Circular RNA Expression Profiles in Plasma from Patients with Heart Failure Related to Platelet Activity


Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Population and Control Groups
2.2. Plasma Isolation and RNA Extraction
2.3. CircRNA Microarray Expression Profiling
2.4. CircRNA Expression Analysis
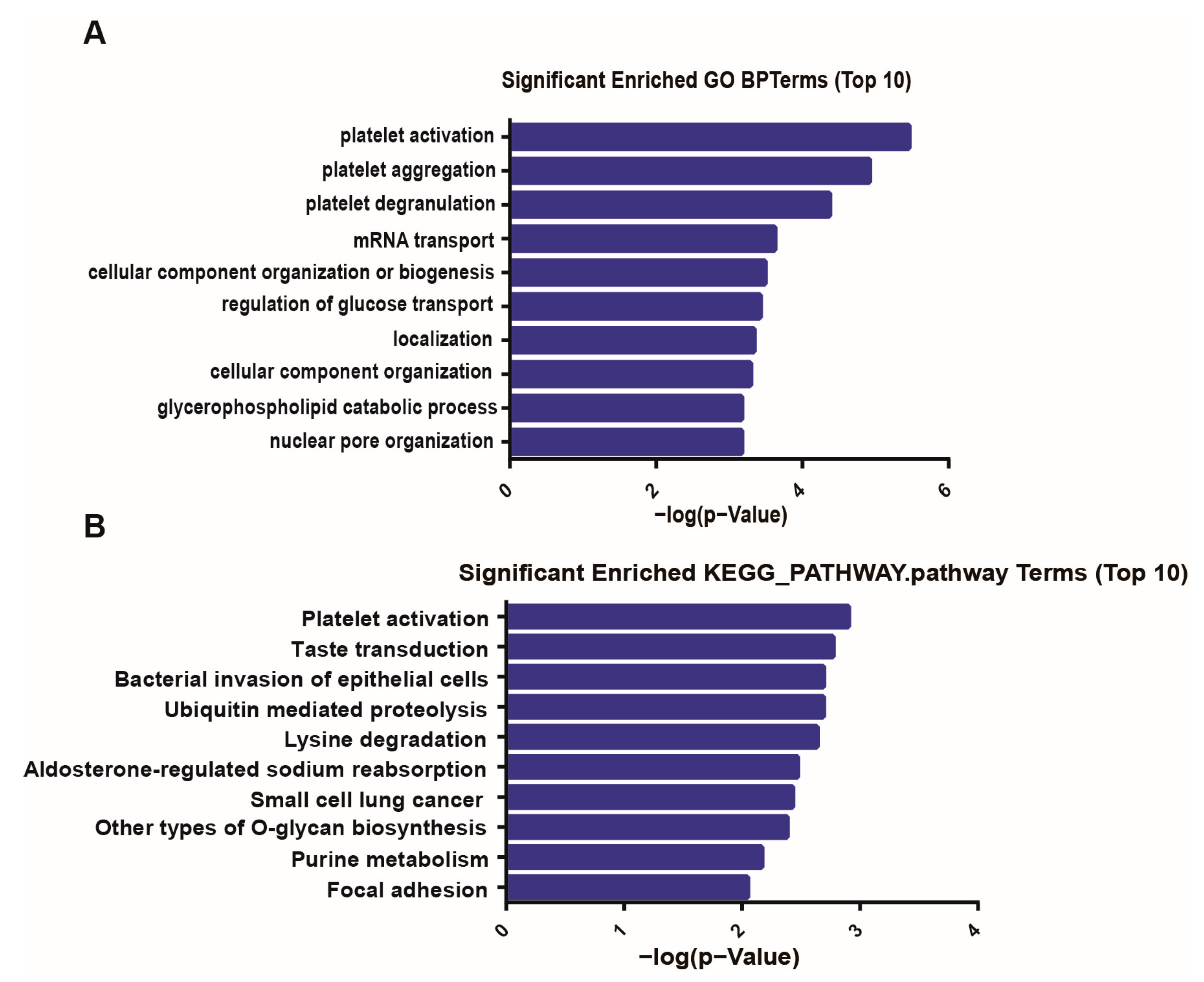
2.5. Gene Ontology and Kyoto Encyclopedia of Genes and Genomes Pathway Analyses
2.6. Statistical Analysis
3. Results
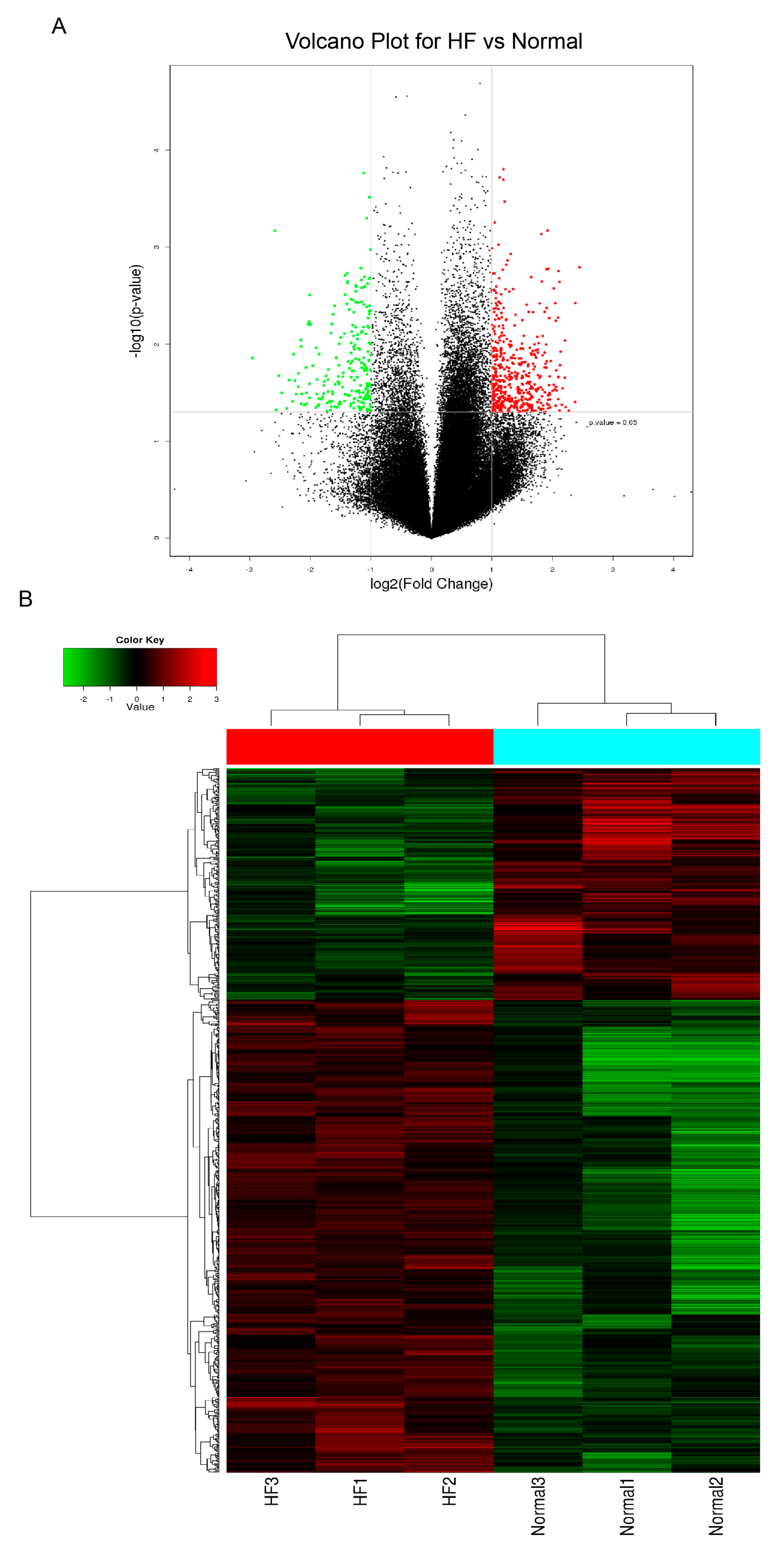
3.1. Expression Profiles of CircRNAs in the Plasma from Patients with HF
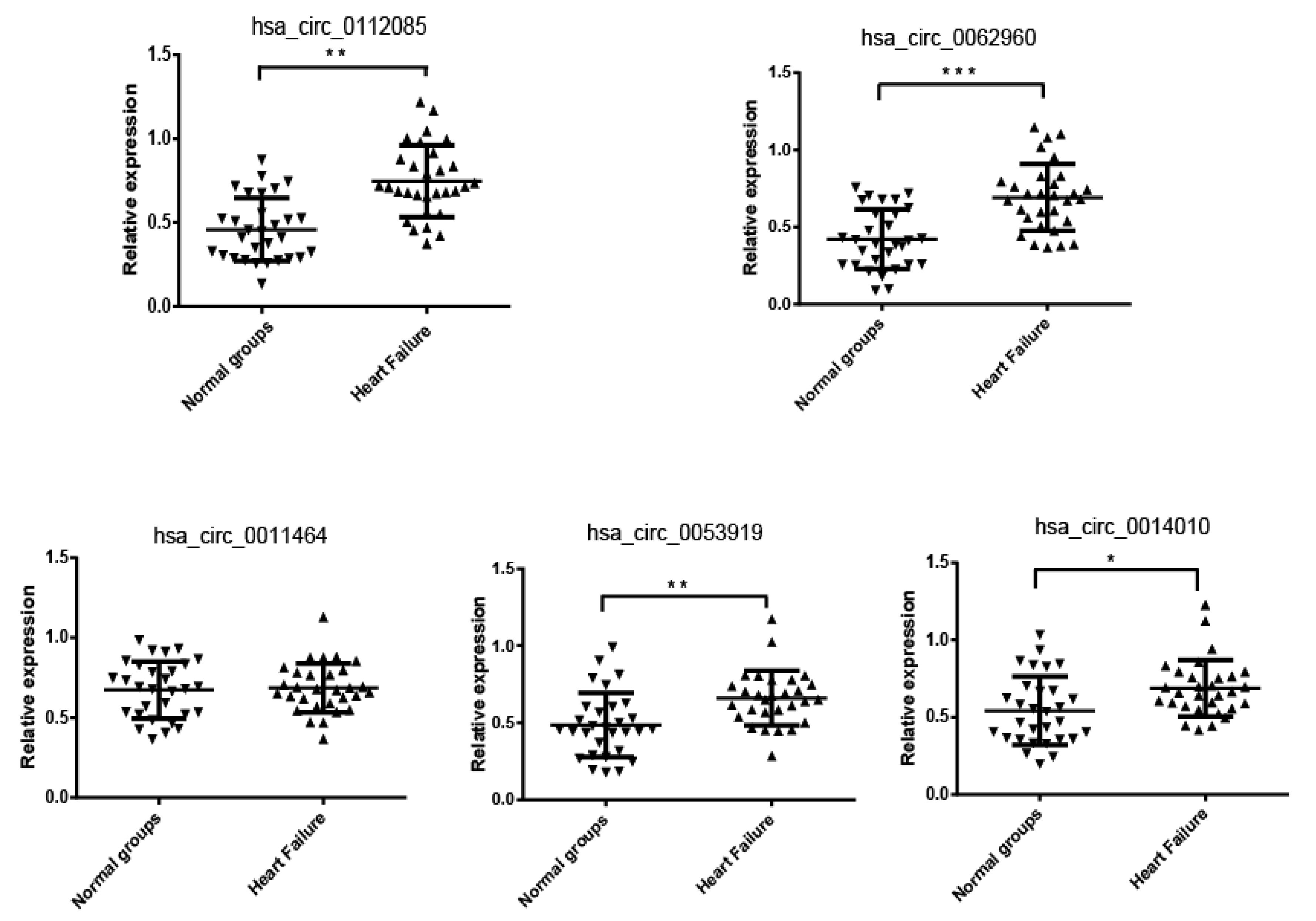
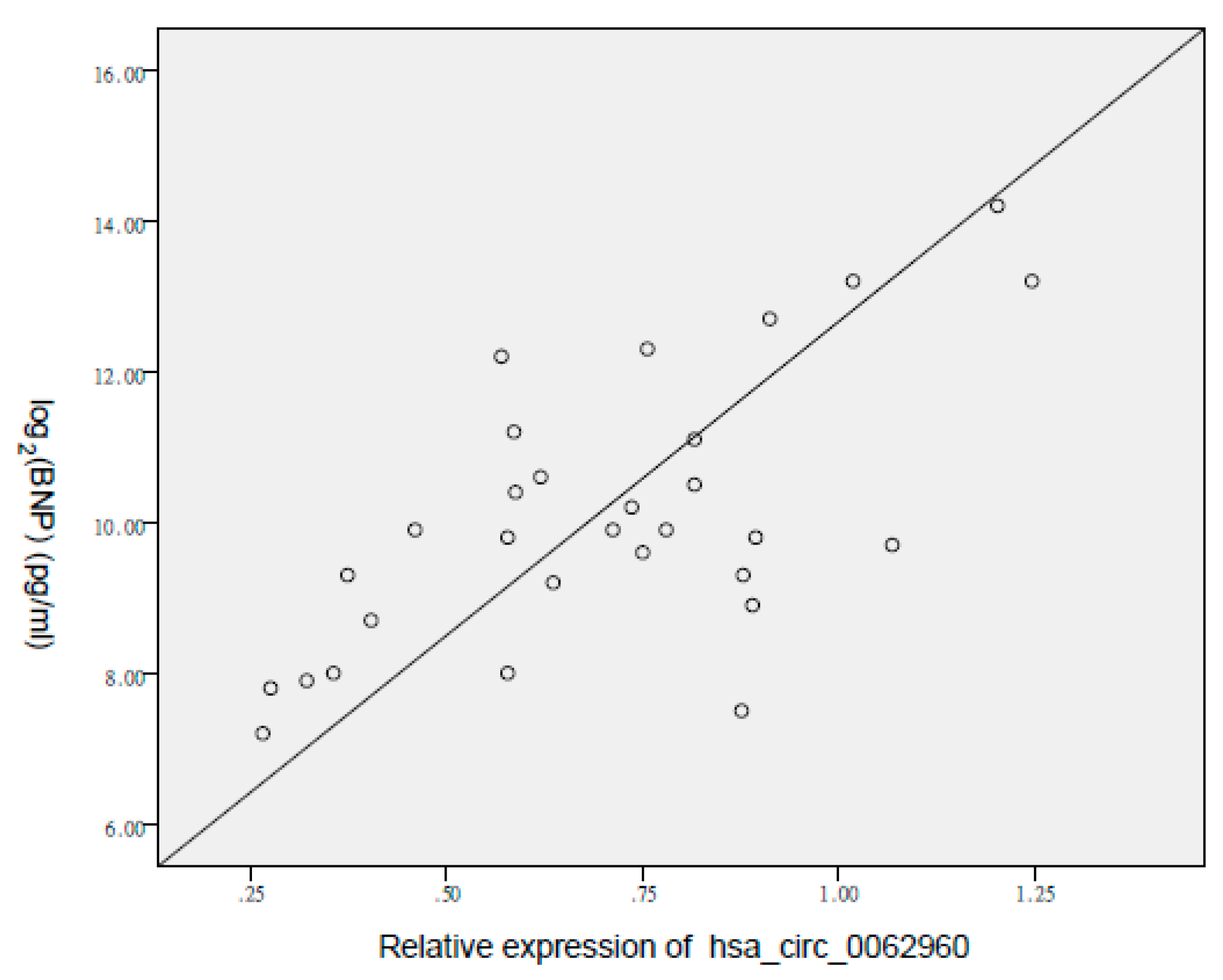
3.2. Verification of the CircRNA Profile by qRT-PCR
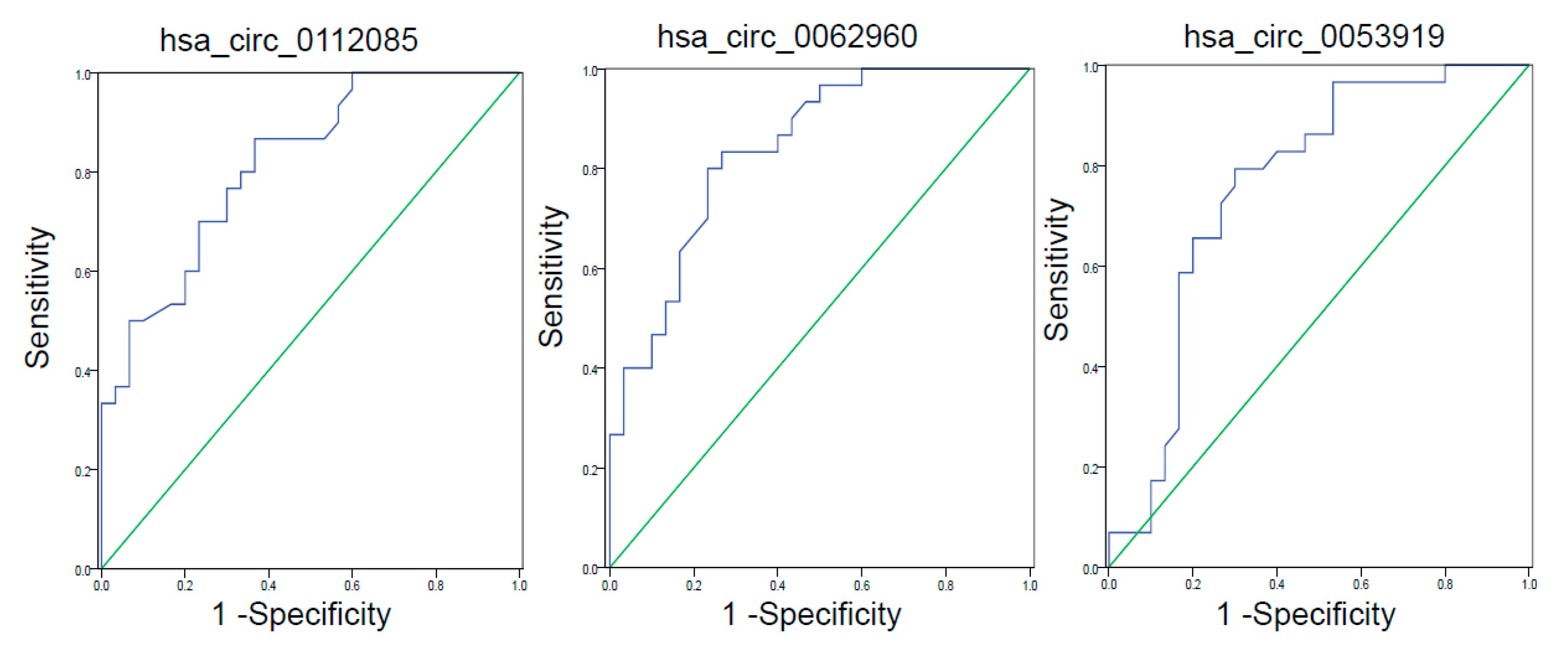
3.3. ROC Curve Analysis of circRNAs with Differential Expression
3.4. GO and KEGG Pathway Analysis of CircRNA Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chadda, K.R.; Fazmin, I.T.; Ahmad, S.; Valli, H.; Edling, C.E.; Huang, C.L.; Jeevaratnam, K. Arrhythmogenic mechanisms of obstructive sleep apnoea in heart failure patients. Sleep 2018, 41, zsy136. [Google Scholar] [CrossRef]
