

A Comparative Study Using Reversed-Phase and Hydrophilic Interaction Liquid Chromatography to Investigate the In Vitro and In Vivo Metabolism of Five Selenium-Containing Cathinone Derivatives

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Enzymes
2.2. Synthesis Procedures
2.3. Nuclear Magnetic Resonance Spectroscopy
2.4. pHLS9 Incubations
2.5. Rat Urine Collection and Sample Preparation
2.6. Monooxygenase Activity Screening
2.7. LC-HRMS/MS and Data Evaluation Conditions
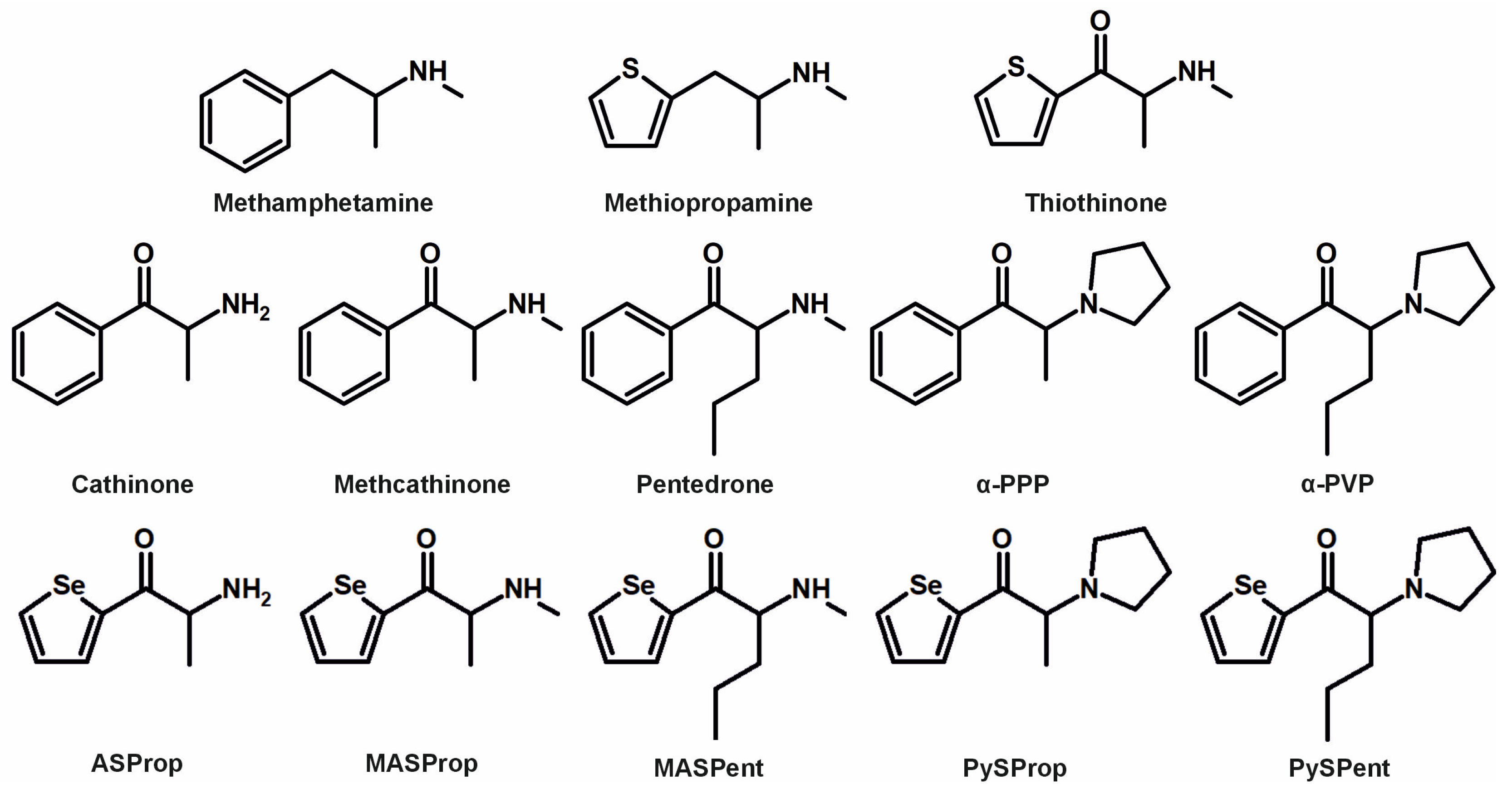
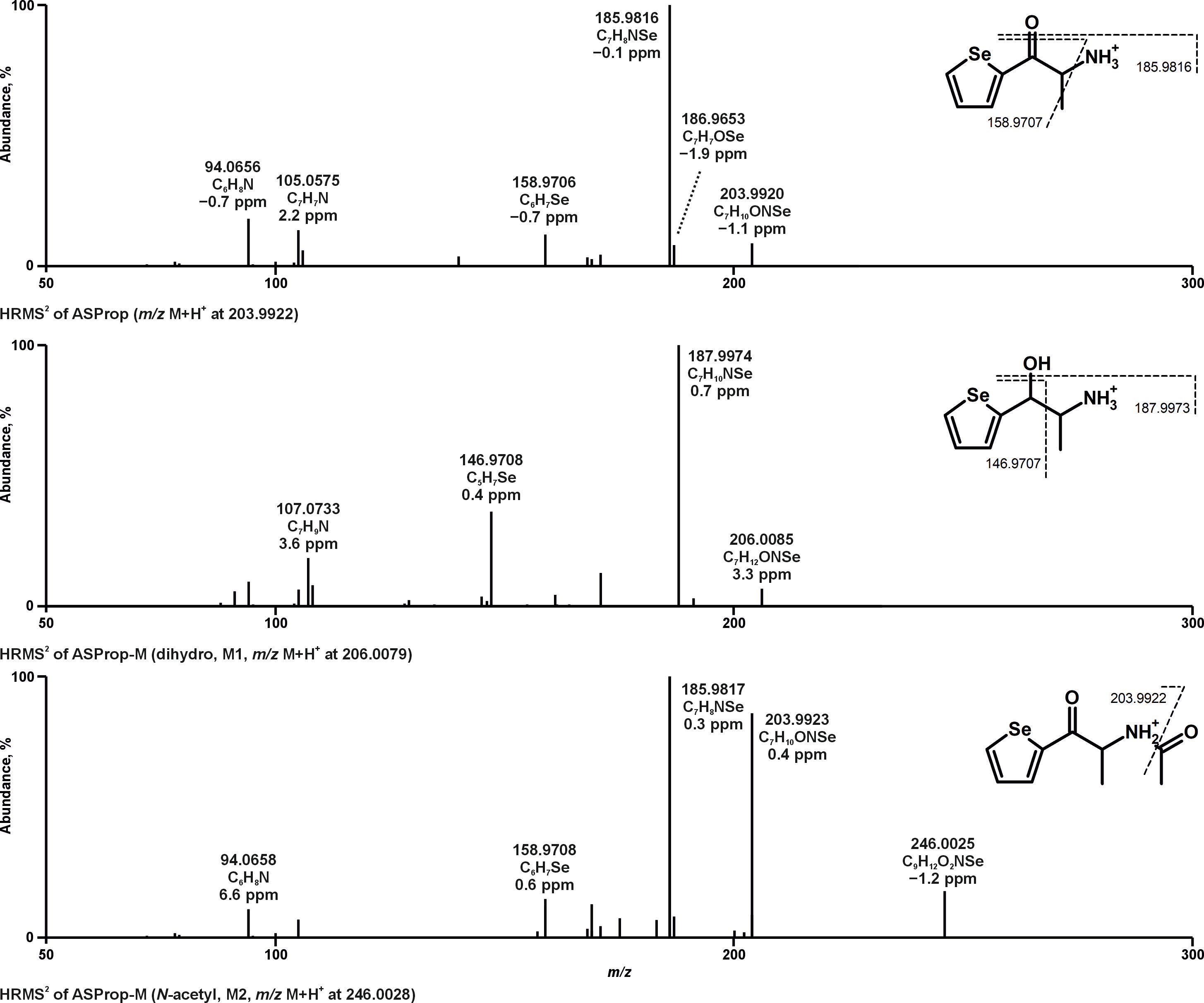
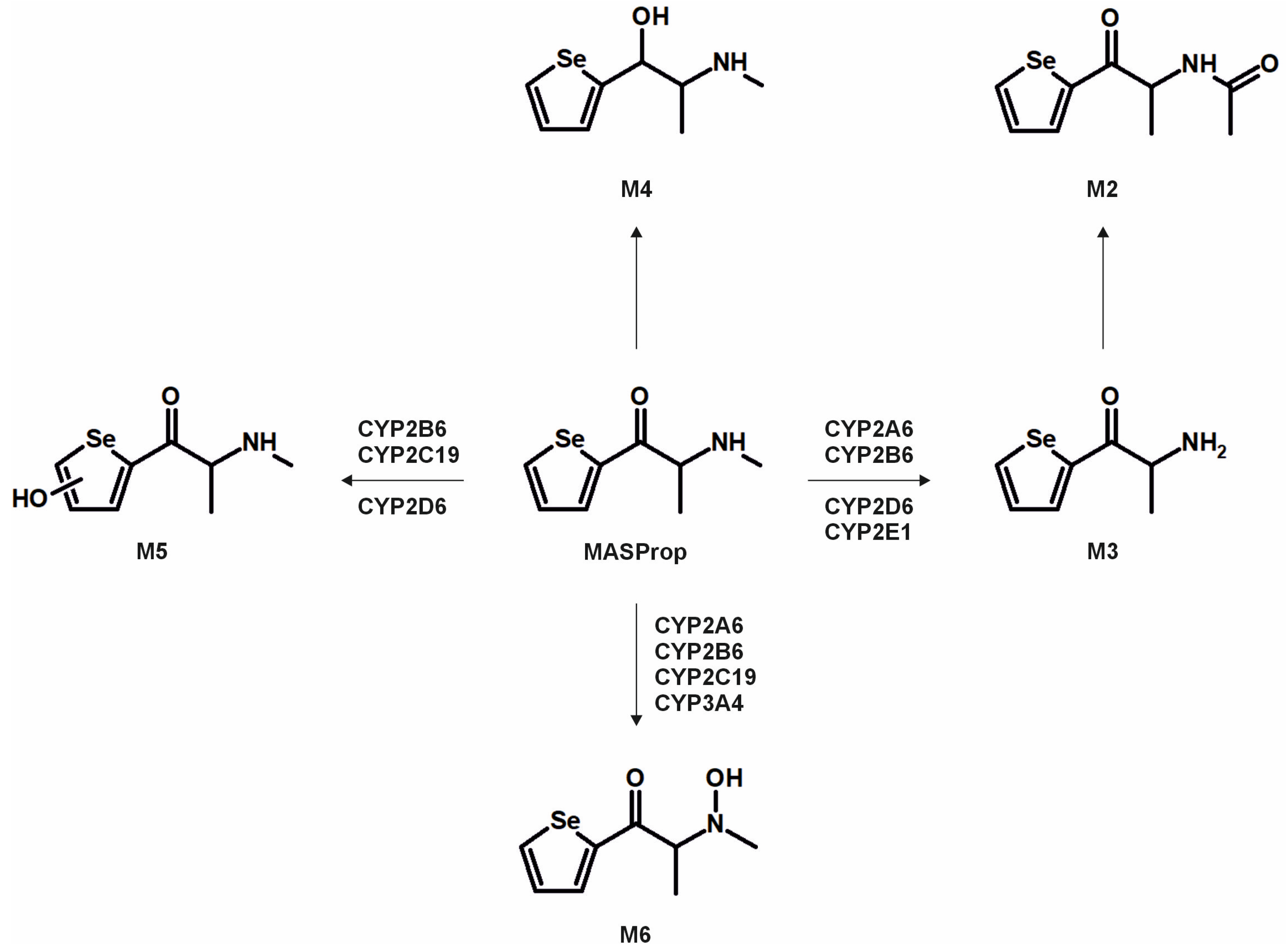
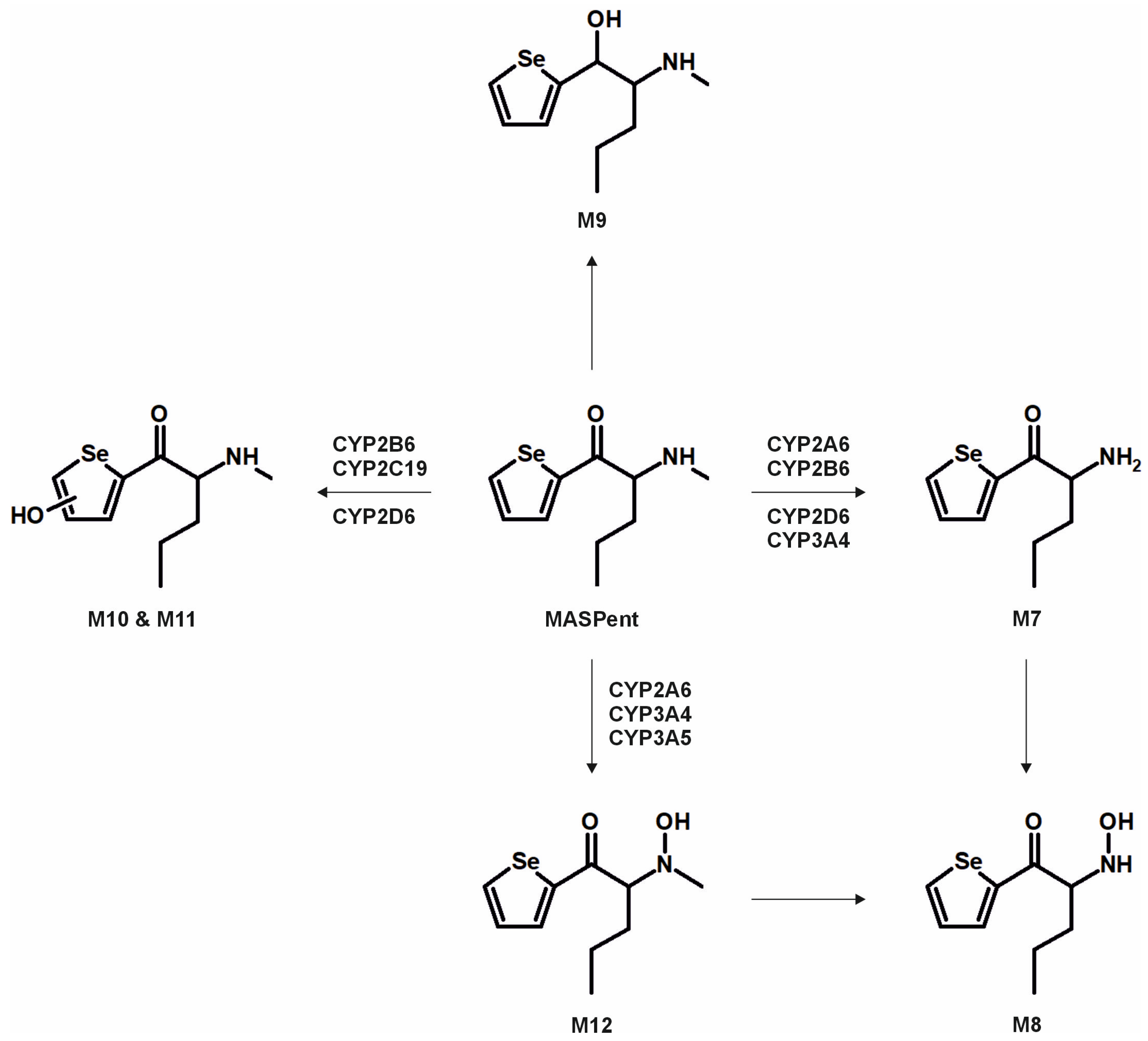
3. Results and Discussion
3.1. Identification of Metabolites
3.2. Metabolic Pathways and Isozyme Mapping
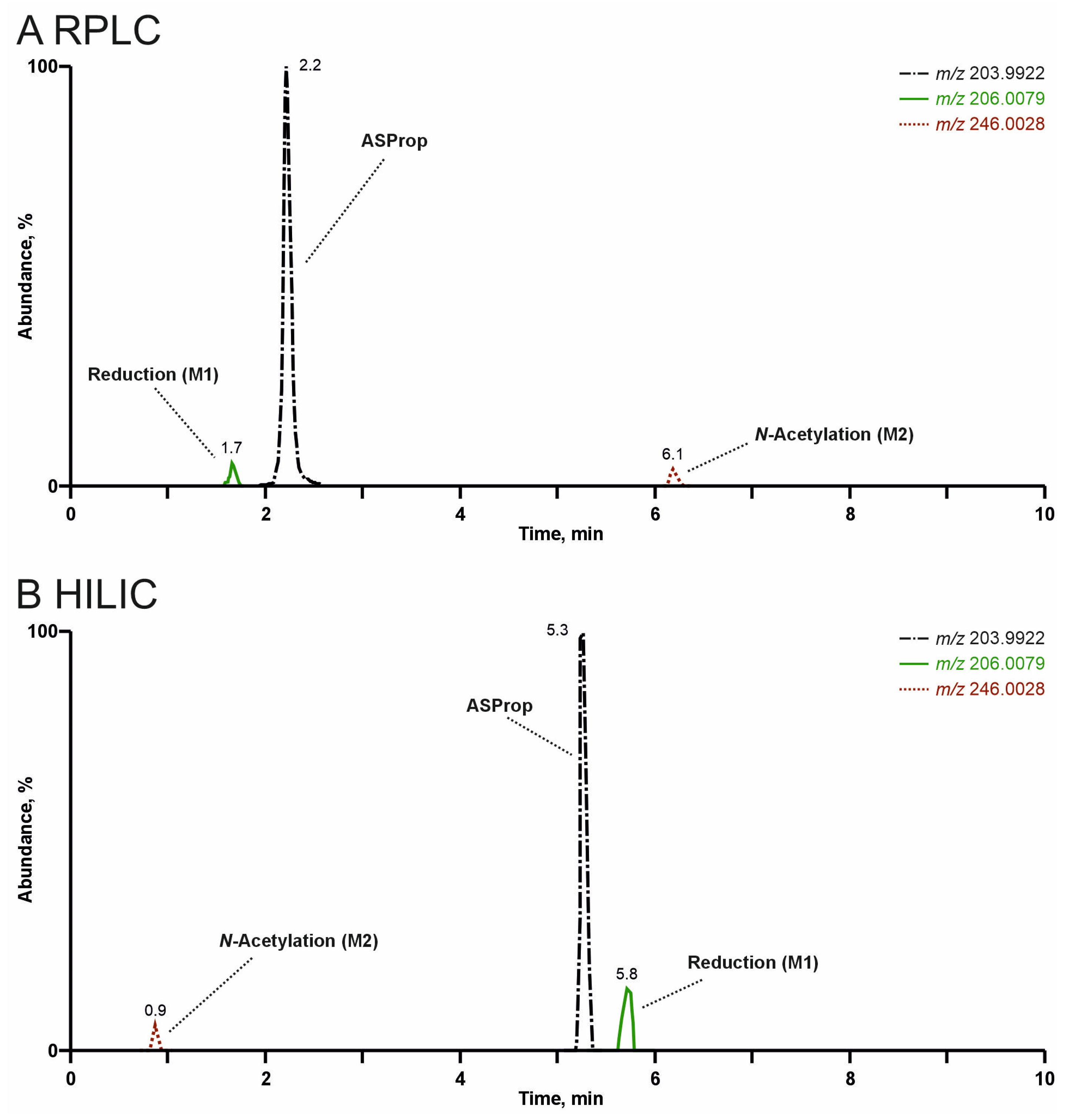
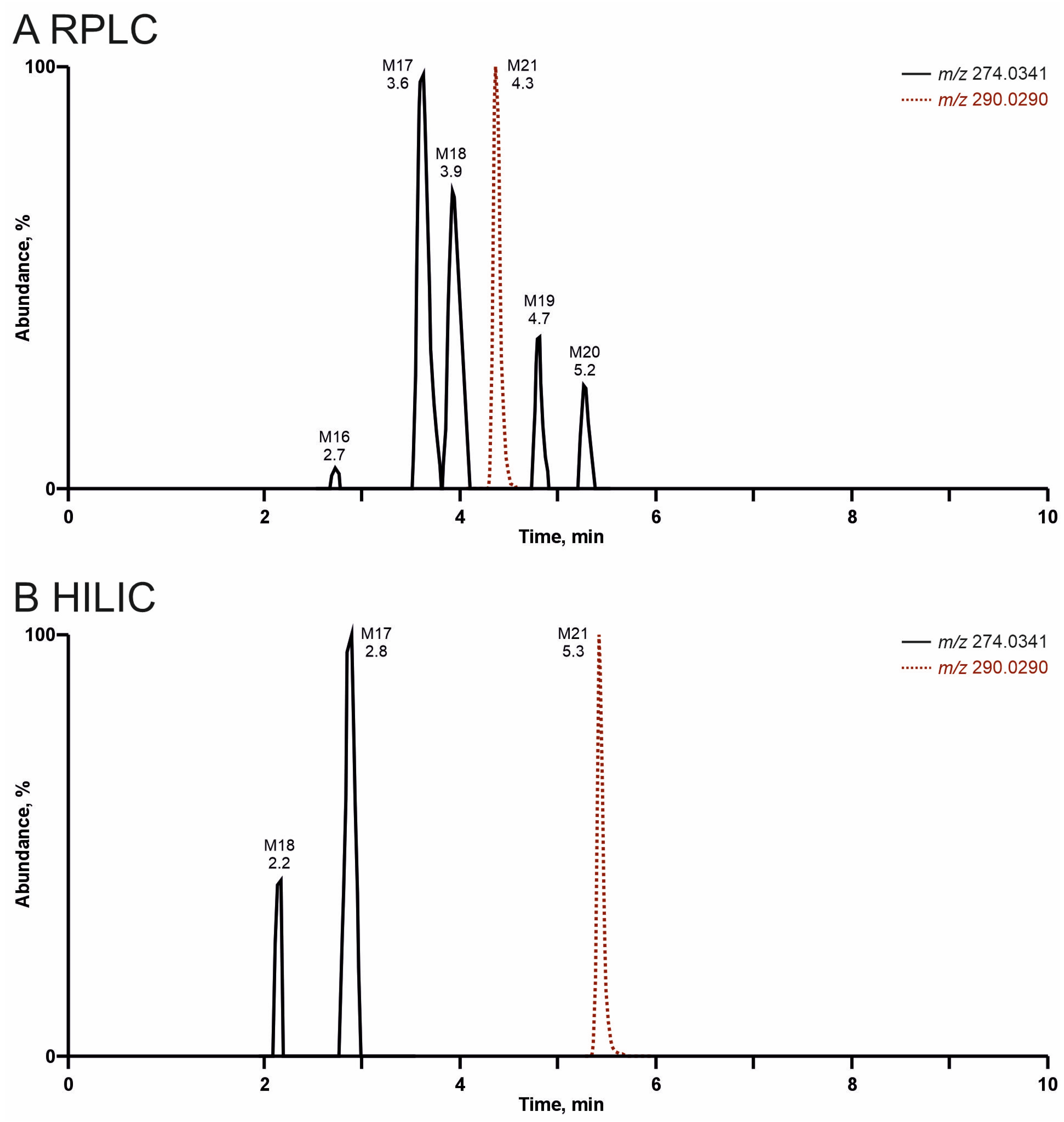
3.3. Comparison of Analyte Separation by RPLC or HILIC
3.4. Comparison of Metabolism Models and Screening Targets
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- UNODC. World Drug Report 2024. Available online: https://www.unodc.org/documents/data-and-analysis/WDR_2024/WDR_2024_SPI.pdf (accessed on 24 January 2025).
