenzie, F.; Trapmann, S.; Emons, H.; Schimmel, H. The importance of commutability of reference materials used as calibrators: The example of ceruloplasmin. Clin. Chem. 2013, 59, 1322–1329. [Google Scholar] [CrossRef]
- Vos, S.J.B.; van Boxtel, M.P.J.; Schiepers, O.J.G.; Deckers, K.; Vugt, M.; Carriere, I.; Dartigues, J.F.; Peres, K.; Artero, S.; Ritchie, K.; et al. Modifiable Risk Factors for Prevention of Dementia in Midlife, Late Life and the Oldest-Old: Validation of the LIBRA Index. J. Alzheimer’s Dis. 2017, 58, 537–547. [Google Scholar] [CrossRef]
- Schiepers, O.J.G.; Kohler, S.; Deckers, K.; Irving, K.; O’Donnell, C.A.; van den Akker, M.; Verhey, F.R.J.; Vos, S.J.B.; Vugt, M.E.; van Boxtel, M.P.J. Lifestyle for Brain Health (LIBRA): A new model for dementia prevention. Int. J. Geriatr. Psychiatry 2018, 33, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Deckers, K.; Barbera, M.; Kohler, S.; Ngandu, T.; van Boxtel, M.; Rusanen, M.; Laatikainen, T.; Verhey, F.; Soininen, H.; Kivipelto, M.; et al. Long-term dementia risk prediction by the LIBRA score: A 30-year follow-up of the CAIDE study. Int. J. Geriatr. Psychiatry 2020, 35, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Barnard, N.D.; Bush, A.I.; Ceccarelli, A.; Cooper, J.; Jager, C.A.; Erickson, K.I.; Fraser, G.; Kesler, S.; Levin, S.; Lucey, M.B.; et al. Dietary and lifestyle guidelines for the prevention of Alzheimer’s disease. Neurobiol. Aging 2014, 35, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, A.; Ngandu, T.; Rusanen, M.; Antikainen, R.; Backman, L.; Havulinna, S.; Hanninen, T.; Laatikainen, T.; Lehtisalo, J.; Levalahti, E. Multidomain lifestyle intervention benefits a large elderly population at risk for cognitive decline and dementia regardless of baseline characteristics: The FINGER trial. Alzheimer’s Dement. 2018, 14, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Licher, S.; Ahmad, S.; Karamujic-Comic, H.; Voortman, T.; Leening, M.J.G.; Ikram, M.A.; Ikram, K. Genetic predisposition, modifiable-risk-factor profile and long-term dementia risk in the general population. Nat. Med. 2019, 25, 1364–1369. [Google Scholar] [CrossRef] [PubMed]
- Multhaup, G.; Schlicksupp, A.; Hesse, L.; Beher, D.; Ruppert, T.; Masters, C.L.; Beyreuther, K. The amyloid precursor protein of Alzheimer’s disease in the reduction of copper (II) to copper (I). Science 1996, 271, 1406–1409. [Google Scholar] [CrossRef] [PubMed]
- Kepp, K.P.; Squitti, R. Copper imbalance in Alzheimer’s disease: Convergence of the chemistry and the clinic. Coord. Chem. Rev. 2019, 397, 168–187. [Google Scholar] [CrossRef]
- Squitti, R.; Malosio, M.L.; Rongioletti, M.C.A.; Tecchio, F. Copper involvement in glutamatergic transmission in physiology and disease as revealed by magnetoencephalography/electroencephalography (MEG/EEG) studies. Aging Clin. Exp. Res. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Sensi, S.L.; Granzotto, A.; Siotto, M.; Squitti, R. Copper and Zinc Dysregulation in Alzheimer’s Disease. Trends Pharmacol. Sci. 2018, 39, 1049–1063. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, K.M.; Hung, Y.H.; Dalziel, A.H.; Li, Q.X.; Laughton, K.; Wikhe, K.; Rembach, A.; Roberts, B.; Masters, C.L.; Bush, A.I.; et al. Copper promotes the trafficking of the amyloid precursor protein. J. Biol. Chem. 2011, 286, 8252–8262. [Google Scholar] [CrossRef]
- Atwood, C.S.; Scarpa, R.C.; Huang, X.; Moir, R.D.; Jones, W.D.; Fairlie, D.P.; Tanzi, R.E.; Ashley, I. Bush Characterization of copper interactions with alzheimer amyloid beta peptides: Identification of an attomolar-affinity copper binding site on amyloid beta1-42. J. Neurochem. 2000, 75, 1219–1233. [Google Scholar] [CrossRef]
- Huang, X.; Atwood, C.S.; Hartshorn, M.A.; Multhaup, G.; Goldstein, L.E.; Scarpa, R.C.; Cuajungco, M.P.; Gray, D.N.; Lim, J.; Moir, R.D.; et al. The A beta peptide of Alzheimer’s disease directly produces hydrogen peroxide through metal ion reduction. Biochemistry 1999, 38, 7609–7616. [Google Scholar] [CrossRef]
- Huang, X.; Cuajungco, M.P.; Atwood, C.S.; Hartshorn, M.A.; Tyndall, J.D.A.; Hanson, G.R.; Stokes, K.C.; Leopold, M.; Multhaup, G.; Goldstein, L.E.; et al. Cu(II) potentiation of Alzheimer abeta neurotoxicity. Correlation with cell-free hydrogen peroxide production and metal reduction. J. Biol. Chem. 1999, 274, 37111–37116. [Google Scholar] [CrossRef]
- Palmer, C.; Menzies, S.L.; Roberts, R.L.; Pavlick, G.; Connor, J.R. Changes in iron histochemistry after hypoxic-ischemic brain injury in the neonatal rat. J. Neurosci. Res. 1999, 56, 60–71. [Google Scholar] [CrossRef]
- Cherny, R.A.; Legg, J.T.; McLean, C.A.; Fairlie, D.P.; Huang, X.; Atwood, C.S.; Beyreuther, K.; Tanzi, R.E.; Masters, C.L.; Bush, A.I. Aqueous dissolution of Alzheimer’s disease Abeta amyloid deposits by biometal depletion. J. Biol. Chem. 1999, 274, 23223–23228. [Google Scholar] [CrossRef] [PubMed]
- Cherny, R.A.; Atwood, C.S.; Xilinas, M.E.; Gray, D.N.; Jones, W.D.; McLean, C.A.; Barnham, K.J.; Volitakis, I.; Fraser, F.W.; Kim, Y.; et al. Treatment with a copper-zinc chelator markedly and rapidly inhibits beta-amyloid accumulation in Alzheimer’s disease transgenic mice. Neuron 2001, 30, 665–676. [Google Scholar] [CrossRef]
