Structure–Activity Relationship Study Reveals the Molecular Basis for Specific Sensing of Hydrophobic Amino Acids by the Campylobacter jejuni Chemoreceptor Tlp3


 ,
,
Abstract
1. Introduction
2. Methods
2.1. In Silico Screening
2.1.1. Preparation of Ligand Library
2.1.2. Virtual Screening for New Tlp3 LBD Ligands
2.2. Thermal Shift Assays
2.3. Isothermal Titration Calorimetry (ITC)
2.4. Construction of Isogenic Δtlp3 Mutant in C. Jejuni NCTC 11168
2.5. Chemotaxis Assay
2.6. Protein Crystallisation, Data Collection and Structure Determination
2.7. RCSB PDB Accession Numbers
3. Results
3.1. Screening for Potential Tlp3 LBD Ligands by Thermal Shift Assays
3.2. Screening for New Potential Isoleucine-Like Chemoeffectors by Molecular Docking
3.3. Ligand Binding Affinity Measurements by Isothermal Titration Calorimetry (ITC)
3.4. C. jejuni Tlp3 Mediates Positive Chemotactic Response to All Identified Ligands
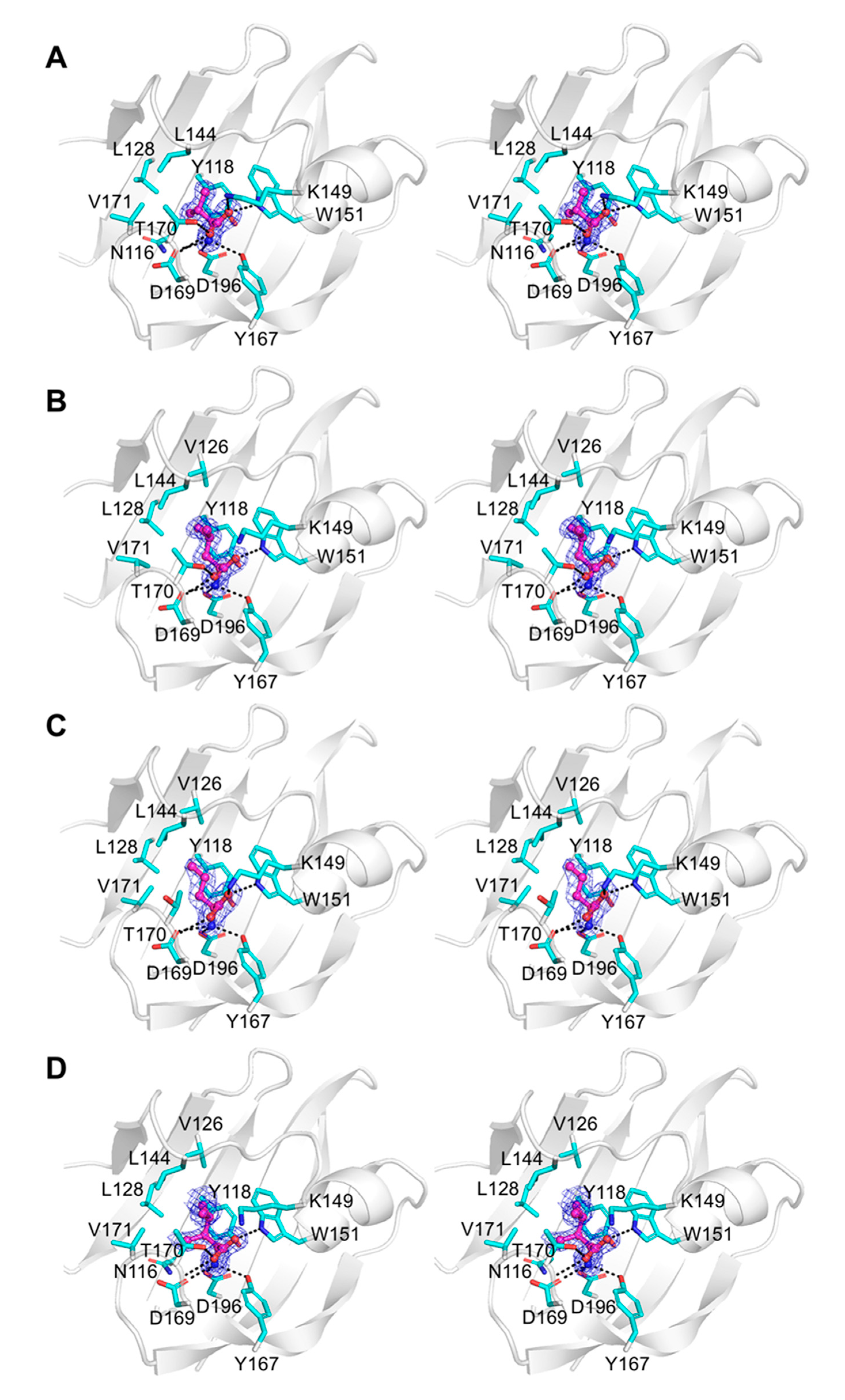
3.5. Crystallographic Analysis of Binding Modes of New Chemoeffectors
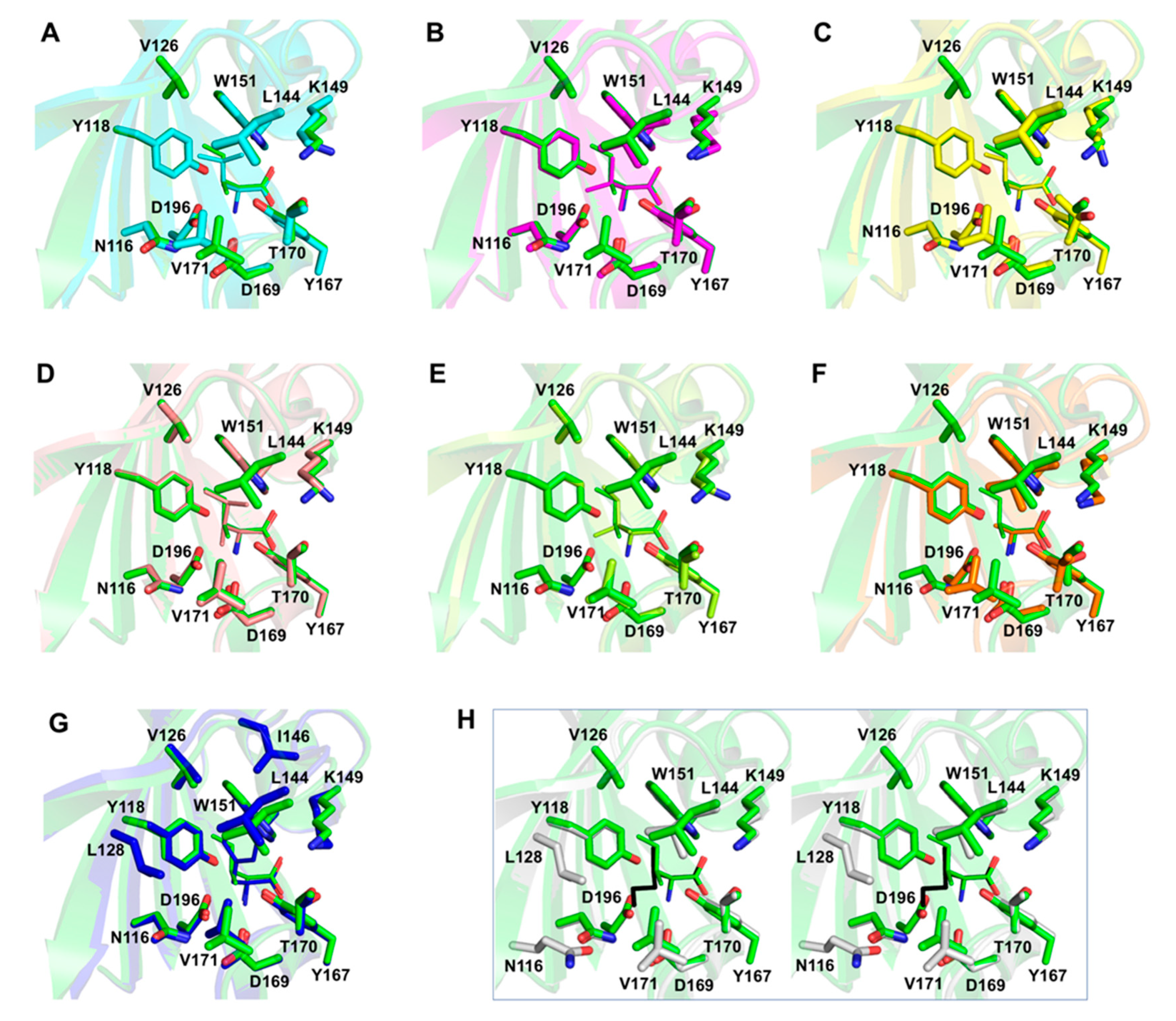
3.6. Analysis of Structure–Activity Relationship of Hydrophobic Amino Acids as Chemoreceptor Tlp3 Ligands
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boes, J.; Nersting, L.; Nielsen, E.M.; Kranker, S.; Enoe, C.; Wachmann, H.C.; Baggesen, D.L. Prevalence and diversity of Campylobacter jejuni in pig herds on farms with and without cattle or poultry. J. Food Prot. 2005, 68, 722–727. [Google Scholar] [CrossRef] [PubMed]
- Oporto, B.; Esteban, J.I.; Aduriz, G.; Juste, R.A.; Hurtado, A. Prevalence and strain diversity of thermophilic campylobacters in cattle, sheep and swine farms. J. Appl. Microbiol. 2007, 103, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Beery, J.T.; Hugdahl, M.B.; Doyle, M.P. Colonization of gastrointestinal tracts of chicks by Campylobacter jejuni. Appl. Environ. Microbiol. 1988, 54, 2365–2370. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.; Nielsen, E.M.; Engberg, J.; On, S.L.; Dietz, H.H. Comparison of genotypes and serotypes of Campylobacter jejuni isolated from Danish wild mammals and birds and from broiler flocks and humans. Appl. Environ. Microbiol. 2001, 67, 3115–3121. [Google Scholar] [CrossRef]
- Shane, S.M. Campylobacter infection of commercial poultry. Rev. Sci. Tech. 2000, 19, 376–395. [Google Scholar] [CrossRef]
- Dingle, K.E.; Colles, F.M.; Ure, R.; Wagenaar, J.A.; Duim, B.; Bolton, F.J.; Fox, A.J.; Wareing, D.R.; Maiden, M.C. Molecular characterization of Campylobacter jejuni clones: A basis for epidemiologic investigation. Emerg. Infect. Dis. 2002, 8, 949–955. [Google Scholar] [CrossRef]
- Friedman, C.R.; Hoekstra, R.M.; Samuel, M.; Marcus, R.; Bender, J.; Shiferaw, B.; Reddy, S.; Ahuja, S.D.; Helfrick, D.L.; Hardnett, F.; et al. Risk factors for sporadic Campylobacter infection in the United States: A case-control study in FoodNet sites. Clin. Infect. Dis. 2004, 38, S285–S296. [Google Scholar] [CrossRef]
