2.2. Conductances in Cycles between Coupled Reactions
In a reaction sequence in which two coupled reactions in series are involved, the output force of the first reaction, A1I, may be used by the second reaction as an input force A1II(A2I is the input affinity, A1II denotes the load affinity). In such a cycle between two coupled reactions, both forces must be equal but of opposite sign. The output power of the first reaction delivers the input power for the second reaction by flowing through A1I, and A2II = - A1I, and back to A1I(at zero power). At steady state, fluxes through A1I and A1II are equal, and hence, both dissipation functions of the cycle, Ф1I and Ф2II, must vanish. From Ф1I = - Ф2II, and Lc1I(A1I)2 = -Lc2II(A2II)2, LIc1 = - LIIc2 is obtained. That is, the partial conductances of two coupled reactions in series are opposite and equal, if under conditions of steady state cycling the magnitudes of the output force of reaction I and the input force of reaction II are equal.
Ф1I = -Ф2II can also be expressed in terms of the steady state flux of cycling and of resistances Rc1I = 1/Lc1I, and Rc2II = 1/Lc2II, yielding Rc1IJ2 + Rc2IIJ2 = 0. It follows that at steady state cycling between two coupled reactions, the sum of the resistances in that cycle must vanish, i.e., that the steady state flux through a cycle between two coupled reactions always occurs at zero overall resistance. Cycling is driven solely by A2I + A1II(A1II negative). Because both reactions are coupled, the conductance (resistance) of the whole cycling process is brought about by both partial (in series) conductances associated with the input and load affinity, respectively.
In oxidative phosphorylation (OP), as described in detail in [
1], a proton cycle is generated over the inner mitochondrial membrane. At steady state, coupled outward proton pumping by redox (NAD
red and FAD
red) reactions of the respiratory chain (
JNA and
JFA) equals the back flow of a given fraction (Q
H) of protons through ATP synthase (
JSY) and ATP/ADP exchange (
JAE) plus H/Pi symport (
JPi). Both partial conductances,
are opposite and equal. For the first reaction (index R)
![Metabolites 02 00667 i039]()
, and for the second
![Metabolites 02 00667 i040]()
, with
vR =
vs+1 (total coupling;
ARo = affinity of
JNA plus
JFA;
vR =
vs+1 = 4protons/extent of reaction).
The remaining fraction of protons (Qr
H) flows back (driven by
![Metabolites 02 00667 i044]()
) through several parallel conductances given by the proton leak flux,
JPL, mitochondrial Na
+/Ca
2+ exchange (with Na
+/H
+ contracted to H
+/Ca
2+ exchange), 2
JHCE, and the malate-aspartate shuttle,
JMS. The partial conductance of this residual proton efflux and the sum of conductances of back flowing fluxes, are also of opposite equality.
Analogously, partial conductances of ATP cycling through the potentials of mitochondrial ATP (ATP
m) production plus ATP transport (contracted to
A1I), and of cytosolic ATP splitting (
A2II) can be formulated. Opposite equality of partial conductances is also fulfilled for this cycle (the above results were obtained by using the simulation SIM
GlOx from reference [
1]).
For a further illustration, an analytically solvable example is given in the Appendix section. Simple electric circuits consisting of one battery connected to an outer conductance, or of two batteries in series, are analysed. These examples show very clearly the behavior of coupled in series reactions.
Further evidence of such an equality of conductances comes from the known fact that for a coupled reaction with an attached load, conductance matching (
LLd =
Lc) is needed to achieve a maximal power output [
1]. At total coupling, the output power is given by:
The maximal Pout is found by differentiation with respect to the variable A1, while A2 remains constant, and by setting the derivative equal to zero:
Inserting A1max into equation (2e) yields Lc1 = - Lc and because - Lc1 = LLd, it follows LLd = Lc.
In addition to cycling at the inner mitochondrial membrane, other types of cycles occur in metabolism. Especially in skeletal muscle cells, the phosphofructokinase (PFK) reaction in conjunction with the fructose-1,6-biphosphatase (FBPase) operating anti-parallel represent a substrate cycle, which may control the pathway of glycolysis (GLY) more sensitively than would be possible by PFK alone. In this cycle, fructose-1,6-biphosphate (FBP), which is produced by ATP-coupled formation from fructose-6-phosphate (F6P), is cycled back via FBPase to F6P. However, usually both fluxes are not equal. Also to demonstrate the opposite equality of partial conductances for this kind of cycle, only equal fluxes can be used for this purpose.
As a further example, the phosphocreatine shuttle will be considered. The creatine kinase (CK) reaction can also be regarded as a coupled reaction. Here, ATP splitting powers phosphocreatine (PCr) formation from creatine (Cr), which may proceed near equilibrium. As described in detail in reference [
1], ATP is shuttled between locations of ATP formation (for instance in the inter-membrane space in mitochondria) and locations of high ATP demand like myofibrils. By analogy to an electric circuit built by two in series batteries with an outer circuit conductance (see Appendix (A4)), the output affinity of PCr formation in the inter-membrane space of mitochondria corresponds to
A1I with associated
LIc1, whereas the affinity of the reverse reaction in myofibrils corresponds to
A1II(with
