Galectin-3: Roles in Neurodevelopment, Neuroinflammation, and Behavior

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
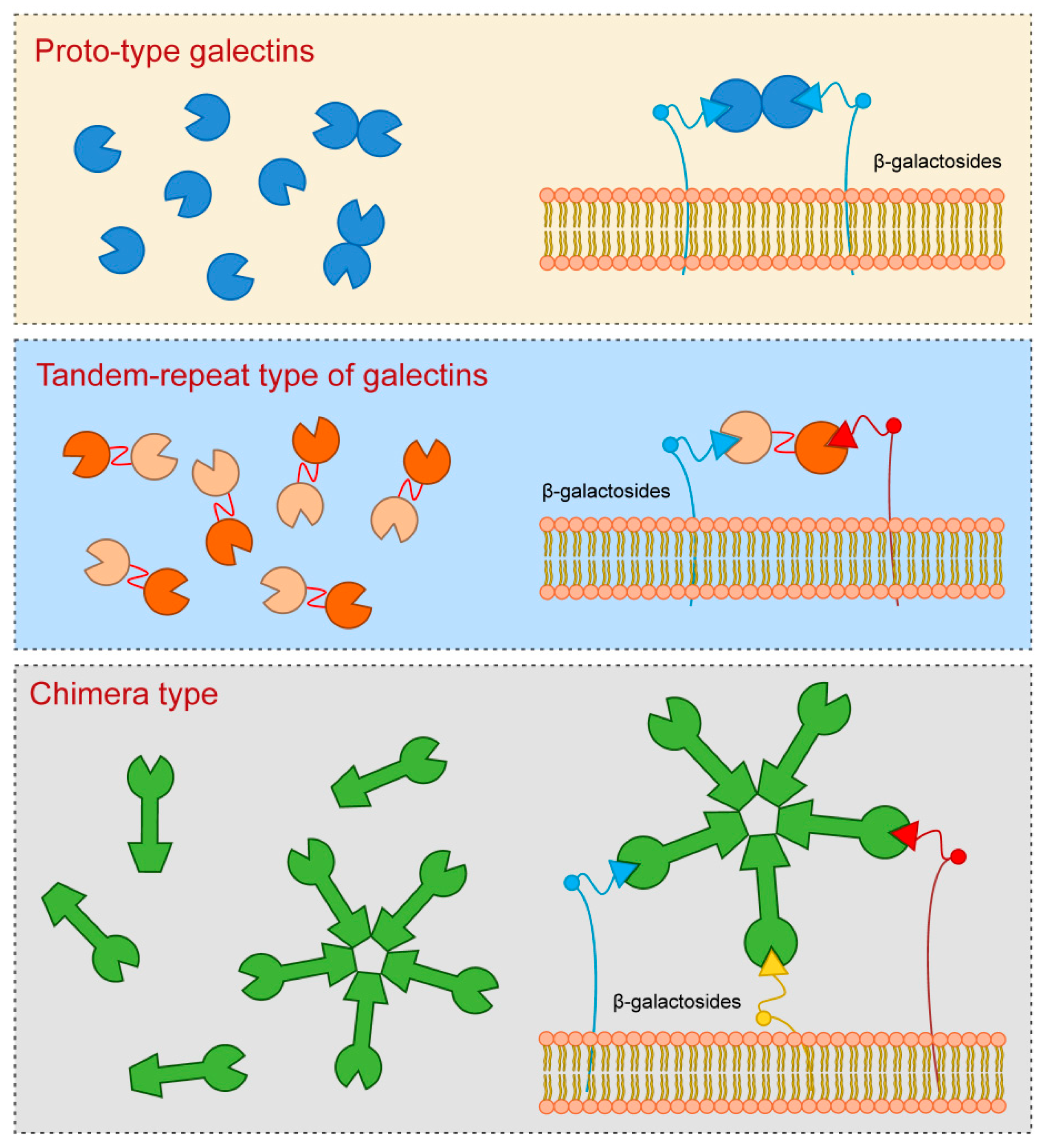
1. Structure of Galectin Proteins
2. Structure and Function of Galectin-3
3. Galectin-3 Ligands and Pattern Recognition Receptors in the CNS
4. Galectin-3 and Matrix Metalloproteinases
5. Galectin-3 and Neurodevelopment
6. Galectin-3 and Neuroinflammation
7. Galectin-3 in the CNS Injury
8. Galectin-3 and the Mechanism of Neurodegeneration
8.1. Prion Disease
8.2. Alzheimer’s Disease
8.3. Parkinson’s Disease
8.4. Huntington’s Disease
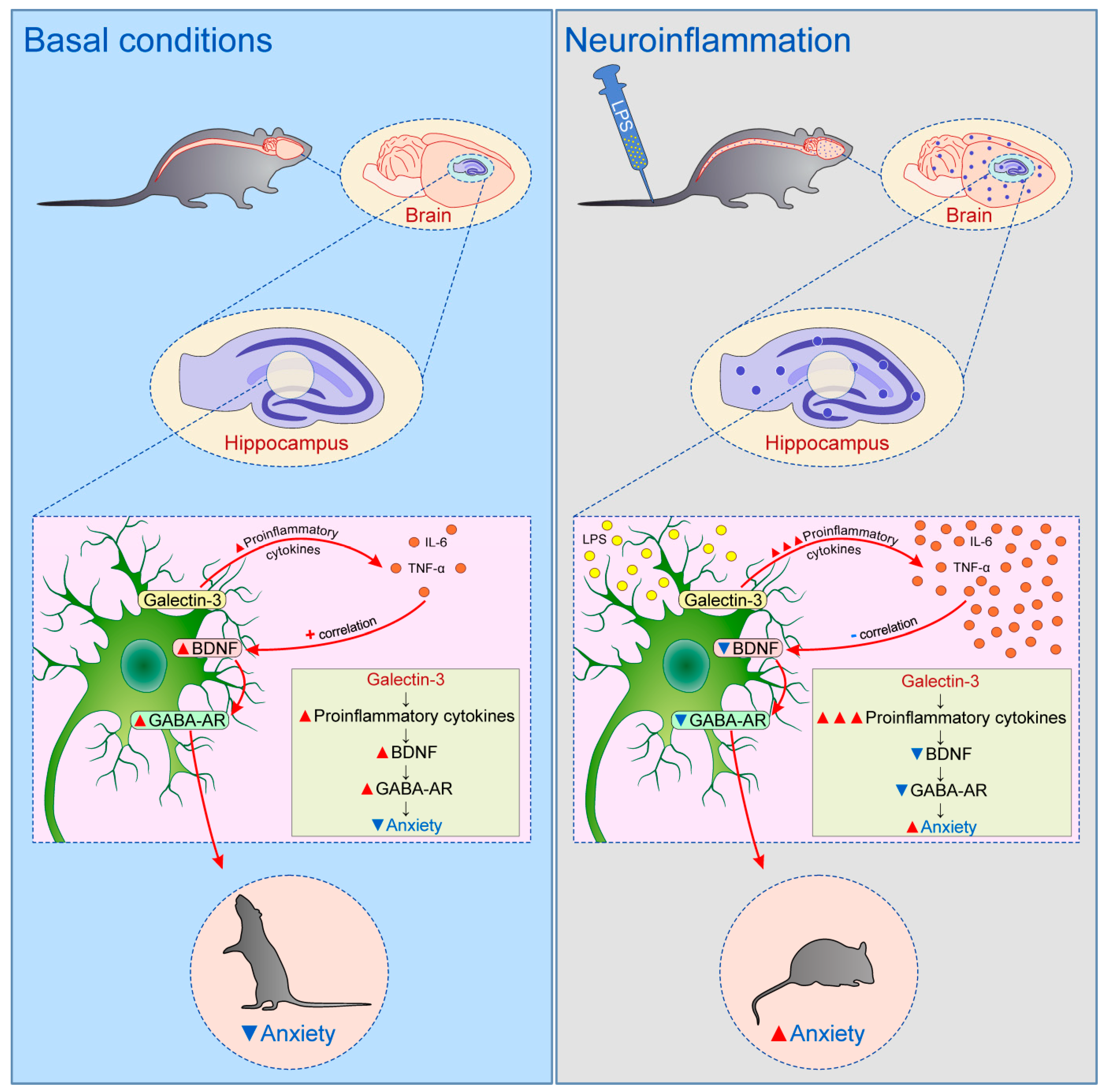
9. Galectin-3 and Behavior
10. Inhibitors of Galectin-3 and Their Potential Therapeutic Use
11. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Barondes, S.H.; Cooper, D.N.; Gitt, M.A.; Leffler, H. Galectins. Structure and function of a large family of animal lectins. J. Biol. Chem. 1994, 269, 20807–20810. [Google Scholar] [PubMed]
- Cooper, D.N. Galectinomics: Finding themes in complexity. Biochim. Biophys. Acta 2002, 1572, 209–231. [Google Scholar] [CrossRef]
- Sciacchitano, S.; Lavra, L.; Morgante, A.; Ulivieri, A.; Magi, F.; De Francesco, G.P.; Bellotti, C.; Salehi, L.B.; Ricci, A. Galectin-3: One Molecule for an Alphabet of Diseases, from A to Z. Int. J. Mol. Sci. 2018, 19, 379. [Google Scholar] [CrossRef]
- Hirabayashi, J.; Kasai, K. The family of metazoan metal–independent beta–galactoside–binding lectins: Structure, function and molecular evolution. Glycobiology 1993, 3, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.Y.; Rabinovich, G.A.; Liu, F.T. Galectins: Structure, function and therapeutic potential. Expert. Rev. Mol. Med. 2008, 10, 17–41. [Google Scholar] [CrossRef] [PubMed]
- Krześlak, A.; Lipińska, A. Galectin-3 as a multifunctional protein. Cell. Mol. Biol. Lett. 2004, 9, 305–328. [Google Scholar]
- Nio–Kobayashi, J. Tissue– and cell–specific localization of galectins, β–galactose–binding animal lectins, and their potential functions in health and disease. Anat. Sci. Int. 2017, 92, 25–36. [Google Scholar] [CrossRef]
- Dumic, J.; Dabelic, S.; Flögel, M. Galectin-3: An open–ended story. Biochim. Biophys. Acta 2006, 1760, 616–635. [Google Scholar] [CrossRef]
- Birdsall, B.; Feeney, J.; Burdett, I.D.; Bawumia, S.; Barboni, E.A.; Hughes, R.C. NMR solution studies of hamster galectin–3 and electron microscopic visualization of surface–adsorbed complexes: Evidence for interactions between the N– and C–terminal domains. Biochemistry 2001, 40, 4859–4866. [Google Scholar] [CrossRef]
- Mazurek, N.; Conklin, J.; Byrd, J.C.; Raz, A.; Bresalier, R.S. Phosphorylation of the beta–galactoside–binding protein galectin–3 modulates binding to its ligands. J. Biol. Chem. 2000, 275, 36311–36315. [Google Scholar] [CrossRef]
- Huflejt, M.E.; Turck, C.W.; Lindstedt, R.; Barondes, S.H.; Leffler, H. L–29, a soluble lactose–binding lectin, is phosphorylated on serine 6 and serine 12 in vivo and by casein kinase I. J. Biol. Chem. 1993, 268, 26712–26718. [Google Scholar] [PubMed]
- Flores–Ibarra, A.; Vértesy, S.; Medrano, F.J.; Gabius, H.J.; Romero, A. Crystallization of a human galectin–3 variant with two ordered segments in the shortened N–terminal tail. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fortuna–Costa, A.; Gomes, A.M.; Kozlowski, E.O.; Stelling, M.P.; Pavão, M.S. Extracellular galectin–3 in tumor progression and metastasis. Front. Oncol. 2014, 4, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Van den Brûle, F.A.; Fernandez, P.L.; Buicu, C.; Liu, F.T.; Jackers, P.; Lambotte, R.; Castronovo, V. Differential expression of galectin–1 and galectin–3 during first trimester human embryogenesis. Dev. Dyn. 1997, 209, 399–405. [Google Scholar] [CrossRef]
- Joo, H.G.; Goedegebuure, P.S.; Sadanaga, N.; Nagoshi, M.; von Bernstorff, W.; Eberlein, T.J. Expression and function of galectin–3, a beta–galactoside–binding protein in activated T lymphocytes. J. Leukoc. Biol. 2001, 69, 555–564. [Google Scholar] [PubMed]
- Di Lella, S.; Sundblad, V.; Cerliani, J.P.; Guardia, C.M.; Estrin, D.A.; Vasta, G.R.; Rabinovich, G.A. When galectins recognize glycans: From biochemistry to physiology and back again. Biochemistry 2011, 50, 7842–7857. [Google Scholar] [CrossRef]
- Liu, F.T.; Rabinovich, G.A. Galectins as modulators of tumour progression. Nat. Rev. Cancer 2005, 5, 29–41. [Google Scholar] [CrossRef]
- Hisrich, B.V.; Young, R.B.; Sansone, A.M.; Bowens, Z.; Green, L.J.; Lessey, B.A.; Blenda, A.V. Role of Human Galectins in Inflammation and Cancers Associated with Endometriosis. Biomolecules 2020, 10, 230. [Google Scholar] [CrossRef]
