EffHunter: A Tool for Prediction of Effector Protein Candidates in Fungal Proteomic Databases

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
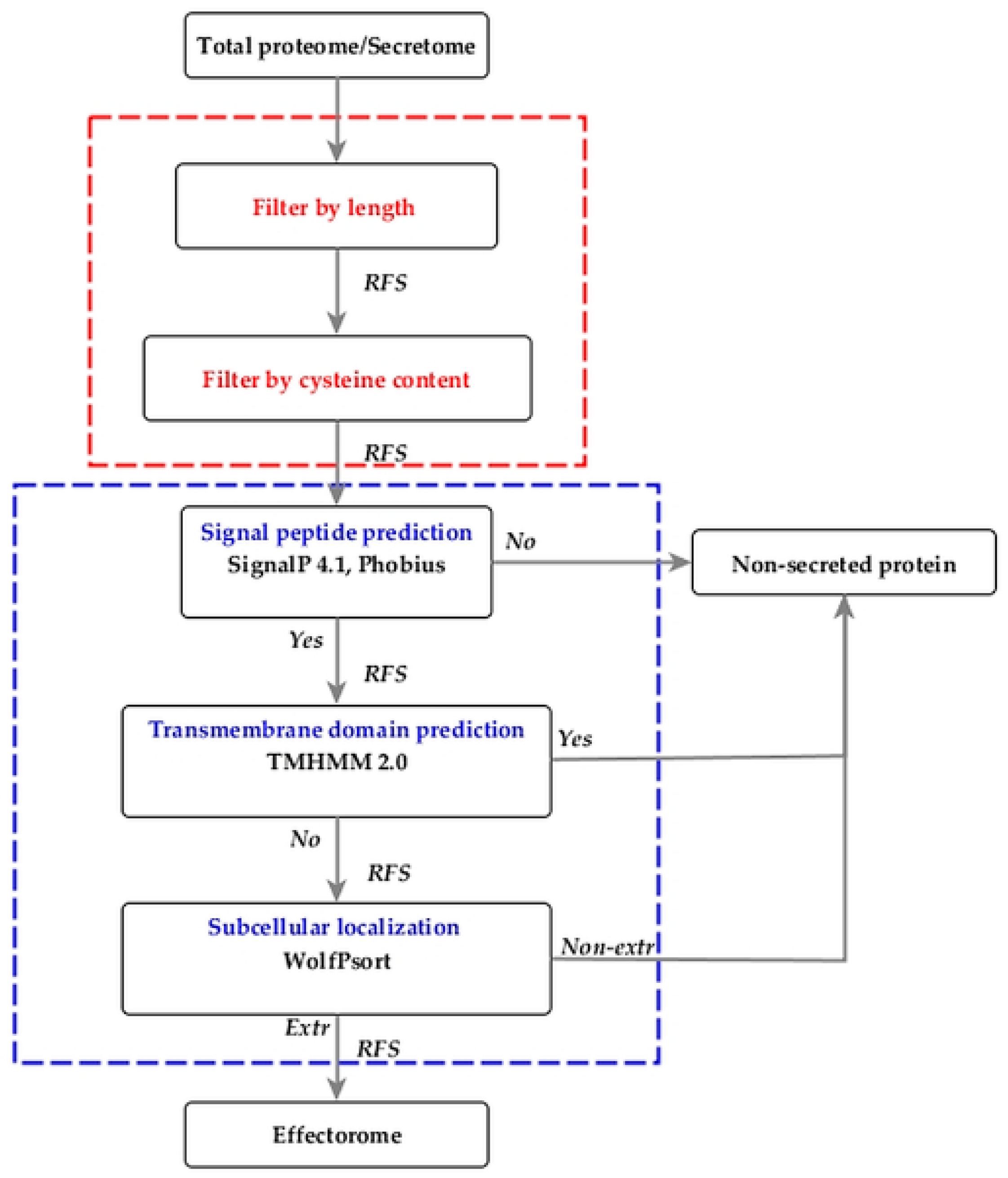
2.1. Architecture of EffHunter Pipeline
2.2. Validation of EffHunter Pipeline in ab Initio Approach
2.3. Validation of EffHunter Pipeline in Comparative Approach
2.4. Prediction of Effector Proteins in Fungal Genomes
3. Results
3.1. EffHunter: A Pipeline to Predict Fungal Effectors Proteins
3.2. Validation ab Initio
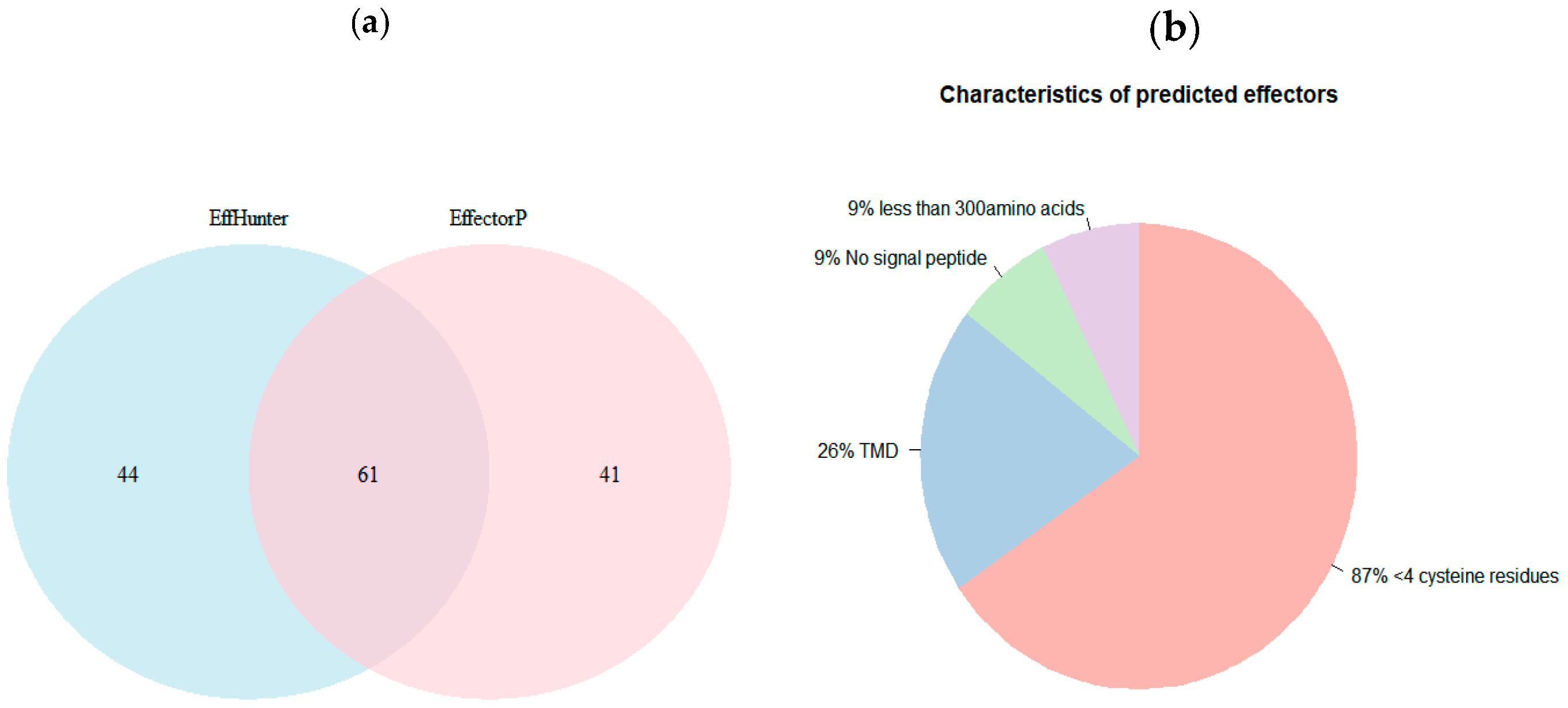
3.3. Validation of EffHunter with Fungal Proteomes and Comparison with Other Effector Prediction Tools
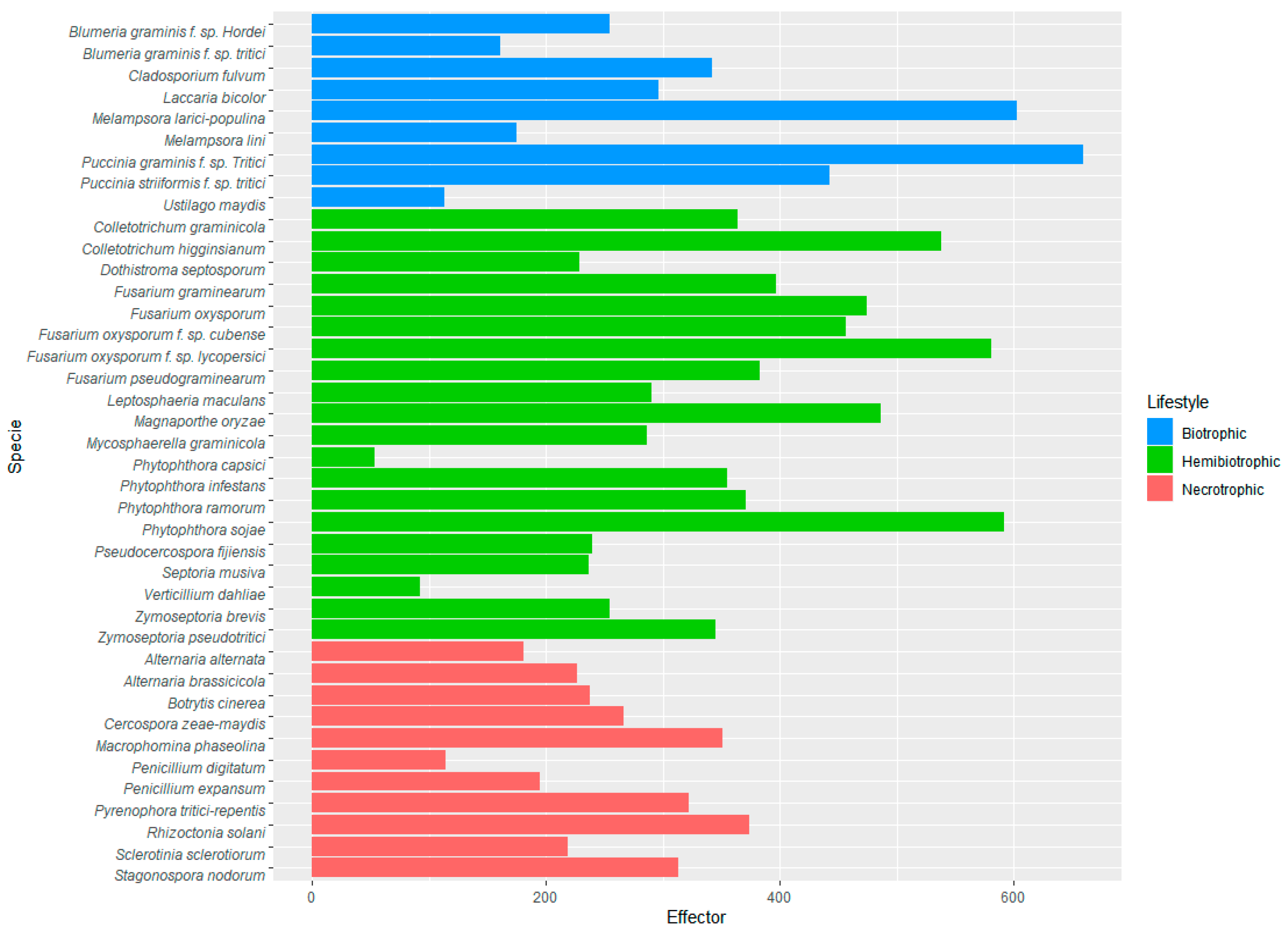
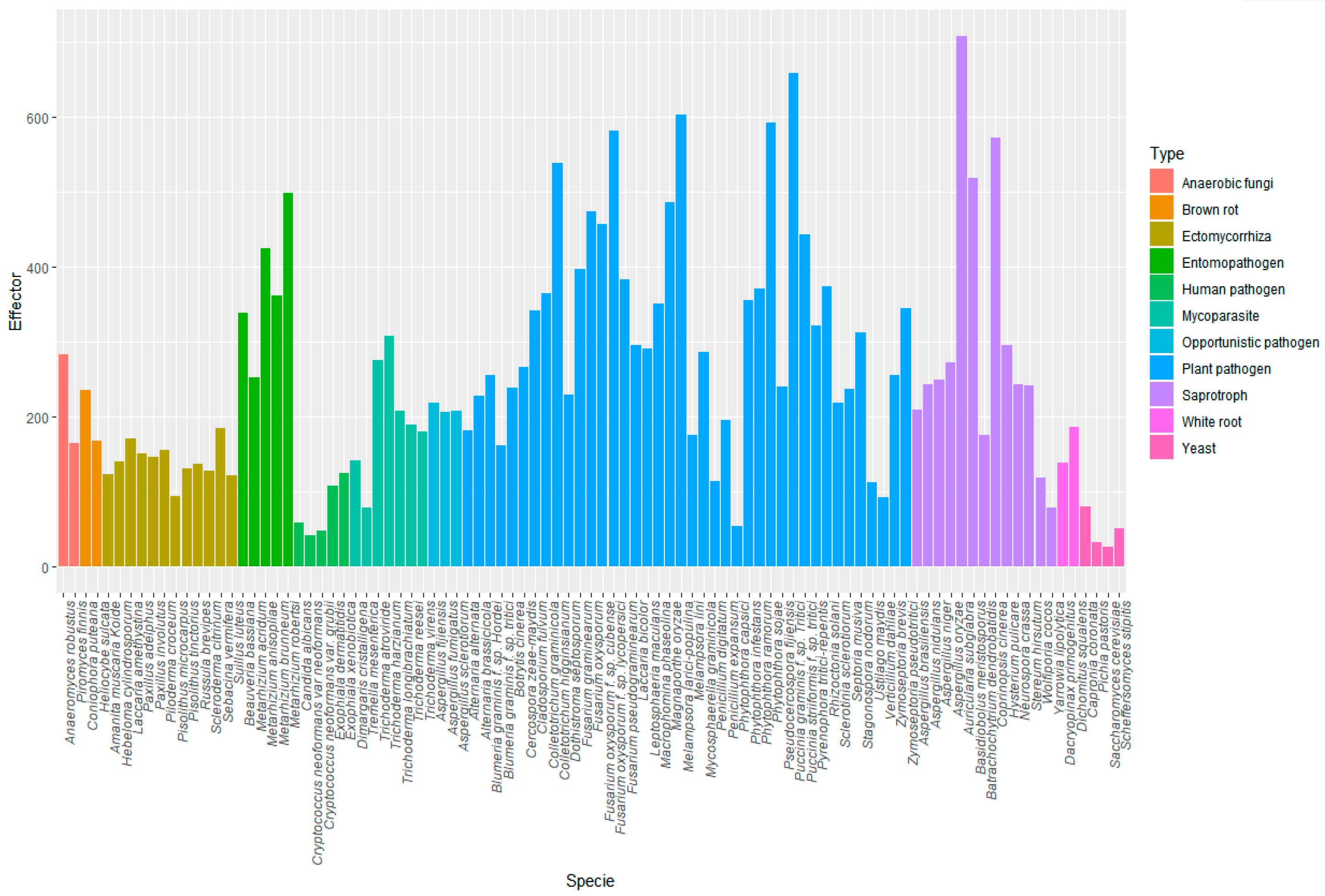
3.4. Prediction of Effector Proteins in Several Fungal and Oomycetes Proteomes with EffHunter
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dalio, R.J.D.; Herlihy, J.; Oliveira, T.S.; McDowell, J.M.; Machado, M. Effector Biology in Focus: A Primer for Computational Prediction and Functional Characterization. Mol. Plant-Microbe Interact. 2018, 31, 22–33. [Google Scholar] [CrossRef] [PubMed]
- de Jonge, R.; Bolton, M.D.; Thomma, B.P. How filamentous pathogens co-opt plants: The ins and outs of fungal effectors. Curr. Opin. Plant Biol. 2011, 14, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, M.C.; Valent, B. Filamentous plant pathogen effectors in action. Nat. Rev. Microbiol. 2013, 11, 800–814. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Edwards, M.C. Genome-Wide Analysis of Small Secreted Cysteine-Rich Proteins Identifies Candidate Effector Proteins Potentially Involved in Fusarium graminearum −Wheat Interactions. Phytopathology 2016, 106, 166–176. [Google Scholar] [CrossRef]
- Neu, E.; Debener, T. Prediction of the Diplocarpon rosae secretome reveals candidate genes for effectors and virulence factors. Fungal. Biol. 2019, 123, 231–239. [Google Scholar] [CrossRef]
- Petre, B.; Saunders, D.G.O.; Sklenar, J.; Lorrain, C.; Win, J.; Duplessis, S.; Kamoun, S. Candidate Effector Proteins of the Rust Pathogen Melampsora larici-populina Target Diverse Plant Cell Compartments. Mol. Plant-Microbe Interact. 2015, 28, 689–700. [Google Scholar] [CrossRef]
- Rep, M. Small proteins of plant-pathogenic fungi secreted during host colonization. Fems Microbiol. Lett. 2005, 253, 19–27. [Google Scholar] [CrossRef]
- de Wit, P.J.; Mehrabi, R.; van den Burg, H.A.; Stergiopoulos, I. Fungal effector proteins: Past, present and future. Mol. Plant Pathol. 2009, 10, 735–747. [Google Scholar] [CrossRef]
- Liu, T.; Song, T.; Zhang, X.; Yuan, H.; Su, L.; Li, W.; Xu, J.; Liu, S.; Chen, L.; Chen, T.; et al. Unconventionally secreted effectors of two filamentous pathogens target plant salicylate biosynthesis. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef]