- Bui, A.L.; Horwich, T.B.; Fonarow, G.C. Epidemiology and risk profile of heart failure. Nat. Rev. Cardiol. 2011, 8, 30–41. [Google Scholar] [CrossRef]
- Cosentino, N.; Campodonico, J. Acute heart failure: Diagnosis first and then treatment. Int. J. Cardiol. 2018, 269, 224–225. [Google Scholar] [CrossRef] [PubMed]
- Kuo, D.C.; Peacock, W.F. Diagnosing and managing acute heart failure in the emergency department. Clin. Exp. Emerg. Med. 2015, 2, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Hamatani, Y.; Nagai, T.; Shiraishi, Y.; Kohsaka, S.; Nakai, M.; Nishimura, K.; Kohno, T.; Nagatomo, Y.; Asaumi, Y.; Goda, A.; et al. Investigators for the WETNCP: Long-Term Prognostic significance of plasma B-Type natriuretic peptide level in patients with acute heart failure with reduced, mid-range, and preserved ejection fractions. Am. J. Cardiol. 2018, 121, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Dahou, A.; Clavel, M.A.; Capoulade, R.; O’Connor, K.; Ribeiro, H.B.; Cote, N.; Le Ven, F.; Rodes-Cabau, J.; Dumesnil, J.G.; Mathieu, P.; et al. B-Type natriuretic peptide and high-sensitivity