LIIc2). To ensure diffusional flow of PCr and Cr between both locations, an additional driving force (corresponding to U
e; see (A4)) with associated conductance must be present. Under such conditions partial conductances do not match. Only when the additional conductance corresponding to the diffusional process (
Le) is added to
LIc1 does this sum become opposite and equal to
LIIc2, as is shown in (A4).
Le depends greatly on structural features. So, to achieve a high diffusional conductance, diffusional paths must be as short as possible, which in turn requires a high grade of structural organization [
26,
27,
28].
It seems worth mentioning that coupled systems like pump and leak cycles are often not in a steady state. For instance, steady state cycling through sarco/endoplasmatic reticulum Ca2+ ATPases (SERCA) and Ca2+ release channels of the sarcoplasmatic reticulum (SR) breaks off during activation of contraction. There is an enormous Ca2+ efflux through release channels; meanwhile the pumping rate of SERCAs may be low. Under these conditions, respective conductances may greatly differ; however, when a new steady state cycling is reached, the partial conductance of SERCA must be opposite and equal to the conductance of the Ca2+ release channels. The opposite has to be expected, when release channels close again, and the Ca2+ pumping rate exceeds the release rate.
2.3. From Chemical Potentials to Mechanical Force Generation
In striated muscle cells like ventricular muscle cells (VMs) or skeletal muscle fibers (SMFs), force generation as well as shortening is brought about by the cyclic action of cross bridges. It is a known fact that this process is powered by ATP splitting. The underlying mechanism of the energy transduction process, however, is not completely understood. Here, a thermodynamic description of the cycle is derived using a formalism recently published [
1]. It takes into account the basic energetics of enzyme-catalysed reactions, which states that the overall affinity of the catalysed and non-catalysed processes must be equal. For an enzyme-catalysed reaction like:
(S = substrate, E = enzyme, ES = enzyme-substrate complex, P = product), this means that at steady state the sum of the affinities of substrate binding, transition, and product release must yield the affinity of the non-catalysed reaction, which is given by the reaction affinity of all involved compounds in the bulk solution:
or, after contraction of the first two terms:
yielding,
With K'r = K'B × K'T × K'R(k'B, k'T, and k'R are equilibrium constants of the binding, transition, and release reaction, respectively, whereas k'r denotes that of the non-catalysed reaction).
An analogous reaction sequence is used here to describe the cross-bridge cycle. The following cycle is given in chemical notation, i.e., the charges of involved species are taken into account. The cycle begins with the splitting reaction of the de-energised actomyosin complex (A-M) by MgATP2- in the diffusional space of myofibrils:
This first reaction yields dissociated actomyosin with MgATP2- bound to myosin (the bold point denotes binding to myosin). Two negative charges develop on the dissociated actin, which are neutralised by potassium ions, K+, stemming from free MgATP2−, which is now bound to myosin heads. On the dissociated myosin heads, it neutralises both emerging positive charges. This first actomyosin dissociation and binding of MgATP2− to myosin is followed by ATP splitting on the myosin heads. This transition reaction is described by
It is coupled to the formation of energised myosin (
![Metabolites 02 00667 i070]()
), which is characterised by a tilting of the myosin head from a more bent arms position by an angle of about 60° towards the respective Z disc, so that now the myosin head builds a right angle with the opposing actin filament.
![Metabolites 02 00667 i070]()
contains free energy from reaction R
2 as conformational energy.
The force generating stroke of the myosin head is triggered by the association reaction to form the energised actomyosin complex (cross-bridge):
Because of uncompensated charges, the resulting intermediate in curly brackets lacks firmness. A more stable conformation is obtained by the last reaction of the cycle, which restores the electro-neutrality of the cross-bridges:
The existence of a less stable interaction of myosin with actin has been shown previously [
29,
30,
31].
In a coupled reaction, de-energised actomyosin is restored by dissociating first MgADP- and H2PO4- from cross-bridges and then by releasing the stored conformational energy. During this reaction the cross-bridge tilts back by 60° towards the sarcomere centre, whereby free energy is transferred to the actin filament as mechanical energy.
From the above scheme (R
1 to R
4) two fluxes can be obtained, which are responsible on the one hand for the production of dissociated and energised myosin heads (
JEn), and on the other hand for the formation of cross-bridges and subsequent mechanical force generation by stroking (
JStr). At steady state, a certain fraction of myosin heads of a half-sarcomere exists in a dissociated and energised state
![Metabolites 02 00667 i077]()
, while the residual fraction interacts as cross-bridges with actin. The resulting fluxes are given by:
(For a more complete description and definition of reference constants (Kref) see (A5); complete conductances (LEn and LStr, respectively) are given in (A14) and (A15).)
If the constraint
![Metabolites 02 00667 i084]()
is fulfilled, contraction of the affinities of both fluxes yields the overall affinity as required (
