Co-Application of Eugenol and QX-314 Elicits the Prolonged Blockade of Voltage-Gated Sodium Channels in Nociceptive Trigeminal Ganglion Neurons

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of TG Neurons
2.3. Whole-Cell Patch-Clamp Recordings
2.4. Drugs
2.5. Statistical Analysis
3. Results
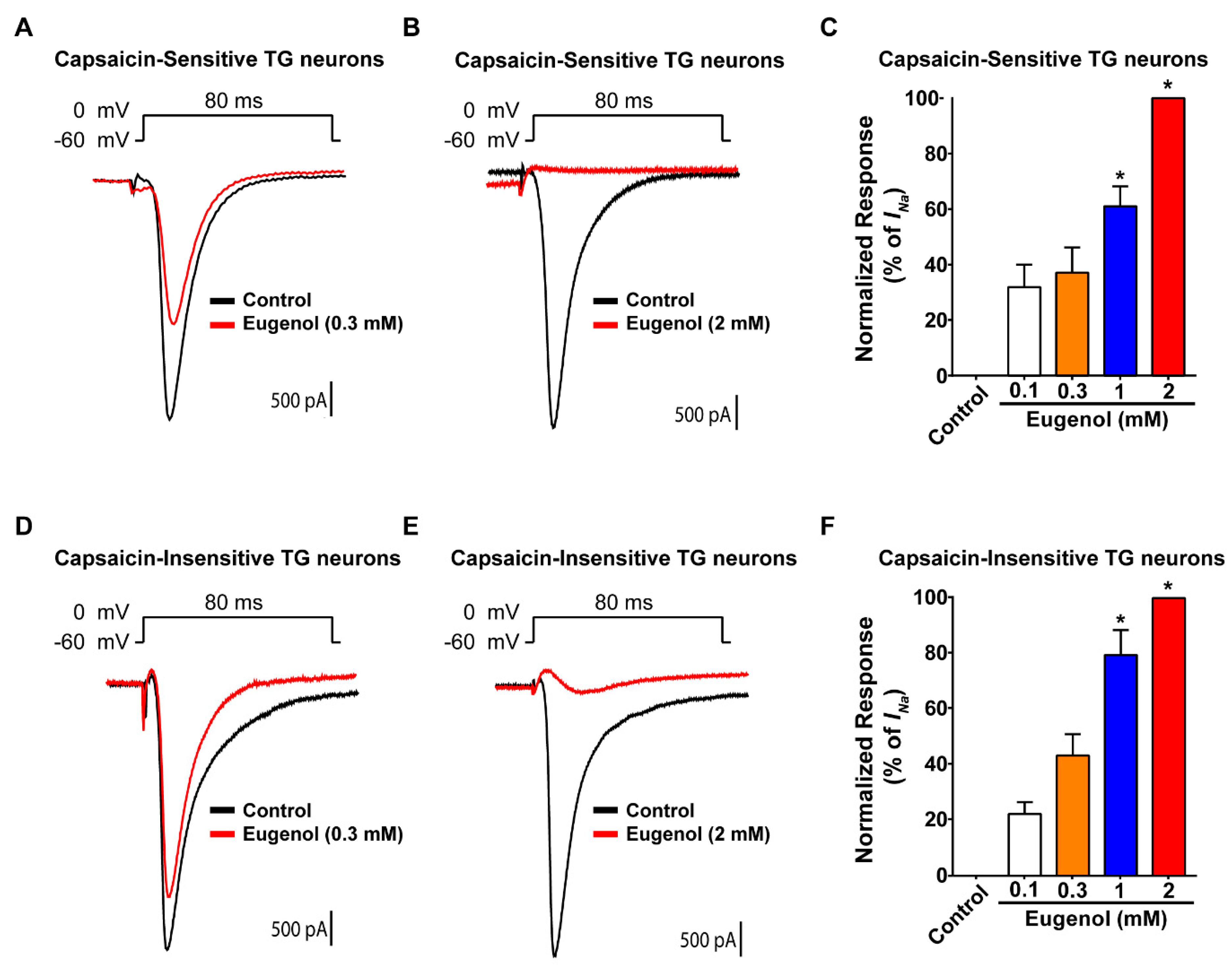
3.1. Eugenol Inhibited VGSC Currents in Small-Sized TG Neurons
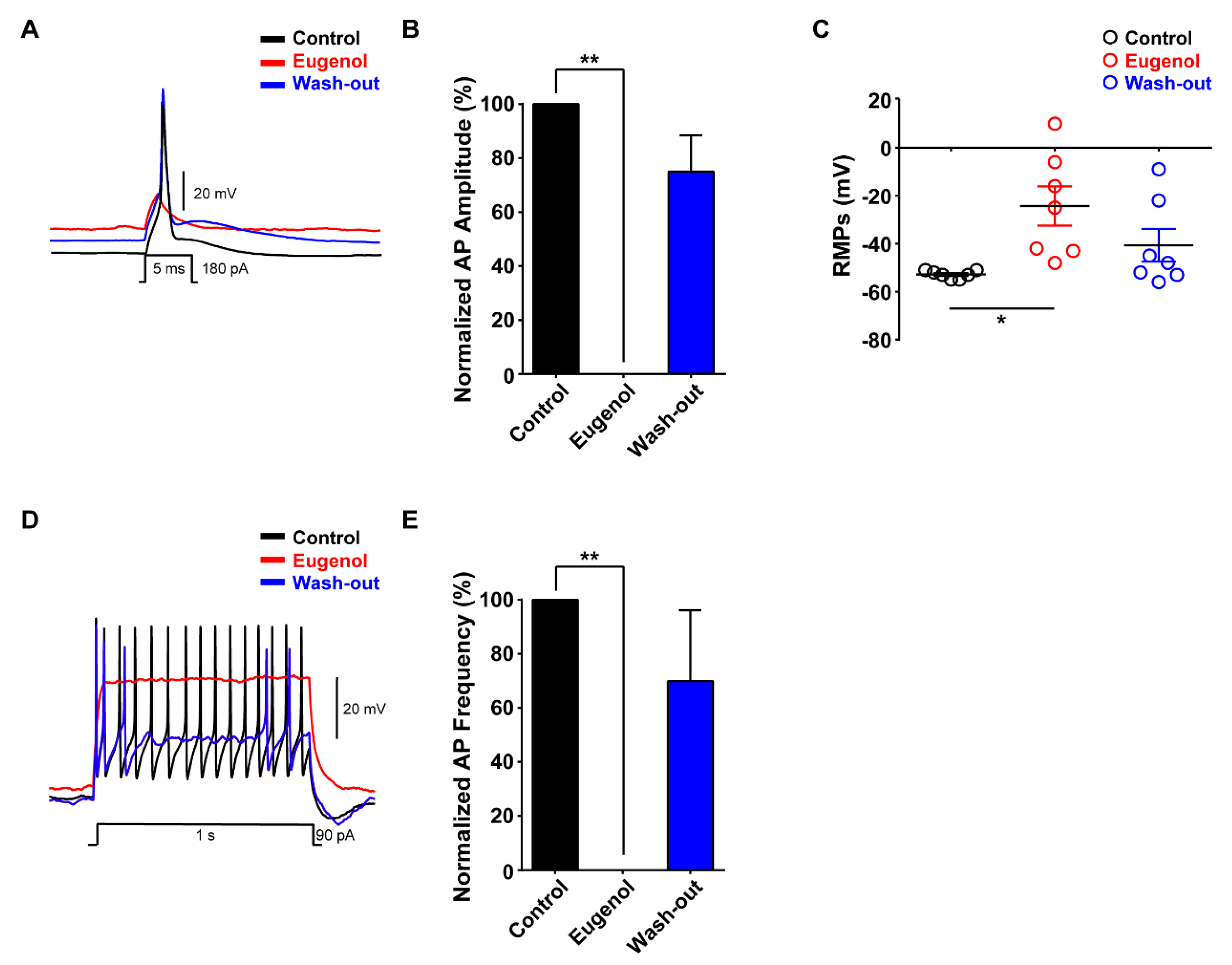
3.2. Eugenol Inhibited Aps in Small-Sized TG Neurons
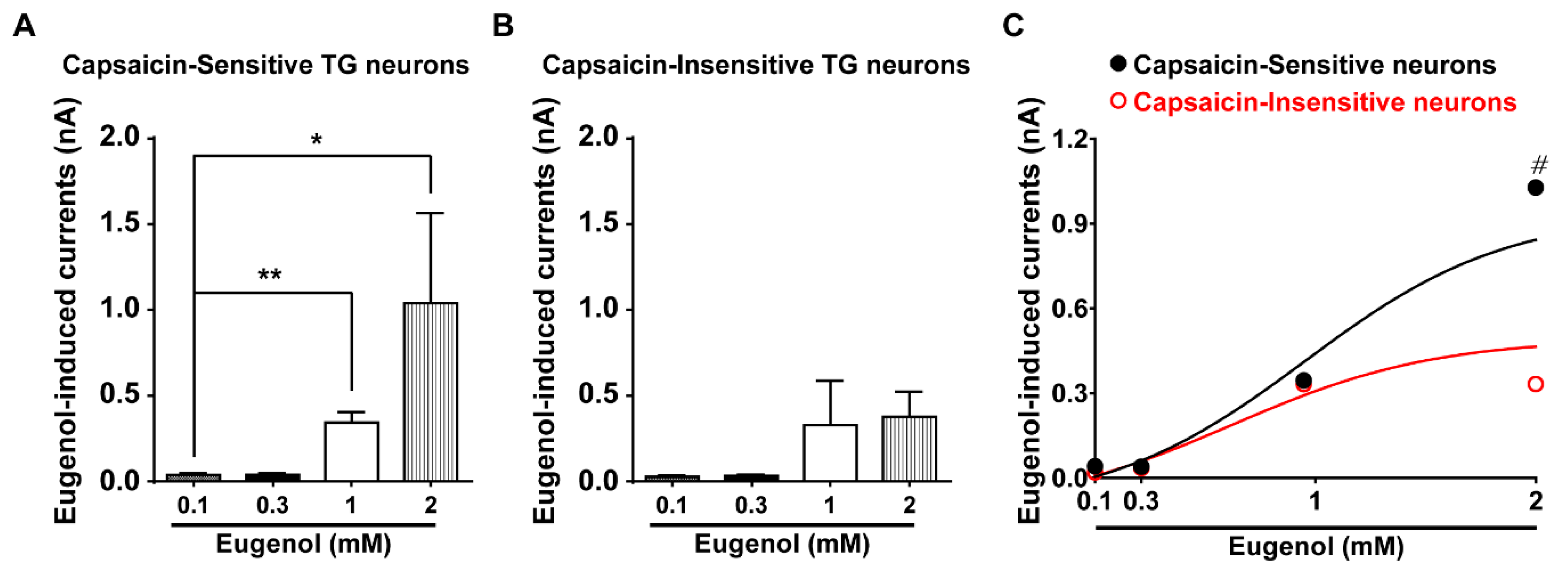
3.3. Eugenol Activated Inward Current in Small-Sized TG Neurons
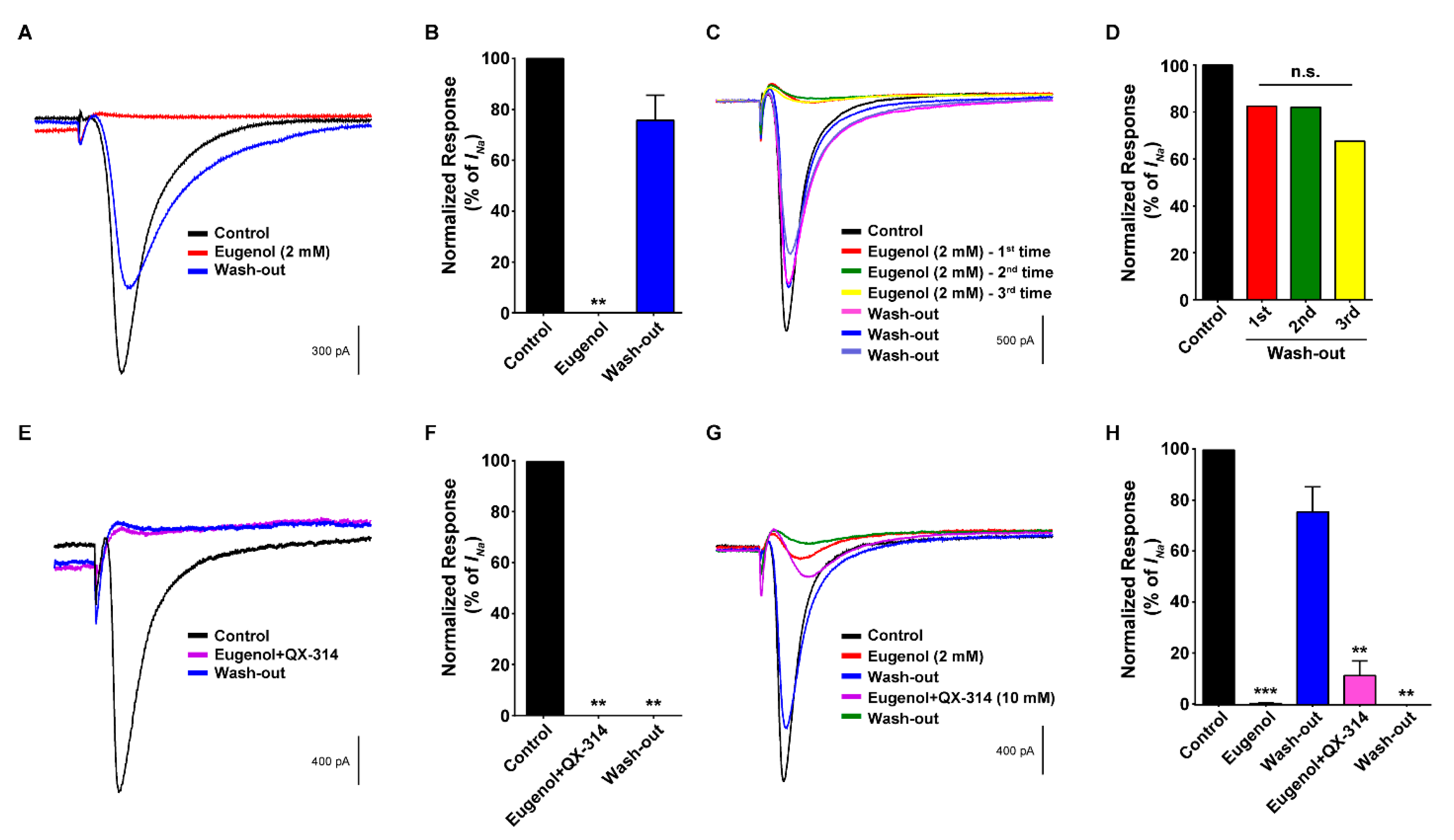
3.4. Co-Administration of Eugenol and QX-314 Blocked VGSC Currents in Small-Sized TG Neurons
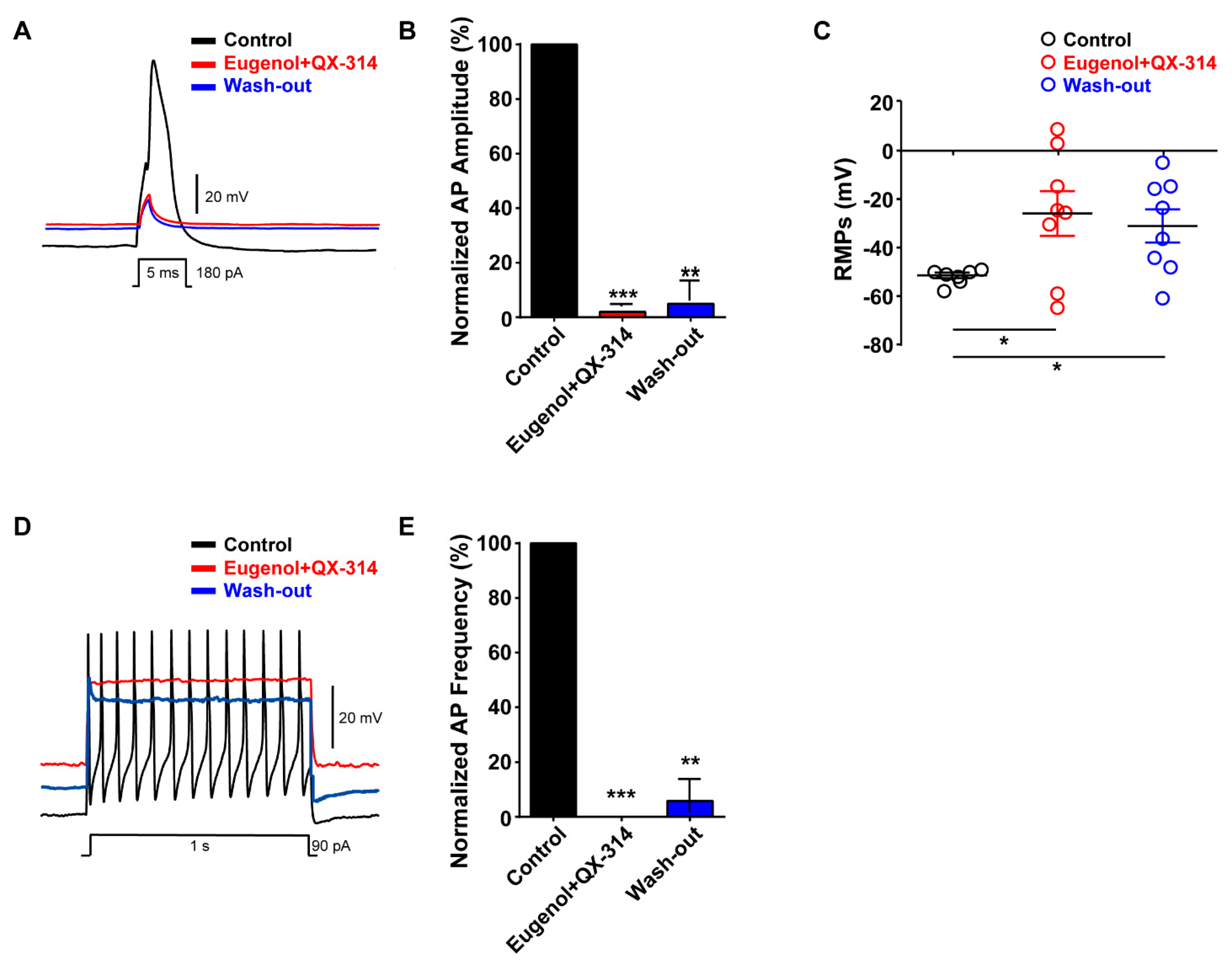
3.5. Co-Administration of Eugenol and QX-314 Blocked Aps in Small-Sized TG Neurons
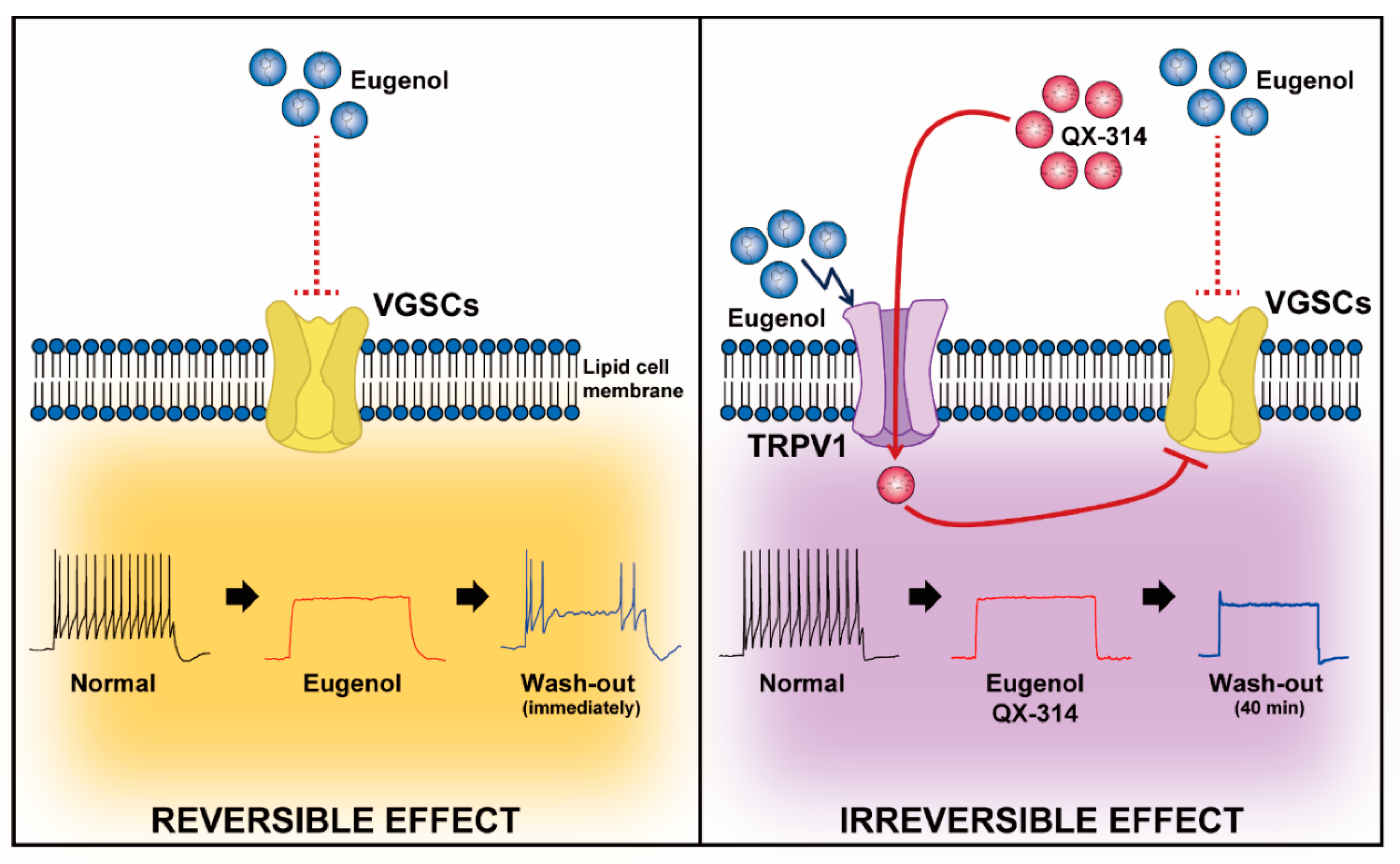
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Roberson, D.P.; Binshtok, A.M.; Blasl, F.; Bean, B.P.; Woolf, C.J. Targeting of sodium channel blockers into nociceptors to produce long-duration analgesia: A systematic study and review. Br. J. Pharmacol. 2011, 164, 48–58. [Google Scholar] [CrossRef]
- Lee, H.S. Recent advances in topical anesthesia. J. Dent. Anesth. Pain Med. 2016, 16, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Acevedo, R.E.; Pless, S.A.; Ahern, C.A.; Schwarz, S.K. The quaternary lidocaine derivative, QX-314, exerts biphasic effects on transient receptor potential vanilloid subtype 1 channels in vitro. Anesthesiology 2011, 114, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Binshtok, A.M.; Gerner, P.; Oh, S.B.; Puopolo, M.; Suzuki, S.; Roberson, D.P.; Herbert, T.; Wang, C.F.; Kim, D.; Chung, G.; et al. Coapplication of lidocaine and the permanently charged sodium channel blocker QX-314 produces a long-lasting nociceptive blockade in rodents. Anesthesiology 2009, 111, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Ellemann, K.; Sjogren, P.; Banning, A.M.; Jensen, T.S.; Smith, T.; Geertsen, P. Trial of intravenous lidocaine on painful neuropathy in cancer patients. Clin. J. Pain 1989, 5, 291–294. [Google Scholar] [CrossRef]
