The Cancer Chemopreventive and Therapeutic Potential of Tetrahydrocurcumin



Abstract
1. Introduction
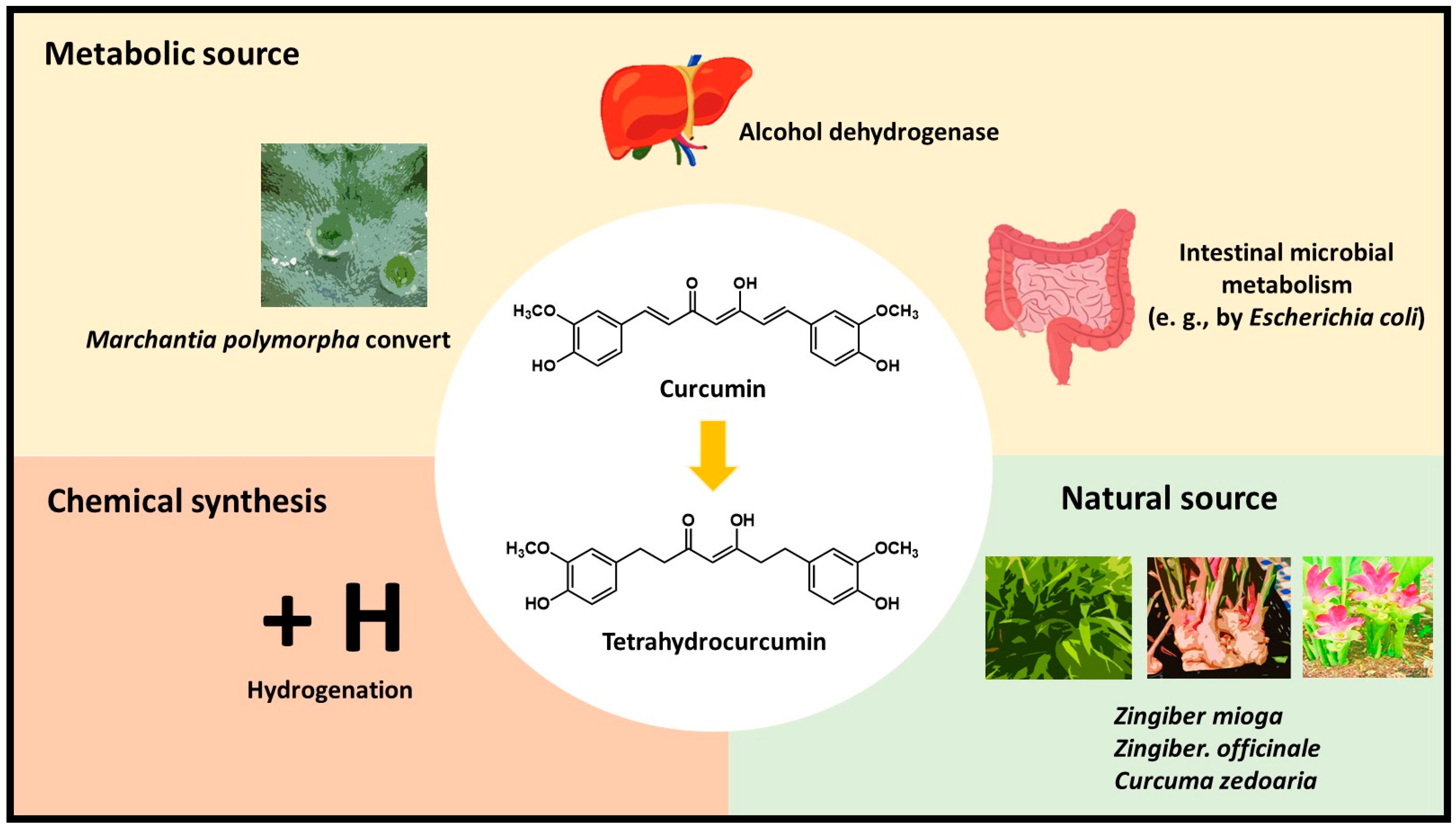
2. Tetrahydrocurcumin, a Candidate Metabolite of Curcumin
2.1. Characterization of Tetrahydrocurcumin
2.2. Beneficial Effects of Tetrahydrocurcumin on Human Diseases
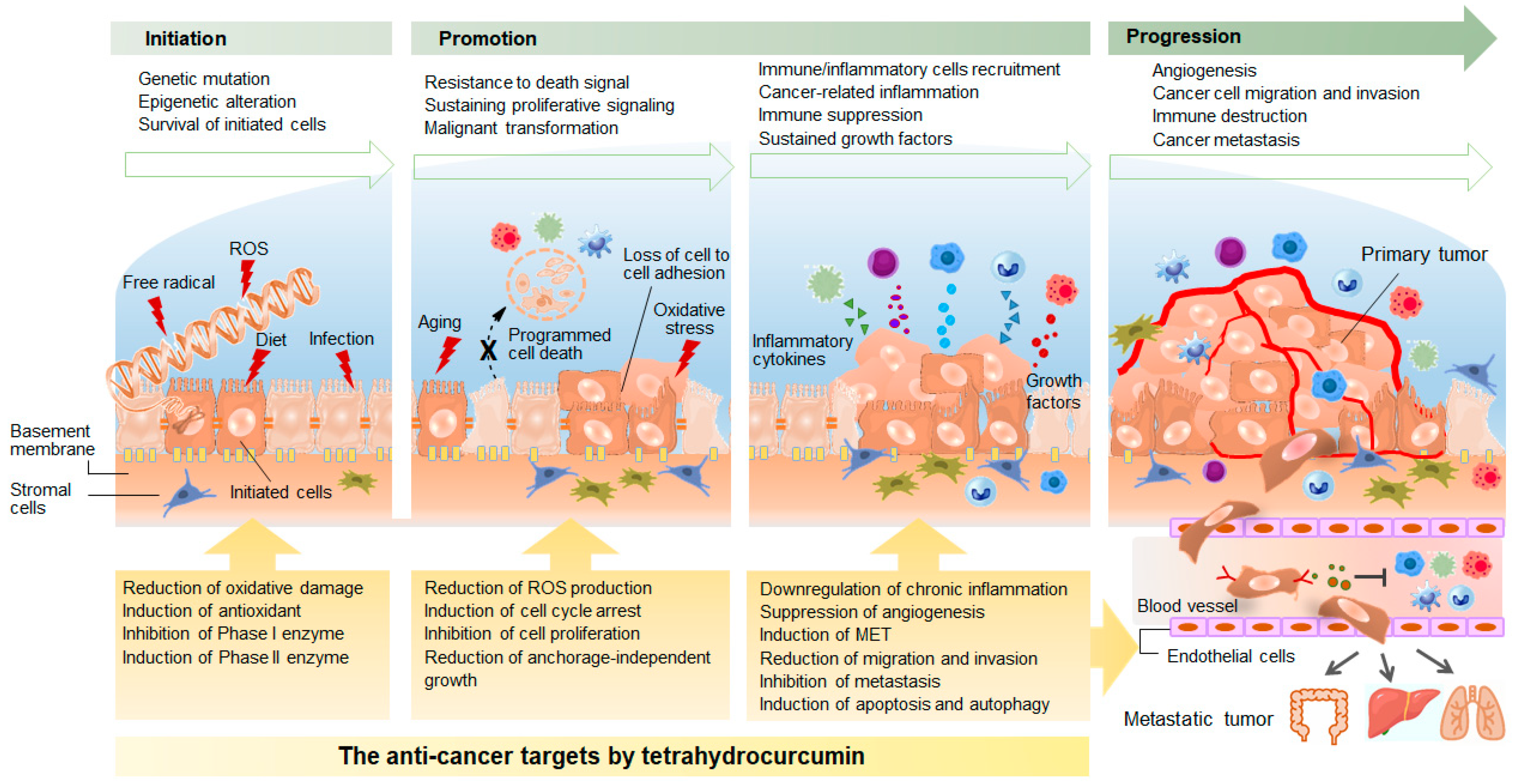
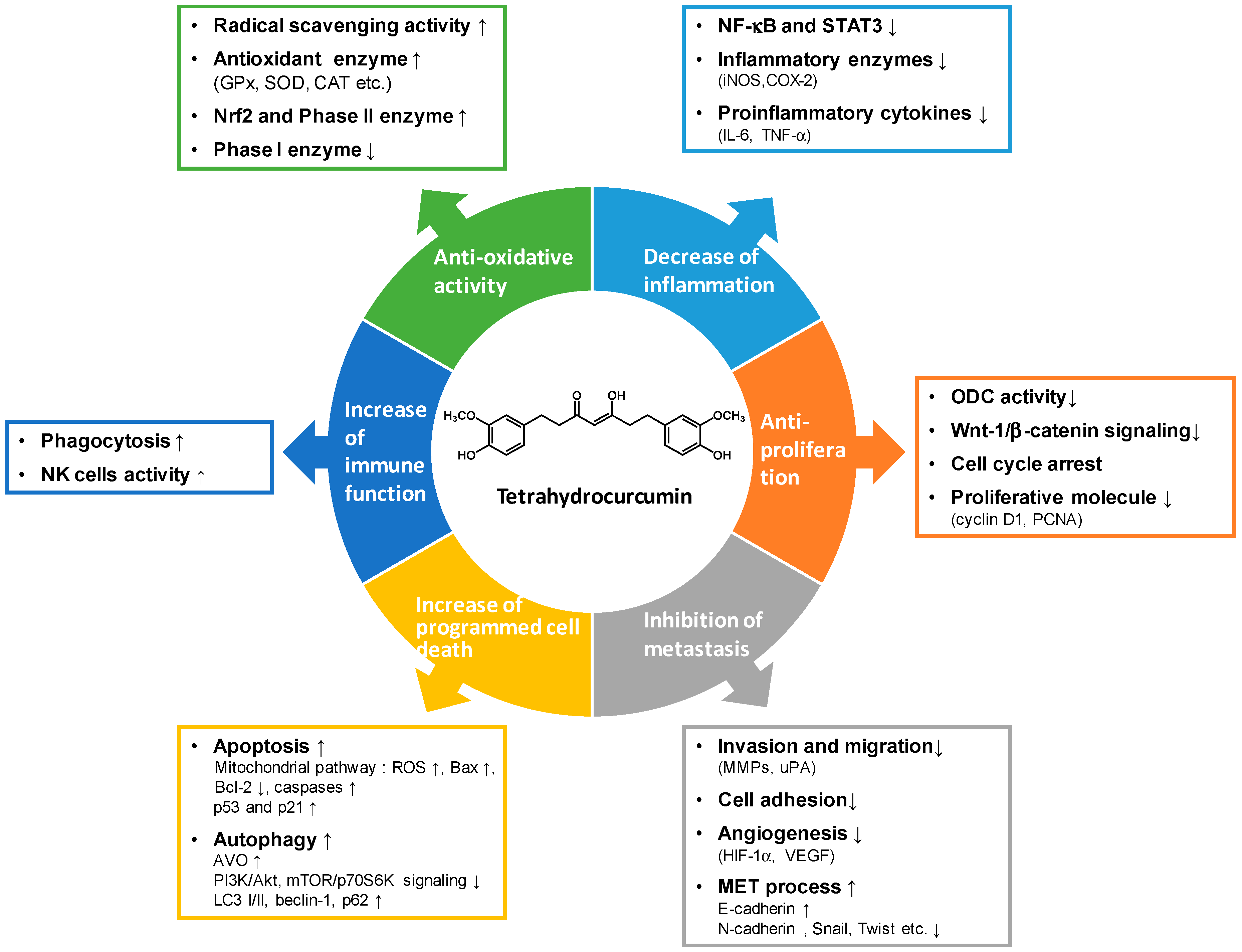
3. Anti-Cancer Effects and the Underlying Mechanisms of Tetrahydrocurcumin Activity
3.1. Anti-Oxidative Activity

{kind=link}
{kind=link}
{kind=link}
| Targeted Cancer | Experimental Model | Tetrahydrocurcumin Concentration | Mechanism of Actions | References | |
|---|---|---|---|---|---|
| Anti-oxidative activity | |||||
| Radical scavenging | - | DPPH radical solution | IC50 = 4.1 ~ 20.7 μM | Radical scavenging | [32,59,60] |
| - | PMA-induced cells | IC50 = 200 μM | O2-• Radical scavenging | [60] | |
| - | Hypoxanthine/xanthine oxidase reaction | 300 μM | O2-• Radical scavenging | [59] | |
| - | Fe2SO4/H2O2 reaction | 200 μM | OH• Radical scavenging | [59] | |
| - | SNAP reaction | IC50 = 104.2 μM | NO Radical scavenging | [59] | |
| Reduction of oxidative damage | - | AAPH-induced linoleic oxidation | 1–12 μM | Reduced lipid peroxidation | [32] |
| Renal | Fe-NTA-induced renal damage in male ddY mice | 0.5% in diet for 4 weeks | Reduced DNA, lipid, and protein oxidative damage | [28] | |
| Induction of antioxidant | Renal | Fe-NTA-induced renal damage in male ddY mice | 0.5% in diet for 4 weeks | Upregulated antioxidant enzymes | [28] |
| Liver | As-induced hepatotoxicity in male albino Wistar rats | 80 mg/kg for 4 weeks | Reduced lipid peroxidation Upregulated antioxidant enzymes | [61] | |
| Liver | Cd-induced hepatotoxicity in male albino Wistar rats | 20, 40, and 80 mg/kg for 4 weeks | Reduced lipid peroxidation Upregulated antioxidant enzymes | [62] | |
| Modulation of Phase I and Phase II enzymes | - | Murine hepatoma cells Hepa 1c1c7 | - | Upregulated NAD(P)H: quinone oxidoreductase | [63] |
| - | Human CYP450 enzymes | 0.01–100 μM | Inhibited CYP2C9 and CYP3A4 activity | [36] | |
| - | Acetaminophen-induced liver injury in male Kunming mice | 25, 50, and 100 mg/kg | Downregulated CYP2E1 gene Downregulated Kepa1 Upregulated Nrf2 and downstream genes | [64] | |
| Anti-inflammatory activity | - | TPA-stimulated HL-60 cells and mouse skin |
| Decreased ROS production Decreased MPO activity | [65] |
| - | LPS-treated RAW264.7 macrophage and C57BL/6 mice |
| Decreased TNF-α production Downregulated phospho-IκB and NF-κB | [66] | |
| - | LPS-treated RAW264.7 macrophage | 3.125–100 μM | Decreased NO, TNF-α, IL-6 production Downregulated NF-κB nuclear translocation | [67] | |
| Colon cancer | DSS-induced colitis in ICR mice | 0.1 and 0.25 mmol/kg for 7 days | Downregulated NF-κB and STAT3 DNA binding activity Downregulated iNOS and COX-2 | [68] | |
| - | Soybean lipooxygenase | 1, 10, and 250 μg/mL | Downregulated lipoxygenase activity | [36] | |
| - | Molecular docking assay | - | Possible phospholipase A2 inhibitor | [52] | |
| Anti-proliferative activity | - | TPA-stimulated mouse skin and JB6 cells |
| Reduced ODC activity Decreased anchorage-independent growth | [69] |
| Colon cancer | DMH-initiated mice | Dietary 0.5% for 7 weeks | Reduced ACF formation and crypt proliferation | [70] | |
| Colon cancer | AOM-treated mice | Dietary 0.005 and 0.02% for 23 weeks | Reduced ACF formation Downregulated iNOS and COX-2 Decreased PCNA Downregulated Wnt-1/β-catenin/p-GSK3β signaling | [71] | |
| Glioma | Glioma cells (alone or combined with radiation) | 3–161 μM | Reduced colony formation Induced G0/G1 cell cycle arrest Downregulated cyclin D1 and PCNA Increased GSH depletion | [72] | |
| Anti-metastatic activity | Fibrosarcoma | HT1080 human fibrosarcoma cells | 5–100 μM | Decreased invasion and migration Decreased cell adhesion Downregulated MMPs and uPA | [73] |
| Liver cancer | HepG2 xenograft model | Oral 3000 mg/kg for 21 days | Decreased angiogenesis | [46] | |
| Cervical cancer | CaSki xenograft model | Oral 100, 300 and 500 mg/kg for 30 days | Decreased angiogenesis Decreased tumor volume Downregulated HIF-1α, VEGF, VEGFR2, EGFR, COX-2 Downregulated p-ERK1/2 and p-AKT | [74,75] | |
| Osteosarcoma | Lung metastasis model Osteosarcoma cell lines |