cardiac troponin for risk stratification in low-flow, low-gradient aortic stenosis: A substudy of the TOPAS study. JACC. Cardiovasc. Imaging 2018, 11, 939–947. [Google Scholar]
- Yamamoto, M.; Hayashida, M.; Kakemizu-Watanabe, M.; Ando, N.; Mukaida, H.; Kawagoe, I.; Yusuke, S.; Inada, E. B-Type Natriuretic Peptide and Hemoglobin are Two Major Factors Significantly Associated With Baseline Cerebral Oxygen Saturation Measured Using the INVOS Oximeter in Patients Undergoing Off-Pump Coronary Artery Bypass Graft Surgery. J. Cardiothorac. Vasc. Anesthesia 2018, 32, 187–196. [Google Scholar] [CrossRef]
- McKie, P.M.; Burnett, J.C. B-Type Natriuretic Peptide as a Biomarker Beyond Heart Failure: Speculations and Opportunities. Mayo Clin. Proc. 2005, 80, 1029–1036. [Google Scholar] [CrossRef]
- Balion, C.M.; Santaguida, P.; McKelvie, R.; Hill, S.A.; McQueen, M.J.; Worster, A.; Raina, P.S. Physiological, pathological, pharmacological, biochemical and hematological factors affecting BNP and NT-proBNP. Clin. Biochem. 2008, 41, 231–239. [Google Scholar] [CrossRef]