- Epps, S.V.; Harvey, R.B.; Hume, M.E.; Phillips, T.D.; Anderson, R.C.; Nisbet, D.J. Foodborne Campylobacter: Infections, metabolism, pathogenesis and reservoirs. Int. J. Environ. Res. Public Health 2013, 10, 6292–6304. [Google Scholar] [CrossRef]
- Crim, S.M.; Iwamoto, M.; Huang, J.Y.; Griffin, P.M.; Gilliss, D.; Cronquist, A.B.; Cartter, M.; Tobin-D’Angelo, M.; Blythe, D.; Smith, K.; et al. Incidence and trends of infection with pathogens transmitted commonly through food—Foodborne Diseases Active Surveillance Network, 10 U.S. sites, 2006–2013. Morb. Mortal. Wkly. Rep. 2014, 63, 328–332. [Google Scholar]
- Blaser, M.J. Epidemiologic and clinical features of Campylobacter jejuni infections. J. Infect. Dis. 1997, 176, S103–S105. [Google Scholar] [CrossRef]
- Friedman, C.; Neimann, J.; Wegener, H.; Tauxe, R. Epidemiology of Campylobacter jejuni infections in the United States and other industrialized nations. In Campylobacter; Nachamkin, I., Szymanski, C.M., Blaser, M.J., Eds.; ASM Press: Washington, DC, USA, 2000; Volume 2, pp. 121–138. [Google Scholar]
- Schmidt-Ott, R.; Schmidt, H.; Feldmann, S.; Brass, F.; Krone, B.; Gross, U. Improved serological diagnosis stresses the major role of Campylobacter jejuni in triggering Guillain-Barre syndrome. Clin. Vaccine Immunol. 2006, 13, 779–783. [Google Scholar] [CrossRef] [PubMed]
- Dasti, J.I.; Tareen, A.M.; Lugert, R.; Zautner, A.E.; Gross, U. Campylobacter jejuni: A brief overview on pathogenicity-associated factors and disease-mediating mechanisms. Int. J. Med. Microbiol. 2010, 300, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Josenhans, C.; Suerbaum, S. The role of motility as a virulence factor in bacteria. Int. J. Med. Microbiol. 2002, 291, 605–614. [Google Scholar] [CrossRef]
- Morooka, T.; Umeda, A.; Amako, K. Motility as an intestinal colonization factor for Campylobacter jejuni. J. Gen. Microbiol. 1985, 131, 1973–1980. [Google Scholar] [CrossRef]
- Hendrixson, D.R.; DiRita, V.J. Identification of Campylobacter jejuni genes involved in commensal colonization of the chick gastrointestinal tract. Mol. Microbiol. 2004, 52, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Tareen, A.M.; Dasti, J.I.; Zautner, A.E.; Gross, U.; Lugert, R. Campylobacter jejuni proteins Cj0952c and Cj0951c affect chemotactic behaviour towards formic acid and are important for invasion of host cells. Microbiology 2010, 156, 3123–3135. [Google Scholar] [CrossRef] [PubMed]
- Zautner, A.E.; Tareen, A.M.; Gross, U.; Lugert, R. Chemotaxis in Campylobacter jejuni. Eur J. Microbiol Immunol (Bp) 2012, 2, 24–31. [Google Scholar] [CrossRef]
- Zhulin, I.B. The superfamily of chemotaxis transducers: From physiology to genomics and back. Adv. Microb. Physiol. 2001, 45, 157–198. [Google Scholar]
- Hazelbauer, G.L.; Falke, J.J.; Parkinson, J.S. Bacterial chemoreceptors: High-performance signaling in networked arrays. Trends Biochem. Sci. 2008, 33, 9–19. [Google Scholar] [CrossRef]