AStrLd +
AATP). Here
AEnLd (stored as conformational energy) denotes the affinity coupled to binding of MgATP
2− to myosin heads (
AEnP), and
AStrLd the affinity which is coupled to the power stroke potential (
AStrP).
AStrLd represents the mechanical work per mole of cross-bridges which has to be overcome during stroking. The quantity
JStr ×
AStrLd is directly related to mechanical power output
PStr =
FLd ×
v(
FLd = load force in Newton (N) of all stroking cross-bridges of a given cross sectional area;
v = velocity of shortening in m/s related to a given fiber length), which as such is conveyed to the surroundings.
In the present model of the cross-bridge cycle, AATP is used at two mechanistically and temporally separated steps. They are given on the one hand by binding of MgATP2− and on the other hand by the release of MgADP− and H2PO4−. Here, most of the free energy of ATP splitting is associated with AEnP, which by the coupling process on myosin heads is transformed first into AEnLd, and then is delivered as AStrP to the power stroke after cross-bridge formation. Therefore, the stroke potential in mechanical units (cross-bridge force × stroke length × NA; NA = Avogadro’s number) must be equal to AStrP (see below).
Because ionic species are involved, the reaction sequence of the cycle should be markedly enforced by electrostatic interactions. So MgATP2− binding can proceed only if actomyosin dissociates, whereas release of products becomes possible only when at the same time cross-bridge formation occurs. Moreover, the conformational change in the myosin head forces it into a new position, which favours an interaction with actin at a new actin binding site displaced a certain distance towards the Z-disc. During stroking, binding of a new MgATP2− molecule and detaching of cross-bridges may preferentially occur at the end of the power stroke, when cross-bridges form an angle of about 60° with the actin filament (see below for uncoupling by stroke shortening).
The contractile performance of whole muscle and of SMFs is exceptionally well reproduced by Hill’s equation [
19]. This equation relates the shortening velocity
v to the mechanical load force
FLd which has to be overcome during shortening.
The above function represents a hyperbola, which fits remarkably well with experimental data obtained under isotonic conditions.
To obtain an equivalent expression from the flux equation JStr, the flux given in mM/s has to be converted into velocity with units of m/s. This is achieved by calculating the stroke frequency for a given concentration of stroking cross-bridges ([CB] = [CB]tot – [MHEn], in mM) and by multiplying with the stroke length lStr(in m) and the number of in series half-sarcomeres Nhs. The result is:
The above expression describes the shortening velocity as a function of
AStrLd at constant
AStrP. It represents a straight line (
Figure 1A). Introducing a Michaelis-Menten like inhibition factor associated with
LStr yields the desired hyperbolic dependency:
Comparing equations 9a and 9d shows that the constant b of Hill’s equation is given by:
As required, the quotient by which b is multiplied is dimensionless. To yield the shortening velocity as a function of force, v(FLd), affinities and KmLd(both in J/mol) have to be converted into units of force. This is achieved by dividing by l Str and by multiplying by the molar number of cross-bridges.
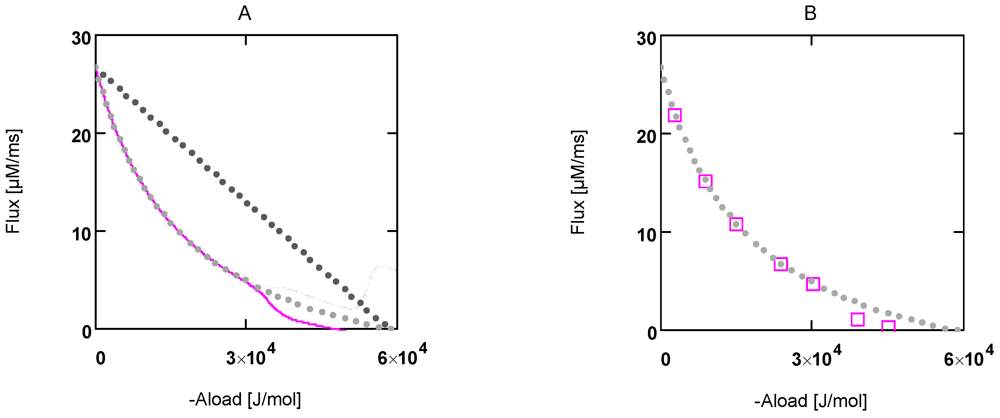
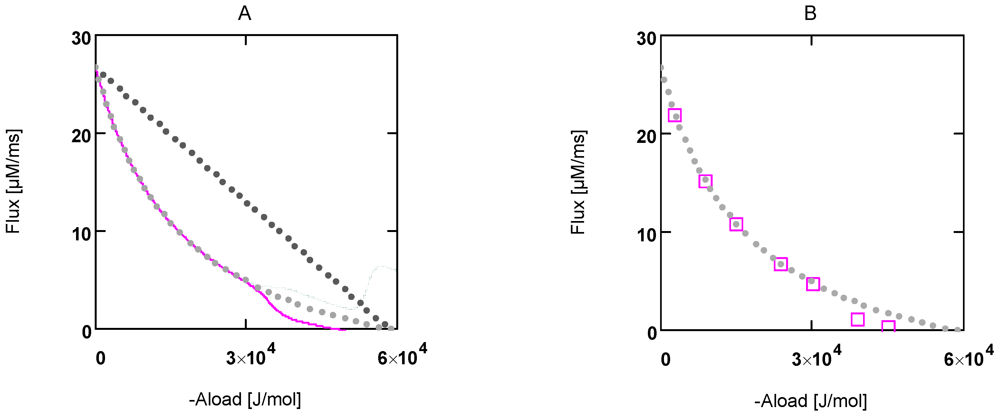
Figure 1.
Flux as a function of load potential at 10.8 µM [Ca2+]. A: (grey dots) according to equation 9b; (light grey dots) according to equation 9c or 9d; (red line) according to equation 11a; (green line) according to equation 11b. B: (light grey dots) according to equation 9c or 9d; (red squares) results from simulation SIMGLYgen.
AStrLd being negative, FLd must also be ≤0. Expressing shortening velocity as a function of a positive variable yields with FLd = - FLd+
Setting -
KfmLd =
a, and -
b =
b+, gives (
Figure 2.)
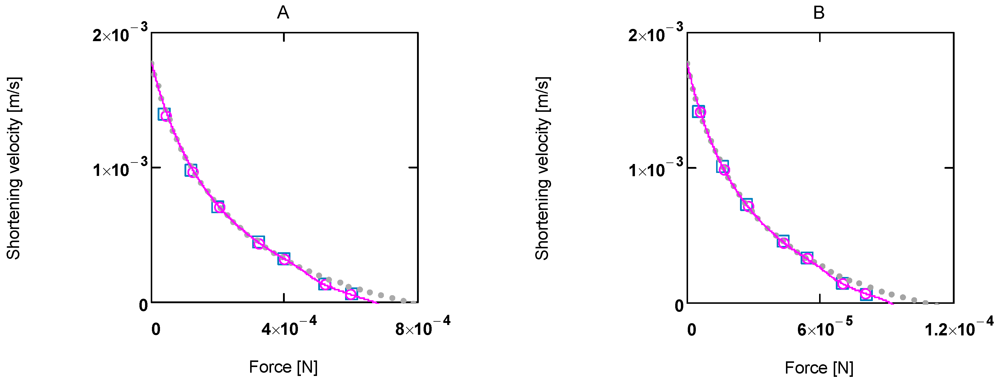
Figure 2.
Shortening velocity as a function of load force at two different Ca2+ concentrations A:[Ca2+] = 1.08 µM; (light grey dots) according to equation 10b; (red line) equation 10b plus uncoupling; (red circles) results from SIMGLYgen versus load force; (blue squares) results from SIMGLYgen versus load force as sensed by cross-bridges; B: as in A, but at [Ca2+] = 0.36µM.
The latter equation formally represents Hill’s equation. In that equation F0 denotes the maximal force obtained under isometric conditions, whereas FP in the latter equation is obtained from the input affinity (AStrP) of JStr by converting it into units of force (see below for a derivation of Fp ≡ F0).
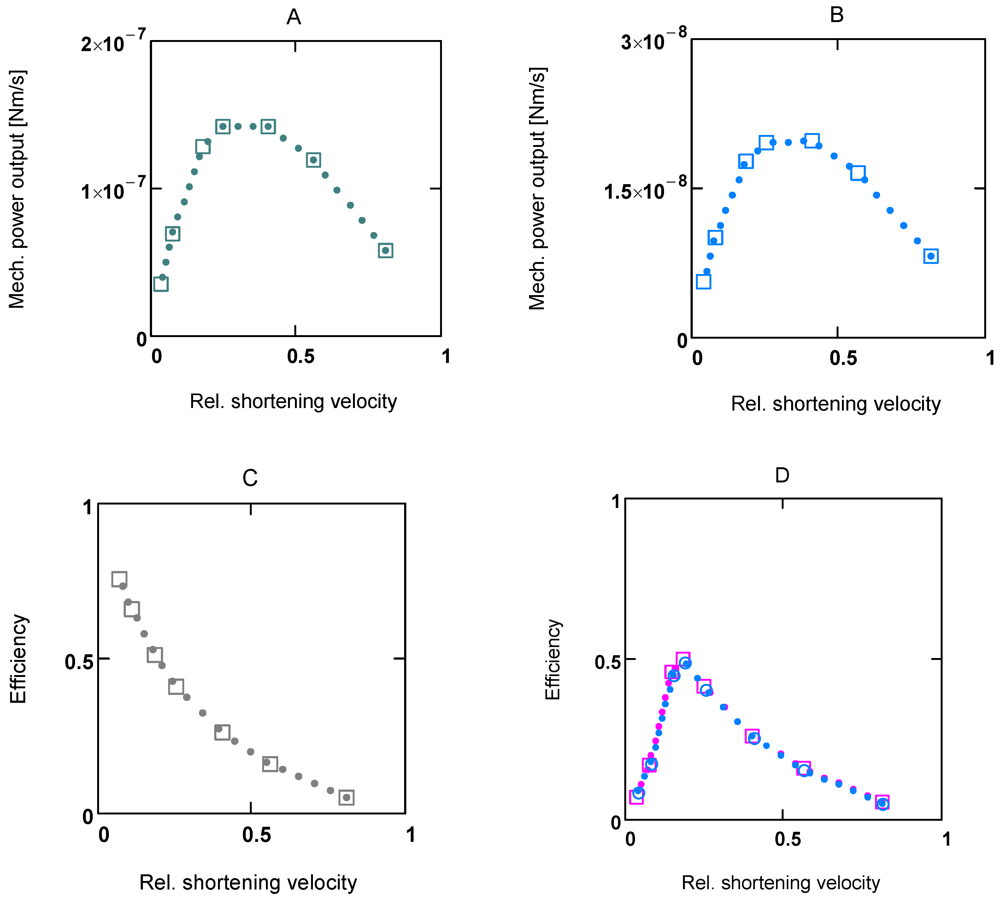
When efficiency is represented as a function of shortening velocity, the experimental data follow a curved line with a maximal efficiency at 0.18 ×
vmax [
12]. From NET it is known that such a maximum is produced by uncoupling. To create such a maximal efficiency, uncoupling terms have to be incorporated into
JStr. Variable, load dependent
λ values (
λ(
AStrLd) instead of constant
λ's) are defined, to preserve the hyperbolic nature of the function. In this way, uncoupling becomes operative only when
AStrLd exceeds a certain value (
Figure 1A). Both flux equations are given by:
These latter equations (for a complete description, especially of conductances, see (A15)) appear in simulations. λ values are given as functions of AStrLd, e.g.,