| Reduced lung metastasis Decreased invasion and migration Promoted MET process Downregulated HIF-1α, VEGF, and MMPs Decreased hypoxia-induced angiogenesis | [76] | |
| Induction of programmed cell death | Breast cancer | MCF-7 cells | 15–130 μM | Induced mitochondria-dependent apoptosis Loss of ΔΨm Increased ROS production Upregulated Bax and Downregulated Bcl-2 Upregulated p21 Activation of caspases | [77,78] |
| Liver cancer | H22 ascites tumor-bearing mouse model | i.p. 5, 10 and 20 mg/kg for 7 days | Induced mitochondria-dependent apoptosis Upregulated Bax and Downregulated Bcl-2 Activation of caspases Upregulated p53 and downregulated MDM2 | [79] | |
| Leukemia | HL-60 cells | 25–100 μM | Induced autophagy Upregulated LC3 I/II and beclin-1 Downregulated PI3K/Akt and mTOR/p70S6K signaling | [80] | |
| Leukemia | Cytarabine-resistance HL-60 cells | 5–100 μM | Induced autophagy Upregulated p62 and beclin-1 | [81] | |
| Lung cancer | A549 cells | 30–130 μM | Induced autophagy Upregulated LC3 I/II and beclin-1 Downregulated PI3K/Akt/mTOR signaling | [82] | |
| Immuno-modulating activity | - | RAW264.7 macrophages and LPS-stimulated mouse splenocytes | 1–10 μg/mL | Increased phagocytosis Increased NK cells activity | [83,84] |
3.2. Modulation of Phase I and Phase II Enzymes
3.3. Anti-Inflammatory Activity
3.4. Anti-Proliferative Activity
3.5. Anti-Metastatic Activity
3.6. Induction of Programmed Cell Death
3.7. Immuno-Modulating Activity
4. Enhancement of Tetrahydrocurcumin Biological Activity Via Structural Modification and Increased Delivery
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AAPH | 2,2′-azobis(2-amidinopropane) dihydrochloride |
| ACF | aberrant crypt foci |
| AOM | azoxymethane |
| AUC | area under curve |
| AVOs | acidic vascular organelles |
| Cmax | maximum plasma concentration |
| COX-2 | cyclooxygenase-2 |
| CYP450 | cytochrome P450 |
| DMH | 1,2-dimethylhydrazine dihydrochloride |
| DSS | dextran sulfate sodium |
| ECM | extracellular matrix |
| EMT | epithelial-mesenchymal transition |
| Fe-NTA | ferric nitrilotriacetate |
| GCLC | glutamate cysteine ligase catalytic subunit |
| GCLM | glutamate-cysteine ligase modifier subunit |
| GPx | glutathione peroxidase |
| GSH | glutathione |
| GST | glutathione S-transferase |
| HIF | hypoxia-inducible factor |
| HNE | hydroxynonenal |
| HO-1 | heme oxygenase-1 |
| Inos | inducible nitric oxide synthase |
| Ir | iridium |
| LPS | lipopolysaccharide |
| MET | mesenchymal-epithelial transition |
| MMPs | matrix metalloproteinases |
| MPO | Myeloperoxidase |
| NF-κB | nuclear factor-κB |
| NO | nitric oxide |
| ODC | ornithine decarboxylase |
| PCNA | proliferating cell nuclear antigen |
| PMA | phorbol 12-myristate 13-acetate |
| SIRPα | signal regulatory protein alpha |
| SNAP | S-nitroso-N-acethylpenicillamine |
| SOD | superoxide dismutase |
| STAT3 | signal transducer and activator of transcription 3 |
| TNF-α | tumor necrosis factor-α |
| TPA | tumor promoter 12-O-tetradecanoylphorbol-13-acetate |
| uPA | urokinase type plasminogen activator |
| VEGF | vascular endothelial growth factor |
| 8-OHdG | hydroxy-2′-deoxyguanosine |
| ΔΨm | mitochondrial transmembrane potential |
References
- Benzie, I.F.F.; Wachtel-Galor, S. Herbal Medicine: Biomolecular and Clinical Aspects, 2nd ed.; CRC Press: Cleveland, OH, USA, 2011. [Google Scholar]
- Vogel, H.; Pelletier, J. Curcumin-biological and medicinal properties. J. macie 1815, I, 289. [Google Scholar]
- Schraufstatter, E.; Bernt, H. Antibacterial action of curcumin and related compounds. Nature 1949, 164, 456. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Patchva, S.; Koh, W.; Aggarwal, B.B. Discovery of curcumin, a component of golden spice, and its miraculous biological activities. Clin. Exp. Pharmacol. Physiol. 2012, 39, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yuan, W.; Deng, G.; Wang, P.; Yang, P. Chemical composition and product quality control of turmeric (Curcuma longa L.). Pharm. Crops 2011, 2, 28–54. [Google Scholar]
- Rajkumari, S. Nutritional value, phytochemical composition, and biological activities of edible Curcuma species: A review. Int. J. Food Prop. 2018, 20, S2668–S2687. [Google Scholar] [CrossRef]
- Amalraj, A.; Pius, A.; Gopi, S.; Gopi, S. Biological activities of curcuminoids, other biomolecules from turmeric and their derivatives—A review. J. Tradit. Complement. Med. 2017, 7, 205–233. [Google Scholar]