- Sonah, H.; Deshmukh, R.K.; Bélanger, R.R. Computational Prediction of Effector Proteins in Fungi: Opportunities and Challenges. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Raffaele, S.; Kamoun, S. Genome evolution in filamentous plant pathogens: Why bigger can be better. Nat. Rev. Microbiol. 2012, 10, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes11Edited by F. Cohen. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, J.; Dodds, P.N.; Gardiner, D.M.; Singh, K.B.; Taylor, J.M. Improved prediction of fungal effector proteins from secretomes with EffectorP 2.0: Prediction of fungal effectors with EffectorP 2.0. Mol. Plant Pathol. 2018, 19, 2094–2110. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, J.; Gardiner, D.M.; Dodds, P.N.; Tini, F.; Covarelli, L.; Singh, K.B.; Manners, J.M.; Taylor, J.M. EffectorP: Predicting fungal effector proteins from secretomes using machine learning. New Phytol. 2016, 210, 743–761. [Google Scholar] [CrossRef]
- White, F.F.; Jones, J.B. One effector at a time. Nat. Plants 2018, 4, 134–135. [Google Scholar] [CrossRef]
- Kall, L.; Krogh, A.; Sonnhammer, E.L.L. Advantages of combined transmembrane topology and signal peptide prediction--the Phobius web server. Nucleic Acids Res. 2007, 35, W429–W432. [Google Scholar] [CrossRef]
- Urban, M.; Cuzick, A.; Rutherford, K.; Irvine, A.; Pedro, H.; Pant, R.; Sadanadan, V.; Khamari, L.; Billal, S.; Mohanty, S.; et al. PHI-base: A new interface and further additions for the multi-species pathogen–host interactions database. Nucleic Acids Res. 2017, 45, D604–D610. [Google Scholar] [CrossRef]
- Winnenburg, R. PHI-base: A new database for pathogen host interactions. Nucleic Acids Res. 2006, 34, D459–D464. [Google Scholar] [CrossRef]
- Bowen, J.K.; Mesarich, C.H.; Rees-George, J.; Cui, W.; Fitzgerald, A.; Win, J.; Plummer, K.M.; Templeton, M.D. Candidate effector gene identification in the ascomycete fungal phytopathogen Venturia inaequalis by expressed sequence tag analysis. Mol. Plant Pathol. 2009, 10, 431–448. [Google Scholar] [CrossRef] [PubMed]
- Dölfors, F.; Holmquist, L.; Dixelius, C.; Tzelepis, G. A LysM effector protein from the basidiomycete Rhizoctonia solani contributes to virulence through suppression of chitin-triggered immunity. Mol. Genet. Genom. 2019. [Google Scholar] [CrossRef]
- Esna Ashari, Z.; Dasgupta, N.; Brayton, K.A.; Broschat, S.L. An optimal set of features for predicting type IV secretion system effector proteins for a subset of species based on a multi-level feature selection approach. PLoS ONE 2018, 13, e0197041. [Google Scholar] [CrossRef] [PubMed]