- Baumann, M.H.; Walters, H.M.; Niello, M.; Sitte, H.H. Neuropharmacology of Synthetic Cathinones. Handb. Exp. Pharmacol. 2018, 252, 113–142. [Google Scholar] [PubMed]
- Adamowicz, P. Blood concentrations of synthetic cathinones. Clin. Toxicol. 2021, 59, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Wagmann, L.; Manier, S.K.; Eckstein, N.; Maurer, H.H.; Meyer, M.R. Toxicokinetic studies of the four new psychoactive substances 4-chloroethcathinone, N-ethylnorpentylone, N-ethylhexedrone, and 4-fluoro-alpha-pyrrolidinohexiophenone. Forensic Toxicol. 2019, 38, 59–69. [Google Scholar] [CrossRef]
- Bunaim, M.K.; Damanhuri, H.A.; Yow, H.Y.; Yaakob, N.S.; Makmor-Bakry, M.; Azmi, N. Understanding methiopropamine, a new psychoactive substance: An in-depth review on its chemistry, pharmacology and implications to human health. Int. J. Legal Med. 2024, 138, 1295–1306. [Google Scholar] [CrossRef] [PubMed]
- Meltzer, P.C.; Butler, D.; Deschamps, J.R.; Madras, B.K. 1-(4-Methylphenyl)-2-pyrrolidin-1-yl-pentan-1-one (Pyrovalerone) analogues: A promising class of monoamine uptake inhibitors. J. Med. Chem. 2006, 49, 1420–1432. [Google Scholar] [CrossRef] [PubMed]