- Ries, C.R.; Pillai, R.; Chung, C.C.; Wang, J.T.; MacLeod, B.A.; Schwarz, S.K. QX-314 produces long-lasting local anesthesia modulated by transient receptor potential vanilloid receptors in mice. Anesthesiology 2009, 111, 122–126. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, J.; Yin, Q.; Yang, L.; Liu, J.; Zhang, W. QX-OH, a QX-314 derivative agent, produces long-acting local anesthesia in rats. Eur. J. Pharm. Sci. 2017, 105, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.; Li, J.; Zheng, Q.; Yang, X.; Lv, R.; Ma, L.; Liu, J.; Zhu, T.; Zhang, W. The quaternary lidocaine derivative QX-314 in combination with bupivacaine for long-lasting nerve block: Efficacy, toxicity, and the optimal formulation in rats. PLoS ONE 2017, 12, e0174421. [Google Scholar] [CrossRef]
- Binshtok, A.M.; Bean, B.P.; Woolf, C.J. Inhibition of nociceptors by TRPV1-mediated entry of impermeant sodium channel blockers. Nature 2007, 449, 607–610. [Google Scholar] [CrossRef]
- Kim, H.Y.; Kim, K.; Li, H.Y.; Chung, G.; Park, C.K.; Kim, J.S.; Jung, S.J.; Lee, M.K.; Ahn, D.K.; Hwang, S.J.; et al. Selectively targeting pain in the trigeminal system. Pain 2010, 150, 29–40. [Google Scholar] [CrossRef]
- Zakir, H.M.; Masuda, Y.; Kitagawa, J. A novel approach for detection of functional expression of TRPV1 channels on regenerated neurons following nerve injury. J. Oral Sci. 2020, 62, 136–139. [Google Scholar] [CrossRef]
- Pavithra, B. Eugenol—A Review. J. Pharm. Sci. Res. 2014, 6, 153–154. [Google Scholar]
- Park, C.K.; Kim, K.; Jung, S.J.; Kim, M.J.; Ahn, D.K.; Hong, S.D.; Kim, J.S.; Oh, S.B. Molecular mechanism for local anesthetic action of eugenol in the rat trigeminal system. Pain 2009, 144, 84–94. [Google Scholar] [CrossRef]
- Li, H.Y.; Lee, B.K.; Kim, J.S.; Jung, S.J.; Oh, S.B. Eugenol Inhibits ATP-induced P2X Currents in Trigeminal Ganglion Neurons. Korean J. Physiol. Pharmacol. 2008, 12, 315–321. [Google Scholar] [CrossRef]
- Park, C.K.; Li, H.Y.; Yeon, K.Y.; Jung, S.J.; Choi, S.Y.; Lee, S.J.; Lee, S.; Park, K.; Kim, J.S.; Oh, S.B. Eugenol inhibits sodium currents in dental afferent neurons. J. Dent. Res. 2006, 85, 900–904. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Moon, J.Y.; Jung, S.J.; Kang, J.G.; Choi, S.P.; Jang, J.H. Eugenol inhibits the GABAA current in trigeminal ganglion neurons. PLoS ONE 2015, 10, e0117316. [Google Scholar] [CrossRef]