- Krijger, J.J.; Thon, M.R.; Deising, H.B.; Wirsel, S.G. Compositions of fungal secretomes indicate a greater impact of phylogenetic history than lifestyle adaptation. BMC Genom. 2014, 15, 722. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Nan, C.; Hu, F. Accurate prediction of bacterial type IV secreted effectors using amino acid composition and PSSM profiles. Bioinformatics 2013, 29, 3135–3142. [Google Scholar] [CrossRef] [PubMed]
- Duplessis, S.; Cuomo, C.A.; Lin, Y.-C.; Aerts, A.; Tisserant, E.; Veneault-Fourrey, C.; Joly, D.L.; Hacquard, S.; Amselem, J.; Cantarel, B.L.; et al. Obligate biotrophy features unraveled by the genomic analysis of rust fungi. Proc. Natl. Acad. Sci. USA 2011, 108, 9166–9171. [Google Scholar] [CrossRef]
- Marton, K.; Flajšman, M.; Radišek, S.; Košmelj, K.; Jakše, J.; Javornik, B.; Berne, S. Comprehensive analysis of Verticillium nonalfalfae in silico secretome uncovers putative effector proteins expressed during hop invasion. PLoS ONE 2018, 13, e0198971. [Google Scholar] [CrossRef]
- Sperschneider, J.; Williams, A.H.; Hane, J.K.; Singh, K.B.; Taylor, J.M. Evaluation of Secretion Prediction Highlights Differing Approaches Needed for Oomycete and Fungal Effectors. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC transporters: The power to change. Nat. Rev. Mol. Cell Biol. 2009, 10, 218–227. [Google Scholar] [CrossRef]
- Wilkens, S. Structure and mechanism of ABC transporters. F1000Prime Reports 2015, 7. [Google Scholar] [CrossRef]
- Werck-Reichhart, D.; Feyereisen, R. Cytochromes P450: A success story. Genome Biol. 2000, 1, reviews3003.1. [Google Scholar] [CrossRef]
- Lin, H.-C.; Yu, P.-L.; Chen, L.-H.; Tsai, H.-C.; Chung, K.-R. A Major Facilitator Superfamily Transporter Regulated by the Stress-Responsive Transcription Factor Yap1 Is Required for Resistance to Fungicides, Xenobiotics, and Oxidants and Full Virulence in Alternaria alternata. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Nagarathinam, K.; Nakada-Nakura, Y.; Parthier, C.; Terada, T.; Juge, N.; Jaenecke, F.; Liu, K.; Hotta, Y.; Miyaji, T.; Omote, H.; et al. Outward open conformation of a Major Facilitator Superfamily multidrug/H+ antiporter provides insights into switching mechanism. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Yan, N. Structural Biology of the Major Facilitator Superfamily Transporters. Annu. Rev. Biophys. 2015, 44, 257–283. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, J.; Catanzariti, A.-M.; DeBoer, K.; Petre, B.; Gardiner, D.M.; Singh, K.B.; Dodds, P.N.; Taylor, J.M. LOCALIZER: Subcellular localization prediction of both plant and effector proteins in the plant cell. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Wang, Q.; Yang, J.; Zhu, X.; Wei, D.-Q. PredT4SE-Stack: Prediction of Bacterial Type IV Secreted Effectors From Protein Sequences Using a Stacked Ensemble Method. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Zeng, C.; Zou, L. An account of in silico identification tools of secreted effector proteins in bacteria and future challenges. Brief. Bioinform. 2019, 20, 110–129. [Google Scholar] [CrossRef]