- Gambaro, V.; Casagni, E.; Dell’Acqua, L.; Roda, G.; Tamborini, L.; Visconti, G.L.; Demartin, F. Identification and characterization of a new designer drug thiothinone in seized products. Forensic Toxicol. 2016, 34, 174–178. [Google Scholar] [CrossRef]
- Norman, C.; Webling, K.; Stalberga, D.; Maas, L.; Tveit, J.; Liu, H.; Watanabe, S.; Vikingsson, S.; Green, H. In vitro metabolism of Benzyl-4CN-BUTINACA and MDMB-4CN-BUTINACA using human hepatocytes and LC-QToF-MS analysis. Arch. Toxicol. 2025, 99, 2355–2366. [Google Scholar] [CrossRef] [PubMed]
- Ji Ram, V.; Sethi, A.; Nath, M.; Pratap, R. Selenium-Based Heterocycles. In The Chemistry of Heterocycles; Ji Ram, V., Sethi, A., Nath, M., Pratap, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 469–484. [Google Scholar]
- Chuai, H.; Zhang, S.Q.; Bai, H.; Li, J.; Wang, Y.; Sun, J.; Wen, E.; Zhang, J.; Xin, M. Small molecule selenium-containing compounds: Recent development and therapeutic applications. Eur. J. Med. Chem. 2021, 223, 113621. [Google Scholar] [CrossRef] [PubMed]
- Debnath, S.; Agarwal, A.; Kumar, N.R.; Bedi, A. Selenium-Based Drug Development for Antioxidant and Anticancer Activity. Future Pharmacol. 2022, 2, 595–607. [Google Scholar] [CrossRef]