- Vatanparast, J.; Khalili, S.; Naseh, M. Dual effects of eugenol on the neuronal excitability: An in vitro study. Neurotoxicology 2017, 58, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, C.M.; Woodruff, A.; Yang, R.; Kohane, D.S. Drug delivery systems for prolonged duration local anesthesia. Mater. Today 2017, 20, 22–31. [Google Scholar] [CrossRef]
- Zhou, C.; Liang, P.; Liu, J.; Zhang, W.; Liao, D.; Chen, Y.; Chen, X.; Li, T. Emulsified isoflurane enhances thermal transient receptor potential vanilloid-1 channel activation-mediated sensory/nociceptive blockade by QX-314. Anesthesiology 2014, 121, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Yeon, K.Y.; Chung, G.; Kim, Y.H.; Hwang, J.H.; Davies, A.J.; Park, M.K.; Ahn, D.K.; Kim, J.S.; Jung, S.J.; Oh, S.B. Eugenol reverses mechanical allodynia after peripheral nerve injury by inhibiting hyperpolarization-activated cyclic nucleotide-gated (HCN) channels. Pain 2011, 152, 2108–2116. [Google Scholar] [CrossRef]
- Hameed, S. Nav1.7 and Nav1.8: Role in the pathophysiology of pain. Mol. Pain 2019, 15, 1744806919858801. [Google Scholar] [CrossRef]
- Mickle, A.D.; Shepherd, A.J.; Mohapatra, D.P. Sensory TRP channels: The key transducers of nociception and pain. Prog. Mol. Biol. Transl. Sci. 2015, 131, 73–118. [Google Scholar]
- Giniatullin, R. Ion Channels of Nociception. Int. J. Mol. Sci. 2020, 21, 3553. [Google Scholar] [CrossRef] [PubMed]
- Deval, E.; Lingueglia, E. Acid-Sensing Ion Channels and nociception in the peripheral and central nervous systems. Neuropharmacology 2015, 94, 49–57. [Google Scholar] [CrossRef]
- Dube, G.R.; Elagoz, A.; Mangat, H. Acid sensing ion channels and acid nociception. Curr. Pharm. Des. 2009, 15, 1750–1766. [Google Scholar] [CrossRef]
- Su, X.; Leon, L.A.; Laping, N.J. Role of spinal Cav2.2 and Cav2.1 ion channels in bladder nociception. J. Urol. 2008, 179, 2464–2469. [Google Scholar] [CrossRef]