- Liang, P.; Liu, S.; Xu, F.; Jiang, S.; Yan, J.; He, Q.; Liu, W.; Lin, C.; Zheng, F.; Wang, X.; et al. Powdery Mildews Are Characterized by Contracted Carbohydrate Metabolism and Diverse Effectors to Adapt to Obligate Biotrophic Lifestyle. Front. Microbiol. 2018, 9, 3160. [Google Scholar] [CrossRef]
- Chang, T.-C.; Salvucci, A.; Crous, P.W.; Stergiopoulos, I. Comparative Genomics of the Sigatoka Disease Complex on Banana Suggests a Link between Parallel Evolutionary Changes in Pseudocercospora fijiensis and Pseudocercospora eumusae and Increased Virulence on the Banana Host. PLoS Genet 2016, 12, e1005904. [Google Scholar] [CrossRef]
- Morais do Amaral, A.; Antoniw, J.; Rudd, J.J.; Hammond-Kosack, K.E. Defining the Predicted Protein Secretome of the Fungal Wheat Leaf Pathogen Mycosphaerella graminicola. PLoS ONE 2012, 7, e49904. [Google Scholar] [CrossRef]
- Cortázar, A.R.; Aransay, A.M.; Alfaro, M.; Oguiza, J.A.; Lavín, J.L. SECRETOOL: Integrated secretome analysis tool for fungi. Amino Acids 2014, 46, 471–473. [Google Scholar] [CrossRef] [PubMed]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucl. Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef] [PubMed]
- Kersey, P.J.; Allen, J.E.; Allot, A.; Barba, M.; Boddu, S.; Bolt, B.J.; Carvalho-Silva, D.; Christensen, M.; Davis, P.; Grabmueller, C.; et al. Ensembl Genomes 2018: An integrated omics infrastructure for non-vertebrate species. Nucleic Acids Res. 2018, 46, D802–D808. [Google Scholar] [CrossRef] [PubMed]
- Wall, L.; Schwartz, R. Programming PERL. In Nutshell Handbooks, 1st ed.; O’Reily & Associates, Inc.: 101 Morris Street, Sebastopol, CA, USA, 1991; ISBN 978-0-937175-64-4. [Google Scholar]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; von Heijne, G. Predicting Subcellular Localization of Proteins Based on their N-terminal Amino Acid Sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef]
- Eisenhaber, B.; Bork, P.; Eisenhaber, F. Prediction of Potential GPI-modification Sites in Proprotein Sequences. J. Mol. Biol. 1999, 292, 741–758. [Google Scholar] [CrossRef]
- Rastogi, S.; Rost, B. LocDB: Experimental annotations of localization for Homo sapiens and Arabidopsis thaliana. Nucleic Acids Res. 2011, 39, D230–D234. [Google Scholar] [CrossRef]
- Solovyev, V.; Sagitov, V. Softberry—ProtComp. Softberry, 1997. Available online: http://www.softberry.com/ (accessed on 15 August 2019).