- Li, X.; Yang, L.; Chen, L.-L. The Biogenesis, Functions, and Challenges of Circular RNAs. Mol. Cell 2018, 71, 428–442. [Google Scholar] [CrossRef]
- Fu, Y.; Jiang, H.; Liu, J.-B.; Sun, X.-L.; Zhang, Z.; Li, S.; Gao, Y.; Yuan, B.; Zhang, J.-B. Genome-wide analysis of circular RNAs in bovine cumulus cells treated with BMP15 and GDF9. Sci. Rep. 2018, 8, 7944. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, L.; Han, G.; Zuo, X.; Zhang, Y.; Zhu, Q.; Wu, J.; Wang, X. A Meta-Analysis of the Diagnostic Accuracy of Circular RNAs in Digestive System Malignancy. Cell. Physiol. Biochem. 2018, 45, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Okholm, T.L.H.; Veno, M.T.; Kjems, J. Circular RNAs are abundantly expressed and upregulated during human epidermal stem cell differentiation. RNA. Biol. 2018, 15, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Devaux, Y.; Creemers, E.E.; Boon, R.A.; Werfel, S.; Thum, T.; Engelhardt, S.; Dimmeler, S.; Squire, I.; Network, O.B.O.T.C. Circular RNAs in heart failure. Eur. J. Hear. Fail. 2017, 19, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-J.; Zhang, C.-Y.; Zhang, S.; Chang, M.; Wang, H.