- Rahmani, A.H.; Alsahli, M.A.; Aly, S.M.; Khan, M.A.; Aldebasi, Y.H. Role of Curcumin in Disease Prevention and Treatment. Adv. Biomed. Res. 2018, 7, 38. [Google Scholar] [CrossRef]
- Hassan, F.U.; Rehman, M.S.; Khan, M.S.; Ali, M.A.; Javed, A.; Nawaz, A.; Yang, C. Curcumin as an Alternative Epigenetic Modulator: Mechanism of Action and Potential Effects. Front. Genet. 2019, 10, 514. [Google Scholar] [CrossRef]
- Singh, S.; Khar, A. Biological effects of curcumin and its role in cancer chemoprevention and therapy. Anticancer Agents Med. Chem. 2006, 6, 259–270. [Google Scholar] [CrossRef]
- Doello, K.; Ortiz, R.; Alvarez, P.J.; Melguizo, C.; Cabeza, L.; Prados, J. Latest in Vitro and in Vivo Assay, Clinical Trials and Patents in Cancer Treatment using Curcumin: A Literature Review. Nutr. Cancer 2018, 70, 569–578. [Google Scholar]
- Liu, W.; Zhai, Y.; Heng, X.; Che, F.Y.; Chen, W.; Sun, D.; Zhai, Z. Oral bioavailability of curcumin: Problems and advancements. J. Drug Target 2016, 24, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Tomeh, M.A.; Hadianamrei, R.; Zhao, X. A Review of Curcumin and Its Derivatives as Anticancer Agents. Int. J. Mol. Sci. 2019, 20, 1033. [Google Scholar] [CrossRef] [PubMed]
- Shoba, G.; Joy, D.; Joseph, T.; Majeed, M.; Rajendran, R.; Srinivas, P.S. Influence of piperine on the pharmacokinetics of curcumin in animals and human volunteers. Planta Med. 1998, 64, 353–356. [Google Scholar] [CrossRef]
- Vyas, A.; Dandawate, P.; Padhye, S.; Ahmad, A.; Sarkar, F. Perspectives on new synthetic curcumin analogs and their potential anticancer properties. Curr. Pharm Des. 2013, 19, 2047–2069. [Google Scholar] [PubMed]
- Lopresti, A.L. The Problem of Curcumin and Its Bioavailability: Could Its Gastrointestinal Influence Contribute to Its Overall Health-Enhancing Effects? Adv. Nutr. 2018, 9, 41–50. [Google Scholar] [PubMed]
- Metzler, M.; Pfeiffer, E.; Schulz, S.I.; Dempe, J.S. Curcumin uptake and metabolism. Biofactors 2013, 39, 14–20. [Google Scholar] [CrossRef]
- Wu, J.C.; Tsai, M.L.; Lai, C.S.; Wang, Y.J.; Ho, C.T.; Pan, M.H. Chemopreventative effects of tetrahydrocurcumin on human diseases. Food Funct. 2014, 5, 12–17. [Google Scholar]
- Aggarwal, B.B.; Deb, L.; Prasad, S. Curcumin differs from tetrahydrocurcumin for molecular targets, signaling pathways and cellular responses. Molecules 2014, 20, 185–205. [Google Scholar]
- Gutierres, V.O.; Campos, M.L.; Arcaro, C.A.; Assis, R.P.; Baldan-Cimatti, H.M.; Peccinini, R.G.; Paula-Gomes, S.; Kettelhut, I.C.; Baviera, A.M.; Brunetti, I.L. Curcumin Pharmacokinetic and Pharmacodynamic Evidences in Streptozotocin-Diabetic Rats Support the Antidiabetic Activity to Be via Metabolite(s). Evid. Based. Complement. Alternat. Med. 2015, 2015, 678218. [Google Scholar]
- Ryu, E.K.; Choe, Y.S.; Lee, K.H.; Choi, Y.; Kim, B.T. Curcumin and dehydrozingerone derivatives: Synthesis, radiolabeling, and evaluation for beta-amyloid plaque imaging. J. Med. Chem. 2006, 49, 6111–6119. [Google Scholar] [CrossRef]
- Pan, M.H.; Huang, T.M.; Lin, J.K. Biotransformation of curcumin through reduction and glucuronidation in mice. Drug Metab. Dispos. 1999, 27, 486–494. [Google Scholar] [PubMed]
- Holder, G.M.; Plummer, J.L.; Ryan, A.J. The metabolism and excretion of curcumin (1,7-bis-(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione) in the rat. Xenobiotica 1978, 8, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Hassaninasab, A.; Hashimoto, Y.; Tomita-Yokotani, K.; Kobayashi, M. Discovery of the curcumin metabolic pathway involving a unique enzyme in an intestinal microorganism. Proc. Natl. Acad. Sci. USA 2011, 108, 6615–6620. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Qiu, F. Curcuminoid metabolism and its contribution to the pharmacological effects. Curr. Drug Metab. 2013, 14, 791–806. [Google Scholar]
- Naito, M.; Wu, X.; Nomura, H.; Kodama, M.; Kato, Y.; Kato, Y.; Osawa, T. The protective effects of tetrahydrocurcumin on oxidative stress in cholesterol-fed rabbits. J. Atheroscler. Thromb. 2002, 9, 243–250. [Google Scholar]
- Matabudul, D.; Pucaj, K.; Bolger, G.; Vcelar, B.; Majeed, M.; Helson, L. Tissue distribution of (Lipocurc) liposomal curcumin and tetrahydrocurcumin following two-and eight-hour infusions in Beagle dogs. Anticancer Res. 2012, 32, 4359–4364. [Google Scholar]