- Ohm, R.A.; Feau, N.; Henrissat, B.; Schoch, C.L.; Horwitz, B.A.; Barry, K.W.; Condon, B.J.; Copeland, A.C.; Dhillon, B.; Glaser, F.; et al. Diverse Lifestyles and Strategies of Plant Pathogenesis Encoded in the Genomes of Eighteen Dothideomycetes Fungi. PLoS Pathog. 2012, 8, e1003037. [Google Scholar] [CrossRef]
- Frantzeskakis, L.; Kracher, B.; Kusch, S.; Yoshikawa-Maekawa, M.; Bauer, S.; Pedersen, C.; Spanu, P.D.; Maekawa, T.; Schulze-Lefert, P.; Panstruga, R. Signatures of host specialization and a recent transposable element burst in the dynamic one-speed genome of the fungal barley powdery mildew pathogen. BMC Genom. 2018, 19, 381. [Google Scholar] [CrossRef]
- de Wit, P.J.G.M.; van der Burgt, A.; Ökmen, B.; Stergiopoulos, I.; Abd-Elsalam, K.A.; Aerts, A.L.; Bahkali, A.H.; Beenen, H.G.; Chettri, P.; Cox, M.P.; et al. The Genomes of the Fungal Plant Pathogens Cladosporium fulvum and Dothistroma septosporum Reveal Adaptation to Different Hosts and Lifestyles But Also Signatures of Common Ancestry. PLoS Genet 2012, 8, e1003088. [Google Scholar] [CrossRef]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet 2012, 44, 1060–1065. [Google Scholar] [CrossRef]
- Armitage, A.D.; Taylor, A.; Sobczyk, M.K.; Baxter, L.; Greenfield, B.P.J.; Bates, H.J.; Wilson, F.; Jackson, A.C.; Ott, S.; Harrison, R.J.; et al. Characterisation of pathogen-specific regions and novel effector candidates in Fusarium oxysporum f. sp. cepae. Sci. Rep. 2018, 8, 13530. [Google Scholar] [CrossRef] [PubMed]
- Rouxel, T.; Grandaubert, J.; Hane, J.K.; Hoede, C.; van de Wouw, A.P.; Couloux, A.; Dominguez, V.; Anthouard, V.; Bally, P.; Bourras, S.; et al. Effector diversification within compartments of the Leptosphaeria maculans genome affected by Repeat-Induced Point mutations. Nat. Commun. 2011, 2, 202. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.A.; Talbot, N.J.; Ebbole, D.J.; Farman, M.L.; Mitchell, T.K.; Orbach, M.J.; Thon, M.; Kulkarni, R.; Xu, J.-R.; Pan, H.; et al. The genome sequence of the rice blast fungus Magnaporthe grisea. Nature 2005, 434, 980–986. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, S.B.; Ben M’Barek, S.; Dhillon, B.; Wittenberg, A.H.J.; Crane, C.F.; Hane, J.K.; Foster, A.J.; Van der Lee, T.A.J.; Grimwood, J.; Aerts, A.; et al. Finished Genome of the Fungal Wheat Pathogen Mycosphaerella graminicola Reveals Dispensome Structure, Chromosome Plasticity, and Stealth Pathogenesis. PLoS Genet 2011, 7, e1002070. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Kamoun, S.; Zody, M.C.; Jiang, R.H.Y.; Handsaker, R.E.; Cano, L.M.; Grabherr, M.; Kodira, C.D.; Raffaele, S.; Torto-Alalibo, T.; et al. Genome sequence and analysis of the Irish potato famine pathogen Phytophthora infestans. Nature 2009, 461, 393–398. [Google Scholar] [CrossRef]
- Manning, V.A.; Pandelova, I.; Dhillon, B.; Wilhelm, L.J.; Goodwin, S.B.; Berlin, A.M.; Figueroa, M.; Freitag, M.; Hane, J.K.; Henrissat, B.; et al. Comparative Genomics of a Plant-Pathogenic Fungus, Pyrenophora tritici-repentis , Reveals Transduplication and the Impact of Repeat Elements on Pathogenicity and Population Divergence. G3 2013, 3, 41–63. [Google Scholar] [CrossRef]
- Kämper, J.; Kahmann, R.; Bölker, M.; Ma, L.-J.; Brefort, T.; Saville, B.J.; Banuett, F.; Kronstad, J.W.; Gold, S.E.; Müller, O.; et al. Insights from the genome of the biotrophic fungal plant pathogen Ustilago maydis. Nature 2006, 444, 97–101. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Z.; Halterman, D.A. Correction: Molecular Determinants of Resistance Activation and Suppression by Phytophthora infestans Effector IPI-O. PLoS Pathog. 2012, 8, e1002595. [Google Scholar] [CrossRef]
- Sánchez-Vallet, A.; Saleem-Batcha, R.; Kombrink, A.; Hansen, G.; Valkenburg, D.-J.; Thomma, B.P.; Mesters, J.R. Fungal effector Ecp6 outcompetes host immune receptor for chitin binding through intrachain LysM dimerization. eLife 2013, 2, e00790. [Google Scholar] [CrossRef]
- Lorang, J.M.; Sweat, T.A.; Wolpert, T.J. Plant disease susceptibility conferred by a “resistance” gene. Proc. Natl. Acad. Sci. USA 2007, 104, 14861–14866. [Google Scholar] [CrossRef]
- Almeida, D.S.; Menezes-Blackburn, D.; Zhang, H.; Haygarth, P.M.; Rosolem, C.A. Phosphorus availability and dynamics in soil affected by long-term ruzigrass cover crop. Geoderma 2019, 337, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Vleeshouwers, V.G.A.A.; Oliver, R.P. Effectors as Tools in Disease Resistance Breeding Against Biotrophic, Hemibiotrophic, and Necrotrophic Plant Pathogens. Mol. Plant-Microbe Interact. 2014, 27, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Oliver, R. Plant breeding for disease resistance in the age of effectors. Phytoparasitica 2009, 37, 1–5. [Google Scholar] [CrossRef]
- Kettles, G.J.; Bayon, C.; Canning, G.; Rudd, J.J.; Kanyuka, K. Apoplastic recognition of multiple candidate effectors from the wheat pathogen Zymoseptoria tritici in the nonhost plant Nicotiana benthamiana. New Phytol. 2017, 213, 338–350. [Google Scholar] [CrossRef]