-Y. Microarray Expression Profile of Circular RNAs in Heart Tissue of Mice with Myocardial Infarction-Induced Heart Failure. Cell. Physiol. Biochem. 2016, 39, 205–216. [Google Scholar] [CrossRef]
- Werfel, S.; Nothjunge, S.; Schwarzmayr, T.; Strom, T.-M.; Meitinger, T.; Engelhardt, S. Characterization of circular RNAs in human, mouse and rat hearts. J. Mol. Cell. Cardiol. 2016, 98, 103–107. [Google Scholar] [CrossRef]
- Memczak, S.; Papavasileiou, P.; Peters, O.; Rajewsky, N. Identification and Characterization of Circular RNAs As a New Class of Putative Biomarkers in Human Blood. PLOS ONE 2015, 10, e0141214. [Google Scholar] [CrossRef]
- Vausort, M.; Salgado-Somoza, A.; Zhang, L.; Leszek, P.; Scholz, M.; Teren, A.; Burkhardt, R.; Thiery, J.; Wagner, D.R.; Devaux, Y. Myocardial Infarction-Associated Circular RNA Predicting Left Ventricular Dysfunction. J. Am. Coll. Cardiol. 2016, 68, 1247–1248. [Google Scholar] [CrossRef]
- Sun, Y.; Jiang, X. Circular RNA Expression Profiles in the Plasma from Patients with Heart Failure; GSE134584; GEO. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE134584 (accessed on 20 July 2019).