- Atwood, C.S.; Moir, R.D.; Huang, X.; Scarpa, R.C.; Bacarra, N.M.; Romano, D.M.; Hartshorn, M.A.; Tanzi, R.E.; Bush, A.I. Dramatic aggregation of Alzheimer abeta by Cu(II) is induced by conditions representing physiological acidosis. J. Biol. Chem. 1998, 273, 12817–12826. [Google Scholar] [CrossRef] [PubMed]
- Barnham, K.J.; Bush, A.I. Metals in Alzheimer’s and Parkinson’s diseases. Curr. Opin. Chem. Biol. 2008, 12, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Bucossi, S.; Ventriglia, M.; Panetta, V.; Salustri, C.; Pasqualetti, P.; Mariani, S.; Siotto, M.; Rossini, P.M.; Squitti, R. Copper in Alzheimer’s disease: A meta-analysis of serum, plasma, and cerebrospinal fluid studies. J. Alzheimer’s Dis. 2011, 24, 175–185. [Google Scholar] [CrossRef]
- Squitti, R.; Simonelli, I.; Ventriglia, M.; Siotto, M.; Pasqualetti, P.; Rembach, A.; Doecke, J.; Bush, A.I. Meta-analysis of serum non-ceruloplasmin copper in Alzheimer’s disease. J. Alzheimer’s Dis. 2014, 38, 809–822. [Google Scholar] [CrossRef]
- Li, D.D.; Zhang, W.; Wang, Z.Y.; Zhao, P. Serum Copper, Zinc, and Iron Levels in Patients with Alzheimer’s Disease: A Meta-Analysis of Case-Control Studies. Front. Aging Neurosci. 2017, 9, 300. [Google Scholar] [CrossRef] [PubMed]
- Schrag, M.; Mueller, C.; Oyoyo, U.; Smith, M.A.; Kirsch, W.M. Iron, zinc and copper in the Alzheimer’s disease brain: A quantitative meta-analysis. Some insight on the influence of citation bias on scientific opinion. Prog. Neurobiol. 2011, 94, 296–306. [Google Scholar] [CrossRef]
- Ventriglia, M.; Bucossi, S.; Panetta, V.; Squitti, R. Copper in Alzheimer’s disease: A meta-analysis of serum, plasma, and cerebrospinal fluid studies. J. Alzheimer’s Dis. 2012, 30, 981–984. [Google Scholar] [CrossRef]
- Wang, Z.X.; Tan, L.; Wang, H.F.; Ma, J.; Liu, J.; Tan, M.S.; Sun, J.H.; Zhu, X.C.; Jiang, T.; Yu, J.T. Serum Iron, Zinc, and Copper Levels in Patients with Alzheimer’s Disease: A Replication Study and Meta-Analyses. J. Alzheimer’s Dis. 2015, 47, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Squitti, R.; Ghidoni, R.; Simonelli, I.; Ivanova, I.D.; Colabufo, N.A.; Zuin, M.; Benussi, L.; Binetti, G.; Cassetta, E.; Rongioletti, M.; et al. Copper dyshomeostasis in Wilson disease and Alzheimer’s disease as shown by serum and urine copper indicators. J. Trace Elem. Med. Biol. 2018, 45, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Squitti, R.; Lupoi, D.; Pasqualetti, P.; Dal Forno, G.; Vernieri, F.; Chiovenda, P.; Rossi, L.; Cortesi, M.; Cassetta, E.; Rossini, P.M. Elevation of serum copper levels in Alzheimer’s disease. Neurology 2002, 59, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Squitti, R.; Fostinelli, S.; Siotto, M.; Ferrari, C.; Binetti, G.; Benussi, L.; Rongioletti, M.; Ghidoni, R. Serum Copper is not Altered in Frontotemporal Lobar Degeneration. J. Alzheimer’s Dis. 2018, 63, 1427–1432. [Google Scholar] [CrossRef] [PubMed]
- Squitti, R.; Pasqualetti, P.; Dal Forno, G.; Moffa, F.; Cassetta, E.; Lupoi, D.; Vernieri, F.; Rossi, L.; Baldassini, M.; Rossini, P.M. Excess of serum copper not related to ceruloplasmin in Alzheimer disease. Neurology 2005, 64, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Squitti, R.; Barbati, G.; Rossi, L.; Ventriglia, M.; Dal Forno, G.; Cesaretti, S.; Moffa, F.; Caridi, I.; Cassetta, E.; Pasqualetti, P.; et al. Excess of nonceruloplasmin serum copper in AD correlates with MMSE, CSF [beta]-amyloid, and h-tau. Neurology 2006, 67, 76–82. [Google Scholar] [CrossRef]
- Squitti, R.; Bressi, F.; Pasqualetti, P.; Bonomini, C.; Ghidoni, R.; Binetti, G.; Cassetta, E.; Moffa, F.; Ventriglia, M.; Vernieri, F.; et al. Longitudinal prognostic value of serum “free” copper in patients with Alzheimer disease. Neurology 2009, 72, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Squitti, R.; Ghidoni, R.; Scrascia, F.; Benussi, L.; Panetta, V.; Pasqualetti, P.; Moffa, F.; Bernardini, S.; Ventriglia, M.; Binetti, G.; et al. Free copper distinguishes mild cognitive impairment subjects from healthy elderly individuals. J. Alzheimer’s Dis. 2011, 23, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Lopez, N.; Tormo, C.; De Blas, I.; Llinares, I.; Alom, J. Oxidative stress in Alzheimer’s disease and mild cognitive impairment with high sensitivity and specificity. J. Alzheimer’s Dis. 2013, 33, 823–829. [Google Scholar] [CrossRef]
- Squitti, R.; Pasqualetti, P.; Polimanti, R.; Salustri, C.; Moffa, F.; Cassetta, E.; Lupoi, D.; Ventriglia, M.; Cortesi, M.; Siotto, M.; et al. Metal-score as a potential non-invasive diagnostic test for Alzheimer’s disease. Curr. Alzheimer Res. 2013, 10, 191–198. [Google Scholar] [CrossRef]
- Squitti, R.; Ghidoni, R.; Siotto, M.; Ventriglia, M.; Benussi, L.; Paterlini, A.; Magri, M.; Binetti, G.; Cassetta, E.; Caprara, D.; et al. Value of serum nonceruloplasmin copper for prediction of mild cognitive impairment conversion to Alzheimer disease. Ann. Neurol. 2014, 75, 574–580. [Google Scholar] [CrossRef] [PubMed]