- Mund, N.L.; Masanta, W.O.; Goldschmidt, A.M.; Lugert, R.; Gross, U.; Zautner, A.E. Association of Campylobacter Jejuni ssp. Jejuni Chemotaxis Receptor Genes with Multilocus Sequence Types and Source of Isolation. Eur. J. Microbiol. Immunol. (Bp) 2016, 6, 162–177. [Google Scholar] [CrossRef]
- Marchant, J.; Wren, B.; Ketley, J. Exploiting genome sequence: Predictions for mechanisms of Campylobacter chemotaxis. Trends Microbiol. 2002, 10, 155–159. [Google Scholar] [CrossRef]
- Day, C.J.; Hartley-Tassell, L.E.; Shewell, L.K.; King, R.M.; Tram, G.; Day, S.K.; Semchenko, E.A.; Korolik, V. Variation of chemosensory receptor content of Campylobacter jejuni strains and modulation of receptor gene expression under different in vivo and in vitro growth conditions. BMC Microbiol. 2012, 12, 128. [Google Scholar] [CrossRef] [PubMed]
- Parkhill, J.; Wren, B.W.; Mungall, K.; Ketley, J.M.; Churcher, C.; Basham, D.; Chillingworth, T.; Davies, R.M.; Feltwell, T.; Holroyd, S.; et al. The genome sequence of the food-borne pathogen Campylobacter jejuni reveals hypervariable sequences. Nature 2000, 403, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Hugdahl, M.B.; Beery, J.T.; Doyle, M.P. Chemotactic behavior of Campylobacter jejuni. Infect. Immun. 1988, 56, 1560–1566. [Google Scholar] [CrossRef]
- Day, C.J.; King, R.M.; Shewell, L.K.; Tram, G.; Najnin, T.; Hartley-Tassell, L.E.; Wilson, J.C.; Fleetwood, A.D.; Zhulin, I.B.; Korolik, V. A direct-sensing galactose chemoreceptor recently evolved in invasive strains of Campylobacter jejuni. Nat. Commun. 2016, 7, 13206. [Google Scholar] [CrossRef]
- Vegge, C.S.; Brondsted, L.; Li, Y.P.; Bang, D.D.; Ingmer, H. Energy taxis drives Campylobacter jejuni toward the most favorable conditions for growth. Appl. Environ. Microbiol. 2009, 75, 5308–5314. [Google Scholar] [CrossRef]
- Hartley-Tassell, L.E.; Shewell, L.K.; Day, C.J.; Wilson, J.C.; Sandhu, R.; Ketley, J.M.; Korolik, V. Identification and characterization of the aspartate chemosensory receptor of Campylobacter jejuni. Mol. Microbiol. 2010, 75, 710–730. [Google Scholar] [CrossRef]
- Rahman, H.; King, R.M.; Shewell, L.K.; Semchenko, E.A.; Hartley-Tassell, L.E.; Wilson, J.C.; Day, C.J.; Korolik, V. Characterisation of a multi-ligand binding chemoreceptor CcmL (Tlp3) of Campylobacter jejuni. PLoS Path. 2014, 10, e1003822. [Google Scholar] [CrossRef]
- Li, Z.; Lou, H.; Ojcius, D.M.; Sun, A.; Sun, D.; Zhao, J.; Lin, X.; Yan, J. Methyl-accepting chemotaxis proteins 3 and 4 are responsible for Campylobacter jejuni chemotaxis and jejuna colonization in mice in response to sodium deoxycholate. J. Med. Microbiol. 2014, 63, 343–354. [Google Scholar] [CrossRef]
- Day, C.; King, R.; Tram, G.; Hartley-Tassell, L.; Fleetwood, A.; Zhulin, I.; Korolik, V. Characterisation of the galactose chemosensory receptor of Campylobacter jejuni. In Proceedings of the Australian Society for Microbiology Annual Scientific Meeting. Australian Society for Microbiology, Melbourne, Austria, 6–9 Jujly 2014. Abstract #107. [Google Scholar]