- Moran, M.M.; Szallasi, A. Targeting nociceptive transient receptor potential channels to treat chronic pain: Current state of the field. Br. J. Pharmacol. 2018, 175, 2185–2203. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Walters, E.T.; Carlton, S.M.; Hu, H. Targeting Pain-evoking Transient Receptor Potential Channels for the Treatment of Pain. Curr. Neuropharmacol. 2013, 11, 652–663. [Google Scholar] [CrossRef]
- Patapoutian, A.; Tate, S.; Woolf, C.J. Transient receptor potential channels: Targeting pain at the source. Nat. Rev. Drug Discov. 2009, 8, 55–68. [Google Scholar] [CrossRef]
- Sun, S.; Sun, J.; Jiang, W.; Wang, W.; Ni, L. Nav1.7 via Promotion of ERK in the Trigeminal Ganglion Plays an Important Role in the Induction of Pulpitis Inflammatory Pain. Biomed. Res. Int. 2019, 2019, 6973932. [Google Scholar] [CrossRef] [PubMed]
- Bennett, D.L.; Clark, A.J.; Huang, J.; Waxman, S.G.; Dib-Hajj, S.D. The Role of Voltage-Gated Sodium Channels in Pain Signaling. Physiol. Rev. 2019, 99, 1079–1151. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.; Porreca, F.; Hunter, J.C.; Gold, M.S. Voltage-gated sodium channels and hyperalgesia. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 371–397. [Google Scholar] [CrossRef]
- Yagura, S.; Onimaru, H.; Kanzaki, K.; Izumizaki, M. Inhibitory effects of eugenol on putative nociceptive response in spinal cord preparation isolated from neonatal rats. Exp. Brain Res. 2018, 236, 1767–1774. [Google Scholar] [CrossRef]
- Lee, M.H.; Yeon, K.Y.; Park, C.K.; Li, H.Y.; Fang, Z.; Kim, M.S.; Choi, S.Y.; Lee, S.J.; Lee, S.; Park, K.; et al. Eugenol inhibits calcium currents in dental afferent neurons. J. Dent. Res. 2005, 84, 848–851. [Google Scholar] [CrossRef]
- Seo, H.; Li, H.Y.; Perez-Reyes, E.; Lee, J.H. Effects of eugenol on T-type Ca2+ channel isoforms. J. Pharmacol. Exp. Ther. 2013, 347, 310–317. [Google Scholar] [CrossRef]
- Li, H.Y.; Park, C.K.; Jung, S.J.; Choi, S.Y.; Lee, S.J.; Park, K.; Kim, J.S.; Oh, S.B. Eugenol inhibits K+ currents in trigeminal ganglion neurons. J. Dent. Res. 2007, 86, 898–902. [Google Scholar] [CrossRef]
- Docherty, R.J.; Farmer, C.E. The pharmacology of voltage-gated sodium channels in sensory neurones. Handb. Exp. Pharmacol. 2009, 194, 519–561. [Google Scholar]
- Im, S.T.; Jo, Y.Y.; Han, G.; Jo, H.J.; Kim, Y.H.; Park, C.K. Dexmedetomidine Inhibits Voltage-Gated Sodium Channels via alpha2-Adrenoceptors in Trigeminal Ganglion Neurons. Mediat. Inflamm. 2018, 2018, 1782719. [Google Scholar] [CrossRef]