- Casarrubia, S.; Daghino, S.; Kohler, A.; Morin, E.; Khouja, H.-R.; Daguerre, Y.; Veneault-Fourrey, C.; Martin, F.M.; Perotto, S.; Martino, E. The Hydrophobin-Like OmSSP1 May Be an Effector in the Ericoid Mycorrhizal Symbiosis. Front. Plant Sci. 2018, 9, 546. [Google Scholar] [CrossRef]
- Guzmán-Guzmán, P.; Alemán-Duarte, M.I.; Delaye, L.; Herrera-Estrella, A.; Olmedo-Monfil, V. Identification of effector-like proteins in Trichoderma spp. and role of a hydrophobin in the plant-fungus interaction and mycoparasitism. BMC Genetics 2017, 18. [Google Scholar] [CrossRef]
- Laur, J.; Ramakrishnan, G.B.; Labbé, C.; Lefebvre, F.; Spanu, P.D.; Bélanger, R.R. Effectors involved in fungal-fungal interaction lead to a rare phenomenon of hyperbiotrophy in the tritrophic system biocontrol agent-powdery mildew-plant. New Phytol. 2018, 217, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Valdespino, C.A.; Casas-Flores, S.; Olmedo-Monfil, V. Trichoderma as a Model to Study Effector-Like Molecules. Front. Microbiol. 2019, 10, 1030. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, D.; Rovenich, H.; Jeena, G.; Nizam, S.; Tissier, A.; Balcke, G.U.; Mahdi, L.; Bonkowski, M.; Langen, G.; Zuccaro, A. The inconspicuous gatekeeper: Endophytic Serendipita vermifera acts as extended plant protection barrier in the rhizosphere. New Phytol. 2019, 224, 886–901. [Google Scholar] [CrossRef]
- Voß, S.; Betz, R.; Heidt, S.; Corradi, N.; Requena, N. RiCRN1, a Crinkler Effector From the Arbuscular Mycorrhizal Fungus Rhizophagus irregularis, Functions in Arbuscule Development. Front. Microbiol. 2018, 9, 2068. [Google Scholar] [CrossRef]
- Xu, Y.; Zhan, J.; Wijeratne, E.M.K.; Burns, A.M.; Gunatilaka, A.A.L.; Molnár, I. Cytotoxic and Antihaptotactic Beauvericin Analogues from Precursor-Directed Biosynthesis with the Insect Pathogen Beauveria bassiana ATCC 7159. J. Nat. Prod. 2007, 70, 1467–1471. [Google Scholar] [CrossRef] [PubMed]
- Bignell, D.R.D.; Fyans, J.K.; Cheng, Z. Phytotoxins produced by plant pathogenic Streptomyces species. J. Appl. Microbiol. 2014, 116, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.-C.; Hui, S.; Choi, J.; Asiegbu, F.O.; Valkonen, J.P.T.; Lee, Y.-H. Secret lifestyles of Neurospora crassa. Sci. Rep. 2015, 4, 5135. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Program | Features | Website | Reference |
|---|---|---|---|
| Perl/Bioperl | International association of users and developers of open-source Perl tools for bioinformatics, genomics and life science. | https://bioperl.org/ | [44] |
| SignalP 4.1 | Predicts the presence of signal peptides and the location of their cleavage sites in proteins from gram-positive bacteria, gram-negative bacteria and eukarya. | http://www.cbs.dtu.dk/services/SignalP/index.php | [12] |
| Phobius | This server is for the prediction of transmembrane topology and signal peptides from the amino acid sequence of a protein. | http://phobius.sbc.su.se/ | [18] |
| WoLFPSORT | Converts protein amino acid sequences into numerical localization features, based on sorting signals, amino acid composition and functional motifs, to predict protein subcellular location. | https://wolfpsort.hgc.jp/ | [13] |
| TMHMM 2.0 | Predicts trans-membrane (TM) domain helices in proteins. | http://www.cbs.dtu.dk/services/TMHMM/ | [14] |
| Species | Effector Proteins |
|---|---|
| Alternaria alternata | Aapg1 |
| Alternaria citri | Acpg1 |
| Aspergillus flavus | PECA |
| Aspergillus fumigatus | CfmB, CfmC |
| Beauveria bassiana | BbCHIT1 |
| Bipolaris maydis | Ppt1 |
| Bipolaris oryzae | Ppt1 |
| Bipolaris zeicola | PGN1 |
| Blumeria graminis f. sp. hordei | Avrk1, Avra1, Avra13, CSEP0105, BEC1005, BEC1040 |
| Blumeria graminis f. sp. tritici | AvrPm2 |
| Botrytis cinerea | CUTA, Nep1, PGIP2, BcPGA1, BcPG2 |
| Botrytis elliptica | BeNEP2 |
| Candida albicans | BGL2, SAP1, sap1, SAP2, SAP3, sap3, Pga26, RBT4 |
| Candida tropicalis | ACP |
| Cladosporium fulvum | Avr2, Avr4, Avr4E, Avr5, Avr9, Ecp1, Ecp2, Ecp4, Ecp5, Ecp6 |
| Claviceps purpurea | CPPG1, CPPG2 |
| Colletotrichum graminicola | CgEP1, Cgfl |
| Fusarium graminearum | FGL1 |
| Fusarium oxysporum | Avr3, PG1, XYL3 |
| Fusarium oxysporum f. sp. lycopersici | Six1, Six2, Six3, Six4, Six5, Six6, Six7, Six8 |
| Fusarium solani | cutA |
| Histoplasma capsulatum | CBP1 |
| Laccaria bicolor | MiSSP7 |
| Leptosphaeria maculans | AvrLm1, AvrLm4–7, AvrLm6, AvrLm11, AvrLmJ1, SP1 |
| Magnaporthe oryzae | Avr1-CO39, Avr-Pia, AvrPib, Avr-Pita, Avr-Pii, Avr-Pik, AvrPi9, AvrPiz-t, Bas1, Bas2, Bas3, Bas4, Bas107, Bas162, GAS1, GAS2, Iug6, Iug9, MC69, MHP1, MoCDIP1, MoCDIP2, MoCDIP3, MoCDIP4, MoCDIP5, MPG1, MoHEG13, Msp1, Pwl1, SPD2, SPD4, SPD7, SPD9, SPD10, XYL-6 |
| Melampsora lini | AvrL2-A, AvrL567-A, AvrM, AvrM14, AvrP4, AvrP123, |
| Metarhizium anisopliae | Pr1 |
| Monilinia fructicola | MfCUT1 |
| Parastagonospora nodorum | SP1 |