- Rozzini, L.; Lanfranchi, F.; Pilotto, A.; Catalani, S.; Gilberti, M.E.; Paganelli, M.; Apostoli, P.; Padovani, A. Serum Non-Ceruloplasmin Non-Albumin Copper Elevation in Mild Cognitive Impairment and Dementia due to Alzheimer’s Disease: A Case Control Study. J. Alzheimer’s Dis. 2018, 61, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Squitti, R.; Polimanti, R.; Bucossi, S.; Ventriglia, M.; Mariani, S.; Manfellotto, D.; Vernieri, F.; Cassetta, E.; Ursini, F.; Rossini, P.M. Linkage disequilibrium and haplotype analysis of the ATP7B gene in Alzheimer’s disease. Rejuvenation Res. 2013, 16, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Bucossi, S.; Polimanti, R.; Ventriglia, M.; Mariani, S.; Siotto, M.; Ursini, F.; Trotta, L.; Scrascia, F.; Callea, A.; Vernieri, F.; et al. Intronic rs2147363 variant in ATP7B transcription factor-binding site associated with Alzheimer’s disease. J. Alzheimer’s Dis. 2013, 37, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.P.; Lin, W.Y.; Wang, W.F.; Tsai, C.H.; Wu, W.C.; Chiou, M.T.; Shen, C.P.; Wu, B.T.; Tsai, F.J. Genetic variability in copper-transporting P-type adenosine triphosphatase (ATP7B) is associated with Alzheimer’s disease in a Chinese population. J. Biol. Regul. Homeost. Agents 2013, 27, 319–327. [Google Scholar] [PubMed]
- Hoogenraad, T. Diagnosis. In Wilson’s disease, 2nd ed.; Intermed Medical Publishers: Amsterdam, The Netherlands, 2001; pp. 109–137. [Google Scholar]
- Squitti, R.; Simonelli, I.; Cassetta, E.; Lupoi, D.; Rongioletti, M.; Ventriglia, M.; Siotto, M. Patients with Increased Non-Ceruloplasmin Copper Appear a Distinct Sub-Group of Alzheimer’s Disease: A Neuroimaging Study. Curr. Alzheimer Res. 2017, 14, 1318–1326. [Google Scholar] [CrossRef]
- James, S.A.; Volitakis, I.; Adlard, P.A.; Duce, J.A.; Masters, C.L.; Cherny, R.A.; Bush, A.I. Elevated labile Cu is associated with oxidative pathology in Alzheimer’s Disease. Free Radic. Biol. Med. 2012, 52, 298–302. [Google Scholar] [CrossRef]
- Bucossi, S.; Mariani, S.; Ventriglia, M.; Polimanti, R.; Gennarelli, M.; Bonvicini, C.; Pasqualetti, P.; Scrascia, F.; Migliore, S.; Vernieri, F.; et al. Association between the c. 2495 A>G ATP7B Polymorphism and Sporadic Alzheimer’s Disease. Int. J. Alzheimer’s Dis. 2011, 2011, 973692. [Google Scholar] [CrossRef]
- Bucossi, S.; Polimanti, R.; Mariani, S.; Ventriglia, M.; Bonvicini, C.; Migliore, S.; Manfellotto, D.; Salustri, C.; Vernieri, F.; Rossini, P.M.; et al. Association of K832R and R952K SNPs of Wilson’s disease gene with Alzheimer’s disease. J. Alzheimer’s Dis. 2012, 29, 913–919. [Google Scholar] [CrossRef] [PubMed]
- McCann, C.J.; Jayakanthan, S.; Siotto, M.; Yang, N.; Osipova, M.; Squitti, R.; Lutsenko, S. Single nucleotide polymorphisms in the human ATP7B gene modify the properties of the ATP7B protein. Metallomics 2019, 11, 1128–1139. [Google Scholar] [CrossRef] [PubMed]
- Mercer, S.W.; Wang, J.; Burke, R. In Vivo Modeling of the Pathogenic Effect of Copper Transporter Mutations That Cause Menkes and Wilson Diseases, Motor Neuropathy, and Susceptibility to Alzheimer’s Disease. J. Biol. Chem. 2017, 292, 4113–4122. [Google Scholar] [CrossRef] [PubMed]
- Salustri, C.; Siotto, M.; Bucossi, S.; Squitti, R. Metals Involvement in Alzheimer’s Disease—A Patho-Genetic View. In Alzheimer’s Disease—Challenges for the Future; Zerr, P.I., Ed.; InTech: London, UK, 2015; Volume 4. [Google Scholar] [CrossRef]
- Squitti, R.; Polimanti, R. Copper hypothesis in the missing hereditability of sporadic Alzheimer’s disease: ATP7B gene as potential harbor of rare variants. J. Alzheimer’s Dis. 2012, 29, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Squitti, R.; Polimanti, R. Copper phenotype in Alzheimer’s disease: Dissecting the pathway. Am. J. Neurodegener. Dis. 2013, 2, 46–56. [Google Scholar] [PubMed]
- Squitti, R.; Polimanti, R.; Siotto, M.; Bucossi, S.; Ventriglia, M.; Mariani, S.; Vernieri, F.; Scrascia, F.; Trotta, L.; Rossini, P.M. ATP7B variants as modulators of copper dyshomeostasis in Alzheimer’s disease. Neuromolecular Med. 2013, 15, 515–522. [Google Scholar] [CrossRef]
- Squitti, R.; Siotto, M.; Bucossi, S.; Polimanti, R. In silico investigation of the ATP7B gene: Insights from functional prediction of non-synonymous substitution to protein structure. Biometals 2014, 27, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Squitti, R.; Siotto, M.; Ivanova, I.; Rongioletti, M. ATP7B and Alzheimer disease. In Clinical and Translational Perspectives on WILSON DISEASE; Kerkar, N., Roberts, E.A., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2019; Volume 42, pp. 427–436. [Google Scholar]
- Squitti, R.; Ventriglia, M.; Gennarelli, M.; Colabufo, N.A.; El Idrissi, I.G.; Bucossi, S.; Mariani, S.; Rongioletti, M.; Zanetti, O.; Congiu, C.; et al. Non-Ceruloplasmin Copper Distincts Subtypes in Alzheimer’s Disease: A Genetic Study of ATP7B Frequency. Mol. Neurobiol. 2017, 54, 671–681. [Google Scholar] [CrossRef]
- Telianidis, J.; Hung, Y.H.; Materia, S.; Fontaine, S.L. Role of the P-Type ATPases, ATP7A and ATP7B in brain copper homeostasis. Front. Aging Neurosci. 2013, 5, 44. [Google Scholar] [CrossRef]
- Pal, A.; Siotto, M.; Prasad, R.; Squitti, R. Towards a unified vision of copper involvement in Alzheimer’s disease: A review connecting basic, experimental, and clinical research. J. Alzheimer’s Dis. 2015, 44, 343–354. [Google Scholar] [CrossRef]
- Squitti, R.; Tecchio, F.; Ventriglia, M. The Role of Copper in Human Diet and Risk of Dementia. Curr. Nutr. Rep. 2015, 4, 114–125. [Google Scholar] [CrossRef]
- Sparks, D.L.; Schreurs, B.G. Trace amounts of copper in water induce-amyloid plaques and learning deficits in a rabbit model of Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2003, 100, 11065–11069. [Google Scholar] [CrossRef]