- Okada, K.; Wangpoengtrakul, C.; Tanaka, T.; Toyokuni, S.; Uchida, K.; Osawa, T. Curcumin and especially tetrahydrocurcumin ameliorate oxidative stress-induced renal injury in mice. J. Nutr. 2001, 131, 2090–2095. [Google Scholar] [CrossRef]
- Begum, A.N.; Jones, M.R.; Lim, G.P.; Morihara, T.; Kim, P.; Heath, D.D.; Rock, C.L.; Pruitt, M.A.; Yang, F.; Hudspeth, B.; et al. Curcumin structure-function, bioavailability, and efficacy in models of neuroinflammation and Alzheimer’s disease. J. Pharmacol. Exp. Ther. 2008, 326, 196–208. [Google Scholar]
- Han, J.S.; Lee, S.; Kim, H.Y.; Lee, C.H. MS-Based Metabolite Profiling of Aboveground and Root Components of Zingiber Mioga and Officinale. Molecules. 2015, 20, 16170–16185. [Google Scholar] [CrossRef]
- Yang, L.; Zhou, C.; Huang, K.; Song, L.; Zheng, Q.; Yu, R.; Zhang, R.; Wu, Y.; Zeng, S.; Cheng, C.H.K.; et al. Antioxidative and cytotoxic properties of diarylheptanoids isolated from Zingiber officinale. Zhongguo Zhong Yao Za Zhi 2009, 34, 319–323. [Google Scholar]
- Somparn, P.; Phisalaphong, C.; Nakornchai, S.; Unchern, S.; Morales, N.P. Comparative antioxidant activities of curcumin and its demethoxy and hydrogenated derivatives. Biol. Pharm. Bull. 2007, 30, 74–78. [Google Scholar] [CrossRef]
- Shimoda, K.; Kubota, N.; Hirano, H.; Matsumoto, M.; Hamada, H.; Hamada, H. Formation of tetrahydrocurcumin by reduction of curcumin with cultured plant cells of Marchantia polymorpha. Nat. Prod. Commun. 2012, 7, 529–530. [Google Scholar] [CrossRef]
- Sato, K.; Iki, N.; Takahashi, T.; Hoshino, H. Evaluation of Stability Against Oxidation and Acid Dissociation Properties for Tetrahydrocurcumin in Aqueous Solution. Bunseki kagaku 2008, 57, 257–263. [Google Scholar] [CrossRef][Green Version]
- Vijaya Saradhi, U.V.; Ling, Y.; Wang, J.; Chiu, M.; Schwartz, E.B.; Fuchs, J.R.; Chan, K.K.; Liu, Z. A liquid chromatography-tandem mass spectrometric method for quantification of curcuminoids in cell medium and mouse plasma. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2010, 878, 3045–3051. [Google Scholar] [CrossRef]
- Novaes, J.T.; Lillico, R.; Sayre, C.L.; Nagabushnam, K.; Majeed, M.; Chen, Y.; Ho, E.A.; Oliveira, A.L.P.; Martinez, S.E.; Alrushaid, S.; et al. Disposition, Metabolism and Histone Deacetylase and Acetyltransferase Inhibition Activity of Tetrahydrocurcumin and Other Curcuminoids. Pharmaceutics 2017, 9, 45. [Google Scholar] [CrossRef]
- Sahebkar, A. Dual effect of curcumin in preventing atherosclerosis: The potential role of pro-oxidant-Antioxidant mechanisms. Nat. Prod. Res. 2015, 29, 491–492. [Google Scholar] [CrossRef]
- Atsumi, T.; Fujisawa, S.; Tonosaki, K. Relationship between intracellular ROS production and membrane mobility in curcumin- and tetrahydrocurcumin-treated human gingival fibroblasts and human submandibular gland carcinoma cells. Oral. Dis. 2005, 11, 236–242. [Google Scholar] [CrossRef]
- Sandur, S.K.; Pandey, M.K.; Sung, B.; Ahn, K.S.; Murakami, A.; Sethi, G.; Limtrakul, P.; Badmaev, V.; Aggarwal, B.B. Curcumin, demethoxycurcumin, bisdemethoxycurcumin, tetrahydrocurcumin and turmerones differentially regulate anti-inflammatory and anti-proliferative responses through a ROS-independent mechanism. Carcinogenesis 2007, 28, 1765–1773. [Google Scholar] [CrossRef]
- Atsumi, T.; Tonosaki, K.; Fujisawa, S. Comparative cytotoxicity and ROS generation by curcumin and tetrahydrocurcumin following visible-light irradiation or treatment with horseradish peroxidase. Anticancer Res. 2007, 27, 363–371. [Google Scholar]
- Sugiyama, Y.; Kawakishi, S.; Osawa, T. Involvement of the beta-diketone moiety in the antioxidative mechanism of tetrahydrocurcumin. Biochem. Pharmacol. 1996, 52, 519–525. [Google Scholar] [CrossRef]
- Pari, L.; Murugan, P. Tetrahydrocurcumin prevents brain lipid peroxidation in streptozotocin-induced diabetic rats. J. Med. Food 2007, 10, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Murugan, P.; Pari, L. Antioxidant effect of tetrahydrocurcumin in streptozotocin-nicotinamide induced diabetic rats. Life Sci. 2006, 79, 1720–1728. [Google Scholar] [CrossRef] [PubMed]
- Khatwani, N.; Adeyeni, T.; Ezekiel, U. The Anti-Proliferative Effects of Curcumin Derivatives, Dimethoxycurcumin, Bisdemethoxycurcumin and Tetrahydrocurcumin, on DLD-1 Colon Cancer Cells. FASEB J. 2016, 30, 1090.6. [Google Scholar]