| Phakopsora pachyrhizi | PpEC23 |
| Phytophthora cactorum | PcF |
| Phytophthora capcisi | Pc129892, Pcipg2 |
| Phytophthora infestans | EPI10, INF1, INF2A, INF2B |
| Phytophthora parasitica | CBEL, Ppxyn1 |
| Phytophthora sojae | GIP2 |
| Puccinia graminis f. sp. tritici | AvrSr50, PGTAUSPE-10-1 |
| Puccinia striiformis f. sp. tritici | Pec6, PstSCR1 |
| Pyrenophora tritici-repentis | ToxB |
| Rhynchosporium secalis | NIP1, NIP2, NIP3 |
| Sclerotinia sclerotiorum | SsSSVP1 |
| Stagonospora nodorum | ToxA, Tox1, Tox3 |
| Trichoderma virens | Sm1 |
| Uromyces fabae | RTP1 |
| Ustilago hordei | UhAvr1 |
| Ustilago maydis | Cmu1, eff1-1, Mig1, Mig2-1, Pep1, Pit2, Tin2, See1 |
| Verticillium dahliae | Ave1, PevD1, Vdlsc1, VdSCP7 |
| Zymoseptoria tritici | AvrStb6, Zt6 |
| Step-by-Step Prediction | ||||||||||||||
| Data* | Proteins in Data set | Total Proteins | Length (30—400aas) | >4 Cysteine | Signal peptide by SignalP/Phobius | Proteins without TMD with TMHMM | Total prediction | Results | Sen/Rec | Spe | PPV/Prec | ACC | FPR | F1 score |
| Set 1 | 150 | 4680 | 765 | 435 | 107 | 105 | 105 | 105 | 70% | 100% | 100% | 99% | 0.00% | 0.82 |
| Set 2 | 2329 | 0 | ||||||||||||
| Set 3 | 476 | 0 | ||||||||||||
| Set 4 | 1725 | 0 | ||||||||||||
| EffHunter | ||||||||||||||
| Data* | Proteins in data set | Total proteins | Total prediction | Prediction | Sen/Rec | Spe | PPV/Prec | ACC | FPR | F1 score | ||||
| Set 1 | 150 | 4680 | 105 | 105 | 70% | 100% | 100% | 99% | 0.00% | 0.82 | ||||
| Set 2 | 2329 | 0 | ||||||||||||
| Set 3 | 476 | 0 | ||||||||||||
| Set 4 | 1725 | 0 | ||||||||||||
| EffectorP 2.0 | ||||||||||||||
| Data* | Proteins in data set | Total proteins | Total prediction | Prediction | Sen/Rec | Spe | PPV/Prec | ACC | FPR | F1 score | ||||
| Set 1 | 150 | 4680 | 166 | 102 | 68% | 98% | 61% | 97% | 1.41% | 0.64 | ||||
| Set 2 | 2329 | 41 | ||||||||||||
| Set 3 | 476 | 22 | ||||||||||||
| Set 4 | 1725 | 1 | ||||||||||||
| EffectorP 1.0 | ||||||||||||||
| Data* | Proteins in data set | Total proteins | Total prediction | Prediction | Sen/Rec | Spe | PPV/Prec | ACC | FPR | F1 score | ||||
| Set 1 | 150 | 4680 | 164 | 91 | 60% | 98% | 55% | 97% | 1.6% | 0.57 | ||||
| Set 2 | 2329 | 49 | ||||||||||||
| Set 3 | 476 | 20 | ||||||||||||
| Set 4 | 1725 | 4 | ||||||||||||
| Sonah et al. [10] (SECRETOOL and filter by length <300 amino acids) | ||||||||||||||
| Data* | Proteins in data set | Total proteins | Total prediction | Prediction | Sen/Rec | Spe | PPV/Prec | ACC | FPR | F1 score | ||||
| Set 1 | 150 | 4680 | 72 | 72 | 48% | 100% | 100% | 98% | 0.00% | 0.64 | ||||
| Set 2 | 2329 | 0 | ||||||||||||
| Set 3 | 476 | 0 | ||||||||||||
| Set 4 | 1725 | 0 | ||||||||||||
| Species | Criteria in Reference | Genome Size (Mbp) | Total Proteome | Secretome | Effectors Prediction in Reference | EffHunter Prediction | Shared | Difference in: Reference(R) EffHunter(E) | Observations for Effectors Predicted by Reference or by EffHunter | Summary of True or False Positives or Negatives, or Ambiguous in the Specific Sets of Effectors (Considering Both Predictions) |
|---|---|---|---|---|---|---|---|---|---|---|
| Blumeria graminis f. sp. hordei [38] | Secretion signal with SignalP 4.1 and SecretomeP, no TMD with TMHMM 2.0, no hits outside powdery mildews with Blastp; subcellular localization with TargetP 1.1 and GPI anchors by Big-PI. | 124.49 | 7118 | 726 | 494 | 490 | 408 | R = 86 | Negatives (86): 62 proteins are larger than 400 amino acids, 5 proteins have GPI binding and 19 proteins have no signal peptide and have TMD. Therefore, all 86 are false positive in this set. TP = 0; FP = 86 | Reference TP = 0 FP = 86 FN = 70 TN = 12 |
| E = 82 | Negatives (12): 3 proteins were predicted by TargetP 1.1 as mitochondria target, 6 proteins were predicted with GPI anchors by big-PI and 3 proteins have homologs in fungal species that are not powdery mildew. Positives (70): 70 meet all criteria of authors; Blastp retrieved homologs only in powdery mildews. TP = 70; FP = 12 | EffHunter TP = 70 FP = 12 FN = 0 TN = 86 | ||||||||
| Pseudocercospora fijiensis [39] | Secretion signal with SignalP 4.1, one or no TM domains with TMHMM 2.0, subcellular localization with TargetP 1.1 and WoLFPSORT, no GPI anchor with PredGPI, length <250aas, >2% cysteine residues | 74.1 | 13,107 | 584 | 105 | 136 | 78 | R = 27 | Negatives (15): 2 proteins are larger than 250 amino acids, 2 have no clear localization prediction with TargetP 1.1, 7 have no signal peptide, 4 have no extracellular location. *Ambiguous (12): 7 have one TMD and 5 small proteins (<60 amino acids) with less than 4 cysteine, but ≥2% cysteine. A = 12; FP = 15 | Reference TP = 0 FP = 15 FN = 32 TN = 16 A = 12 |