- Cho, J.S.; Kim, T.H.; Lim, J.M.; Song, J.H. Effects of eugenol on Na+ currents in rat dorsal root ganglion neurons. Brain Res. 2008, 1243, 53–62. [Google Scholar] [CrossRef]
- Chung, G.; Rhee, J.N.; Jung, S.J.; Kim, J.S.; Oh, S.B. Modulation of CaV2.3 calcium channel currents by eugenol. J. Dent. Res. 2008, 87, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Virginio, C.; MacKenzie, A.; Rassendren, F.A.; North, R.A.; Surprenant, A. Pore dilation of neuronal P2X receptor channels. Nat. Neurosci. 1999, 2, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Immke, D.C.; Gavva, N.R. The TRPV1 receptor and nociception. Semin. Cell Dev. Biol. 2006, 17, 582–591. [Google Scholar] [CrossRef]
- Jardin, I.; Lopez, J.J.; Diez, R.; Sanchez-Collado, J.; Cantonero, C.; Albarran, L.; Woodard, G.E.; Redondo, P.C.; Salido, G.M.; Smani, T.; et al. TRPs in Pain Sensation. Front. Physiol. 2017, 8, 392. [Google Scholar] [CrossRef]
- Buffum, M.D.; Hutt, E.; Chang, V.T.; Craine, M.H.; Snow, A.L. Cognitive impairment and pain management: Review of issues and challenges. J. Rehabil. Res. Dev. 2007, 44, 315–330. [Google Scholar] [CrossRef]
- Storozhuk, M.V.; Moroz, O.F.; Zholos, A.V. Multifunctional TRPV1 Ion Channels in Physiology and Pathology with Focus on the Brain, Vasculature, and Some Visceral Systems. Biomed. Res. Int. 2019, 2019, 5806321. [Google Scholar] [CrossRef] [PubMed]
- Gerner, P.; Binshtok, A.M.; Wang, C.F.; Hevelone, N.D.; Bean, B.P.; Woolf, C.J.; Wang, G.K. Capsaicin combined with local anesthetics preferentially prolongs sensory/nociceptive block in rat sciatic nerve. Anesthesiology 2008, 109, 872–878. [Google Scholar] [CrossRef]
- Haas, D.A. An update on local anesthetics in dentistry. J. Can. Dent. Assoc. 2002, 68, 546–551. [Google Scholar] [PubMed]
- Butterworth, J.F.; Strichartz, G.R. Molecular mechanisms of local anesthesia: A review. Anesthesiology 1990, 72, 711–734. [Google Scholar] [CrossRef]
- Myrdal, S.E.; Steyger, P.S. TRPV1 regulators mediate gentamicin penetration of cultured kidney cells. Hear. Res. 2005, 204, 170–182. [Google Scholar] [CrossRef]