- Somparna, N.; Kukongviriyapanb, V.; Kukongviriyapanc, U.; Senggunpraib, L.; Prawanb, A. Tetrahydrocurcumin protection against doxorubicin-induced apoptosis. Sci. Asia 2015, 41, 114. [Google Scholar] [CrossRef][Green Version]
- Yoysungnoen, P.; Wirachwong, P.; Changtam, C.; Suksamrarn, A.; Patumraj, S. Anti-cancer and anti-angiogenic effects of curcumin and tetrahydrocurcumin on implanted hepatocellular carcinoma in nude mice. World J. Gastroenterol. 2008, 14, 2003–2009. [Google Scholar] [CrossRef]
- Kitani, K.; Osawa, T.; Yokozawa, T. The effects of tetrahydrocurcumin and green tea polyphenol on the survival of male C57BL/6 mice. Biogerontology 2007, 8, 567–573. [Google Scholar] [CrossRef]
- Zhang, Z.B.; Luo, D.D.; Xie, J.H.; Xian, Y.F.; Lai, Z.Q.; Liu, Y.H.; Liu, W.H.; Chen, J.N.; Lai, X.P.; Lin, Z.X.; et al. Curcumin’s Metabolites, Tetrahydrocurcumin and Octahydrocurcumin, Possess Superior Anti-inflammatory Effects in vivo Through Suppression of TAK1-NF-κB Pathway. Front. Pharmacol. 2018, 9, 1181. [Google Scholar]
- Majeed, M.; Natarajan, S.; Pandey, A.; Bani, S.; Mundkur, L. Subchronic and Reproductive/Developmental Toxicity Studies of Tetrahydrocurcumin in Rats. Toxicol. Res. 2019, 35, 65–74. [Google Scholar] [CrossRef]
- Xiang, L.; Nakamura, Y.; Lim, Y.M.; Yamasaki, Y.; Kurokawa-Nose, Y.; Maruyama, W.; Osawa, T.; Matsuura, A.; Motoyama, N.; Tsuda, L. Tetrahydrocurcumin extends life span and inhibits the oxidative stress response by regulating the FOXO forkhead transcription factor. Aging (Albany N. Y.) 2011, 3, 1098–1109. [Google Scholar] [CrossRef]
- Girija, C.R.; Karunakar, P.; Poojari, C.S.; Begum, N.S.; Syed, A.A. Molecular Docking Studies of Curcumin Derivatives with Multiple Protein Targets for Procarcinogen Activating Enzyme Inhibition. J. Proteom. Bioinform. 2010, 3, 200–203. [Google Scholar] [CrossRef]
- Dileep, K.V.; Tintu, I.; Sadasivan, C. Molecular docking studies of curcumin analogs with phospholipase A2. Interdiscip. Sci. 2011, 3, 189–197. [Google Scholar] [PubMed]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Gogo-Jack, I.; Shaw, A.T. Tumour heterogeneity and resistance to cancer therapies. Nat. Rev. Clin. Oncol. 2018, 15, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Ramon, Y.C.; Sese, M.; Capdevila, C.; Aasen, T.; Mattos-Arruda, L.D.; Diaz-Cano, S.J.; Hernández-Losa, J.; Castellví, J. Clinical implications of intratumor heterogeneity: Challenges and opportunities. J. Mol. Med. (Berlin) 2020, 98, 161–177. [Google Scholar] [CrossRef]
- Liou, G.Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar]
- Strzelczyk, J.K.; Wiczkowski, A. Oxidative damage and carcinogenesis. Contemp. Oncol. (Pozn.) 2012, 16, 230–233. [Google Scholar]
- Morales, N.P.; Sirijaroonwong, S.; Yamanont, P.; Phisalaphong, C. Electron Paramagnetic Resonance Study of the Free Radical Scavenging Capacity of Curcumin and Its Demethoxy and Hydrogenated Derivatives. Biol. Pharm Bull. 2015, 38, 1478–1483. [Google Scholar]
- Suzuki, M.; Nakamura, T.; Iyoki, S.; Fujiwara, A.; Watanabe, Y.; Mohri, K.; Isobe, K.; Ono, K.; Yano, S. Elucidation of anti-allergic activities of curcumin-related compounds with a special reference to their anti-oxidative activities. Biol. Pharm. Bull. 2005, 28, 1438–1443. [Google Scholar]
- Muthumani, M.; Miltonprabu, S. Ameliorative efficacy of tetrahydrocurcumin against arsenic induced oxidative damage, dyslipidemia and hepatic mitochondrial toxicity in rats. Chem. Biol. Interact. 2015, 235, 95–105. [Google Scholar]
- Ramakrishnan, R.; Elangovan, P.; Pari, L. Protective Role of Tetrahydrocurcumin: An Active Polyphenolic Curcuminoid on Cadmium-Induced Oxidative Damage in Rats. Appl. Biochem. Biotechnol. 2017, 183, 51–69. [Google Scholar] [PubMed]
- Dinkova-Kostova, A.T.; Talalay, P. Relation of structure of curcumin analogs to their potencies as inducers of Phase 2 detoxification enzymes. Carcinogenesis 1999, 20, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.D.; Chen, J.F.; Liu, J.J.; Xie, J.H.; Zhang, Z.B.; Gu, J.Y.; Zhuo, J.Y.; Huang, S.; Su, Z.R.; Sun, Z.H. Tetrahydrocurcumin and octahydrocurcumin, the primary and final hydrogenated metabolites of curcumin, possess superior hepatic-protective effect against acetaminophen-induced liver injury: Role of CYP2E1 and Keap1-Nrf2 pathway. Food Chem. Toxicol. 2019, 123, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Ohto, Y.; Murakami, A.; Osawa, T.; Ohigashi, H. Inhibitory effects of curcumin and tetrahydrocurcuminoids on the tumor promoter-induced reactive oxygen species generation in leukocytes in vitro and in vivo. Jpn. J. Cancer Res. 1998, 89, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Nishida, M.; Nishiumi, S.; Mizushina, Y.; Fujishima, Y.; Yamamoto, K.; Masuda, A.; Mizuno, S.; Fujita, T.; Morita, Y.; Kutsumi, H.; et al. Monoacetylcurcumin strongly regulates inflammatory responses through inhibition of NF-κB activation. Int. J. Mol. Med. 2010, 25, 761–767. [Google Scholar] [PubMed]