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Meng, S.; Zhou, H.; Feng, Z.; Xu, Z.; Tang, Y.; Li, P.; Wu, M. CircRNA: Functions and properties of a novel potential biomarker for cancer. Mol. Cancer 2017, 16, 94. [Google Scholar] [CrossRef]
- Jiang, M.; Lash, G.E.; Zhao, X.; Long, Y.; Guo, C.; Yang, H. CircRNA-0004904, CircRNA-0001855, and PAPP-A: Potential Novel Biomarkers for the Prediction of Preeclampsia. Cell. Physiol. Biochem. 2018, 46, 2576–2586. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.-M.; Wen, X.; Han, X.-R.; Wang, S.; Wang, Y.-J.; Shen, M.; Fan, S.-H.; Zhang, Z.-F.; Shan, Q.; Li, M.-Q.; et al. Role of circRNA-DLEU2 in human acute myeloid leukemia. Mol. Cell. Boil. 2018, 38, e00259-18. [Google Scholar]
- Soslau, G. Circular RNA (circRNA) was an important bridge in the switch from the RNA world to the DNA world. J. Theor. Boil. 2018, 447, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, C.-X.; Xue, W.; Zhang, Y.; Jiang, S.; Yin, Q.-F.; Wei, J.; Yao, R.-W.; Yang, L.; Chen, L.-L. Coordinated circRNA Biogenesis and Function with NF90/NF110 in Viral Infection. Mol. Cell 2017, 67, 214–227.e7. [Google Scholar] [CrossRef]
- Wu, N.; Jin, L.; Cai, J. Profiling and bioinformatics analyses reveal differential circular RNA expression in hypertensive patients. Clin. Exp. Hypertens. 2017, 39, 454–459. [Google Scholar] [CrossRef]
- Zhang, S.; Zeng, X.; Ding, T.; Guo, L.; Li, Y.; Ou, S.; Yuan, H. Microarray profile of circular RNAs identifies hsa_circ_0014130 as a new circular RNA biomarker in non-small cell lung cancer. Sci. Rep. 2018, 8, 2878. [Google Scholar] [CrossRef]
- Zheng, J.; Li, Z.; Wang, T.; Zhao, Y.; Wang, Y. Microarray Expression Profile of Circular RNAs in Plasma from Primary Biliary Cholangitis Patients. Cell. Physiol. Biochem. 2017, 44, 1271–1281. [Google Scholar] [CrossRef]
- Tan, W.L.; Lim, B.T.; Anene-Nzelu, C.G.; Ackers-Johnson, M.; Dashi, A.; See, K.; Tiang, Z.; Lee, D.P.; Chua, W.W.; Luu, T.D.; et al. A landscape of circular RNA expression in the human heart. Cardiovasc. Res. 2017, 113, 298–309. [Google Scholar] [CrossRef]
- Wang, K.; Long, B.; Liu, F.; Wang, J.-X.; Liu, C.-Y.; Zhao, B.; Zhou, L.-Y.; Sun, T.; Wang, M.; Yu, T.; et al. A circular RNA protects the heart from pathological hypertrophy and heart failure by targeting miR-223. Eur. Hear. J. 2016, 37, 2602–2611. [Google Scholar] [CrossRef]
- Du, W.W.; Yang, W.; Chen, Y.; Wu, Z.K.; Foster, F.S.; Yang, Z.; Li, X.; Yang, B.B. Foxo3 circular RNA promotes cardiac senescence by modulating multiple factors associated with stress and senescence responses. Eur. Heart, J. 2017, 38, 1402–1412. [Google Scholar] [CrossRef]
- Tijsen, A.J.; Creemers, E.E.; Moerland, P.D.; De Windt, L.J.; Van Der Wal, A.C.; Kok, W.E.; Pinto, Y.M. MiR423-5p As a Circulating Biomarker for Heart Failure. Circ. Res. 2010, 106, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Goren, Y.; Kushnir, M.; Zafrir, B.; Tabak, S.; Lewis, B.S.; Amir, O. Serum levels of microRNAs in patients with heart failure. Eur. J. Hear. Fail. 2012, 14, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Gurbel, P.A.; Gattis, W.A.; Fuzaylov, S.F.; Gaulden, L.; Hasselblad, V.; Serebruany, V.L.; O’Connor, C.M. Evaluation of platelets in heart failure: is platelet activity related to etiology, functional class, or clinical outcomes? Am. Hear. J. 2002, 143, 1068–1075. [Google Scholar] [CrossRef] [PubMed]