- Wang, J.L.; Gray, R.M.; Haudek, K.C.; Patterson, R.J. Nucleocytoplasmic lectins. Biochim. Biophys. Acta 2004, 1673, 75–93. [Google Scholar] [CrossRef]
- Hughes, R.C. Secretion of the galectin family of mammalian carbohydrate–binding proteins. Biochim. Biophys. Acta 1999, 1473, 172–185. [Google Scholar] [CrossRef]
- Théry, C.; Boussac, M.; Véron, P.; Ricciardi–Castagnoli, P.; Raposo, G.; Garin, J.; Amigorena, S. Proteomic analysis of dendritic cell–derived exosomes: A secreted subcellular compartment distinct from apoptotic vesicles. J. Immunol. 2001, 166, 7309–7318. [Google Scholar] [CrossRef] [PubMed]
- Mehul, B.; Hughes, R.C. Plasma membrane targetting, vesicular budding and release of galectin 3 from the cytoplasm of mammalian cells during secretion. J. Cell. Sci. 1997, 110, 1169–1178. [Google Scholar] [PubMed]
- Welton, J.L.; Khanna, S.; Giles, P.J.; Brennan, P.; Brewis, I.A.; Staffurth, J.; Mason, M.D.; Clayton, A. Proteomics analysis of bladder cancer exosomes. Mol. Cell. Proteom. 2010, 9, 1324–1338. [Google Scholar] [CrossRef] [PubMed]
- Bänfer, S.; Schneider, D.; Dewes, J.; Strauss, M.T.; Freibert, S.A.; Heimerl, T.; Maier, U.G.; Elsässer, H.P.; Jungmann, R.; Jacob, R. Molecular mechanism to recruit galectin–3 into multivesicular bodies for polarized exosomal secretion. Proc. Natl. Acad. Sci. USA 2018, 115, 4396–4405. [Google Scholar] [CrossRef]
- Arsenijevic, A.; Milovanovic, M.; Milovanovic, J.; Stojanovic, B.; Zdravkovic, N.; Leung, P.S.; Liu, F.T.; Gershwin, M.E.; Lukic, M.L. Deletion of Galectin-3 Enhances Xenobiotic Induced Murine Primary Biliary Cholangitis by Facilitating Apoptosis of BECs and Release of Autoantigens. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef]
- Arsenijevic, A.; Milovanovic, J.; Stojanovic, B.; Djordjevic, D.; Stanojevic, I.; Jankovic, N.; Vojvodic, D.; Arsenijevic, N.; Lukic, M.L.; Milovanovic, M. Gal–3 Deficiency Suppresses Novosphyngobium aromaticivorans Inflammasome Activation and IL–17 Driven Autoimmune Cholangitis in Mice. Front. Immunol. 2019, 10, 1–18. [Google Scholar] [CrossRef]
- Kovacevic, M.M.; Pejnovic, N.; Mitrovic, S.; Jovicic, N.; Petrovic, I.; Arsenijevic, N.; Lukic, M.L.; Ljujic, B. Galectin-3 deficiency enhances type 2 immune cell–mediated myocarditis in mice. Immunol. Res. 2018, 66, 491–502. [Google Scholar] [CrossRef]
- Ahmed, H.; AlSadek, D.M. Galectin-3 as a Potential Target to Prevent Cancer Metastasis. Clin. Med. Insights Oncol. 2015, 9, 113–121. [Google Scholar] [CrossRef]
- Hara, A.; Niwa, M.; Noguchi, K.; Kanayama, T.; Niwa, A.; Matsuo, M.; Hatano, Y.; Tomita, H. Galectin-3 as a Next–Generation Biomarker for Detecting Early Stage of Various Diseases. Biomolecules 2020, 10, 389. [Google Scholar] [CrossRef]
- Yang, R.Y.; Hsu, D.K.; Liu, F.T. Expression of galectin–3 modulates T–cell growth and apoptosis. Proc. Natl. Acad. Sci. USA 1996, 93, 6737–6742. [Google Scholar] [CrossRef]
- Díaz–Alvarez, L.; Ortega, E. The Many Roles of Galectin-3, a Multifaceted Molecule, in Innate Immune Responses against Pathogens. Mediators. Inflamm. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Vasta, G.R. Galectins as pattern recognition receptors: Structure, function, and evolution. Adv. Exp. Med. Biol. 2012, 946, 21–36. [Google Scholar] [PubMed]
- Meylan, E.; Tschopp, J.; Karin, M. Intracellular pattern recognition receptors in the host response. Nature 2006, 442, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; St–Pierre, C.; Bhaumik, P.; Nieminen, J. Galectins in innate immunity: Dual functions of host soluble beta–galactoside–binding lectins as damage–associated molecular patterns (DAMPs) and as receptors for pathogen–associated molecular patterns (PAMPs). Immunol. Rev. 2009, 230, 172–187. [Google Scholar] [CrossRef]
- Mey, A.; Leffler, H.; Hmama, Z.; Normier, G.; Revillard, J.P. The animal lectin galectin–3 interacts with bacterial lipopolysaccharides via two independent sites. J. Immunol. 1996, 156, 1572–1577. [Google Scholar]
- John, C.M.; Jarvis, G.A.; Swanson, K.V.; Leffler, H.; Cooper, M.D.; Huflejt, M.E.; Griffiss, J.M. Galectin-3 binds lactosaminylated lipooligosaccharides from Neisseria gonorrhoeae and is selectively expressed by mucosal epithelial cells that are infected. Cell. Microbiol. 2002, 4, 649–662. [Google Scholar] [CrossRef]
- Li, Y.; Komai–Koma, M.; Gilchrist, D.S.; Hsu, D.K.; Liu, F.T.; Springall, T.; Xu, D. Galectin-3 is a negative regulator of lipopolysaccharide–mediated inflammation. J. Immunol. 2008, 181, 2781–2789. [Google Scholar] [CrossRef]
- Park, A.M.; Hagiwara, S.; Hsu, D.K.; Liu, F.T.; Yoshie, O. Galectin-3 Plays an Important Role in Innate Immunity to Gastric Infection by Helicobacter pylori. Infect. Immun. 2016, 84, 1184–1193. [Google Scholar] [CrossRef]