- Sparks, D.L.; Friedland, R.; Petanceska, S.; Schreurs, B.G.; Shi, J.; Perry, G.; Smith, M.A.; Sharma, A.; Derosa, S.; Ziolkowski, C.; et al. Trace copper levels in the drinking water, but not zinc or aluminum influence CNS Alzheimer-like pathology. J. Nutr. Health Aging. 2006, 10, 247–254. [Google Scholar] [PubMed]
- Lu, J.; Zheng, Y.L.; Wu, D.M.; Sun, D.X.; Shan, Q.; Fan, S.H. Trace amounts of copper induce neurotoxicity in the cholesterol-fed mice through apoptosis. FEBS Lett. 2006, 580, 6730–6740. [Google Scholar] [CrossRef] [PubMed]
- Arnal, N.; Castillo, O.; de Alaniz, M.J.; Marra, C.A. Effects of Copper and/or Cholesterol Overload on Mitochondrial Function in a Rat Model of Incipient Neurodegeneration. Int. J. Alzheimer’s Dis. 2013, 2013, 645379. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Jing, T.; Niu, L.; Huang, X.; Wang, Y.; Deng, X.; Wang, M. Amyloidogenesis induced by diet cholesterol and copper in a model mouse for Alzheimer’s disease and protection effects of zinc and fluvastatin. Brain Res. Bull. 2018, 143, 1–8. [Google Scholar] [CrossRef]
- Abolaji, A.O.; Fasae, K.D.; Iwezor, C.E.; Aschner, M.; Farombi, E.O. Curcumin attenuates copper-induced oxidative stress and neurotoxicity in Drosophila melanogaster. Toxicol. Rep. 2020, 7, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Lamtai, M.; Zghari, O.; Ouakki, S.; Marmouzi, I.; Mesfioui, A.; EI Hessni, A.; Ouichou, A. Chronic copper exposure leads to hippocampus oxidative stress and impaired learning and memory in male and female rats. Toxicol. Res. 2020. [Google Scholar] [CrossRef]
- Singh, I.; Sagare, A.P.; Coma, M.; Perlmutter, D.; Gelein, R.; Bell, R.D.; Deane, R.J.; Zhong, E.; Parisi, M.; Ciszewski, J.; et al. Low levels of copper disrupt brain amyloid-beta homeostasis by altering its production and clearance. Proc. Natl. Acad. Sci. USA 2013, 110, 14771–14776. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Luo, X.; Xu, H.; Ma, Q.; Yuan, J.; Li, X.; Chang, R.C.; Qu, Z.; Huang, X.; Zhuang, Z.; et al. Identification of the Key Molecules Involved in Chronic Copper Exposure-Aggravated Memory Impairment in Transgenic Mice of Alzheimer’s Disease Using Proteomic Analysis. J. Alzheimer’s Dis. 2015, 44, 455–469. [Google Scholar] [CrossRef]
- Huat, T.J.; Camats-Perna, J.E.; Newcombe, A.; Valmas, N.; Kitazawa, M.; Medeiros, R. Metal Toxicity Links to Alzheimer’s Disease and Neuroinflammation. J. Mol. Biol. 2019, 431, 1843–1868. [Google Scholar] [CrossRef]
- Brewer, G.J. Copper-2 Hypothesis for Causation of the Current Alzheimer’s Disease Epidemic Together With Dietary Changes That Enhance the Epidemic. Chem. Res. Toxicol. 2017, 30, 763–768. [Google Scholar] [CrossRef]
- Esmieu, C.; Guettas, D.; Conte-Daban, A.; Sabater, L.; Faller, P.; Hureau, C. Copper-Targeting Approaches in Alzheimer’s Disease: How To Improve the Fallouts Obtained From in Vitro Studies. Inorg. Chem. 2019, 58, 13509–13527. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, J.R.; Osdaghi, E.; Behlau, F.; Köhl, J.; Jones, J.B.; Aubertot, J.N. Thirteen decades of antimicrobial copper compounds applied in agriculture. A review. Agron. Sustain. Dev. 2018, 38, 1–18. [Google Scholar] [CrossRef]
- Fishel, F.M. Pest Management and Pesticides: A Historical Perspective. Available online: http://moodle.toxoer.com/pluginfile.php/8421/mod_imscp/content/1/Pest_Management_and_Pesticides.pdf (accessed on 24 April 2020).
- Lara, W.H.; Toledo, M.; Takahashi, M.Y. Teores de cobre em café torrado e moído e em café bebida. Rev. Inst. Adolfo Lutz 1975, 35, 17–22. [Google Scholar]
- Almeida, O.C.; Almeida, L.C.C.; Bezerra, J.L. Estudo de fungicidas à base de cobre no controle da vassourade-bruxa do cacaueiro no sudeste da Bahia. Agrotrópica 2002, 14, 97–100. [Google Scholar]
- Can Organic Agriculture Give Up Copper As a Crop Protection Product? Available online: https://inra-dam-front-resources-cdn.brainsonic.com/ressources/afile/444379-9fbc6-resource-expertise-cuivre-en-ab-8-pages-anglais.pdf (accessed on 24 April 2020).
- FAO—Guidelines for the Production, Processing, Labelling and Marketing of Organically Produced Foods1. Available online: http://www.fao.org/docs/eims/upload/230124/CXG_032e.pdf (accessed on 24 April 2020).
- Instrução Normativa Interministerial Nº 28 de 08 de Junho de 2011 (Produção de Organismos Aquáticos).pdf—Português (Brasil). Available online: https://www.gov.br/agricultura/pt-br/assuntos/sustentabilidade/organicos/legislacao/portugues/instrucao-normativa-interministerial-no-28-de-08-de-junho-de-2011-producao-de-organismos-aquaticos.pdf/view (accessed on 27 April 2020).
- Ministério da Agricultura, Pecuária e Abastecimento. Available online: http://www.agricultura.gov.br/assuntos/sustentabilidade/organicos/legislacao/portugues/instrucao-normativa-no-46-de-06-de-outubro-de-2011-producao-vegetal-e-animal-regulada-pela-in-17-2014.pdf/view (accessed on 24 January 2020).
- Ministério da Agricultura, Pecuária e Abastecimento. Available online: http://www.agricultura.gov.br/assuntos/sustentabilidade/organicos/legislacao/portugues/instrucao-normativa-no-17-de-18-de-junho-de-2014.pdf/view (accessed on 24 January 2020).
- ifoam_eu_copper_minimisation_in_organic_farming_may2018_0.pdf. Available online: https://www.ifoam-eu.org/sites/default/files/ifoam_eu_copper_minimisation_in_organic_farming_may2018_0.pdf (accessed on 30 January 2020).