- Ali, W.; Benedetti, R.; Handzlik, J.; Zwergel, C.; Battistelli, C. The innovative potential of selenium-containing agents for fighting cancer and viral infections. Drug Discov. Today 2021, 26, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Angelov, D.; O’Brien, J.; Kavanagh, P. The syntheses of 1-(2-thienyl)-2-(methylamino) propane (methiopropamine) and its 3-thienyl isomer for use as reference standards. Drug Test. Anal. 2013, 5, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Wagmann, L.; Hempel, N.; Richter, L.H.J.; Brandt, S.D.; Stratford, A.; Meyer, M.R. Phenethylamine-derived new psychoactive substances 2C-E-FLY, 2C-EF-FLY, and 2C-T-7-FLY: Investigations on their metabolic fate including isoenzyme activities and their toxicological detectability in urine screenings. Drug Test. Anal. 2019, 11, 1507–1521. [Google Scholar] [CrossRef] [PubMed]
- Massano, M.; Nunez-Montero, M.; Papaseit, E.; Hladun, O.; Perez-Mana, C.; Ventura, M.; Marchei, E.; Alladio, E.; Gerace, E.; Pichini, S.; et al. Metabolic profile of N-ethylhexedrone, N-ethylpentedrone, and 4-chloromethcathinone in urine samples by UHPLC-QTOF-HRMS. J. Pharm. Biomed. Anal. 2024, 241, 115994. [Google Scholar] [CrossRef] [PubMed]