- Brenneis, C.; Kistner, K.; Puopolo, M.; Jo, S.; Roberson, D.; Sisignano, M.; Segal, D.; Cobos, E.J.; Wainger, B.J.; Labocha, S.; et al. Bupivacaine-induced cellular entry of QX-314 and its contribution to differential nerve block. Br. J. Pharmacol. 2014, 171, 438–451. [Google Scholar] [CrossRef]
- Puopolo, M.; Binshtok, A.M.; Yao, G.L.; Oh, S.B.; Woolf, C.J.; Bean, B.P. Permeation and block of TRPV1 channels by the cationic lidocaine derivative QX-314. J. Neurophysiol. 2013, 109, 1704–1712. [Google Scholar] [CrossRef]
- Stueber, T.; Eberhardt, M.J.; Hadamitzky, C.; Jangra, A.; Schenk, S.; Dick, F.; Stoetzer, C.; Kistner, K.; Reeh, P.W.; Binshtok, A.M.; et al. Quaternary Lidocaine Derivative QX-314 Activates and Permeates Human TRPV1 and TRPA1 to Produce Inhibition of Sodium Channels and Cytotoxicity. Anesthesiology 2016, 124, 1153–1165. [Google Scholar] [CrossRef]
- Zakir, H.M.; Mostafeezur, R.M.; Suzuki, A.; Hitomi, S.; Suzuki, I.; Maeda, T.; Seo, K.; Yamada, Y.; Yamamura, K.; Lev, S.; et al. Expression of TRPV1 channels after nerve injury provides an essential delivery tool for neuropathic pain attenuation. PLoS ONE 2012, 7, e44023. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Fox, L.E.; Cheng, J. Differential effects of peripheral versus central coadministration of QX-314 and capsaicin on neuropathic pain in rats. Anesthesiology 2012, 117, 365–380. [Google Scholar] [CrossRef]
- Schwarz, S.K.; Cheung, H.M.; Ries, C.R.; Lee, S.M.; Wang, J.T.; MacLeod, B.A. Lumbar intrathecal administration of the quaternary lidocaine derivative, QX-314, produces irritation and death in mice. Anesthesiology 2010, 113, 438–444. [Google Scholar] [CrossRef]
- Koo, C.H.; Shin, H.J.; Han, S.H.; Ryu, J.H. Lidocaine vs. Other Local Anesthetics in the Development of Transient Neurologic Symptoms (TNS) Following Spinal Anesthesia: A Meta-Analysis of Randomized Controlled Trials. J. Clin. Med. 2020, 9, 493. [Google Scholar] [CrossRef] [PubMed]
- Barletta, M.; Reed, R. Local Anesthetics: Pharmacology and Special Preparations. Vet. Clin. N. Am. Small Anim. Pract. 2019, 49, 1109–1125. [Google Scholar] [CrossRef] [PubMed]