(see below for a mechanistic interpretation of uncoupling and λ values). Conversion to mechanical units can then be done in the same way as shown above.
In
Figure 1 the effects of uncoupling are shown. At a load of about −3.0×10
4 J/mol, deviations from the hyperbolic (coupled) curve begin to arise. From the plots it can be seen that uncoupling leads to a shift of the intersection with the abscissa to less negative values of
AStrLd, whereas - (
JStrP) is still maintained, even at
AStrLd = - (
AStrP), where the coupled flux must be zero and only uncoupled fluxes are possible.
In the following, an attempt has been made to interpret the above results, which were gained from a phenomenological approach, mechanistically by relating coupled and uncoupled fluxes to possible cross-bridge actions.
At AStrLd = - (AStrP), coupled reactions with associated actin filament movement come to a halt, because the driving force has vanished. As already mentioned above, now only uncoupled fluxes can occur. Such a situation may also be realised with isometric contraction, which is known to be associated with ATP splitting and heat production, but without power output. That is, a mechanism has to be found which explains the identity of the isometric force F0 with FP, which was merely formally derived from the input affinity AStrP by a conversion factor. This is achieved by defining the uncoupling mechanism by a shortening of the stroke length lStr of the power stroke. Total uncoupling is reached when lStr = 0. This may be realised under isometric conditions. Free energy corresponding to AStrP ≈ AATP is delivered to actin filaments as mechanical work, i. e., F0 × lStr × NA = AStrP. Shortening may be brought about through splitting of actomyosin bonds before the whole stroke length is transferred to an actin filament. When AStrLd = - AStrP, as is realised under isometric conditions, actomyosin splitting already occurs at zero stroke length, so that no energy can be delivered to the actin filaments. Only force development by cross-bridges during the time interval between bond formation and bond splitting is possible under these conditions. This may be achieved by the torque every myosin head exerts on an actin filament after bond formation and release of H2PO4− and MgADP−. The associated force then acts on these filaments, but without being able to bring about filament movement, since this is hindered by the equal and opposite load force. Therefore, if the force remains constant over the whole stroke length, then FP is equal to the isometric force F0.
When compared with a cross-bridge cycle during contractions, the cross-bridge cycle under isometric conditions becomes altered insofar as coupled stroking is impossible; the power stroke occurs completely uncoupled, so that all free energy associated with AStrP becomes dissipated as heat. Moreover, dissipative stroking under these conditions may occur in the presence of bound MgATP2−. The following derivation shows how stroke shortening may be involved with uncoupling.
Stroke shortening is given by:
this leads to,
Under totally coupled conditions, the input flux is given by:
Uncoupling by stroke shortening dissipates free energy, which can be expressed by a leak dissipation function:
The leak conductance LPStrL can be replaced by LStr, because this latter conductance may depend mainly on the formation mechanism of the actomyosin bond. The stroke reaction associated with conformational changes of the myosin head is assumed to proceed at a high conductance, since the energising reaction (JEn), which is coupled to the same conformational change in the reverse direction, also proceeds at a very high conductance. So an increase by stroke shortening of a high conductance (stroking) in series with a low conductance (bond formation) may be negligible, so that ФPStrL can be expressed as:
Comparing this latter equation with that used in the simulation,
![Metabolites 02 00667 i131]()
, yields:
The input flux then is given by:
The output flux is reduced by stroke shortening as if it were uncoupled. The same dissipation function LStr(ΔAStrLP)2 is associated with output reactions, yielding:
and the output flux:
It follows,
For Δl = 0, identical coupled fluxes arise, and for AStrLd = - AStrP, both λ values are equal. Moreover, if λStrLd = λStrP = 1, (Δl/lStr)2 is also equal to 1.0, which means that now isometric conditions do exist.
From equation (13i) it can be taken that uncoupling by stroke shortening reduces JStrLd as if there were a leak flux through AStrLd. On the other hand, JStrP increases (equation (13g)) as if there were an additional leak flux through AStrP. The above derivations demonstrate that stroke shortening obviously leads to the same effects as uncoupling by leak fluxes. It seems justified, therefore, to also describe uncoupling by stroke shortening by lambda values, as was done previously mainly in the context of oxidative phosphorylation.
The degree of coupling is given by,
with above results this yields:
Under the limiting conditions of isometric contraction (AStrLd = - AStrP; (Δl/lStr)2 = 1.0), qStr is given by:
At loads ≈ -3 × 10
4 <
AStrLd < −
AStrP,
λStrP(
AStrLd) will be smaller than 1.0 but not zero, so that the process in this region would proceed at a higher degree of coupling (e. g. at
AStrLd = −4.5 10
4,
qStr = 0.833). At values of
AStrLd > ≈ -3 × 10
4 the process is totally coupled ((
Δl/lStr)
2 = 0), that is, cross-bridges work at full stroke length. Only this part of the performance curve (
Figure 1 and