- Boziki, M.; Polyzos, S.A.; Deretzi, G.; Kazakos, E.; Katsinelos, P.; Doulberis, M.; Kotronis, G.; Giartza–Taxidou, E.; Laskaridis, L.; Tzivras, D.; et al. A potential impact of Helicobacter pylori–related galectin–3 in neurodegeneration. Neurochem. Int. 2018, 113, 137–151. [Google Scholar] [CrossRef]
- Farnworth, S.L.; Henderson, N.C.; Mackinnon, A.C.; Atkinson, K.M.; Wilkinson, T.; Dhaliwal, K.; Hayashi, K.; Simpson, A.J.; Rossi, A.G.; Haslett, C.; et al. Galectin-3 reduces the severity of pneumococcal pneumonia by augmenting neutrophil function. Am. J. Pathol. 2008, 172, 395–405. [Google Scholar] [CrossRef]
- Linden, J.R.; De Paepe, M.E.; Laforce–Nesbitt, S.S.; Bliss, J.M. Galectin-3 plays an important role in protection against disseminated candidiasis. Med. Mycol. 2013, 51, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Kohatsu, L.; Hsu, D.K.; Jegalian, A.G.; Liu, F.T.; Baum, L.G. Galectin-3 induces death of Candida species expressing specific beta–1, 2–linked mannans. J. Immunol. 2006, 177, 4718–4726. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.B.; Li, Q.; Steichen, A.L.; Binstock, B.J.; Metzger, D.W.; Teale, J.M.; Sharma, J. Galectin-3 functions as an alarmin: Pathogenic role for sepsis development in murine respiratory tularemia. PLoS ONE 2013, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Baseras, B.; Gaida, M.M.; Kahle, N.; Schuppel, A.K.; Kathrey, D.; Prior, B.; Wente, M.; Hänsch, G.M. Galectin-3 inhibits the chemotaxis of human polymorphonuclear neutrophils in vitro. Immunobiology 2012, 217, 83–90. [Google Scholar] [CrossRef]
- Bhaumik, P.; St–Pierre, G.; Milot, V.; St–Pierre, C.; Sato, S. Galectin-3 facilitates neutrophil recruitment as an innate immune response to a parasitic protozoa cutaneous infection. J. Immunol. 2013, 190, 630–640. [Google Scholar] [CrossRef]
- Ochieng, J.; Fridman, R.; Nangia–Makker, P.; Kleiner, D.E.; Liotta, L.A.; Stetler–Stevenson, W.G.; Raz, A. Galectin-3 is a novel substrate for human matrix metalloproteinases–2 and –9. Biochemistry 1994, 33, 14109–14114. [Google Scholar] [CrossRef]
- Ochieng, J.; Green, B.; Evans, S.; James, O.; Warfield, P. Modulation of the biological functions of galectin–3 by matrix metalloproteinases. Biochim. Biophys. Acta 1998, 1379, 97–106. [Google Scholar] [CrossRef]
- Takenaka, Y.; Fukumori, T.; Raz, A. Galectin-3 and metastasis. Glycoconj. J. 2002, 19, 543–549. [Google Scholar] [CrossRef]
- Wang, Y.G.; Kim, S.J.; Baek, J.H.; Lee, H.W.; Jeong, S.Y.; Chun, K.H. Galectin-3 increases the motility of mouse melanoma cells by regulating matrix metalloproteinase–1 expression. Exp. Mol. Med. 2012, 44, 387–393. [Google Scholar] [CrossRef]
- Kim, S.J.; Shin, J.Y.; Lee, K.D.; Bae, Y.K.; Choi, I.J.; Park, S.H.; Chun, K.H. Galectin-3 facilitates cell motility in gastric cancer by up–regulating protease–activated receptor–1 (PAR–1) and matrix metalloproteinase–1 (MMP–1). PLoS ONE 2011, 6, 1–10. [Google Scholar] [CrossRef]
- Eisa, N.H.; Ebrahim, M.A.; Ragab, M.; Eissa, L.A.; El–Gayar, A.M. Galectin-3 and matrix metalloproteinase–9: Perspective in management of hepatocellular carcinoma. J. Oncol. Pharm. Pract. 2015, 21, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Mauris, J.; Woodward, A.M.; Cao, Z.; Panjwani, N.; Argüeso, P. Molecular basis for MMP9 induction and disruption of epithelial cell–cell contacts by galectin–3. J. Cell. Sci. 2014, 127, 3141–3148. [Google Scholar] [CrossRef] [PubMed]
- Svedin, P.; Hagberg, H.; Sävman, K.; Zhu, C.; Mallard, C. Matrix metalloproteinase–9 gene knock–out protects the immature brain after cerebral hypoxia–ischemia. J. Neurosci. 2007, 27, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Doverhag, C.; Hedtjärn, M.; Poirier, F.; Mallard, C.; Hagberg, H.; Karlsson, A.; Sävman, K. Galectin-3 contributes to neonatal hypoxic–ischemic brain injury. Neurobiol. Dis. 2010, 38, 36–46. [Google Scholar] [CrossRef]
- Mensah–Brown, E.P.; Al Rabesi, Z.; Shahin, A.; Al Shamsi, M.; Arsenijevic, N.; Hsu, D.K.; Liu, F.T.; Lukic, M.L. Targeted disruption of the galectin–3 gene results in decreased susceptibility to multiple low dose streptozotocin–induced diabetes in mice. Clin. Immunol. 2009, 130, 83–88. [Google Scholar] [CrossRef]
- Mensah–Brown, E.; Shahin, A.; Parekh, K.; Hakim, A.A.; Shamisi, M.A.; Hsu, D.K.; Lukic, M.L. Functional capacity of macrophages determines the induction of type 1 diabetes. Ann. N. Y. Acad. Sci. 2006, 1084, 49–57. [Google Scholar] [CrossRef]