- Berardinelli, D.; Taoussi, O.; Daziani, G.; Tavoletta, F.; Ricci, G.; Tronconi, L.P.; Adamowicz, P.; Busardo, F.P.; Carlier, J. 3-CMC, 4-CMC, and 4-BMC Human Metabolic Profiling: New Major Pathways to Document Consumption of Methcathinone Analogues? AAPS J. 2024, 26, 70. [Google Scholar] [CrossRef] [PubMed]
- Niessen, W.M.; Correa, R.A. Interpretation of MS-MS Mass Spectra of Drugs and Pesticides; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Cashman, J.R.; Xiong, Y.N.; Xu, L.; Janowsky, A. N-oxygenation of amphetamine and methamphetamine by the human flavin-containing monooxygenase (form 3): Role in bioactivation and detoxication. J. Pharmacol. Exp. Ther. 1999, 288, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Manier, S.K.; Richter, L.H.J.; Schaper, J.; Maurer, H.H.; Meyer, M.R. Different in vitro and in vivo tools for elucidating the human metabolism of alpha-cathinone-derived drugs of abuse. Drug Test. Anal. 2018, 10, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Welter-Luedeke, J.; Maurer, H.H. New Psychoactive Substances: Chemistry, Pharmacology, Metabolism, and Detectability of Amphetamine Derivatives With Modified Ring Systems. Ther. Drug Monit. 2016, 38, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Tyrkko, E.; Pelander, A.; Ketola, R.A.; Ojanpera, I. In silico and in vitro metabolism studies support identification of designer drugs in human urine by liquid chromatography/quadrupole-time-of-flight mass spectrometry. Anal. Bioanal. Chem. 2013, 405, 6697–6709. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y. Potential of HILIC-MS in quantitative bioanalysis of drugs and drug metabolites. J. Sep. Sci. 2008, 31, 1481–1491. [Google Scholar] [CrossRef] [PubMed]
- Isokawa, M.; Kanamori, T.; Funatsu, T.; Tsunoda, M. Recent advances in hydrophilic interaction chromatography for quantitative analysis of endogenous and pharmaceutical compounds in plasma samples. Bioanalysis 2014, 6, 2421–2439. [Google Scholar] [CrossRef] [PubMed]
- Peters, F.T. Recent developments in urinalysis of metabolites of new psychoactive substances using LC-MS. Bioanalysis 2014, 6, 2083–2107. [Google Scholar] [CrossRef] [PubMed]