- Liu, Y.C.; Machuca, M.A.; Beckham, S.A.; Gunzburg, M.J.; Roujeinikova, A. Structural basis for amino-acid recognition and transmembrane signalling by tandem Per–Arnt–Sim (tandem PAS) chemoreceptor sensory domains. Acta Crystallogr. D Biol. Crystallogr. 2015, 71, 2127–2136. [Google Scholar] [CrossRef]
- Machuca, M.A.; Liu, Y.C.; Beckham, S.A.; Gunzburg, M.J.; Roujeinikova, A. The crystal structure of the tandem-PAS sensing domain of Campylobacter jejuni chemoreceptor Tlp1 suggests indirect mechanism of ligand recognition. J. Struct. Biol. 2016, 194, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, A.A.; Fleetwood, A.D.; Adebali, O.; Finn, R.D.; Zhulin, I.B. Cache domains that are homologous to, but different from PAS domains comprise the largest superfamily of extracellular sensors in prokaryotes. PLoS Comp. Biol. 2016, 12, e1004862. [Google Scholar] [CrossRef] [PubMed]
- Glekas, G.D.; Foster, R.M.; Cates, J.R.; Estrella, J.A.; Wawrzyniak, M.J.; Rao, C.V.; Ordal, G.W. A PAS domain binds asparagine in the chemotaxis receptor McpB in Bacillus subtilis. J. Biol. Chem. 2010, 285, 1870–1878. [Google Scholar] [CrossRef]
- Glekas, G.D.; Mulhern, B.J.; Kroc, A.; Duelfer, K.A.; Lei, V.; Rao, C.V.; Ordal, G.W. The Bacillus subtilis chemoreceptor McpC senses multiple ligands using two discrete mechanisms. J. Biol. Chem. 2012, 287, 39412–39418. [Google Scholar] [CrossRef]
- Oku, S.; Komatsu, A.; Tajima, T.; Nakashimada, Y.; Kato, J. Identification of chemotaxis sensory proteins for amino acids in Pseudomonas fluorescens Pf0-1 and their involvement in chemotaxis to tomato root exudate and root colonization. Microbes Environ. 2012, 27, ME12005. [Google Scholar] [CrossRef]
- Nishiyama, S.; Suzuki, D.; Itoh, Y.; Suzuki, K.; Tajima, H.; Hyakutake, A.; Homma, M.; Butler-Wu, S.M.; Camilli, A.; Kawagishi, I. Mlp24 (McpX) of Vibrio cholerae implicated in pathogenicity functions as a chemoreceptor for multiple amino acids. Infect. Immun. 2012, 80, 3170–3178. [Google Scholar] [CrossRef] [PubMed]
- Rico-Jiménez, M.; Muñoz-Martínez, F.; García-Fontana, C.; Fernandez, M.; Morel, B.; Ortega, Á.; Ramos, J.L.; Krell, T. Paralogous chemoreceptors mediate chemotaxis towards protein amino acids and the non-protein amino acid gamma-aminobutyrate (GABA). Mol. Microbiol. 2013, 88, 1230–1243. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.A.; Hildreth, S.; Helm, R.F.; Scharf, B.E. Sinorhizobium meliloti chemoreceptor McpU mediates chemotaxis toward host plant exudates through direct proline sensing. Appl. Environ. Microbiol. 2014, 80, 3404–3415. [Google Scholar] [CrossRef]
- McKellar, J.L.; Minnell, J.J.; Gerth, M.L. A high-throughput screen for ligand binding reveals the specificities of three amino acid chemoreceptors from Pseudomonas syringae pv. actinidiae. Mol. Microbiol. 2015, 96, 694–707. [Google Scholar] [CrossRef]