- Kuhne, S.; Roßberg, D.; Rohrig, P.; Mering, F.; Weihrauch, F.; Kanthak, S.; Kienzle, J.; Patzwahl, W.; Reiners, E.; Gitzel, J. The Use of Copper Pesticides in Germany and the Search for Minimization and Replacement Strategies. Org. Farming 2017, 3, 66–75. [Google Scholar] [CrossRef]
- USDA—United States Department of Agriculture. Available online: https://www.ecfr.gov/cgi-bin/text-idx?c=ecfr&SID=9874504b6f1025eb0e6b67cadf9d3b40&rgn=div6&view=text&node=7:3.1.1.9.32.7&idno=7 (accessed on 30 January 2020).
- Calda Bordalesa: Utilidades e Preparo. Ministério da Agricultura. Available online: https://ainfo.cnptia.embrapa.br/digital/bitstream/item/38833/1/FOL200837.pdf (accessed on 20 January 2020).
- EAOPS—East African Organic Products Standard. Available online: https://law.resource.org/pub/eac/ibr/eas.456.2007.html (accessed on 20 January 2020).
- EAC. Aid for Trade Case Story: The East African Organic Products Standard Submission by the United Nations Environment Programme. Available online: https://www.oecd.org/aidfortrade/47719232.pdf. (accessed on 24 January 2020).
- Aplicação de calda bordalesa em frutíferas. Available online: http://www.agricultura.gov.br/assuntos/sustentabilidade/organicos/fichas-agroecologicas/arquivos-sanidade-vegetal/2-aplicacao-de-calda-bordalesa-em-frutiferas.pdf (accessed on 24 January 2020).
- Sistema de produção de uva de mesa do norte de Minas Gerais. Available online: https://sistemasdeproducao.cnptia.embrapa.br/FontesHTML/Uva/MesaNorteMinas/tecnologia.htm (accessed on 12 January 2020).
- Rusjan, D.; Strlič, M.; Pucko, D.; Korošec-Koruza, Z. Copper accumulation regarding the soil characteristics in Sub-Mediterranean vineyards of Slovenia. Geoderma 2007, 141, 111–118. [Google Scholar] [CrossRef]
- Calviño, D.F.; Nóvoa-Muñoz, J.C.; Díaz-Raviña, M.; Arias-Estévez, M. Copper accumulation and fractionation in vineyard soils from temperate humid zone (NW Iberian Peninsula). Geoderma 2009, 153, 119–129. [Google Scholar] [CrossRef]
- Tóth, G.; Hermann, T.; Silva, M.R. Heavy metals in agricultural soils of the European Union with implications for food safety. Environ. Int. 2016, 88, 299–309. [Google Scholar] [CrossRef]
- Sacristán, D.; Carbó, E. Copper Contamination in Mediterranean Agricultural Soils: Soil Quality Standards and Adequate Soil Management Practices for Horticultural Crops. In Soil Contamination—Current Consequences and Further Solutions; Larramendy, M.L., Soloneski, S., Eds.; InTech: London, UK, 2016; Volume 4. [Google Scholar] [CrossRef]
- Wu, C.; Luo, Y.; Zhang, L. Variability of copper availability in paddy fields in relation to selected soil properties in southeast China. Geoderma 2010, 156, 200–206. [Google Scholar] [CrossRef]
- Ávila, G.; Gaete, H.; Sauvé, S.; Neaman, A. Organic matter reduces copper toxicity for the earthworm Eisenia fetida in soils from mining areas in Central Chile. Chil. J. Agric. Res. 2009, 69, 252–259. [Google Scholar] [CrossRef]
- Pietrzak, U.; McPhail, D.C. Copper accumulation, distribution and fractionation in vineyard soils of Victoria, Australia. Geoderma 2004, 122, 151–166. [Google Scholar] [CrossRef]
- Xu, J.; Yang, L.; Wang, Z.; Dong, G.; Huang, J.; Wang, Y. Toxicity of copper on rice growth and accumulation of copper in rice grain in copper contaminated soil. Chemosphere 2006, 62, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Mallmann, F.J.K.; Rheinheimer, D.S.; Ceretta, C.A.; Cella, C.; Minella, J.P.G.; Guma, R.L.; Filipovi, V.; Oort, F.; Šimunek, J. Soil tillage to reduce surface metal contamination—Model development and simulations of zinc and copper content profiles in a pig slurry-amended soil. Agric. Ecosyst. Environ. 2014, 196, 59–68. [Google Scholar] [CrossRef]
- National Research Council (US) Committee on Copper in Drinking Water. Copper in Drinking Water. Available online: https://www.ncbi.nlm.nih.gov/books/NBK225402/ (accessed on 12 January 2020).
- Shen, X.L.; Yu, J.H.; Zhang, D.F.; Xie, J.X.; Jiang, H. Positive relationship between mortality from Alzheimer’s disease and soil metal concentration in mainland China. J. Alzheimer’s Dis. 2014, 42, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Evans, D.A.; Tangney, C.C.; Bienias, J.L.; Schneider, J.A.; Wilson, R.S.; Scherr, P.A. Dietary Copper and High Saturated and trans Fat Intakes Associated With Cognitive Decline. Arch. Neurol. 2006, 63, 1085–1088. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority) Peer review of the pesticide risk assessment of the active substance copper compounds. EFSA J. 2018, 16, 5152.
- Napoli, M.; Cecchi, S.; Grassi, C.; Baldi, A.; Zanchi, C.A.; Orlandini, S. Phytoextraction of copper from a contaminated soil using arable and vegetable crops. Chemosphere 2019, 219, 122–129. [Google Scholar] [CrossRef]
- Rahman, M.M.; Azirun, S.M.; Boyce, A.N. Enhanced Accumulation of Copper and Lead in Amaranth (Amaranthus paniculatus), Indian Mustard (Brassica juncea) and Sunflower (Helianthus annuus). PLoS ONE 2013, 8, e62941. [Google Scholar] [CrossRef]
- Hummes, A.P.; Bortoluzzi, E.C.; Tonini, V.; Silva, L.; Petry, C. Transfer of Copper and Zinc from Soil to Grapevine-Derived Products in Young and Centenarian Vineyards. Water Air Soil Pollut. 2019, 230, 150–161. [Google Scholar] [CrossRef]
- Santos, C.E.; Debastiani, R.; Souza, V.S.; Peretti, D.E.; Jobim, P.F.; Yoneama, M.L.; Amaral, L.; Dias, J.F. The influence of the winemaking process on the elemental composition of the Marselan red wine. J. Sci. Food Agric. 2019, 99, 4642–4650. [Google Scholar] [CrossRef]
- Donici, A.C.; Bunea, I.; Călugăr, A.; Harsan, E.; Bora, F.D. Investigation of the Copper Content in Vineyard Soil, Grape, Must and Wine in the Main Vineyards of Romania: A Preliminary Study. Bull. UASVM Hortic. 2019, 76, 31–46. [Google Scholar] [CrossRef]
- Rehman, M.; Liu, L.; Wang, Q.; Saleem, M.H.; Bashir, S.; Ullah, S.; Peng, D. Copper environmental toxicology, recent advances, and future outlook: A review. Environ. Sci. Pollut. Res. 2019, 26, 18003–18016. [Google Scholar] [CrossRef] [PubMed]
- Alexander, P.D.; Alloway, B.J.; Dourado, A.M. Genotypic variations in the accumulation of Cd, Cu, Pb and Zn exhibited by six commonly grown vegetables. Environ. Pollut. 2006, 144, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Garrido, L.R.; Botton, M. Recomendações Técnicas para o Manejo das Pragas e Doenças Fúngicas da Videira na Região Sul do Brasil. In Circula Téchnica; Empresa Brasileira de Pesquisa Agropecuária: Bento Gonçalves, RS, Brasil, 2015; Volume 117, pp. 3–23. [Google Scholar]
- 1516729946. Available online: http://www.ihara.com.br/upload/produtos/fispq/1516729946.pdf (accessed on 24 January 2020).