Figure 2) is hyperbolic and fulfils Hill’s formalism. Between the intersection (
AStrLd = −4.756×10
4) and
AStrLd = -
AStrP,
JStrLd formally could be negative, which would mean that actin filaments were moving in the direction of stretching. This is, however, impossible, because actomyosin bonds would have to be broken by a load force, which is smaller than
F0. Therefore, in this region of loads,
JStrLd cannot be negative; it must remain zero.
2.5. Calcium Ions and Force Development
In the previous section it was shown, how shortening velocity depends on
AStrLd at a given [Ca
2+]. On the one hand, the driving force is changed by the load potential (see
Figure 1, linear dependence), and on the other hand the conductance
LStr depends on
AStrLd through the hyperbolic inhibition factor. At a given [Ca
2+]
, both effects are responsible for the characteristic appearance of the performance curve under coupled conditions.
In the present model of the cross-bridge cycle, interference of [Ca
2+] with
AEnP as well as with
LStr is necessary. In the latter case, [Ca
2+] can activate
JStr through a sigmoid activation factor (A15). This takes into account the fact that Ca
2+ binding to troponin C removes the inhibition of cross-bridge cycling, so that binding of myosin heads to actin binding sites becomes possible [
32,
33]. On the other hand, [Ca
2+] is known to strongly activate force development. Here it is assumed that this may be caused by an increase in cross-bridge concentration [CB]. By introducing a [Ca
2+] dependent
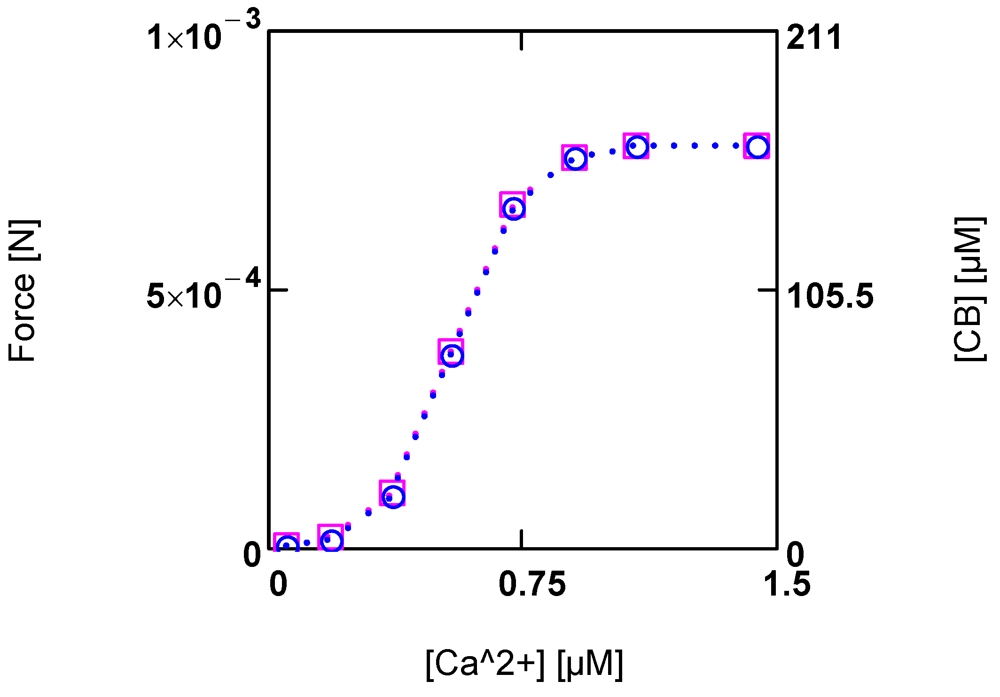
KBref (see (A14)), a sigmoid variation in both [CB] and force
F by [Ca
2+] can be obtained (
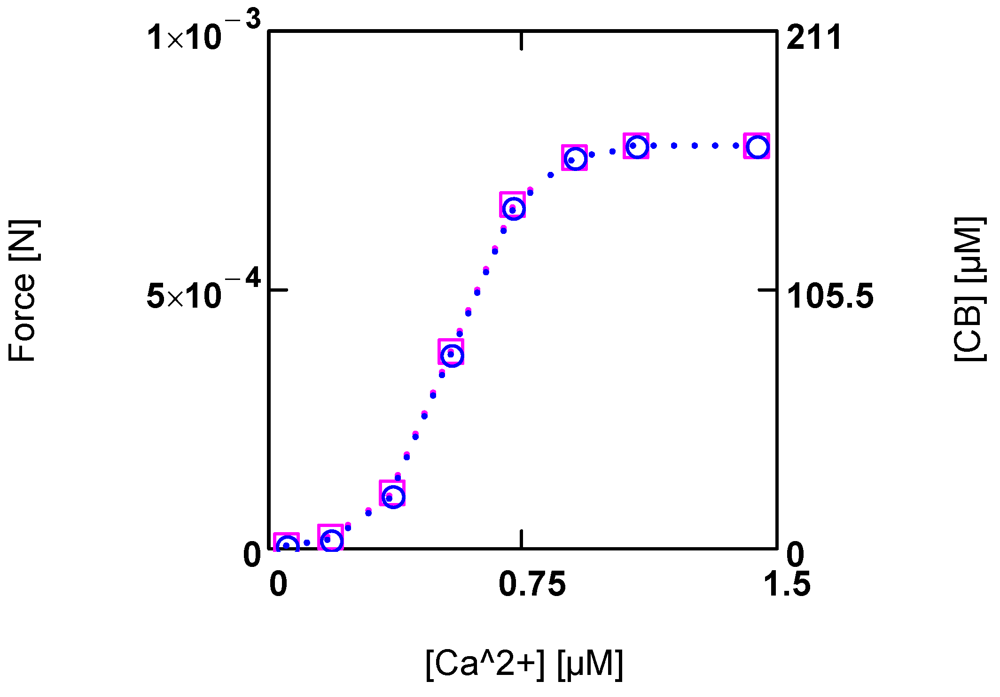
Figure 4.).
Figure 4.
Developed force and cross-bridge concentration [CB] and their dependence on [Ca2+]. (red squares) force; (blue circles) [CB]. Notice that at the given dimensioning of the right ordinate a matching of results is obtained.
At steady state, a certain [CB] is produced by [Ca2+] activated JStr, and in addition by [Ca2+] inhibited JEn (see (A14)). The inhibition of JEn by [Ca2+] is brought about by a decrease of AEnP with increasing [Ca2+]. This is possible because this reaction proceeds at a very high conductance and therefore, is close to equilibrium. So already a small variation of the driving force can produce a large change in the reaction velocity. In this way, a sensible, [Ca2+] dependent adjustment of [CB] and force can be achieved. An elevation of [Ca2+] thus increases both shortening velocity as well as force development.
The total myosin head concentration ([MHEn] + [CB]) of a half-sarcomere amounts to 656 µM (see Methods). At a saturating [Ca2+] of 1.08 µM, fluxes JEn and JStr are so adjusted as to yield a concentration of [CB] = 0.25 ([MHEn] + [CB]), i.e., at this [Ca2+], 25% of myosin heads form cross-bridges and thus are involved with cycling and force generation. At [Ca2+] = 0.36 µM, only about 3% of cross-bridges are engaged, and at 0.09µM [Ca2+], [CB] is further markedly reduced, which means that now near resting conditions are reached.
It seems plausible to suggest that during shortening it is not always the same group of cross-bridges that is active, but that, e.g., at 1.08 µM [Ca
2+], four different groups may alternately be involved with contraction. The cycling frequency of an individual cross-bridge would then be much lower than the frequency of ATP splitting, which might be advantageous, especially at high velocities. Furthermore, an alternating involvement of groups may be absolutely necessary for a smooth shortening. How this might be accomplished is so far not known. An involvement of special filaments of the sarcomere cytoskeleton [
34,
35,
36], which may be responsible for a subtle sensing of load forces and an undisturbed takeover of a given load by a new fraction of cross-bridges during synchronous stroking, seems indispensable.
The values of maximal tension (=force/unit area in N/m
2 = Pa, Pa = Pasqual) obtained from SIM
GLYgen (A16) in the present study are comparable to experimental values. For instance, a value of 372 kPa (from
F0 = 7.756 × 10
−4 N, [Ca
2+] = 1.06 µM, 37°C) found here, seems to be in reasonable agreement with about 320 kPa resulting from measurements with a fast-twitch mouse fiber at 25°C [
37]. The extrapolated value of maximal shortening velocity,
vmaxHS = 1.95 µm × HS
−1 × s
−1 ([Ca
2+]= 1.06 µM,
AL = 0 J/mol, HS = half-sarcomere) compares to 1.6 µm × HS
−1 × s
−1 of frog fibers at 0°C [
12]. A value of
ηmax of about 50% at about 0.18
vmax ([Ca
2+] = 1.06µM) results from adjustment. It compares to the experimental values of 35–45 % for the same value of
v for frog muscles at 0 °C [
12].
All these parameters of contractile performance may, however, be reduced to a certain extent by dissipative frictional processes associated with v which are not addressed in the present simulation. Such dissipation during fiber shortening may be produced mainly by viscous deformations of membranes and the filament lattice.
2.6. [H+], [Mg2+], and Fatigue
Enzyme-catalysed ATP splitting by myosin heads is formulated here with respect to the ATP species MgATP2−. By using a reference constant and binding polynomials, an [H+] and [Mg2+] dependent K'ATP of this reaction can be formulated (see A6 and A7). In simulations of fatigue, in addition to [H+], [Mg2+] has also been included as a variable, especially because this ion may interfere with ATP species and so may influence JEn through a change in [MgATP2−], which in turn would alter [CB].