- Gavira, J.A.; Gumerov, V.M.; Rico-Jiménez, M.; Petukh, M.; Upadhyay, A.A.; Ortega, A.; Matilla, M.A.; Zhulin, I.B.; Krell, T. How Bacterial Chemoreceptors Evolve Novel Ligand Specificities. mBio 2020, 11, 11. [Google Scholar] [CrossRef]
- Ud-Din, A.; Khan, M.; Roujeinikova, A. Broad specificity of amino acid chemoreceptor CtaA of Pseudomonas fluorescens is afforded by plasticity of its amphipathic ligand-binding pocket. Mol. Plant.-Microbe Interact. 2020, 33, 4. [Google Scholar] [CrossRef]
- Machuca, M.A.; Johnson, K.S.; Liu, Y.C.; Steer, D.L.; Ottemann, K.M.; Roujeinikova, A. Helicobacter pylori chemoreceptor TlpC mediates chemotaxis to lactate. Sci. Rep. 2017, 7, 14089. [Google Scholar] [CrossRef] [PubMed]
- Gavira, J.A.; Ortega, Á.; Martín-Mora, D.; Conejero-Muriel, M.T.; Corral-Lugo, A.; Morel, B.; Matilla, M.A.; Krell, T. Structural basis for polyamine binding at the dCACHE domain of the McpU chemoreceptor from Pseudomonas putida. J. Mol. Biol. 2018, 430, 1950–1963. [Google Scholar] [CrossRef] [PubMed]
- Corral-Lugo, A.; Matilla, M.A.; Martín-Mora, D.; Jiménez, H.S.; Torres, N.M.; Kato, J.; Hida, A.; Oku, S.; Conejero-Muriel, M.; Gavira, J.A. High-affinity chemotaxis to histamine mediated by the TlpQ chemoreceptor of the human pathogen Pseudomonas aeruginosa. mBio 2018, 9, e01894-18. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.J.; Shoichet, B.K. ZINC− a free database of commercially available compounds for virtual screening. J. Chem. Inf. Model. 2005, 45, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.J.; Sterling, T.; Mysinger, M.M.; Bolstad, E.S.; Coleman, R.G. ZINC: A free tool to discover chemistry for biology. J. Chem. Inf. Model. 2012, 52, 1757–1768. [Google Scholar] [CrossRef]
- Sterling, T.; Irwin, J.J. ZINC 15–ligand discovery for everyone. J. Chem. Inf. Model. 2015, 55, 2324–2337. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminformatics 2011, 3, 33. [Google Scholar] [CrossRef]
- Rappé, A.K.; Casewit, C.J.; Colwell, K.; Goddard, W., III; Skiff, W. UFF, a full periodic table force field for molecular mechanics and molecular dynamics simulations. J. Am. Chem. Soc. 1992, 114, 10024–10035. [Google Scholar] [CrossRef]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with PyRx. Meth. Mol. Biol. 2015, 243–250. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Kooli, F.; Liu, Y.; Abboudi, M.; Rakass, S.; Hassani, H.O.; Ibrahim, S.M.; Al-Faze, R. Application of organo-magadiites for the removal of eosin dye from aqueous solutions: Thermal treatment and regeneration. Molecules 2018, 23, 2280. [Google Scholar] [CrossRef] [PubMed]
- Machuca, M.A.; Liu, Y.C.; Roujeinikova, A. Cloning, expression, refolding, purification and preliminary crystallographic analysis of the sensory domain of the Campylobacter chemoreceptor for aspartate A (CcaA). Acta Crystallogr. F 2015, 71, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Bochner, B.R.; Gadzinski, P.; Panomitros, E. Phenotype microarrays for high-throughput phenotypic testing and assay of gene function. Genome Res. 2001, 11, 1246–1255. [Google Scholar] [CrossRef]