- Massie, H.R.; Aiello, V.R.; Iodice, A.A. Changes with age in copper and superoxide dismutase levels in brains of C57BL/6J mice. Mech. Ageing Dev. 1979, 10, 93–99. [Google Scholar] [CrossRef]
- Zatta, P.; Drago, D.; Zambenedetti, P.; Bolognin, S.; Nogara, E.; Peruffo, A.; Cozzi, B. Accumulation of copper and other metal ions, and metallothionein I/II expression in the bovine brain as a function of aging. J. Chem. Neuroanat. 2008, 36, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Vasudevaraju, P.; Bharathi, T.J.; Shamasundar, N.M.; Subba Rao, K.; Balaraj, B.M.; Rao, K.S.J.; Sathyanarayana Rao, T.S. New evidence on Iron, Copper accumulation and Zinc depletion and its correlation with DNA integrity in aging human brain regions. Indian J. Psychiatry 2010, 52, 104–144. [Google Scholar]
- Vanacore, R.; Eskew, J.D.; Sung, L.; Davis, T.; Smith, A. Safe Coordinated Trafficking of Heme and Iron With Copper Maintain Cell Homeostasis: Modules From the Hemopexin System. Biometals 2019, 32, 355–367. [Google Scholar] [CrossRef]
- Pohanka, M. Copper and copper nanoparticles toxicity and their impact on basic functions in the body. Bratisl Lek Listy 2019, 120, 397–409. [Google Scholar] [CrossRef]
- Lavado, L.K.; Zhang, M.H.; Patel, K.; Khan, S.; Patel, U.K. Biometals as Potential Predictors of the Neurodegenerative Decline in Alzheimer’s Disease. Cureus 2019, 5, 1–11. [Google Scholar] [CrossRef]
- Hsu, H.W.; Bondy, S.C.; Kitazawa, M. Environmental and Dietary Exposure to Copper and Its Cellular Mechanisms Linking to Alzheimer’s Disease. Toxicol. Sci. 2018, 163, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Pal, I.; Roy, M.; Dey, S.G. Active-site Environment of Cu Bound Amyloid β and Amylin Peptides. J. Biol. Inorg. Chem. 2019, 24, 1245–1259. [Google Scholar] [CrossRef] [PubMed]
- La Torre, A.; Iovino, V.; Caradonia, F. Copper in plant protection: Current situation and prospects. Phytopathol. Mediterr. 2018, 57, 201–236. [Google Scholar]
- Pertot, I.; Bilali, H.E.; Simeone, V.; Vecchione, A.; Zulini, L. Efficacy evaluation and phytotoxicity assessment of traditional and new copper compounds used in copper reduction strategies in organic viticulture in northern and southern Italy environments. Integr. Prot. Vitic. IOBC/wprs Bull. 2006, 29, 61–65. [Google Scholar]
- 278–284 Smith J et al.—SRUC. Available online: http://www.sruc.ac.uk/downloads/file/2022/278-284 (accessed on 24 March 2020).
- Torre, A.L.; Righi, L.; Iovino, V.; Battaglia, V. Control of late blight in organic farming with low copper dosages or natural products as alternatives to copper. Eur. J. Plant Pathol. 2019, 155, 769–778. [Google Scholar] [CrossRef]
- Moonen, A.C.; Bàrberi, P. Functional biodiversity: An agroecosystem approach. Agric. Ecosyst. Environ. 2008, 127, 7–21. [Google Scholar] [CrossRef]
- Bankina, B.; Bimsteine, G.; Paulovska, L.; Paura, L.; Pavlovica, O.; Kaneps, J.; Neusa-Luca, I.; Roga, A.; Fridmanis, D. Effects of soil tillage and crop rotation on the development of wheat stem base diseases. Can. J. Plant Pathol. 2019, 41, 435–442. [Google Scholar] [CrossRef]
- Runno-Paurson, E.; Laaniste, P.; Eremeev, V.; Tahtjarv, T.; Kaurilind, E.; Tosens, T.; Niinemets, U.; Williams, I.H. Does winter oilseed rape as a winter cover crop influence potato late blight development in an organic crop rotation? Biol. Agric. Hortic. 2019, 36, 71–83. [Google Scholar] [CrossRef]
- Page, K.; Dang, Y.; Dalal, R. Impacts of conservation tillage on soil quality, including soil-borne crop diseases, with a focus on semi-arid grain cropping systems. Australas. Plant Pathol. 2013, 42, 363–377. [Google Scholar] [CrossRef]
- Lage, D.A.C.; Marouelli, W.A.; Cafe, A.C. Management of powdery mildew and behaviour of late blight under different irrigation configurations in organic tomato. Crop Prot. 2019, 125, 104886. [Google Scholar] [CrossRef]
- Merwin, I.A.; Wilcox, W.F.; Stiles, W.C. Influence of orchard ground cover management on the development of Phytophthora crown and root rots of apple. Plant Dis. 1992, 76, 199–205. [Google Scholar] [CrossRef]
- Vukicevich, E.; Lowery, D.T.; Urbez-Torres, J.R.; Bowen, P.; Hart, M. Groundcover management changes grapevine root fungal communities and plant-soil feedback. Plant Soil 2018, 424, 419–433. [Google Scholar] [CrossRef]
- Holb, I.J. Effect of pruning on apple scab in organic apple production. Plant Dis. 2005, 89, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Commission Implementing Regulation (EU) 2015/232 of 13 February 2015. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32015R0232&from=EN (accessed on 24 April 2020).