Changes in [H+] in the sarcosol are brought about mainly by two different mechanisms, which are both related to metabolic activity. One source of protons is manifest when metabolism is switched from rest to high power output. Fluxes in ATP consumption and production, JATPCon and JATPPro, respectively, must then both increase to the same extent to reach a new steady state. During the adjustment, a phase of disturbed steady state occurs, during which both fluxes do not match. When power output increases, JATPCon always leads JATPPro, i.e., there is an uncompensated ATP splitting until a new steady state is reached, at which point ATP production again equals ATP consumption.
According to Alberty [
20], this reaction is associated with proton production in dependence of [H
+] and [Mg
2+] (see (A6) and (A7)) for derivation of [H
+] changes and pH buffering). In addition, the CK and adenylate kinase (AK) reactions are involved, because these equilibria are also changed under these conditions and, as with ATP splitting, H
+ and Mg
2+ binding species are involved. Buffering of both ion concentrations is brought about mainly by sites intrinsic to the sarcosol. For Mg
2+ binding sites, an additional release of Mg
2+ by interfering [H
+] has to be expected.
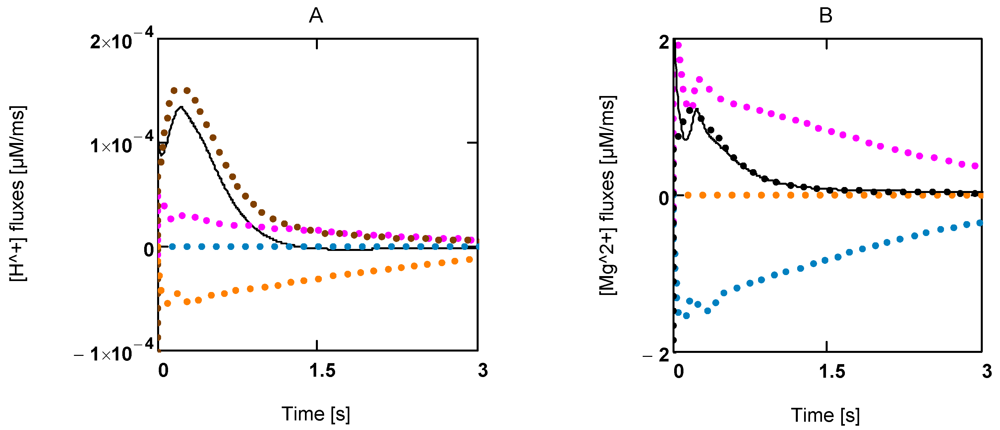
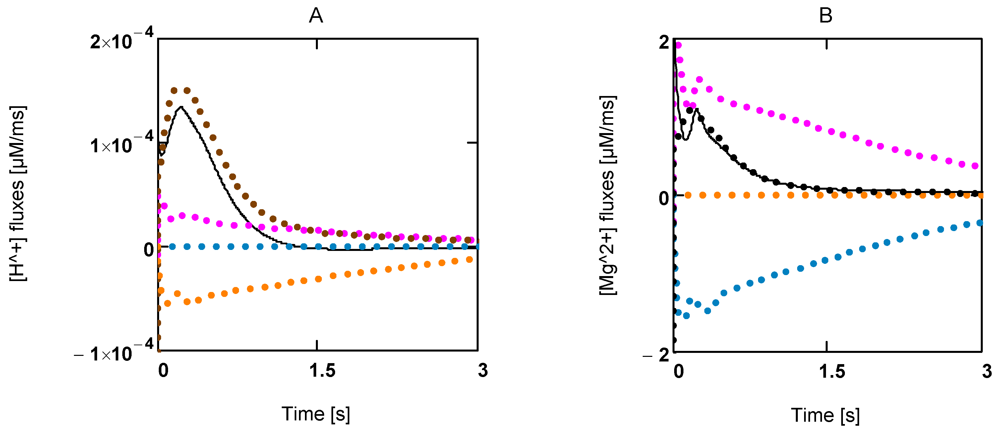
Figure 5A shows the time courses of rates of [H
+] changes. Interestingly, [H
+] production by ATP splitting is practically compensated by [H
+] consumption by the CK reaction. The contribution by the AK reaction is negligible. A similar behavior is found for Mg
2+ (
Figure 5B). A concentration increase in this ion is mainly brought about by acidification.
Figure 5.
Time courses of [H+] and [Mg2+] during extreme power output. A: [H+] fluxes; (brown) mainly LDH reaction and lactate transport; (red) ATP splitting; (blue) JAK; (yellow) JCK; (black line) resultant [H+] flux; B: [Mg2+] fluxes of the same reactions.
A second source of protons is given by the disturbance of lactate production by glycogenolysis (or glycolysis) and lactate efflux via lactate/H symport at the sarcolemma. Especially when lactate and H+ accumulate in the glycocalyx (the outer aspect of the sarcolemma), the concentrations of these compounds also increase drastically in the sarcosol. This seems to be the main mechanism of sarcosolic acidification.
Muscular fatigue at the cellular level can be defined as a phase of markedly reduced contractile performance, which largely recovers after a period of rest [
38]. Because metabolites like creatine, ADP, Pi, H
+, and lactate accumulate during conditions of fatigue in a similar way as can be observed during ischemia or hypoxia, which are known to be the result of impaired ATP production, it seems justified to suggest that the preconditioning for fatigue may also be initiated by a deterioration of the energy metabolism of the muscle fibers. Whenever ATP delivery does not match ATP consumption, such a situation may arise.
These effects can be easily demonstrated with a simulation of glycogenolytic or glycolytic ATP production in the absence of mitochondrial metabolism (SIMGLYgen, see (A16)), which is related to the energy metabolism of fast muscle fibers. At 1.08 µM [Ca2+] and a load of –1.5 × 104 J (constant glycogen content and glucose concentration [Glu] = 4.0 mM), efficiency of glycogenolytic ATP production is ηGLYgen = 0.722, that of glycolytic ATP ηGLY = 0.525. The higher efficiency is mainly caused by the stoichiometric coefficients of coupled ATP production of 3.0 and 2.0 for the glycogenolytic and glycolytic pathways, respectively.
Under these conditions of high power output, metabolite concentrations change only moderately compared to resting conditions (at 1.06 µM [Ca2+] and a load potential of −1.5 × 104 J/mol, [ADP] = 113, [Pi] = 8.32 × 103, phosphocreatine concentration [PCr] = 9.7 × 103, lactate concentration [Lac] = 3.0 × 103, [Mg2+] = 832, and pH = 7.09).
However, when a back pressure on glycogenolysis (or glycolysis) is produced by accumulated extracellular [Lac]e and [H+]e, the flux through this pathway may become reduced. In addition, efficiency has been reduced by switching from glycogenolysis to glycolysis. The power output of ATP production is markedly reduced by these combined effects. As a result, the power of ATP production begins to fall, so that ATP consumption may overcome ATP production. Steady state cycling through ATP consuming and producing pathways can now no longer be maintained.
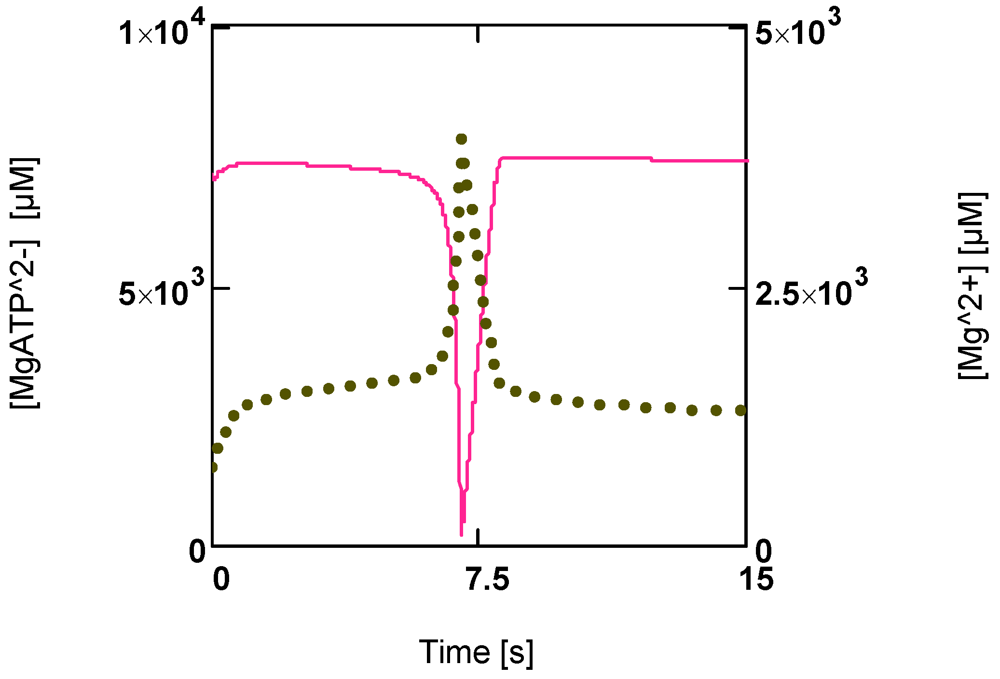
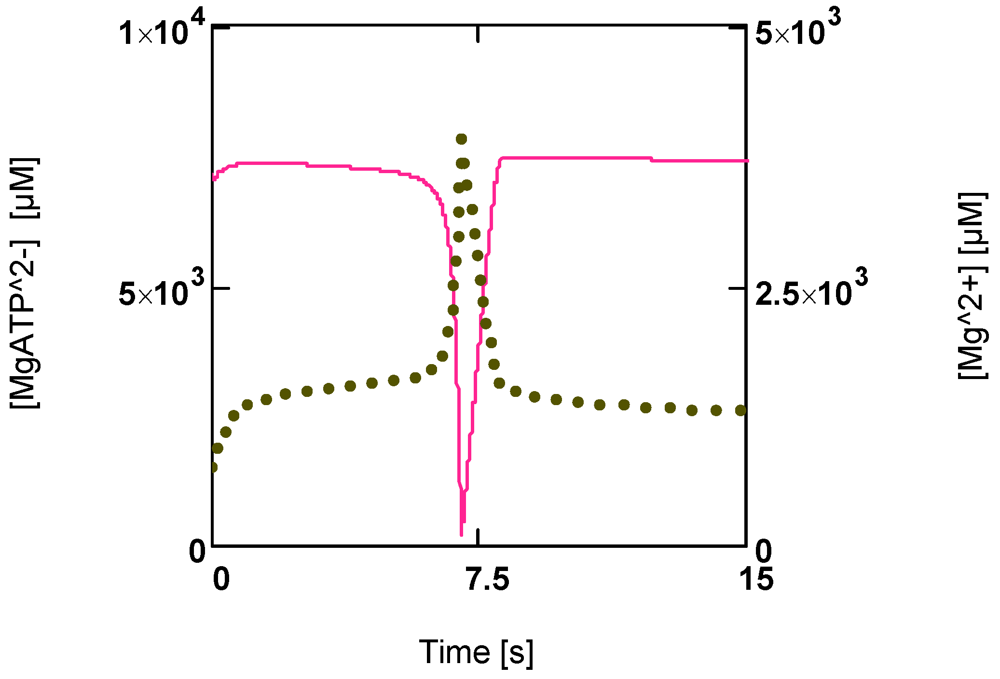
Figure 6 shows that a first phase of slowly falling [MgATP
2−] is followed by a phase of continuously enhanced reduction of this ATP species to low values ([MgATP
2−] = 230.0 µM; [PCr] = 1.6 µM). Immediately after reaching a minimum, a rapid recovery of [ATP] (up to starting values) begins. [Mg
2+] shows a corresponding behavior. During the first phase it increases because of acidification, and then a sharp peak is produced by the onset of an extreme uncompensated ATP splitting (
Figure 6). An increased [Mg
2+] may counteract the switch off of cross-bridge cycling and may aid recovery by increasing [MgATP
2−].
Figure 6.
Time courses of [MgATP2−] and [Mg2+] during development of fatigue. (red line) [MgATP2−]; (green points) [Mg2+].
Other parameters such as [PCr], [Pi], [Lac], and pH only partially recover under these conditions of extreme power output. An almost complete recovery, however, is possible under conditions of markedly reduced power output near resting [Ca2+].