- Volarevic, V.; Markovic, B.S.; Jankovic, M.G.; Djokovic, B.; Jovicic, N.; Harrell, C.R.; Fellabaum, C.; Djonov, V.; Arsenijevic, N.; Lukic, M.L. Galectin 3 protects from cisplatin–induced acute kidney injury by promoting TLR–2–dependent activation of IDO1/Kynurenine pathway in renal DCs. Theranostics 2019, 9, 5976–6001. [Google Scholar] [CrossRef]
- Jovanovic, M.; Gajovic, N.; Zdravkovic, N.; Jovanovic, M.; Jurisevic, M.; Vojvodic, D.; Maric, V.; Arsenijevic, A.; Jovanovic, I. Fecal Galectin-3: A New Promising Biomarker for Severity and Progression of Colorectal Carcinoma. Mediators. Inflamm. 2018, 2018, 1–11. [Google Scholar] [CrossRef]
- Radosavljevic, G.; Volarevic, V.; Jovanovic, I.; Milovanovic, M.; Pejnovic, N.; Arsenijevic, N.; Hsu, D.K.; Lukic, M.L. The roles of Galectin-3 in autoimmunity and tumor progression. Immunol. Res. 2012, 52, 100–110. [Google Scholar] [CrossRef]
- Pejnovic, N.; Jeftic, I.; Jovicic, N.; Arsenijevic, N.; Lukic, M.L. Galectin-3 and IL–33/ST2 axis roles and interplay in diet–induced steatohepatitis. World J. Gastroenterol. 2016, 22, 9706–9717. [Google Scholar] [CrossRef]
- Li, Y.S.; Li, X.T.; Yu, L.G.; Wang, L.; Shi, Z.Y.; Guo, X.L. Roles of galectin–3 in metabolic disorders and tumor cell metabolism. Int. J. Biol. Macromol. 2020, 142, 463–473. [Google Scholar] [PubMed]
- Yoo, H.I.; Kim, E.G.; Lee, E.J.; Hong, S.Y.; Yoon, C.S.; Hong, M.J.; Park, S.J.; Woo, R.S.; Baik, T.K.; Song, D.Y. Neuroanatomical distribution of galectin–3 in the adult rat brain. J. Mol. Histol. 2017, 48, 133–146. [Google Scholar] [PubMed]
- Comte, I.; Kim, Y.; Young, C.C.; van der Harg, J.M.; Hockberger, P.; Bolam, P.J.; Poirier, F.; Szele, F.G. Galectin-3 maintains cell motility from the subventricular zone to the olfactory bulb. J. Cell. Sci. 2011, 124, 2438–2447. [Google Scholar] [PubMed]
- Partridge, E.A.; Le Roy, C.; Di Guglielmo, G.M.; Pawling, J.; Cheung, P.; Granovsky, M.; Nabi, I.R.; Wrana, J.L.; Dennis, J.W. Regulation of cytokine receptors by Golgi N–glycan processing and endocytosis. Science 2004, 306, 120–124. [Google Scholar] [PubMed]
- Liu, W.; Hsu, D.K.; Chen, H.Y.; Yang, R.Y.; Carraway, K.L., 3rd; Isseroff, R.R.; Liu, F.T. Galectin-3 regulates intracellular trafficking of EGFR through Alix and promotes keratinocyte migration. J. Invest. Dermatol. 2012, 132, 2828–2837. [Google Scholar]
- Pasquini, L.A.; Millet, V.; Hoyos, H.C.; Giannoni, J.P.; Croci, D.O.; Marder, M.; Liu, F.T.; Rabinovich, G.A.; Pasquini, J.M. Galectin-3 drives oligodendrocyte differentiation to control myelin integrity and function. Cell Death Differ. 2011, 18, 1746–1756. [Google Scholar]
- Thomas, L.; Pasquini, L.A. Galectin-3–Mediated Glial Crosstalk Drives Oligodendrocyte Differentiation and (Re)myelination. Front. Cell. Neurosci. 2018, 12, 297–313. [Google Scholar] [CrossRef]
- Lee, A.C.; Kozuki, N.; Blencowe, H.; Vos, T.; Bahalim, A.; Darmstadt, G.L.; Niermeyer, S.; Ellis, M.; Robertson, N.J.; Cousens, S.; et al. Intrapartum–related neonatal encephalopathy incidence and impairment at regional and global levels for 2010 with trends from 1990. Pediatr. Res. 2013, 74, 50–72. [Google Scholar] [CrossRef]
- Schiering, I.A.; de Haan, T.R.; Niermeijer, J.M.; Koelman, J.H.; Majoie, C.B.; Reneman, L.; Aronica, E. Correlation between clinical and histologic findings in the human neonatal hippocampus after perinatal asphyxia. J. Neuropathol. Exp. Neurol. 2014, 73, 324–334. [Google Scholar]
- Vlassaks, E.; Brudek, T.; Pakkenberg, B.; Gavilanes, A.W. Cerebellar cytokine expression in a rat model for fetal asphyctic preconditioning and perinatal asphyxia. Cerebellum 2014, 13, 471–478. [Google Scholar] [CrossRef]
- Torres–Cuevas, I.; Corral–Debrinski, M.; Gressens, P. Brain oxidative damage in murine models of neonatal hypoxia/ischemia and reoxygenation. Free Radic. Biol. Med. 2019, 142, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Lalancette–Hébert, M.; Swarup, V.; Beaulieu, J.M.; Bohacek, I.; Abdelhamid, E.; Weng, Y.C.; Sato, S.; Kriz, J. Galectin-3 is required for resident microglia activation and proliferation in response to ischemic injury. J. Neurosci. 2012, 32, 10383–10395. [Google Scholar] [CrossRef] [PubMed]
- Rahimian, R.; Béland, L.C.; Kriz, J. Galectin-3: Mediator of microglia responses in injured brain. Drug. Discov. Today 2018, 23, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Doverhag, C.; Keller, M.; Karlsson, A.; Hedtjarn, M.; Nilsson, U.; Kapeller, E.; Sarkozy, G.; Klimaschewski, L.; Humpel, C.; Hagberg, H.; et al. Pharmacological and genetic inhibition of NADPH oxidase does not reduce brain damage in different models of perinatal brain injury in newborn mice. Neurobiol. Dis. 2008, 31, 133–144. [Google Scholar]