- Serebruany, V.; McKenzie, M.; Meister, A.; Fuzaylov, S.; Gurbel, P.; Atar, D.; Gattis, W.; O’Connor, C. Whole blood impedance aggregometry for the assessment of platelet function in patients with congestive heart failure (EPCOT Trial). Eur. J. Hear. Fail. 2002, 4, 461–467. [Google Scholar] [CrossRef]
- Serebruany, V.L.; Murugesan, S.R.; Pothula, A.; Atar, D.; Lowry, D.R.; O’Connor, C.M.; Gurbel, P.A. Increased soluble platelet/endothelial cellular adhesion molecule-1 and osteonectin levels in patients with severe congestive heart failure. Independence of disease etiology, and antecedent aspirin therapy. Eur. J. Hear. Fail. 1999, 1, 243–249. [Google Scholar] [CrossRef]
- Montenont, E.; Echagarruga, C.; Allen, N.; Araldi, E.; Suarez, Y.; Berger, J.S. Platelet WDR1 suppresses platelet activity and is associated with cardiovascular disease. Blood 2016, 128, 2033–2042. [Google Scholar] [CrossRef]
- Hughes, J.; Dawson, R.; Tea, M.; McAninch, D.; Piltz, S.; Jackson, D.; Stewart, L.; Ricos, M.G.; Dibbens, L.M.; Harvey, N.L.; et al. Knockout of the epilepsy gene Depdc5 in mice causes severe embryonic dysmorphology with hyperactivity of mTORC1 signalling. Sci. Rep. 2017, 7, 12618. [Google Scholar] [CrossRef]
- Raines, E.W.; Lane, T.F.; Iruela-Arispe, M.L.; Ross, R.; Sage, E.H. The extracellular glycoprotein SPARC interacts with platelet-derived growth factor (PDGF)-AB and -BB and inhibits the binding of PDGF to its receptors. Proc. Natl. Acad. Sci. 1992, 89, 1281–1285. [Google Scholar] [CrossRef]
- Wang, Z.; Holly, S.P.; Larson, M.K.; Liu, J.; Yuan, W.; Chrzanowska-Wodnicka, M.; White, G.C.; Parise, L.V. Rap1b is critical for glycoprotein VI-mediated but not ADP receptor-mediated α2β1 activation. J. Thromb. Haemost. 2009, 7, 693–700. [Google Scholar] [CrossRef]
- Al Hawas, R.; Ren, Q.; Ye, S.; Karim, Z.A.; Filipovich, A.H.; Whiteheart, S.W. Munc18b/STXBP2 is required for platelet secretion. Blood 2012, 120, 2493–2500. [Google Scholar] [CrossRef]
- Rao, A.K.; Freishtat, R.J.; Jalagadugula, G.; Singh, A.; Mao, G.; Wiles, A.; Cheung, P.; Boden, G. Alterations in insulin-signaling and coagulation pathways in platelets during hyperglycemia-hyperinsulinemia in healthy non-diabetic subject. Thromb. Res. 2014, 134, 704–710. [Google Scholar] [CrossRef] [PubMed]
- Zainaghi, I.A.; Forlenza, O.V.; Gattaz, W.F. Abnormal APP processing in platelets of patients with Alzheimer’s disease: Correlations with membrane fluidity and cognitive decline. Psychopharmacol. 2007, 192, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Schedel, A.; Kaiser, K.; Uhlig, S.; Lorenz, F.; Sarin, A.; Starigk, J.; Hassmann, D.; Bieback, K.; Bugert, P. Megakaryocytes and platelets express nicotinic acetylcholine receptors but nicotine does not affect megakaryopoiesis or platelet function. Platelets 2016, 27, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Falet, H.; Pollitt, A.Y.; Begonja, A.J.; Weber, S.E.; Duerschmied, D.; Wagner, D.D.; Watson, S.P.; Hartwig, J.H. A novel interaction between FlnA and Syk regulates platelet ITAM-mediated receptor signaling and function. J. Exp. Med. 2010, 207, 1967–1979. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Patients Group (n = 30) | Control Group (n = 30) | p-Value |
|---|---|---|---|
| Gender (male) | 24 (80.0%) | 22 (73.3%) | 0.65 |
| Age (year) | 65 ± 10.2 | 62 ± 8.7 | 0.39 |