- Zhao, F.; Gong, Y.; Hu, Y.; Lu, M.; Wang, J.; Dong, J.; Chen, D.; Chen, L.; Fu, F.; Qiu, F. Curcumin and its major metabolites inhibit the inflammatory response induced by lipopolysaccharide: Translocation of nuclear factor-κB as potential target. Mol. Med. Rep. 2015, 11, 3087–3093. [Google Scholar] [CrossRef]
- Yang, J.Y.; Zhong, X.; Kim, S.J.; Kim, D.H.; Kim, H.S.; Lee, J.S.; Yum, H.W.; Lee, J.; Na, H.K.; Surh, Y.J. Comparative Effects of Curcumin and Tetrahydrocurcumin on Dextran Sulfate Sodium-induced Colitis and Inflammatory Signaling in Mice. J. Cancer Prev. 2018, 23, 18–24. [Google Scholar]
- Huang, M.T.; Ma, W.; Lu, Y.P.; Chang, R.L.; Fisher, C.; Manchand, P.S.; Newmark, H.L.; Conney, A.H. Effects of curcumin, demethoxycurcumin, bisdemethoxycurcumin and tetrahydrocurcumin on 12-O-tetradecanoylphorbol-13-acetate-induced tumor promotion. Carcinogenesis 1995, 16, 2493–2497. [Google Scholar] [CrossRef]
- Kim, J.M.; Araki, S.; Kim, D.J.; Park, C.B.; Takasuka, N.; Baba-Toriyama, H.; Ota, T.; Nir, Z.; Khachik, F.; Shimidzu, N.; et al. Chemopreventive effects of carotenoids and curcumins on mouse colon carcinogenesis after 1,2-dimethylhydrazine initiation. Carcinogenesis 1998, 19, 81–85. [Google Scholar]
- Lai, C.S.; Wu, J.C.; Yu, S.F.; Badmaev, V.; Nagabhushanam, K.; Ho, C.T.; Pan, M.H. Tetrahydrocurcumin is more effective than curcumin in preventing azoxymethane-induced colon carcinogenesis. Mol. Nutr. Food Res. 2011, 55, 1819–1828. [Google Scholar]
- Zhang, X.; Peng, L.; Liu, A.; Ji, J.; Zhao, L.; Zhai, G. The enhanced effect of tetrahydrocurcumin on radiosensitivity of glioma cells. J. Pharm. Pharmacol. 2018, 70, 749–759. [Google Scholar] [CrossRef]
- Yodkeeree, S.; Garbisa, S.; Limtrakul, P. Tetrahydrocurcumin inhibits HT1080 cell migration and invasion via downregulation of MMPs and uPA. Acta Pharmacol. Sin. 2008, 29, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Yoysungnoen, B.; Bhattarakosol, P.; Patumraj, S.; Changtam, C. Effects of tetrahydrocurcumin on hypoxia-inducible factor-1α and vascular endothelial growth factor expression in cervical cancer cell-induced angiogenesis in nude mice. Biomed. Res. Int. 2015, 2015, 391748. [Google Scholar] [CrossRef] [PubMed]
- Yoysungnoen, B.; Bhattarakosol, P.; Changtam, C.; Patumraj, S. Effects of Tetrahydrocurcumin on Tumor Growth and Cellular Signaling in Cervical Cancer Xenografts in Nude Mice. Biomed. Res. Int. 2016, 2016, 1781208. [Google Scholar] [PubMed]
- Zhang, Y.; Liu, Y.; Zou, J.; Yan, L.; Du, W.; Zhang, Y.; Sun, H.; Lu, P.; Geng, S.; Gu, R.; et al. Tetrahydrocurcumin induces mesenchymal-epithelial transition and suppresses angiogenesis by targeting HIF-1alpha and autophagy in human osteosarcoma. Oncotarget 2017, 8, 91134–91149. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.; Wang, M.M.; Wang, Y.H.; Zhang, Z.N.; Cao, H.R.; Lv, Y.H.; Yang, Y.; Fan, P.H.; Qiu, F.; Gao, X.M. Tetrahydrocurcumin induces G2/M cell cycle arrest and apoptosis involving p38 MAPK activation in human breast cancer cells. Food Chem. Toxicol. 2014, 67, 193–200. [Google Scholar]
- Han, X.; Deng, S.; Wang, N.; Liu, Y.; Yang, X. Inhibitory effects and molecular mechanisms of tetrahydrocurcumin against human breast cancer MCF-7 cells. Food Nutr. Res. 2016, 60, 30616. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Z.; Lin, G.; Luo, D.; Chen, H.; Yang, H.; Liang, J.; Liu, Y.; Xie, J.; Su, Z.; et al. Tetrahydrocurcumin is more effective than curcumin in inducing the apoptosis of H22 cells via regulation of a mitochondrial apoptosis pathway in ascites tumor-bearing mice. Food Funct. 2017, 8, 3120–3129. [Google Scholar] [CrossRef]