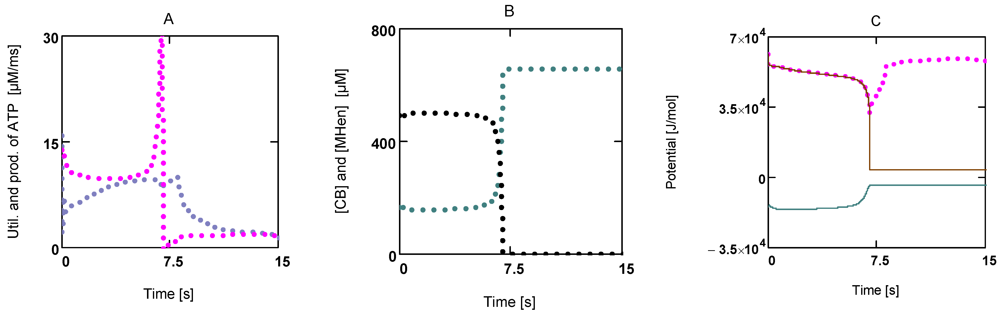
How this switch back to normal [ATP] is brought about can be seen from
Figure 7. Not only have fluxes of ATP consumption and production,
JATPCon and
JATPPro, become different now (
JATPcon >
JATPpro;
Figure 7A), both fluxes of the cross-bridge cycle,
JEn and
JStr, have also changed. These fluxes determine concentrations of [MH
En] and [CB], respectively. An increase in
JEn and a decrease in
JStr would lower [MH
En] (whereby [CB] would be increased). Both concentrations always change reciprocally (
Figure 7B).
AStrP and
AStrLd are also affected.
AStrP in particular is rapidly reduced until it is equal to −
AStrLd. Now all fluxes of the cycle must vanish, because the driving force of
JStr has become zero. As a result, ATP consumption by cross-bridge cycling is switched off.
Figure 7.
Time courses of JATPCon and JATPPro, of [CB] and [MHEn], and potentials of the cross-bridge cycle during fatigue development. (A) (red) JATPCon; (blue) JATPPro; (B) (black) [CB]; (blue) [MHEn]; (C) (red points) AATP; (brown line) AStrP; (blue line) AStrLd; notice that after 7.5 s AStrP and AStrLd become opposite and equal.
Under these conditions all myosin heads form cross-bridges, which however are unable to perform the power stroke, since the input force is equal to the opposed load force. In such a situation myosin heads may be bound to actin and may have dissociated H2PO4− and MgADP− similar to an isometric contraction, but in contrast to those latter conditions, equilibrium of forces is now brought about at a much lower load force (AStrP = −AStrLd = 0.375 × 104 J/mol at 1.08 µM [Ca2+]). A load-dependent actomyosin splitting by MgATP2− at the beginning of the stroke, that is uncoupling, is impossible under these conditions. So cross-bridge cycling with concomitant ATP consumption may be completely prevented. [ATP], therefore, can recover rapidly, even if the conditions leading to fatigue first remain unchanged.
By this mechanism the fatigued skeletal muscle fiber is capable of protecting itself from the dangerous risk of irreversible cell damage. This seems to be necessary, since this cell type is voluntarily controlled without any protecting mechanism against an unbridled consumption of ATP, as is known to occur with other ATP coupled reactions such as, for instance, ion pumps. These are controlled mainly by the ion concentration that they are transporting. For example, when [Na+] in the sarcosol is lowered by the Na/K pump to values below 10.0 mM, the reaction rate of this transport process is increasingly deactivated by the decreasing [Na+], so that ATP consumption also is reduced.
Such a protective mechanism is not known, however, for the cross-bridge cycle. Contractions with concomitant ATP splitting would be incessantly initiated, as long as firing of nervous impulses persisted. The voluntary muscle fiber would obey this parent command up to exhaustion or even up to cell death, if the fatigue producing mechanism were absent. Obviously it represents that special control mechanism which is necessary to protect voluntary muscle from dangerous ATP depletion during phases of high energetic demands.
The above results are obtained from a simulation, in which ATP production is confined solely to glycolysis. A whole muscle, however, is constructed from many types of functionally different fibers, with slow fibers having densely packed mitochondria, and fast fibers in which mitochondrial density can be very low. It is a known fact that especially fast fibers with a very low content of mitochondria and, therefore, a mainly anaerobic ATP production, are much more liable to be affected by overpowering than slow fibers. This may be brought about primarily by the preconditioning effects especially associated with this fiber type. At very high energetic demands, fast fibers produce much more lactate and protons through glycolysis than slow fibers, which can oxidise pyruvate by mitochondria. That is, the glycolytic ATP production rate of fast fibers may be decelerated by a back pressure, which may be generated by accumulating lactate and protons during high power output. In slow fibers with a high rate of oxidative glucose metabolism, such a back pressure cannot be produced as easily. Therefore, the metabolic changes leading to fatigue are simulated here with respect to fast fibers with a low resistance to fatigue. This weakness may be best demonstrated with an extreme fiber type, which can produce ATP solely by GLY. However, fibers completely devoid of mitochondria may not exist in vertebrate muscle. The results of this fatigue model, therefore, can only be taken as an approximation of real fast muscle fibers. Muscular contraction on the basis of NET has been treated theoretically by Caplan and Essig [
13]. These authors explained the curvature of Hill’s equation by an uncoupling. They defined the degree of coupling by
![Metabolites 02 00667 i170]()
with
θ =
a/
F0 =
b/
vmax(
θ between 0.2 and 0.3). This latter equality is also fulfilled by the present model (
θ = 0.309). However, in contrast to the approach of the above authors, the hyperbolic form of Hill’s equation is produced here by introducing a Michaelis-Menten-like inhibition factor into the respective conductance, as already mentioned above. Moreover, uncoupling in the present model is produced through a load-dependent stroke shortening, which generates the maximum obtained with power plots.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 , and
, and 