- Pesheva, P.; Kuklinski, S.; Biersack, H.J.; Probstmeier, R. Nerve growth factor–mediated expression of galectin–3 in mouse dorsal root ganglion neurons. Neurosci. Lett. 2000, 293, 37–40. [Google Scholar] [CrossRef]
- Pesheva, P.; Kuklinski, S.; Schmitz, B.; Probstmeier, R. Galectin-3 promotes neural cell adhesion and neurite growth. J. Neurosci. Res. 1998, 54, 639–654. [Google Scholar] [CrossRef]
- Kuklinski, S.; Vladimirova, V.; Waha, A.; Kamata, H.; Pesheva, P.; Probstmeier, R. Expression of galectin–3 in neuronally differentiating PC12 cells is regulated both via Ras/MAPK–dependent and –independent signalling pathways. J. Neurochem. 2003, 87, 1112–1124. [Google Scholar]
- Umekawa, T.; Osman, A.M.; Han, W.; Ikeda, T.; Blomgren, K. Resident microglia, rather than blood–derived macrophages, contribute to the earlier and more pronounced inflammatory reaction in the immature compared with the adult hippocampus after hypoxia–ischemia. Glia 2015, 63, 2220–2230. [Google Scholar] [CrossRef]
- Chip, S.; Fernández–López, D.; Li, F.; Faustino, J.; Derugin, N.; Vexler, Z.S. Genetic deletion of galectin–3 enhances neuroinflammation, affects microglial activation and contributes to sub–chronic injury in experimental neonatal focal stroke. Brain. Behav. Immun. 2017, 60, 270–281. [Google Scholar]
- Al–Dalahmah, O.; Campos Soares, L.; Nicholson, J.; Draijer, S.; Mundim, M.; Lu, V.M.; Sun, B.; Tyler, T.; Adorján, I.; O’Neill, E.; et al. Galectin-3 modulates postnatal subventricular zone gliogenesis. Glia 2020, 68, 435–450. [Google Scholar]
- Stajic, D.; Selakovic, D.; Jovicic, N.; Joksimovic, J.; Arsenijevic, N.; Lukic, M.L.; Rosic, G. The role of galectin–3 in modulation of anxiety state level in mice. Brain. Behav. Immun. 2019, 78, 177–187. [Google Scholar] [CrossRef]
- Shin, T. The pleiotropic effects of galectin–3 in neuroinflammation: A review. Acta. Histochem. 2013, 115, 407–411. [Google Scholar] [CrossRef]
- Steinman, L.; Zamvil, S.S. How to successfully apply animal studies in experimental allergic encephalomyelitis to research on multiple sclerosis. Ann. Neurol. 2006, 60, 12–21. [Google Scholar] [CrossRef]
- Jiang, H.R.; Al Rasebi, Z.; Mensah–Brown, E.; Shahin, A.; Xu, D.; Goodyear, C.S.; Fukada, S.Y.; Liu, F.T.; Liew, F.Y.; Lukic, M.L. Galectin-3 deficiency reduces the severity of experimental autoimmune encephalomyelitis. J. Immunol. 2009, 182, 1167–1173. [Google Scholar] [CrossRef]
- Wolf, S.A.; Boddeke, H.W.; Kettenmann, H. Microglia in Physiology and Disease. Annu. Rev. Physiol. 2017, 79, 619–643. [Google Scholar]
- Reichert, F.; Rotshenker, S. Galectin-3/MAC–2 in experimental allergic encephalomyelitis. Exp. Neurol. 1999, 160, 508–514. [Google Scholar] [CrossRef]
- Rotshenker, S.; Reichert, F.; Gitik, M.; Haklai, R.; Elad–Sfadia, G.; Kloog, Y. Galectin-3/MAC–2, Ras and PI3K activate complement receptor–3 and scavenger receptor–AI/II mediated myelin phagocytosis in microglia. Glia 2008, 56, 1607–1613. [Google Scholar]
- Rotshenker, S. The role of Galectin-3/MAC–2 in the activation of the innate–immune function of phagocytosis in microglia in injury and disease. J. Mol. Neurosci. 2009, 39, 99–103. [Google Scholar] [CrossRef]
- Rotshenker, S. Wallerian degeneration: The innate–immune response to traumatic nerve injury. J. Neuroinflam. 2011, 8, 109. [Google Scholar]
- Mietto, B.S.; Jurgensen, S.; Alves, L.; Pecli, C.; Narciso, M.S.; Assunção-Miranda, I.; Villa–Verde, D.M.; de Souza Lima, F.R.; de Menezes, J.R.; Benjamim, C.F.; et al. Lack of galectin–3 speeds Wallerian degeneration by altering TLR and pro–inflammatory cytokine expressions in injured sciatic nerve. Eur. J. Neurosci. 2013, 37, 1682–1690. [Google Scholar]
- Chaudhary, P.; Marracci, G.; Galipeau, D.; Pocius, E.; Morris, B.; Bourdette, D. Lipoic acid reduces inflammation in a mouse focal cortical experimental autoimmune encephalomyelitis model. J. Neuroimmunol. 2015, 289, 68–74. [Google Scholar]
- Itabashi, T.; Arima, Y.; Kamimura, D.; Higuchi, K.; Bando, Y.; Takahashi–Iwanaga, H.; Murakami, M.; Watanabe, M.; Iwanaga, T.; Nio–Kobayashi, J. Cell– and stage–specific localization of galectin–3, a β–galactoside–binding lectin, in a mouse model of experimental autoimmune encephalomyelitis. Neurochem. Int. 2018, 118, 176–184. [Google Scholar] [CrossRef]
- Petrovic, I.; Pejnovic, N.; Ljujic, B.; Pavlovic, S.; Miletic Kovacevic, M.; Jeftic, I.; Djukic, A.; Draginic, N.; Andjic, M.; Arsenijevic, N.; et al. Overexpression of Galectin 3 in Pancreatic β Cells Amplifies β–Cell Apoptosis and Islet Inflammation in Type–2 Diabetes in Mice. Front. Endocrinol. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Jaquenod De Giusti, C.; Alberdi, L.; Frik, J.; Ferrer, M.F.; Scharrig, E.; Schattner, M.; Gomez, R.M. Galectin-3 is upregulated in activated glia during Junin virus–induced murine encephalitis. Neurosci. Lett. 2011, 501, 163–166. [Google Scholar]