- Farming without plant protection products Can we grow without using herbicides, fungicides and insecticides? Available online: http://www.europarl.europa.eu/cmsdata/185760/EPRS_IDA(2019)634416_EN.pdf (accessed on 24 April 2020).


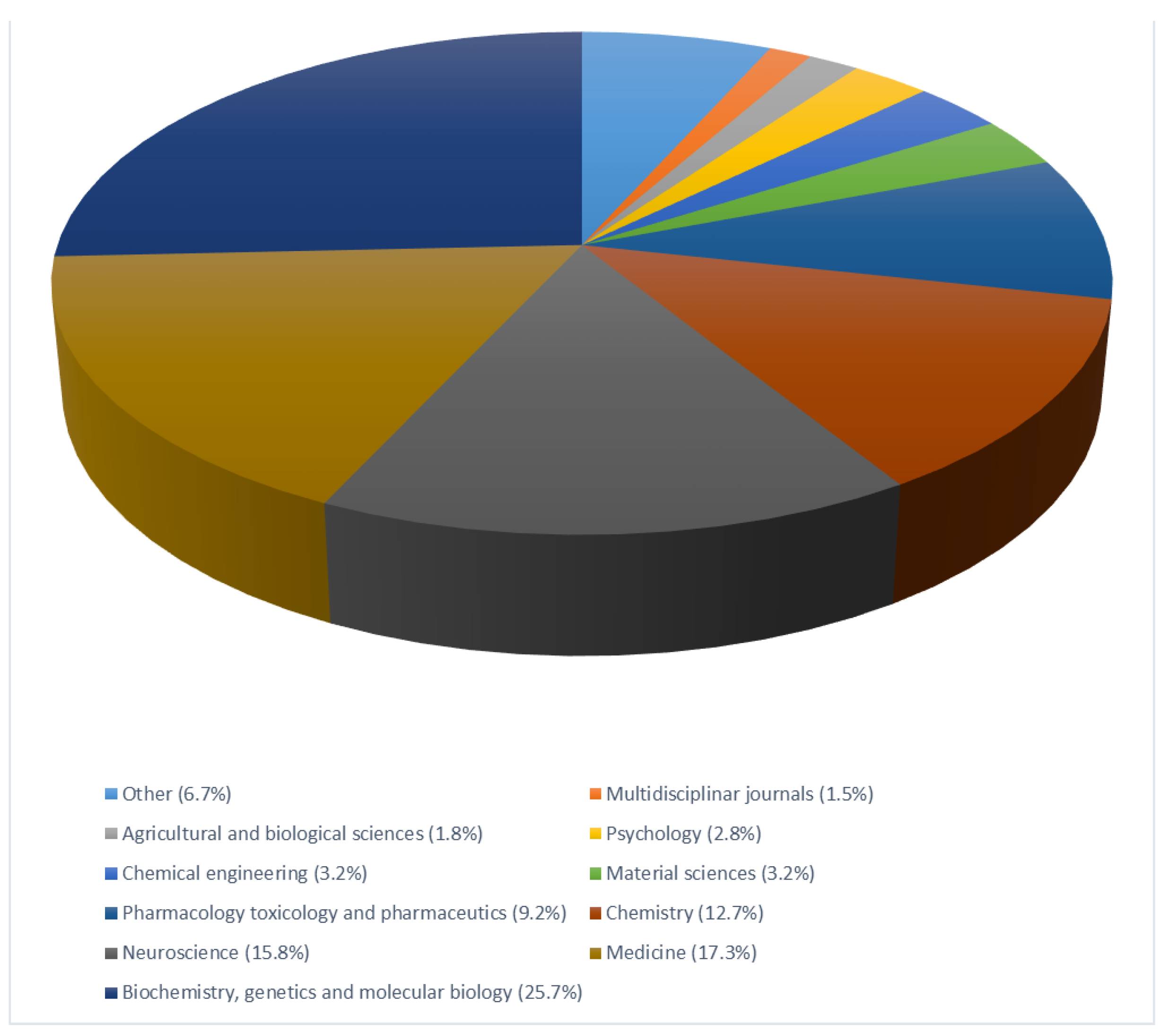{kind=link}
| Subjects | Risk (OR 1, RR 2, HR 3) | CI 4 95% | p Value | |
|---|---|---|---|---|
| Serum Copper and Risk for Alzheimer’s Disease | ||||
| Copper level was higher in subjects with AD than in control subjects and correlated with poor neuropsychological performance and medial temporal lobe atrophy [35] | 76 AD vs. 79 healthy subjects | 1.8 | 1.36–2.43 | p < 0.05 |
| Copper level was higher in subjects with AD than in patients with vascular dementia subjects (VaD) [36] | 48 AD vs. 20 VaD | 2.06 | 1.28–3.31 | p < 0.003 |
| Non-ceruloplasmin copper was higher in AD than in healthy controls and VaD and correlated with poor neuropsychological performance [37] | 47 AD vs. 44 healthy subjects and 24 VaD subjects | p < 0.001 | ||
| Non-ceruloplasmin copper was higher in AD than in healthy controls; Cerebrospinal (CSF) β-amyloid and H-Tau correlated with serum non-ceruloplasmin copper; copper in the CSF was partially dependent on the serum Non-ceruloplasmin copper (t = 2.2, p = 0.04). Mini-Mental State Examination (MMSE) and verbal memory scores correlated positively with β-amyloid (r = 0.46, p = 0.002) and inversely with nonceruloplasmin-Cu (= 0.45, p = 0.003) [38] | 28 AD vs. 25 healthy subjects | p < 0.001 | ||
| Non-ceruloplasmin copper predicted the annual change in MMSE; when the annual change in MMSE was divided into <3 or ≥3 points, Non-ceruloplasmin copper was the only predictor of a more severe decline [39] | 81 AD subjects, 1 year longitudinal study | 1.23 | 1.03–1.47 | p < 0.022 |
| Non-ceruloplasmin copper was higher in MCI than in healthy subjects [40] | 83 MCI subjects, 100 healthy subjects | 1.22 | 1.05–1.41 | p < 0.01 |
| Copper level showed a significant increase in the serum of AD and MCI compared to control (p = 0.038) [41] | 36 AD, 18 MCI vs. 25 healthy subjects | p < 0.05 | ||
| Non-ceruloplasmin copper increased the risk of having AD; when combined in an algorithm with sex, APOE, Cp/Tf, TAS, the ability to discriminate AD patients vs. controls was high (ROC 5, AUC 6 = 0.9) [42] | 93 AD, 45 VaD, 48 healthy subjects | 3.21 | 1.53–6.71 | p < 0.002 |
| Non-ceruloplasmin copper was a predictor of conversion to AD: MCI subjects with nonceruloplasmin-Cu levels > 1.6 µmol/L had a hazard conversion rate (50% conversion in 4 years) that was ~3 higher than those with values ≤ 1.6 µmol/L (< 20% in 4 years) [43] | 131 MCI subjects, 6 years longitudinal study | 3.3 | 1.21–9.24 | p = 0.02 |
| Non-ceruloplasmin copper levels higher in MCI and AD with respect to control (p < 0.0001) [44] | 44 AD and 36 MCI vs. 28 healthy subjects | p < 0.001 | ||
| Non-ceruloplasmin copper and Cu:Cp resulted higher in AD and in Wilson disease (WD) than in healthy controls; while nCp-Cu was similar between AD and WD, Cu:Cp was higher in WD. 24 h urinary copper excretion in AD patients (12.05 μg/day) was higher than in healthy controls (4.82 μg/day); 77.8% of the AD patients under D-penicillamine treatment had a 24 h urinary excretion higher than 200 μg/day, suggestive of a failure of copper control [34] | 385 AD, 9 WD, 336 healthy subjects | p < 0.0001 | ||
| Non-ceruloplasmin copper does not change in frontotemporal lobar degeneration (FTLD) [36] | 85 FTLD, 55 healthy subjects | p < 0.001 | ||
| ATP7B Gene Variants and Risk for Alzheimer’s Disease | ||||
| Specific genetic variants in the ATP7B gene, namely rs1801243 (OR = 1.52, 95% CI = 1.10–2.09), rs2147363 (OR = 1.58, 95% CI = 1.11–2.25), rs1061472 (OR = 1.73, 95% CI = 1.23–2.43), and rs732774 (OR = 2.31, 95% CI = 1.41–3.77) increased the risk of having AD [45] | 285 AD vs. 230 healthy subjects | 2.3 | 1.41–3.77 | p < 0.001 |