- Vadhanan, P.; Tripaty, D.K.; Adinarayanan, S. Physiological and pharmacologic aspects of peripheral nerve blocks. J. Anaesthesiol. Clin. Pharmacol. 2015, 31, 384–393. [Google Scholar] [CrossRef]
- Becker, D.E.; Reed, K.L. Local anesthetics: Review of pharmacological considerations. Anesth. Prog. 2012, 59, 90–101. [Google Scholar] [CrossRef]
- Swain, A.; Nag, D.S.; Sahu, S.; Samaddar, D.P. Adjuvants to local anesthetics: Current understanding and future trends. World J. Clin. Cases 2017, 5, 307–323. [Google Scholar] [CrossRef]
- Ma, L.; Mu, Y.; Zhang, Z.; Sun, Q. Eugenol promotes functional recovery and alleviates inflammation, oxidative stress, and neural apoptosis in a rat model of spinal cord injury. Restor. Neurol. Neurosci. 2018, 36, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.J.; Tabakoff, B.; Levinson, S.R.; Heinbockel, T. Inhibition of Nav1.7 channels by methyl eugenol as a mechanism underlying its antinociceptive and anesthetic actions. Acta Pharmacol. Sin. 2015, 36, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Ghofran, O.; Safari, T.; Shahraki, M.R. Effects of Eugenol on Pain Response to the Formalin Test and Plasma Antioxidant Activity in High Fructose Drinking Water in Male Rats. Int. J. Prev. Med. 2019, 10, 151–157. [Google Scholar] [PubMed]
- Chung, G.; Im, S.T.; Kim, Y.H.; Jung, S.J.; Rhyu, M.R.; Oh, S.B. Activation of transient receptor potential ankyrin 1 by eugenol. Neuroscience 2014, 261, 153–160. [Google Scholar] [CrossRef]
- Park, C.K.; Kim, M.S.; Fang, Z.; Li, H.Y.; Jung, S.J.; Choi, S.Y.; Lee, S.J.; Park, K.; Kim, J.S.; Oh, S.B. Functional expression of thermo-transient receptor potential channels in dental primary afferent neurons: Implication for tooth pain. J. Biol. Chem. 2006, 281, 17304–17311. [Google Scholar] [CrossRef]
- Nakagawa, H.; Hiura, A. Comparison of the transport of QX-314 through TRPA1, TRPM8, and TRPV1 channels. J. Pain Res. 2013, 6, 223–230. [Google Scholar] [CrossRef]
- Jabbari, N.; Eftekhari, Z.; Roodbari, N.H.; Parivar, K. Evaluation of Encapsulated Eugenol by Chitosan Nanoparticles on the aggressive model of rheumatoid arthritis. Int. Immunopharmacol. 2020, 85, 106554–106562. [Google Scholar] [CrossRef]