- Van Wichelen, N.; Estevez-Danta, A.; Belova, L.; den Ouden, F.; Verougstraete, N.; Roggeman, M.; Boogaerts, T.; Quireyns, M.; Robeyns, R.; De Brabanter, N.; et al. In vitro biotransformation of 3-methylmethcathinone (3-MMC) through incubation with human liver microsomes and cytosol and application to in vivo samples. J. Pharm. Biomed. Anal. 2024, 248, 116335. [Google Scholar] [CrossRef] [PubMed]
- Di Trana, A.; La Maida, N.; de la Rosa, G.; Di Giorgi, A.; Graziano, S.; Aldhaehri, K.; Papaseit, E.; Hladun, O.; Farre, M.; Perez, C.; et al. Early and Mid-Term Disposition of alpha-PVP and its unknown Metabolites in Urine and Oral Fluid Through a Multi-Analytical Hyphenated Approach Following a Single Non-Controlled Administration to Healthy Volunteers. AAPS J. 2025, 27, 25. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagmann, L.; Schmitt, J.H.; Gampfer, T.M.; Brandt, S.D.; Scott, K.; Kavanagh, P.V.; Meyer, M.R. A Comparative Study Using Reversed-Phase and Hydrophilic Interaction Liquid Chromatography to Investigate the In Vitro and In Vivo Metabolism of Five Selenium-Containing Cathinone Derivatives. Metabolites 2025, 15, 497. https://doi.org/10.3390/metabo15080497