| E = 58 | Negatives (16): 11 proteins are GPI-anchored, 5 proteins have mitochondria target (by TargetP 1.1). Positives (32): 30 meet all criteria and have 2.0-8.2% cysteine; 2 proteins are predicted with signal peptide by SignalP 4.1 and Phobius, and as extracellular with WoLFPSORT, but TargetP 1.1 (reference) fails to predict localization. *Ambiguous (10): 10 are >200 amino acids and have 4 (but <2%) cysteine. Meet all other criteria. TP = 32; FP = 16; A = 10 | EffHunter TP = 32 FP = 16 FN = 0 TN = 15 A = 10 | ||||||||
| Mycosphaerella graminicola [40] | Size <200 amino acids; secretion signal with SignalP 4.1, one or no TM domain with TMHMM 2.0, secreted by TargetP 1.1, no GPI-anchor with big-PI, subcellular localization with WoLFPSORT and ProtComp, and no functional information | 39.7 | 10933 | 492 | 171 | 183 | 110 | R = 61 | Negatives (60): 37 proteins are larger than 200 amino acids; 3 do not have signal peptide; 6 are GPI-anchored, 14 are not predicted as secreted by ProtComp. Positives (1): Protein ID 82029 matches all criteria. This protein is not in the nonredundant set of M. graminicola database at JGI, therefore EffHunter could not analyze it. TP = 1; FP = 60 | Reference TP = 1 FP = 60 FN=50 TN=24 |
| E = 74 | Negatives (24): 4 proteins have GPI anchors; 1 has mitochondria target; 6 are predicted cytosolic, mitochondrial or nuclear by ProtComp and 13 proteins have functional annotation in PFAM Positives (50): 50 match all criteria and have no functional annotation or known protein domains. TP = 50; FP = 24 | EffHunter TP = 50 FP = 24 FN=1 TN=60 |
| Species | Lifestyle | Genome | Total Proteins | Effector Predictions | Effectors in Reference | Reference Genome | |||
|---|---|---|---|---|---|---|---|---|---|
| Mb | Coverage | EffHunter | EffectorP 2.0 | *SECRETOOL | |||||
| Alternaria brassicicola | Necrotroph | 31.03 | 120× | 10688 | 227 | 113 | 228 | 139 | [49] |
| Blumeria graminis | Biotroph | 158.94 | 13× | 6526 | 255 | 109 | 143 | 437 | [50] |
| Cladosporium fulvum | Biotroph | 61.11 | 21× | 14127 | 342 | 151 | 296 | 271 | [51] |
| Colletotrichum graminicola | Hemibiotroph | 51.6 | 9× | 12006 | 364 | 159 | 352 | 177 | [52] |
| Fusarium oxysporum | Hemibiotroph | 55.72 | 186.1× | 17726 | 474 | 256 | 361 | 364 | [53] |
| Leptosphaeria maculans | Hemibiotroph | 44.81 | 8.31× | 12469 | 290 | 162 | 263 | 529 | [54] |
| Magnaporthe oryzae | Hemibiotroph | 41.7 | 7× | 12755 | 273 | 368 | 528 | 163 | [55] |
| Mycosphaerella graminicola | Hemibiotroph | 39.7 | 8.9× | 10933 | 286 | 166 | 235 | NS | [56] |
| Phytophtora infestans | Hemibiotroph | 228.54 | 7.6× | 17787 | 355 | 404 | 343 | 563 | [57] |
| Puccinia graminis f. sp. tritici | Biotroph | 88.64 | 6.9× | 15979 | 659 | 605 | 612 | 1106 | [26] |
| Pyrenophora tritici-repentis | Necrotroph | 37.84 | 98× | 12169 | 322 | 182 | 328 | 317 | [58] |
| Ustilago maydis | Biotroph | 19.66 | 10× | 6785 | 113 | 107 | 142 | 426 | [59] |
| Species | Effector | Length | No. of Cysteine | Signal Peptide | *TMD | EffHunter | EffectorP 2.0 |
|---|---|---|---|---|---|---|---|
| Sporisorium reilianum | SAD1 | 626 | 4 | No | 0 | Non-effector | Effector |
| Zymoseptoria tritici | Mg3LysM | 232 | 9 | Yes | 0 | Effector | Non-effector |
| Blumeria graminis f. sp. hordei | BEC1054 | 118 | 2 | Yes | 0 | Non-effector | Effector |
| BEC1011 | 118 | 3 | Yes | 0 | Non-effector | Effector | |
| BEC1019 | 316 | 8 | Yes | 0 | Effector | Non-effector | |
| CSEP0055 | 122 | 3 | Yes | 0 | Non-effector | Effector | |
| Bcg1 | 146 | 2 | Yes | 0 | Non-effector | Effector | |
| CSEP0105 | 128 | 6 | Yes | 0 | Effector | Non-effector | |
| Rhizophagus irregularis | SIS1 | 149 | 2 | Yes | 1 | Non-effector | Effector |
| Fusarium graminearum | Xyla | 231 | 1 | Yes | 0 | Non-effector | Effector |
| Piriformospora indica | PIIN 08944 | 120 | 0 | Yes | 0 | Non-effector | Non-effector |
| Blumeria graminis f. sp. tritici | AvrPm3 | 130 | 2 | Yes | 0 | Non-effector | Effector |
| Puccinia graminis f. sp. tritici | AvrSr35 | 577 | 3 | Yes | 0 | Non-effector | Non-effector |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carreón-Anguiano, K.G.; Islas-Flores, I.; Vega-Arreguín, J.; Sáenz-Carbonell, L.; Canto-Canché, B. EffHunter: A Tool for Prediction of Effector Protein Candidates in Fungal Proteomic Databases. Biomolecules 2020, 10, 712. https://doi.org/10.3390/biom10050712
Carreón-Anguiano KG, Islas-Flores I, Vega-Arreguín J, Sáenz-Carbonell L, Canto-Canché B. EffHunter: A Tool for Prediction of Effector Protein Candidates in Fungal Proteomic Databases. Biomolecules. 2020; 10(5):712. https://doi.org/10.3390/biom10050712
Chicago/Turabian StyleCarreón-Anguiano, Karla Gisel, Ignacio Islas-Flores, Julio Vega-Arreguín, Luis Sáenz-Carbonell, and Blondy Canto-Canché. 2020. "EffHunter: A Tool for Prediction of Effector Protein Candidates in Fungal Proteomic Databases" Biomolecules 10, no. 5: 712. https://doi.org/10.3390/biom10050712
APA StyleCarreón-Anguiano, K. G., Islas-Flores, I., Vega-Arreguín, J., Sáenz-Carbonell, L., & Canto-Canché, B. (2020). EffHunter: A Tool for Prediction of Effector Protein Candidates in Fungal Proteomic Databases. Biomolecules, 10(5), 712. https://doi.org/10.3390/biom10050712