- Moreira Vasconcelos, C.F.; da Cunha Ferreira, N.M.; Hardy Lima Pontes, N.; de Sousa Dos Reis, T.D.; Basto Souza, R.; Aragao Catunda Junior, F.E.; Vasconcelos Aguiar, L.M.; Maranguape Silva da Cunha, R. Eugenol and its association with levodopa in 6-hydroxydopamine-induced hemiparkinsonian rats: Behavioural and neurochemical alterations. Basic Clin. Pharmacol. Toxicol. 2020, 127, 287–302. [Google Scholar] [CrossRef]
- Mesole, S.B.; Alfred, O.O.; Yusuf, U.A.; Lukubi, L.; Ndhlovu, D. Apoptotic Inducement of Neuronal Cells by Aluminium Chloride and the Neuroprotective Effect of Eugenol in Wistar Rats. Oxid. Med. Cell. Longev. 2020, 2020, 8425643–8425650. [Google Scholar] [CrossRef]
- Mohammadi Nejad, S.; Ozgunes, H.; Basaran, N. Pharmacological and Toxicological Properties of Eugenol. Turk. J. Pharm. Sci. 2017, 14, 201–206. [Google Scholar] [CrossRef]
- Hermanns, H.; Hollmann, M.W.; Stevens, M.F.; Lirk, P.; Brandenburger, T.; Piegeler, T.; Werdehausen, R. Molecular mechanisms of action of systemic lidocaine in acute and chronic pain: A narrative review. Br. J. Anaesth. 2019, 123, 335–349. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, S.-M.; Lee, K.; Im, S.-T.; Go, E.J.; Kim, Y.H.; Park, C.-K. Co-Application of Eugenol and QX-314 Elicits the Prolonged Blockade of Voltage-Gated Sodium Channels in Nociceptive Trigeminal Ganglion Neurons. Biomolecules 2020, 10, 1513. https://doi.org/10.3390/biom10111513
Hwang S-M, Lee K, Im S-T, Go EJ, Kim YH, Park C-K. Co-Application of Eugenol and QX-314 Elicits the Prolonged Blockade of Voltage-Gated Sodium Channels in Nociceptive Trigeminal Ganglion Neurons. Biomolecules. 2020; 10(11):1513. https://doi.org/10.3390/biom10111513
Chicago/Turabian StyleHwang, Sung-Min, Kihwan Lee, Sang-Taek Im, Eun Jin Go, Yong Ho Kim, and Chul-Kyu Park. 2020. "Co-Application of Eugenol and QX-314 Elicits the Prolonged Blockade of Voltage-Gated Sodium Channels in Nociceptive Trigeminal Ganglion Neurons" Biomolecules 10, no. 11: 1513. https://doi.org/10.3390/biom10111513
APA StyleHwang, S.-M., Lee, K., Im, S.-T., Go, E. J., Kim, Y. H., & Park, C.-K. (2020). Co-Application of Eugenol and QX-314 Elicits the Prolonged Blockade of Voltage-Gated Sodium Channels in Nociceptive Trigeminal Ganglion Neurons. Biomolecules, 10(11), 1513. https://doi.org/10.3390/biom10111513