- Wu, J.C.; Lai, C.S.; Badmaev, V.; Nagabhushanam, K.; Ho, C.T.; Pan, M.H. Tetrahydrocurcumin, a major metabolite of curcumin, induced autophagic cell death through coordinative modulation of PI3K/Akt-mTOR and MAPK signaling pathways in human leukemia HL-60 cells. Mol. Nutr. Food Res. 2011, 55, 1646–1654. [Google Scholar] [CrossRef]
- Tseng, Y.H.; Chiou, S.S.; Weng, J.P.; Lin, P.C. Curcumin and tetrahydrocurcumin induce cell death in Ara-C-resistant acute myeloid leukemia. Phytother. Res. 2019, 33, 1199–1207. [Google Scholar]
- Song, G.; Lu, H.; Chen, F.; Wang, Y.; Fan, W.; Shao, W.; Lu, H.; Lin, B. Tetrahydrocurcumin induced autophagy via suppression of PI3K/Akt/mTOR in non-small cell lung carcinoma cells. Mol. Med. Rep. 2018, 17, 5964–5969. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.M.; Kim, H.J.; Kim, S.; Choi, W.H.; Choi, Y.H.; Ryu, S.Y.; Kim, Y.S.; Koh, W.S.; Park, S.Y. Modulation of macrophage functions by compounds isolated from Zingiber officinale. Planta Med. 2009, 75, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Trivedia, K.M.; Parthasarathi, P.; Sethib, K.K.; Gangwarb, M.; Mondal, S.C.; Jana, S. Solid and liquid state characterization of tetrahydrocurcumin using XRPD, FT-IR, DSC, TGA, LC-MS, GC-MS, NMR and its biological activities. J. Pharm. Anal. 2020. [Google Scholar] [CrossRef]
- Borsari, M.; Ferrari, E.; Grandi, M.; Saladini, M. Curcuminoids as potential new iron-chelating agents: Spectroscopic, polarographic and potentiometric study on theirFe(III) complexing ability. Inorg. Chim. Acta 2001, 328, 61–68. [Google Scholar] [CrossRef]
- Guengerich, F.P. Metabolism of chemical carcinogens. Carcinogenesis 2000, 21, 345–351. [Google Scholar]
- Iyanagi, T. Molecular mechanism of phase I and phase II drug-metabolizing enzymes: Implications for detoxification. Int. Rev. Cytol. 2007, 260, 35–112. [Google Scholar]
- Colotta, F.; Allavena, P.; Sica, A.; Garlanda, C.; Mantovani, A. Cancer-related inflammation, the seventh hallmark of cancer: Links to genetic instability. Carcinogenesis 2009, 30, 1073–1081. [Google Scholar] [CrossRef]
- Crusz, S.M.; Balkwill, F.R. Inflammation and cancer: Advances and new agents. Nat. Rev. Clin. Oncol. 2015, 12, 584–596. [Google Scholar]
- Padhye, S.; Chavan, D.; Pandey, S.; Deshpande, J.; Swamy, K.V.; Sarkar, F.H. Perspectives on chemopreventive and therapeutic potential of curcumin analogs in medicinal chemistry. Mini. Rev. Med. Chem. 2010, 10, 372–387. [Google Scholar]
- Bairwa, K.; Grover, J.; Kania, M.; Jachak, S.M. Recent developments in chemistry and biology of curcumin analogues. RSC Adv. 2014, 4, 13946–13978. [Google Scholar] [CrossRef]
- Liang, G.; Shao, L.; Wang, Y.; Zhao, C.; Chu, Y.; Xiao, J.; Zhao, Y.; Li, X.; Yang, S. Exploration and synthesis of curcumin analogues with improved structural stability both in vitro and in vivo as cytotoxic agents. Bioorg. Med. Chem 2009, 17, 2623–2631. [Google Scholar] [PubMed]
- Khudhayer, O.M.; Fakri, M.Y. Curcumin analogs: Synthesis and biological activities. Med. Chem Res. 2020, 29, 479–486. [Google Scholar] [CrossRef]
- Ramayanti, O.; Brinkkemper, M.; Verkuijlen, S.A.W.M.; Ritmaleni, L.; Go, M.L.; Middeldorp, J.M. Curcuminoids as EBV Lytic Activators for Adjuvant Treatment in EBV-Positive Carcinomas. Cancers (Basel) 2018, 10, 89. [Google Scholar]
- Pae, H.O.; Jeong, S.O.; Kim, H.S.; Kim, S.H.; Song, Y.S.; Kim, S.K.; Chai, K.Y.; Chung, H.T. Dimethoxycurcumin, a synthetic curcumin analogue with higher metabolic stability, inhibits NO production, inducible NO synthase expression and NF-kappaB activation in RAW264.7 macrophages activated with LPS. Mol. Nutr. Food Res. 2008, 52, 1082–1091. [Google Scholar] [PubMed]
- Jeong, S.O.; Oh, G.S.; Ha, H.Y.; Soon, K.B.; Kim, H.S.; Kim, Y.C.; Kim, E.C.; Lee, K.M.; Chung, H.T.; Pae, H.O. Dimethoxycurcumin, a Synthetic Curcumin Analogue, Induces Heme Oxygenase-1 Expression through Nrf2 Activation in RAW264.7 Macrophages. J. Clin. Biochem Nutr. 2009, 44, 79–84. [Google Scholar]
- Feitelson, M.A.; Arzumanyan, A.; Kulathinal, R.J.; Blain, S.W.; Holcombe, R.F.; Mahajna, J.; Marino, M.; Martinez-Chantar, M.L.; Nawroth, R.; Sanchez-Garcia, I.; et al. Sustained proliferation in cancer: Mechanisms and novel therapeutic targets. Semin. Cancer Biol. 2015, 35, S25–S54. [Google Scholar] [CrossRef]
- Murray-Stewart, T.R.; Woster, P.M.; Casero, R.A., Jr. Targeting polyamine metabolism for cancer therapy and prevention. Biochem. J. 2016, 473, 2937–2953. [Google Scholar] [CrossRef]