- Lo, M.C.; Aulabaugh, A.; Jin, G.; Cowling, R.; Bard, J.; Malamas, M.; Ellestad, G. Evaluation of fluorescence-based thermal shift assays for hit identification in drug discovery. Anal. Biochem. 2004, 332, 153–159. [Google Scholar] [CrossRef]
- Pantoliano, M.W.; Petrella, E.C.; Kwasnoski, J.D.; Lobanov, V.S.; Myslik, J.; Graf, E.; Carver, T.; Asel, E.; Springer, B.A.; Lane, P.; et al. High-density miniaturized thermal shift assays as a general strategy for drug discovery. J. Biomol. Screen. 2001, 6, 429–440. [Google Scholar] [CrossRef]
- Trieu-Cuot, P.; Gerbaud, G.; Lambert, T.; Courvalin, P. In vivo transfer of genetic information between gram-positive and gram-negative bacteria. EMBO J. 1985, 4, 3583–3587. [Google Scholar] [CrossRef]
- Van Vliet, A.H.M.; Wood, A.C.; Wooldridge, K.; Ketley, J.M. Genetic Manipulation of Enteric Campylobacter Species. Methods Microbiol. 1998, 27, 407–419. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Elgamoudi, B.A.; Ketley, J.M.; Korolik, V. New approach to distinguishing chemoattractants, chemorepellents and catabolised chemoeffectors for Campylobacter jejuni. J. Microbiol. Methods 2018, 146, 83–91. [Google Scholar] [CrossRef]
- Machuca, M.A.; Liu, Y.C.; Beckham, S.A.; Roujeinikova, A. Cloning, refolding, purification and preliminary crystallographic analysis of the sensory domain of the Campylobacter chemoreceptor for multiple ligands (CcmL). Acta Crystallogr. F 2015, 71, 211–216. [Google Scholar] [CrossRef] [PubMed]
- McPhillips, T.M.; McPhillips, S.E.; Chiu, H.J.; Cohen, A.E.; Deacon, A.M.; Ellis, P.J.; Garman, E.; Gonzalez, A.; Sauter, N.K.; Phizackerley, R.P.; et al. Blu-Ice and the Distributed Control System: Software for data acquisition and instrument control at macromolecular crystallography beamlines. J. Synchrotron Radiat. 2002, 9, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Battye, T.G.; Kontogiannis, L.; Johnson, O.; Powell, H.R.; Leslie, A.G. iMOSFLM: A new graphical interface for diffraction-image processing with MOSFLM. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.R.; Murshudov, G.N. How good are my data and what is the resolution? Acta Crystallogr. D Biol. Crystallogr. 2013, 69, 1204–1214. [Google Scholar] [CrossRef] [PubMed]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.; McCoy, A.; et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Storoni, L.C.; Read, R.J. Likelihood-enhanced fast translation functions. Acta Crystallogr. D Biol. Crystallogr. 2005, 61, 458–464. [Google Scholar] [CrossRef]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef]
- Chen, V.B.; Arendall, W.B., 3rd; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef]
- DeLano, W.L. The PyMOL Molecular Graphics System. Available online: http://www.pymol.Org (accessed on 20 April 2020).