- James, R.E.; Hillis, J.; Adorján, I.; Gration, B.; Mundim, M.V.; Iqbal, A.J.; Majumdar, M.M.; Yates, R.L.; Richards, M.M.; Goings, G.E.; et al. Loss of galectin–3 decreases the number of immune cells in the subventricular zone and restores proliferation in a viral model of multiple sclerosis. Glia 2016, 64, 105–121. [Google Scholar] [CrossRef]
- Venkatraman, A.; Hardas, S.; Patel, N.; Singh Bajaj, N.; Arora, G.; Arora, P. Galectin-3: An emerging biomarker in stroke and cerebrovascular diseases. Eur. J. Neurol. 2018, 25, 238–246. [Google Scholar] [CrossRef]
- Yan, Y.P.; Lang, B.T.; Vemuganti, R.; Dempsey, R.J. Galectin-3 mediates post–ischemic tissue remodeling. Brain. Res. 2009, 1288, 116–124. [Google Scholar] [CrossRef]
- Rahimian, R.; Lively, S.; Abdelhamid, E.; Lalancette–Hebert, M.; Schlichter, L.; Sato, S.; Kriz, J. Delayed Galectin-3–Mediated Reprogramming of Microglia After Stroke is Protective. Mol. Neurobiol. 2019, 56, 6371–6385. [Google Scholar] [CrossRef]
- Satoh, K.; Niwa, M.; Goda, W.; Binh, N.H.; Nakashima, M.; Takamatsu, M.; Hara, A. Galectin-3 expression in delayed neuronal death of hippocampal CA1 following transient forebrain ischemia, and its inhibition by hypothermia. Brain. Res. 2011, 1382, 266–274. [Google Scholar]
- Cheng, X.; Boza–Serrano, A.; Turesson, M.F.; Deierborg, T.; Ekblad, E.; Voss, U. Galectin-3 causes enteric neuronal loss in mice after left sided permanent middle cerebral artery occlusion, a model of stroke. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef]
- Deierborg, T.; Burguillos, M.A. A new “sweet” ligand for Toll–like receptor 4. Oncotarget 2015, 6, 19928–19929. [Google Scholar] [CrossRef]
- Burguillos, M.A.; Svensson, M.; Schulte, T.; Boza–Serrano, A.; Garcia–Quintanilla, A.; Kavanagh, E.; Santiago, M.; Viceconte, N.; Oliva–Martin, M.J.; Osman, A.M.; et al. Galectin-3 Acts as a Toll–like Receptor 4 Ligand and Contributes to Microglial Activation. Cell. Rep. 2015, 10, 1626–1630. [Google Scholar] [CrossRef]
- Wu, Z.S.; Lo, J.J.; Wu, S.H.; Wang, C.Z.; Chen, R.F.; Lee, S.S.; Chai, C.Y.; Huang, S.H. Early Hyperbaric Oxygen Treatment Attenuates Burn–Induced Neuroinflammation by Inhibiting the Galectin-3–Dependent Toll–Like Receptor–4 Pathway in a Rat Model. Int. J. Mol. Sci. 2018, 19, 2195. [Google Scholar] [CrossRef]
- Stojanovic, B.; Jovanovic, I.; Stojanovic, B.S.; Stojanovic, M.D.; Gajovic, N.; Radosavljevic, G.; Pantic, J.; Arsenijevic, N.; Lukic, M.L. Deletion of Galectin-3 attenuates acute pancreatitis in mice by affecting activation of innate inflammatory cells. Eur. J. Immunol. 2019, 49, 940–946. [Google Scholar] [CrossRef]
- Nomura, K.; Vilalta, A.; Allendorf, D.H.; Hornik, T.C.; Brown, G.C. Activated Microglia Desialylate and Phagocytose Cells via Neuraminidase, Galectin-3, and Mer Tyrosine Kinase. J. Immunol. 2017, 198, 4792–4801. [Google Scholar] [CrossRef]
- Caberoy, N.B.; Alvarado, G.; Bigcas, J.L.; Li, W. Galectin-3 is a new MerTK–specific eat–me signal. J. Cell. Physiol. 2012, 227, 401–407. [Google Scholar] [CrossRef]
- Yip, P.K.; Carrillo–Jimenez, A.; King, P.; Vilalta, A.; Nomura, K.; Chau, C.C.; Egerton, A.M.; Liu, Z.H.; Shetty, A.J.; Tremoleda, J.L.; et al. Galectin-3 released in response to traumatic brain injury acts as an alarmin orchestrating brain immune response and promoting neurodegeneration. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Shen, Y.F.; Yu, W.H.; Dong, X.Q.; Du, Q.; Yang, D.B.; Wu, G.Q.; Zhang, Z.Y.; Wang, H.; Jiang, L. The change of plasma galectin–3 concentrations after traumatic brain injury. Clin. Chim. Acta 2016, 456, 75–80. [Google Scholar] [CrossRef]
- Shan, R.; Szmydynger–Chodobska, J.; Warren, O.U.; Mohammad, F.; Zink, B.J.; Chodobski, A. A New Panel of Blood Biomarkers for the Diagnosis of Mild Traumatic Brain Injury/Concussion in Adults. J. Neurotrauma 2016, 33, 49–57. [Google Scholar] [CrossRef]
- Byrnes, K.R.; Washington, P.M.; Knoblach, S.M.; Hoffman, E.; Faden, A.I. Delayed inflammatory mRNA and protein expression after spinal cord injury. J. Neuroinflam. 2011, 8, 130–141. [Google Scholar] [CrossRef]
- Pajoohesh–Ganji, A.; Knoblach, S.M.; Faden, A.I.; Byrnes, K.R. Characterization of inflammatory gene expression and galectin–3 function after spinal cord injury in mice. Brain. Res. 2012, 1475, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Mostacada, K.; Oliveira, F.L.; Villa–Verde, D.M.; Martinez, A.M. Lack of galectin–3 improves the functional outcome and tissue sparing by modulating inflammatory response after a compressive spinal cord injury. Exp. Neurol. 2015, 271, 390–400. [Google Scholar] [CrossRef] [PubMed]