 , and
, and 

 , and
, and 
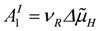 , and for the second
, and for the second 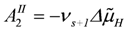 , with vR = vs+1 (total coupling; ARo = affinity of JNA plus JFA; vR = vs+1 = 4protons/extent of reaction).
, with vR = vs+1 (total coupling; ARo = affinity of JNA plus JFA; vR = vs+1 = 4protons/extent of reaction).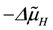 ) through several parallel conductances given by the proton leak flux, JPL, mitochondrial Na+/Ca2+ exchange (with Na+/H+ contracted to H+/Ca2+ exchange), 2JHCE, and the malate-aspartate shuttle, JMS. The partial conductance of this residual proton efflux and the sum of conductances of back flowing fluxes, are also of opposite equality.
) through several parallel conductances given by the proton leak flux, JPL, mitochondrial Na+/Ca2+ exchange (with Na+/H+ contracted to H+/Ca2+ exchange), 2JHCE, and the malate-aspartate shuttle, JMS. The partial conductance of this residual proton efflux and the sum of conductances of back flowing fluxes, are also of opposite equality.

 leads to
leads to 






 ), which is characterised by a tilting of the myosin head from a more bent arms position by an angle of about 60° towards the respective Z disc, so that now the myosin head builds a right angle with the opposing actin filament.
), which is characterised by a tilting of the myosin head from a more bent arms position by an angle of about 60° towards the respective Z disc, so that now the myosin head builds a right angle with the opposing actin filament. 

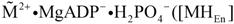 , while the residual fraction interacts as cross-bridges with actin. The resulting fluxes are given by:
, while the residual fraction interacts as cross-bridges with actin. The resulting fluxes are given by: , and
, and

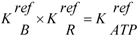 is fulfilled, contraction of the affinities of both fluxes yields the overall affinity as required (AStrLd + AATP). Here AEnLd (stored as conformational energy) denotes the affinity coupled to binding of MgATP2− to myosin heads (AEnP), and AStrLd the affinity which is coupled to the power stroke potential (AStrP). AStrLd represents the mechanical work per mole of cross-bridges which has to be overcome during stroking. The quantity JStr × AStrLd is directly related to mechanical power output PStr = FLd × v(FLd = load force in Newton (N) of all stroking cross-bridges of a given cross sectional area; v = velocity of shortening in m/s related to a given fiber length), which as such is conveyed to the surroundings.
is fulfilled, contraction of the affinities of both fluxes yields the overall affinity as required (AStrLd + AATP). Here AEnLd (stored as conformational energy) denotes the affinity coupled to binding of MgATP2− to myosin heads (AEnP), and AStrLd the affinity which is coupled to the power stroke potential (AStrP). AStrLd represents the mechanical work per mole of cross-bridges which has to be overcome during stroking. The quantity JStr × AStrLd is directly related to mechanical power output PStr = FLd × v(FLd = load force in Newton (N) of all stroking cross-bridges of a given cross sectional area; v = velocity of shortening in m/s related to a given fiber length), which as such is conveyed to the surroundings. 

 , or
, or

 (in m/s)
(in m/s)






 (negative)
(negative)
 (at constant force)
(at constant force)



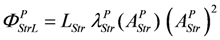 , yields:
, yields:







 , and
, and  .
.





 with θ = a/F0 = b/vmax(θ between 0.2 and 0.3). This latter equality is also fulfilled by the present model (θ = 0.309). However, in contrast to the approach of the above authors, the hyperbolic form of Hill’s equation is produced here by introducing a Michaelis-Menten-like inhibition factor into the respective conductance, as already mentioned above. Moreover, uncoupling in the present model is produced through a load-dependent stroke shortening, which generates the maximum obtained with power plots.
with θ = a/F0 = b/vmax(θ between 0.2 and 0.3). This latter equality is also fulfilled by the present model (θ = 0.309). However, in contrast to the approach of the above authors, the hyperbolic form of Hill’s equation is produced here by introducing a Michaelis-Menten-like inhibition factor into the respective conductance, as already mentioned above. Moreover, uncoupling in the present model is produced through a load-dependent stroke shortening, which generates the maximum obtained with power plots.  , and
, and
 (=1.415027×10−15 m3 or 1415.027 pL)
(=1.415027×10−15 m3 or 1415.027 pL)




 or 0.972453 pL
or 0.972453 pL




 , and
, and 
 , and from (A1),
, and from (A1),  .
.
 , or
, or 

 , and
, and 






 , and
, and 
 , and
, and 

 is identical to κHBU if binding sites contain only a single site with only one proton dissociation constant.
is identical to κHBU if binding sites contain only a single site with only one proton dissociation constant.


 , is adjusted to 9.0 mM plus a variable [ATP]. The change of [Mg2+] is given then by:
, is adjusted to 9.0 mM plus a variable [ATP]. The change of [Mg2+] is given then by:







 .
. 

 .
. 
 .
.  .
.  .
.  .
.  .
.  .
.  .
.  .
.  .
.  .
.  .
. 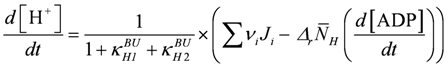 .
. 