| BMI | 28.7 ± 5.2 | 26.3 ± 4.8 | 0.22 |
| Diabetes | 13 (43.3%) | 2 (6.7%) | 0.008 |
| Hypertension | 16 (53.3%) | 4 (13.3%) | 0.009 |
| BNP (pg/mL) | 1070 (145–17000) | 12 (0–23.8) | <0.0001 |
| Ejection fraction (%) | 33 (25–39) | 59 (52–67) | <0.001 |
| NYHA grade | NA | ||
| ClassI | 1 (3.3%) | ||
| ClassII | 4 (13.3%) | ||
| ClassIII | 20 (66.7%) | ||
| ClassIV | 5 (16.7%) |
| CircRNA ID | Location | Gene Symbol | qRT-CR primers | Product Size(bp) |
|---|---|---|---|---|
| hsa_circ_0112085 | chr1:220928288-220936392 | MARC2 | F: 5’-TATTGTGGTGACCGGCTGTG-3′ R: 5′-TGTTTGAGGAAGGCTGCTTG-3′ | 192 |
| hsa_circ_0062960 | chr22:32242819-32242953 | DEPDC5 | F: 5′-GGGAAGAAGGGAACCTCAGC-3′ R: 5′-GGGGTGCAGTTGACTCCAGA-3′ | 129 |
| hsa_circ_0011464 | chr1:33557650-33560314 | ADC | F:5′-CGGCTCGTGTTTGAAATGGG-3′ R:5′-GGCGCAGATGATCTTACTGG-3′ | 162 |
| hsa_circ_0053919 | chr2:33488360-33586582 | LTBP1 | F:5′-CTGAAGCTGGTGGTGAGAAC-3′ R:5′-ACTTGGCTGGCACTAGACGT-3′ | 103 |
| hsa_circ_0014010 | chr1:150312873-150318612 | PRPF3 | F:5′-GGATGTCAACGTGGTAGTAGTGG-3′ R:5′-GTGCTTCCCTCCTTGTTTGT-3′ | 196 |
| circRNA | AUC | 95% CI | p-value | Sensitivity | Specificity |
|---|---|---|---|---|---|
| hsa_circ_0112085 | 0.817 | 0.713–0.921 | <0.0001 | 0.867 | 0.533 |
| hsa_circ_0062960 | 0.838 | 0.740–0.937 | <0.0001 | 0.867 | 0.400 |
| hsa_circ_0053919 | 0.759 | 0.631–0.887 | 0.001 | 0.862 | 0.467 |
| CircRNAID | Chromsome | CircStart | CircEnd | Strand | Gene | Ensembl ID | Reference |
|---|---|---|---|---|---|---|---|
| hsa_circ_0069197 | chr4 | 10075962 | 10099515 | - | WDR1 | ENSG00000071127 | [37] |
| hsa-circRNA14488-21 | chr5 | 151054172 | 151055762 | - | SPARC | ENSG00000113140 | [39] |
| hsa_circ_0027463 | chr12 | 69044179 | 69045831 | + | RAP1B | ENSG00000127314 | [40] |
| hsa_circ_0088697 | chr9 | 130438922 | 130439032 | + | STXBP1 | ENSG00000136854 | [41] |
| hsa-circRNA11337-4 | chr17 | 8808093 | 8812491 | - | PIK3R5 | ENSG00000141506 | [42] |
| hsa_circ_0001966 | chr21 | 27347382 | 27372497 | - | APP | ENSG00000142192 | [43] |
| hsa_circ_0085166 | chr8 | 101936362 | 101936526 | - | YWHAZ | ENSG00000164924 | [44] |
| hsa_circ_0091966 | chrX | 153587613 | 153590516 | - | FLNA | ENSG00000196924 | [45] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Jiang, X.; Lv, Y.; Liang, X.; Zhao, B.; Bian, W.; Zhang, D.; Jiang, J.; Zhang, C. Circular RNA Expression Profiles in Plasma from Patients with Heart Failure Related to Platelet Activity. Biomolecules 2020, 10, 187. https://doi.org/10.3390/biom10020187
Sun Y, Jiang X, Lv Y, Liang X, Zhao B, Bian W, Zhang D, Jiang J, Zhang C. Circular RNA Expression Profiles in Plasma from Patients with Heart Failure Related to Platelet Activity. Biomolecules. 2020; 10(2):187. https://doi.org/10.3390/biom10020187
Chicago/Turabian StyleSun, Yeying, Xiaoli Jiang, Yan Lv, Xinyue Liang, Bingrui Zhao, Weihua Bian, Daolai Zhang, Jing Jiang, and Chunxiang Zhang. 2020. "Circular RNA Expression Profiles in Plasma from Patients with Heart Failure Related to Platelet Activity" Biomolecules 10, no. 2: 187. https://doi.org/10.3390/biom10020187
APA StyleSun, Y., Jiang, X., Lv, Y., Liang, X., Zhao, B., Bian, W., Zhang, D., Jiang, J., & Zhang, C. (2020). Circular RNA Expression Profiles in Plasma from Patients with Heart Failure Related to Platelet Activity. Biomolecules, 10(2), 187. https://doi.org/10.3390/biom10020187