- David, S.; Kroner, A. Repertoire of microglial and macrophage responses after spinal cord injury. Nat. Rev. Neurosci. 2011, 12, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Mok, S.W.; Riemer, C.; Madela, K.; Hsu, D.K.; Liu, F.T.; Gültner, S.; Heise, I.; Baier, M. Role of galectin–3 in prion infections of the CNS. Biochem. Biophys. Res. Commun. 2007, 359, 672–678. [Google Scholar] [CrossRef]
- Šimić, G.; Babić Leko, M.; Wray, S.; Harrington, C.; Delalle, I.; Jovanov–Milošević, N.; Bažadona, D.; Buée, L.; de Silva, R.; Di Giovanni, G.; et al. Protein Hyperphosphorylation and Aggregation in Alzheimer’s Disease and Other Tauopathies, and Possible Neuroprotective Strategies. Biomolecules 2016, 6, 6. [Google Scholar] [CrossRef]
- Boza–Serrano, A.; Ruiz, R.; Sanchez–Varo, R.; García–Revilla, J.; Yang, Y.; Jimenez–Ferrer, I.; Paulus, A.; Wennström, M.; Vilalta, A.; Allendorf, D.; et al. Galectin-3, a novel endogenous TREM2 ligand, detrimentally regulates inflammatory response in Alzheimer’s disease. Acta Neuropathol. 2019, 138, 251–273. [Google Scholar] [CrossRef]
- Tao, C.C.; Cheng, K.M.; Ma, Y.L.; Hsu, W.L.; Chen, Y.C.; Fuh, J.L.; Lee, W.J.; Chao, C.C.; Lee, E.H.Y. Galectin-3 promotes Aβ oligomerization and Aβ toxicity in a mouse model of Alzheimer’s disease. Cell Death Differ. 2020, 27, 192–209. [Google Scholar] [CrossRef]
- Beitz, J.M. Parkinson’s disease: A review. Front. Biosci. (Schol. Ed.) 2014, 6, 65–74. [Google Scholar] [CrossRef]
- Boza–Serrano, A.; Reyes, J.F.; Rey, N.L.; Leffler, H.; Bousset, L.; Nilsson, U.; Brundin, P.; Venero, J.L.; Burguillos, M.A.; Deierborg, T. The role of Galectin-3 in α–synuclein–induced microglial activation. Acta Neuropathol. Commun. 2014, 2, 156. [Google Scholar]
- Yazar, H.O.; Yazar, T.; Cihan, M. A preliminary data: Evaluation of serum Galectin-3 levels in patients with Idiopathic Parkinson’s Disease. J. Clin. Neurosci. 2019, 70, 164–168. [Google Scholar] [CrossRef]
- Siew, J.J.; Chen, H.M.; Chen, H.Y.; Chen, H.L.; Chen, C.M.; Soong, B.W.; Wu, Y.R.; Chang, C.P.; Chan, Y.C.; Lin, C.H.; et al. Galectin-3 is required for the microglia–mediated brain inflammation in a model of Huntington’s disease. Nat. Commun. 2019, 10, 1–18. [Google Scholar]
- Chaudoin, T.R.; Bonasera, S.J. Mice lacking galectin–3 (Lgals3) function have decreased home cage movement. BMC Neurosci. 2018, 19, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Orr, S.L.; Le, D.; Long, J.M.; Sobieszczuk, P.; Ma, B.; Tian, H.; Fang, X.; Paulson, J.C.; Marth, J.D.; Varki, N. A phenotype survey of 36 mutant mouse strains with gene–targeted defects in glycosyltransferases or glycan–binding proteins. Glycobiology 2013, 23, 363–380. [Google Scholar] [CrossRef] [PubMed]
- Trompet, S.; Jukema, W.; Mooijaart, S.P.; Ford, I.; Stott, D.J.; Westendorp, R.G.; de Craen, A.J. Genetic variation in galectin–3 gene associates with cognitive function at old age. Neurobiol. Aging 2012, 33, 2232.e1–2232.e9. [Google Scholar] [CrossRef]
- Torkildsen, O.; Brunborg, L.A.; Myhr, K.M.; Bø, L. The cuprizone model for demyelination. Acta Neurol. Scand. Suppl. 2008, 188, 72–76. [Google Scholar] [CrossRef]
- Ramírez, E.; Sánchez–Maldonado, C.; Mayoral, M.A.; Mendieta, L.; Alatriste, V.; Patricio–Martínez, A.; Limón, I.D. Neuroinflammation induced by the peptide amyloid–β (25–35) increase the presence of galectin–3 in astrocytes and microglia and impairs spatial memory. Neuropeptides 2019, 74, 11–23. [Google Scholar]
- Müller, N. Inflammation in Schizophrenia: Pathogenetic Aspects and Therapeutic Considerations. Schizophr. Bull. 2018, 44, 973–982. [Google Scholar] [CrossRef]
- Kajitani, K.; Yanagimoto, K.; Nakabeppu, Y. Serum galectin–3, but not galectin–1, levels are elevated in schizophrenia: Implications for the role of inflammation. Psychopharmacology (Berl.) 2017, 234, 2919–2927. [Google Scholar] [CrossRef]
- Borovcanin, M.M.; Janicijevic, S.M.; Jovanovic, I.P.; Gajovic, N.; Arsenijevic, N.N.; Lukic, M.L. IL–33/ST2 Pathway and Galectin-3 as a New Analytes in Pathogenesis and Cardiometabolic Risk Evaluation in Psychosis. Front. Psychiatry 2018, 9, 271–280. [Google Scholar] [CrossRef]
- Woelfer, M.; Kasties, V.; Kahlfuss, S.; Walter, M. The Role of Depressive Subtypes within the Neuroinflammation Hypothesis of Major Depressive Disorder. Neuroscience 2019, 403, 93–110. [Google Scholar] [CrossRef]