- Bansal, A.; Simon, M.C. Glutathione metabolism in cancer progression and treatment resistance. J. Cell Biol. 2018, 217, 2291–2298. [Google Scholar] [CrossRef]
- Jiang, W.G.; Sanders, A.J.; Katoh, M.; Ungefroren, H.; Gieseler, F.; Prince, M.; Thompson, S.K.; Zollo, M.; Spano, D.; Dhawan, P.; et al. Tissue invasion and metastasis: Molecular, biological and clinical perspectives. Semin. Cancer Biol. 2015, 35, S244–S275. [Google Scholar] [CrossRef]
- Carneiro, B.A.; El-Deiry, W.S. Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 2020, 10, 1038. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, L.; Zhai, G.; Ji, J.; Liu, A. Multifunctional Polyethylene Glycol (PEG)-Poly (Lactic-Co-Glycolic Acid) (PLGA)-Based Nanoparticles Loading Doxorubicin and Tetrahydrocurcumin for Combined Chemoradiotherapy of Glioma. Med. Sci. Monit. 2019, 25, 9737–9751. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.M.M.; Towers, C.G.; Thorburn, A. Targeting autophagy in cancer. Nat. Rev. Cancer 2017, 17, 528–542. [Google Scholar] [CrossRef] [PubMed]
- Limtrakul, P.; Chearwae, W.; Shukla, S.; Phisalphong, C.; Ambudkar, S.V. Modulation of function of three ABC drug transporters, P-glycoprotein (ABCB1), mitoxantrone resistance protein (ABCG2) and multidrug resistance protein 1 (ABCC1) by tetrahydrocurcumin, a major metabolite of curcumin. Mol. Cell Biochem. 2007, 296, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Jiang, W.; Kim, B.Y.S.; Zhang, C.C.; Fu, Y.S.; Weissman, I.L. Phagocytosis checkpoints as new targets for cancer immunotherapy. Nat. Rev. Cancer 2019, 19, 568–586. [Google Scholar] [CrossRef]
- Colombo, A.; Fontani, M.; Dragonetti, C.; Roberto, D.; Williams, J.A.G.; Perrotolo, R.S.; Casagrande, F.; Barozzi, S.; Polo, S. A Highly Luminescent Tetrahydrocurcumin Ir(III) Complex with Remarkable Photoactivated Anticancer Activity. Chemistry. 2019, 25, 7948–7952. [Google Scholar] [CrossRef]
- Mahal, A.; Wu, P.; Jiang, Z.H.; Wei, X. Synthesis and Cytotoxic Activity of Novel Tetrahydrocurcumin Derivatives Bearing Pyrazole Moiety. Nat. Prod. Bioprospect. 2017, 7, 461–469. [Google Scholar] [CrossRef]
- Mahal, A.; Wu, P.; Jiang, Z.H.; Wei, X. Schiff Bases of Tetrahydrocurcumin as Potential Anticancer Agents. Chemistryselect 2019, 4, 366–369. [Google Scholar] [CrossRef]
- Astuti, I.Y.; Suliatin, T.; Wahyuningrum, R. Dissolution enhancement of tetrahydrocurcumin using optimized self-nanoemulsifying drug delivery system. Int. J. App. Pharm. 2019, 11, 97–102. [Google Scholar] [CrossRef]
- Setthacheewakul, S.; Kedjinda, W.; Maneenuan, D.; Wiwattanapatapee, R. Controlled release of oral tetrahydrocurcumin from a novel self-emulsifying floating drug delivery system (SEFDDS). AAPS. Pharm. Sci. Tech. 2011, 12, 152–164. [Google Scholar] [CrossRef]
- Kakkar, V.; Kaur, I.P.; Kaur, A.P.; Saini, K.; Singh, K.K. Topical delivery of tetrahydrocurcumin lipid nanoparticles effectively inhibits skin inflammation: In vitro and in vivo study. Drug Dev. Ind. Pharm. 2018, 44, 1701–1712. [Google Scholar] [CrossRef]
- Plyduang, T.; Lomlim, L.; Yuenyongsawad, S.; Wiwattanapatapee, R. Carboxymethylcellulose-tetrahydrocurcumin conjugates for colon-specific delivery of a novel anti-cancer agent, 4-amino tetrahydrocurcumin. Eur. J. Pharm. Biopharm. 2014, 88, 351–360. [Google Scholar] [PubMed]
- Bhaskar, R.A.; Prasad, E.; Deepthi, S.S.; Haritha, V.; Ramakrishna, S.; Madhusuda, K.; Surekha, M.V.; Venkata Rao, Y.S.R. Wound healing: A new perspective on glucosylated tetrahydrocurcumin. Drug Des. Dev. Ther. 2015, 9, 3579–3588. [Google Scholar]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, C.-S.; Ho, C.-T.; Pan, M.-H. The Cancer Chemopreventive and Therapeutic Potential of Tetrahydrocurcumin. Biomolecules 2020, 10, 831. https://doi.org/10.3390/biom10060831
Lai C-S, Ho C-T, Pan M-H. The Cancer Chemopreventive and Therapeutic Potential of Tetrahydrocurcumin. Biomolecules. 2020; 10(6):831. https://doi.org/10.3390/biom10060831
Chicago/Turabian StyleLai, Ching-Shu, Chi-Tang Ho, and Min-Hsiung Pan. 2020. "The Cancer Chemopreventive and Therapeutic Potential of Tetrahydrocurcumin" Biomolecules 10, no. 6: 831. https://doi.org/10.3390/biom10060831
APA StyleLai, C.-S., Ho, C.-T., & Pan, M.-H. (2020). The Cancer Chemopreventive and Therapeutic Potential of Tetrahydrocurcumin. Biomolecules, 10(6), 831. https://doi.org/10.3390/biom10060831