- Corral-Lugo, A.; De la Torre, J.; Matilla, M.A.; Fernandez, M.; Morel, B.; Espinosa-Urgel, M.; Krell, T. Assessment of the contribution of chemoreceptor-based signalling to biofilm formation. Environ. Microbiol. 2016, 18, 3355–3372. [Google Scholar] [CrossRef]
- Fernandez, M.; Morel, B.; Corral-Lugo, A.; Krell, T. Identification of a chemoreceptor that specifically mediates chemotaxis toward metabolizable purine derivatives. Mol. Microbiol. 2016, 99, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Dundas, J.; Ouyang, Z.; Tseng, J.; Binkowski, A.; Turpaz, Y.; Liang, J. CASTp: Computed atlas of surface topography of proteins with structural and topographical mapping of functionally annotated residues. Nucleic Acids Res. 2006, 34, W116–W118. [Google Scholar] [CrossRef] [PubMed]
- Ribardo, D.A.; Hendrixson, D.R. Analysis of the LIV system of Campylobacter jejuni reveals alternative roles for LivJ and LivK in commensalism beyond branched-chain amino acid transport. J. Bacteriol. 2011, 193, 6233–6243. [Google Scholar] [CrossRef] [PubMed]
- Rahmanian, M.; Claus, D.R.; Oxender, D.L. Multiplicity of leucine transport systems in Escherichia coli K-12. J. Bacteriol. 1973, 116, 1258–1266. [Google Scholar] [CrossRef] [PubMed]
- Koyanagi, T.; Katayama, T.; Suzuki, H.; Kumagai, H. Identification of the LIV-I/LS system as the third phenylalanine transporter in Escherichia coli K-12. J. Bacteriol. 2004, 186, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yu, Y.; Chen, X.; Diab, A.A.; Ruan, L.; He, J.; Wang, H.; He, Y.-W. The multiple DSF-family QS signals are synthesized from carbohydrate and branched-chain amino acids via the FAS elongation cycle. Sci. Rep. 2015, 5, 13294. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ZINC ID | Chemical Name | Predicted Free Energy of Binding (kcal mol−1) | Aqueous Solubility at pH 7.4 (mM) |
|---|---|---|---|
| ZINC000001707292 | 4-methyl-L-isoleucine | −7.1 | 2030 |
| ZINC000005841418 | β-methyl-L-norleucine | −6.7 | 1743 |
| ZINC000014967114 | 3-methyl-L-isoleucine | −7.2 | 61 |
| Dataset | Tlp3 LBD/ L-Leucine | Tlp3 LBD/ L-Valine | Tlp3 LBD/α-Amino-N-Valeric Acid | Tlp3 LBD/4-Methyl-L-Isoleucine | Tlp3 LBD/β-Methyl-L-Norleucine | Tlp3 LBD/3-Methyl-L-Isoleucine | Tlp3 LBD/L-Alanine | Tlp3 LBD/L-Phenylalanine |
|---|---|---|---|---|---|---|---|---|
| Space group | P21 | P21 | P21 | P21 | P21 | P21 | P21 | P21 |
| a, b, c (Å) | 42.6, 137.5, 48.9 | 42.6, 137.5, 49.1 | 41.9, 137.8, 49.2 | 42.4, 137.7, 48.8 | 42.1, 138.1, 48.8 | 42.3, 138.1, 48.8 | 42.6, 137.6, 49.1 | 42.4, 137.4, 48.8 |
| β (°) | 94.3 | 94.5 | 93.9 | 94.0 | 94.7 | 93.8 | 94.7 | 94.0 |
| Resolution range (Å) | 28.21–1.40 (1.42–1.40) | 28.18–1.40 (1.42–1.40) | 46.23–2.10 (2.16–2.10) | 40.45–1.42 (1.50–1.42) | 35.86–1.38 (1.46–1.38) | 35.99–1.38 (1.46–1.38) | 46.12–1.32 (1.34–1.32) | 45.85–1.38 (1.40–1.38) |
| Rmerge | 0.048 (0.399) | 0.047 (0.308) | 0.077 (0.313) | 0.042 (0.147) | 0.056 (0.333) | 0.068 (0.342) | 0.026 (0.152) | 0.032 (0.292) |
| Average I/σ(I) | 10.7 (2.6) | 11.4 (3.1) | 10.6 (3.3) | 17.1 (6.4) | 10.1 (2.8) | 9.1 (2.6) | 15.9 (4.1) | 12.4 (2.6) |
| Completeness (%) | 99.2 (99.4) | 94.3 (88.0) | 97.6 (96.6) | 98.5 (98.4) | 89.1 (87.6) | 81.1 (74.8) | 97.5 (89.5) | 98.7 (99.8) |
| Redundancy | 3.7 | 3.2 | 4.3 | 3.6 | 3.1 | 3.5 | 3.5 | 3.2 |
| Observed reflections | 403, 905 | 328, 682 | 134, 763 | 375, 443 | 311, 496 | 320, 095 | 445, 181 | 354, 924 |
| Unique reflections | 109, 203 | 103, 884 | 31, 594 | 102, 980 | 100, 314 | 91, 649 | 128, 662 | 964, 43 |