Wagmann L, Schmitt JH, Gampfer TM, Brandt SD, Scott K, Kavanagh PV, Meyer MR. A Comparative Study Using Reversed-Phase and Hydrophilic Interaction Liquid Chromatography to Investigate the In Vitro and In Vivo Metabolism of Five Selenium-Containing Cathinone Derivatives. Metabolites. 2025; 15(8):497. https://doi.org/10.3390/metabo15080497
Chicago/Turabian StyleWagmann, Lea, Jana H. Schmitt, Tanja M. Gampfer, Simon D. Brandt, Kenneth Scott, Pierce V. Kavanagh, and Markus R. Meyer. 2025. "A Comparative Study Using Reversed-Phase and Hydrophilic Interaction Liquid Chromatography to Investigate the In Vitro and In Vivo Metabolism of Five Selenium-Containing Cathinone Derivatives" Metabolites 15, no. 8: 497. https://doi.org/10.3390/metabo15080497
APA StyleWagmann, L., Schmitt, J. H., Gampfer, T. M., Brandt, S. D., Scott, K., Kavanagh, P. V., & Meyer, M. R. (2025). A Comparative Study Using Reversed-Phase and Hydrophilic Interaction Liquid Chromatography to Investigate the In Vitro and In Vivo Metabolism of Five Selenium-Containing Cathinone Derivatives. Metabolites, 15(8), 497. https://doi.org/10.3390/metabo15080497