- Melin, E.O.; Dereke, J.; Thunander, M.; Hillman, M. Depression in type 1 diabetes was associated with high levels of circulating galectin–3. Endocr. Connect. 2018, 7, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Kantak, K.M.; Singh, T.; Kerstetter, K.A.; Dembro, K.A.; Mutebi, M.M.; Harvey, R.C.; Deschepper, C.F.; Dwoskin, L.P. Advancing the spontaneous hypertensive rat model of attention deficit/hyperactivity disorder. Behav. Neurosci. 2008, 122, 340–357. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhao, Q.; Zhu, X.; Peng, M.; Jia, C.; Wu, W.; Zheng, J.; Wu, X.Z. A novel function of microRNA let–7d in regulation of galectin–3 expression in attention deficit hyperactivity disorder rat brain. Brain Pathol. 2010, 20, 1042–1054. [Google Scholar] [PubMed]
- Wu, L.H.; Peng, M.; Yu, M.; Zhao, Q.L.; Li, C.; Jin, Y.T.; Jiang, Y.; Chen, Z.Y.; Deng, N.H.; Sun, H.; et al. Circulating MicroRNA Let–7d in Attention–Deficit/Hyperactivity Disorder. Neuromolecular. Med. 2015, 17, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Stegmayr, J.; Zetterberg, F.; Carlsson, M.C.; Huang, X.; Sharma, G.; Kahl–Knutson, B.; Schambye, H.; Nilsson, U.J.; Oredsson, S.; Leffler, H. Extracellular and intracellular small–molecule galectin–3 inhibitors. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Dahlqvist, A.; Mandal, S.; Peterson, K.; Håkansson, M.; Logan, D.T.; Zetterberg, F.R.; Leffler, H.; Nilsson, U.J. 3–Substituted 1–Naphthamidomethyl–C–galactosyls Interact with Two Unique Sub–sites for High–Affinity and High–Selectivity Inhibition of Galectin-3. Molecules 2019, 24, 4554. [Google Scholar] [CrossRef]
- Chen, W.S.; Cao, Z.; Leffler, H.; Nilsson, U.J.; Panjwani, N. Galectin-3 Inhibition by a Small–Molecule Inhibitor Reduces Both Pathological Corneal Neovascularization and Fibrosis. Invest. Ophthalmol. Vis. Sci. 2017, 58, 9–20. [Google Scholar] [CrossRef]
- Rajput, V.K.; MacKinnon, A.; Mandal, S.; Collins, P.; Blanchard, H.; Leffler, H.; Sethi, T.; Schambye, H.; Mukhopadhyay, B.; Nilsson, U.J. A Selective Galactose–Coumarin–Derived Galectin-3 Inhibitor Demonstrates Involvement of Galectin-3–glycan Interactions in a Pulmonary Fibrosis Model. J. Med. Chem. 2016, 59, 8141–8147. [Google Scholar] [CrossRef]
- Mackinnon, A.C.; Gibbons, M.A.; Farnworth, S.L.; Leffler, H.; Nilsson, U.J.; Delaine, T.; Simpson, A.J.; Forbes, S.J.; Hirani, N.; Gauldie, J.; et al. Regulation of transforming growth factor–β1–driven lung fibrosis by galectin–3. Am. J. Respir. Crit. Care Med. 2012, 185, 537–546. [Google Scholar] [CrossRef]
- Volarevic, V.; Markovic, B.S.; Bojic, S.; Stojanovic, M.; Nilsson, U.; Leffler, H.; Besra, G.S.; Arsenijevic, N.; Paunovic, V.; Trajkovic, V.; et al. Gal–3 regulates the capacity of dendritic cells to promote NKT–cell–induced liver injury. Eur. J. Immunol. 2015, 45, 531–543. [Google Scholar] [CrossRef]
- Volarevic, V.; Milovanovic, M.; Ljujic, B.; Pejnovic, N.; Arsenijevic, N.; Nilsson, U.; Leffler, H.; Lukic, M.L. Galectin-3 deficiency prevents concanavalin A–induced hepatitis in mice. Hepatology 2012, 55, 1954–1964. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.I.; Whang, E.E.; Donner, D.B.; Jiang, X.; Price, B.D.; Carothers, A.M.; Delaine, T.; Leffler, H.; Nilsson, U.J.; Nose, V.; et al. Galectin-3 targeted therapy with a small molecule inhibitor activates apoptosis and enhances both chemosensitivity and radiosensitivity in papillary thyroid cancer. Mol. Cancer Res. 2009, 7, 1655–1662. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, H.; Liu, L.; Nakano, F.; Kawakita, F.; Kanamaru, H.; Nakatsuka, Y.; Okada, T.; Suzuki, H. Modified Citrus Pectin Prevents Blood–Brain Barrier Disruption in Mouse Subarachnoid Hemorrhage by Inhibiting Galectin-3. Stroke 2018, 49, 2743–2751. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srejovic, I.; Selakovic, D.; Jovicic, N.; Jakovljević, V.; Lukic, M.L.; Rosic, G. Galectin-3: Roles in Neurodevelopment, Neuroinflammation, and Behavior. Biomolecules 2020, 10, 798. https://doi.org/10.3390/biom10050798
Srejovic I, Selakovic D, Jovicic N, Jakovljević V, Lukic ML, Rosic G. Galectin-3: Roles in Neurodevelopment, Neuroinflammation, and Behavior. Biomolecules. 2020; 10(5):798. https://doi.org/10.3390/biom10050798
Chicago/Turabian StyleSrejovic, Ivan, Dragica Selakovic, Nemanja Jovicic, Vladimir Jakovljević, Miodrag L. Lukic, and Gvozden Rosic. 2020. "Galectin-3: Roles in Neurodevelopment, Neuroinflammation, and Behavior" Biomolecules 10, no. 5: 798. https://doi.org/10.3390/biom10050798
APA StyleSrejovic, I., Selakovic, D., Jovicic, N., Jakovljević, V., Lukic, M. L., & Rosic, G. (2020). Galectin-3: Roles in Neurodevelopment, Neuroinflammation, and Behavior. Biomolecules, 10(5), 798. https://doi.org/10.3390/biom10050798