| Dataset | Tlp3 LBD/ L-Leucine | Tlp3 LBD/ L-Valine | Tlp3 LBD/α-Amino-N-Valeric Acid | Tlp3 LBD/4-Methyl-L-Isoleucine | Tlp3 LBD/β-Methyl-L-Norleucine | Tlp3 LBD/3-Methyl-L-Isoleucine | Tlp3 LBD/L-Alanine | Tlp3 LBD/L-Phenylalanine |
|---|---|---|---|---|---|---|---|---|
| Rwork/Rfree a | 0.168/0.189 | 0.169/0.191 | 0.168/0.214 | 0.115/0.162 | 0.127/0.180 | 0.121/0.180 | 0.124/0.156 | 0.132/0.174 |
| No. protein atoms | 4465 | 4489 | 4221 | 4146 | 4186 | 4102 | 4207 | 4102 |
| No. water molecules | 820 | 944 | 241 | 936 | 845 | 833 | 799 | 758 |
| Average B (protein atoms, Å2) | 15 | 14 | 26 | 19 | 21 | 24 | 19 | 25 |
| Average B (water molecules, Å2) | 29 | 29 | 30 | 40 | 41 | 43 | 38 | 43 |
| Average B (ligand, Å2) | 10 | 10 | 15 | 17 | 20 | 18 | 14 | 25 |
| r.m.s. deviations from ideality | ||||||||
| Bond lengths (Å) | 0.005 | 0.005 | 0.006 | 0.012 | 0.012 | 0.012 | 0.012 | 0.012 |
| Bond angles (°) | 0.8 | 1.8 | 0.8 | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 |
| Ramachandran plot (%) Favoured | 99 | 99 | 100 | 100 | 99 | 100 | 99 | 99 |
| Allowed | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 1 |
| Outliers | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Compound | Thermal Shift | Thermodynamic Parameters | ||
|---|---|---|---|---|
| ∆Tm (°C) | Kd (µM) | ∆H (kcal mol−1) | ∆S (cal mol−1 deg−1) | |
| L-isoleucine a | 2.1 ± 0.8 | 86 ± 10 | −4.4 ± 0.2 | 3.6 |
| L-leucine | 2.4 ± 0.2 | 105 ± 6 | −4.6 ± 0.1 | 2.9 |
| L-valine | 1.3 ± 0.1 | 405 ± 27 | −4.4 ± 0.2 | 0.8 |
| α-amino-N-valeric acid | 1.4 ± 0.3 | 168 ± 9 | −6.5 ± 0.2 | −4.5 |
| Ala-Thr | 1.1 ± 0.3 | NB b | ||
| 2-deoxycytidine | 1.4 ± 0.8 | NB | ||
| methyl pyruvate | −1.8 ± 0.0 | NB | ||
| L-lysine | −0.5 ± 0.1 | NB | ||
| L-arginine | 0.1 ± 0.2 | NB | ||
| L-aspartic acid | −0.2 ± 0.7 | NB | ||
| succinic acid | −0.1 ± 0.6 | NP | ||
| malic acid | −0.8 ± 0.4 | NP | ||
| thiamine | −0.1 ± 0.4 | NP | ||
| purine | NP c | NB | ||
| 4-methyl-L-isoleucine | 2.8 ± 0.3 | 324 ± 20 | −5.1 ± 0.2 | −1.01 |
| β-methyl-L-norleucine | 3.7 ± 0.2 | 294 ± 14 | −6.2 ± 0.2 | −4.77 |
| 3-methyl-L-isoleucine | 2.0 ± 0.1 | 484 ± 30 | −5.7 ± 0.2 | −4.03 |
| L-alanine | −0.6 ± 0.7 | 4980 ± 917 | −150.2 ± 54.5 | −495.50 |
| L-phenylalanine | −0.3 ± 0.4 | 730 ± 55 | −5.9 ± 0.3 | −5.44 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.F.; Machuca, M.A.; Rahman, M.M.; Koç, C.; Norton, R.S.; Smith, B.J.; Roujeinikova, A. Structure–Activity Relationship Study Reveals the Molecular Basis for Specific Sensing of Hydrophobic Amino Acids by the Campylobacter jejuni Chemoreceptor Tlp3. Biomolecules 2020, 10, 744. https://doi.org/10.3390/biom10050744
Khan MF, Machuca MA, Rahman MM, Koç C, Norton RS, Smith BJ, Roujeinikova A. Structure–Activity Relationship Study Reveals the Molecular Basis for Specific Sensing of Hydrophobic Amino Acids by the Campylobacter jejuni Chemoreceptor Tlp3. Biomolecules. 2020; 10(5):744. https://doi.org/10.3390/biom10050744
Chicago/Turabian StyleKhan, Mohammad F., Mayra A. Machuca, Mohammad M. Rahman, Cengiz Koç, Raymond S. Norton, Brian J. Smith, and Anna Roujeinikova. 2020. "Structure–Activity Relationship Study Reveals the Molecular Basis for Specific Sensing of Hydrophobic Amino Acids by the Campylobacter jejuni Chemoreceptor Tlp3" Biomolecules 10, no. 5: 744. https://doi.org/10.3390/biom10050744
APA StyleKhan, M. F., Machuca, M. A., Rahman, M. M., Koç, C., Norton, R. S., Smith, B. J., & Roujeinikova, A. (2020). Structure–Activity Relationship Study Reveals the Molecular Basis for Specific Sensing of Hydrophobic Amino Acids by the Campylobacter jejuni Chemoreceptor Tlp3. Biomolecules, 10(5), 744. https://doi.org/10.3390/biom10050744