| Wilson disease-causing variant rs7334118 in linkage disequilibrium with the intronic rs2147363 (associated with AD risk) was detected in two AD patients but in no healthy individuals. However, this Wilson disease mutation did not explain the observed genetic association of rs2147363. Conversely, in silico analyses of rs2147363 functionality highlighted that this variant is located in a binding site of a transcription factor and is associated with regulatory functions [46] | 286 AD vs. 283 healthy subjects | 1.3 | 1.06–1.69 | p = 0.015 |
| Haplotype TGC in specific genetic variants in the ATP7B gene, namely rs1801243, rs1801249, rs1801244, and rs1801243 was associated with an increased risk of having AD [47] | 120 AD vs. 111 healthy subjects | 5.16 | 2.54–10.5 | p < 0.001 |
| Authors, Year | Animal Model | Dose and Route | Duration | Effects |
|---|---|---|---|---|
| Animal Models of Copper Neurotoxicity Induced by Altered Diet | ||||
| Sparks and Schreurs, 2003 [65] | New Zealand white rabbits | 12 mg/L copper in DW 1 + 2% cholesterol − oral | 10 weeks | Accumulation of Aβ in brain; deficit in complex memory acquisition |
| Sparks et al., 2006 [66] | Beagle dogs | 200 mg/L CuSO4 in DW + high fat diet − oral | 4 months | Extracellular Aβ deposits |
| Lu et al., 2006 [67] | Kumming strain mice | 0.21 mg/L copper in DW + 2% cholesterol − oral | 8 weeks | Cognitive deficits; neuronal apoptosis |
| Arnal et al., 2013 [68] | Wistar rat | 3 mg/L copper in tap water + 2% cholesterol − oral | 2 months | Increased oxidative stress in brain; increased non-ceruloplasmin copper in hippocampus; increased Aβ (1–42)/ Aβ (1–40) in cortex and hippocampus |
| Arnal et al., 2013 [68] | Wistar rat | 3 mg/L copper in tap water + 2% cholesterol − oral | 8 weeks | Slight nut noticeable change in visuo-spatial memory |
| Yao et al., 2018 [69] | Tg2567 mouse | 0.1 mg/L copper in drinking water and 2% cholesterol in the food | 3 months | Significant deposit of Aβ and senile-plaque formation in hippocampus and temporal cortex regions |
| Abolaji et al., 2020 [70] | D. melanogaster flies | Cu2+ (1 mM) | 7 days | reduced survival |
| Lamtai et al., 2020 [71] | Rat | CuCl2 (0.25 mg/kg, 0.5 mg/kg and 1 mg/kg) injected intraperitoneally | 8 weeks | Working memory, spatial learning and memory were significantly impaired in rats treated with Cu at dose of 1 mg/kg |
| Models of Copper Neurotoxicity in Genetically Compromised Animals | ||||
| Sparks et al., 2006 [66] | Watanable rabbits | 0.13 mg/L copper in DW − oral | 10 weeks | Accumulation of Aβ in superior temporal cortex and hippocampus |
| Sparks et al., 2006 [66] | PS1/APP transgenic mice | 0.12 mg/L Cu in DW − oral | 6 weeks | Deposition of Aβ |
| Singh et al., 2013 [72] | APP sw/0 mice | 0.13 mg/L copper in DW − oral | 90 days | Increase brain Aβ production; increased neuroinflammation; memory impairment; increased Cu levels in brain capillaries and parenchyma |
| Yu et al., 2014 [73] | 3xTg-AD | 250 mg/L CuSO4 in drinking water | 6 months | memory impairment |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coelho, F.C.; Squitti, R.; Ventriglia, M.; Cerchiaro, G.; Daher, J.P.; Rocha, J.G.; Rongioletti, M.C.A.; Moonen, A.-C. Agricultural Use of Copper and Its Link to Alzheimer’s Disease. Biomolecules 2020, 10, 897. https://doi.org/10.3390/biom10060897
Coelho FC, Squitti R, Ventriglia M, Cerchiaro G, Daher JP, Rocha JG, Rongioletti MCA, Moonen A-C. Agricultural Use of Copper and Its Link to Alzheimer’s Disease. Biomolecules. 2020; 10(6):897. https://doi.org/10.3390/biom10060897
Chicago/Turabian StyleCoelho, Fábio C., Rosanna Squitti, Mariacarla Ventriglia, Giselle Cerchiaro, João P. Daher, Jaídson G. Rocha, Mauro C. A. Rongioletti, and Anna-Camilla Moonen. 2020. "Agricultural Use of Copper and Its Link to Alzheimer’s Disease" Biomolecules 10, no. 6: 897. https://doi.org/10.3390/biom10060897
APA StyleCoelho, F. C., Squitti, R., Ventriglia, M., Cerchiaro, G., Daher, J. P., Rocha, J. G., Rongioletti, M. C. A., & Moonen, A.-C. (2020). Agricultural Use of Copper and Its Link to Alzheimer’s Disease. Biomolecules, 10(6), 897. https://doi.org/10.3390/biom10060897