
Compound 3d Attenuates Metabolic Dysfunction-Associated Steatohepatitis via Peroxisome Proliferator-Activated Receptor Pathway Activation and Inhibition of Inflammatory and Apoptotic Signaling

 , , , , , , , , and
, , , , , , , , and
Abstract

1. Introduction
2. Materials and Methods
2.1. In Vitro Lipid Accumulation Model
2.2. In Vitro Inflammation Model
2.3. Cell Apoptosis-Related Assays
2.4. Animal Studies and Histological Analysis
2.5. Biochemical Analysis of Serum and Liver Tissue
2.6. The Sequencing of the Proteome and the 16S rDNA
2.7. Western Blotting Analysis
2.8. In Vitro Stability and Pharmacokinetic Studies of 3d
2.9. Statistical Analysis
3. Results
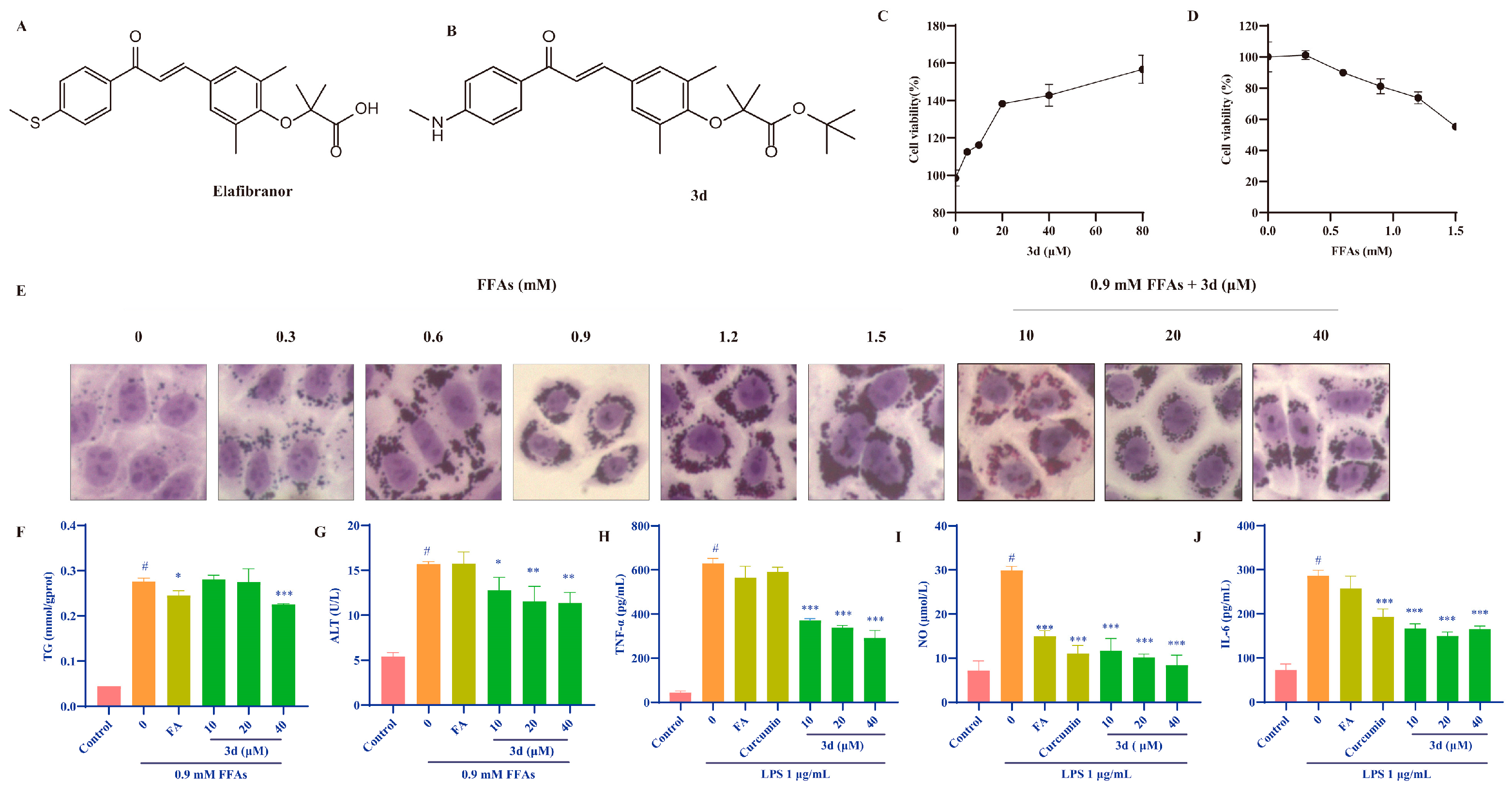
3.1. D Reduced Lipid Accumulation and Inhibited Inflammatory Response In Vitro
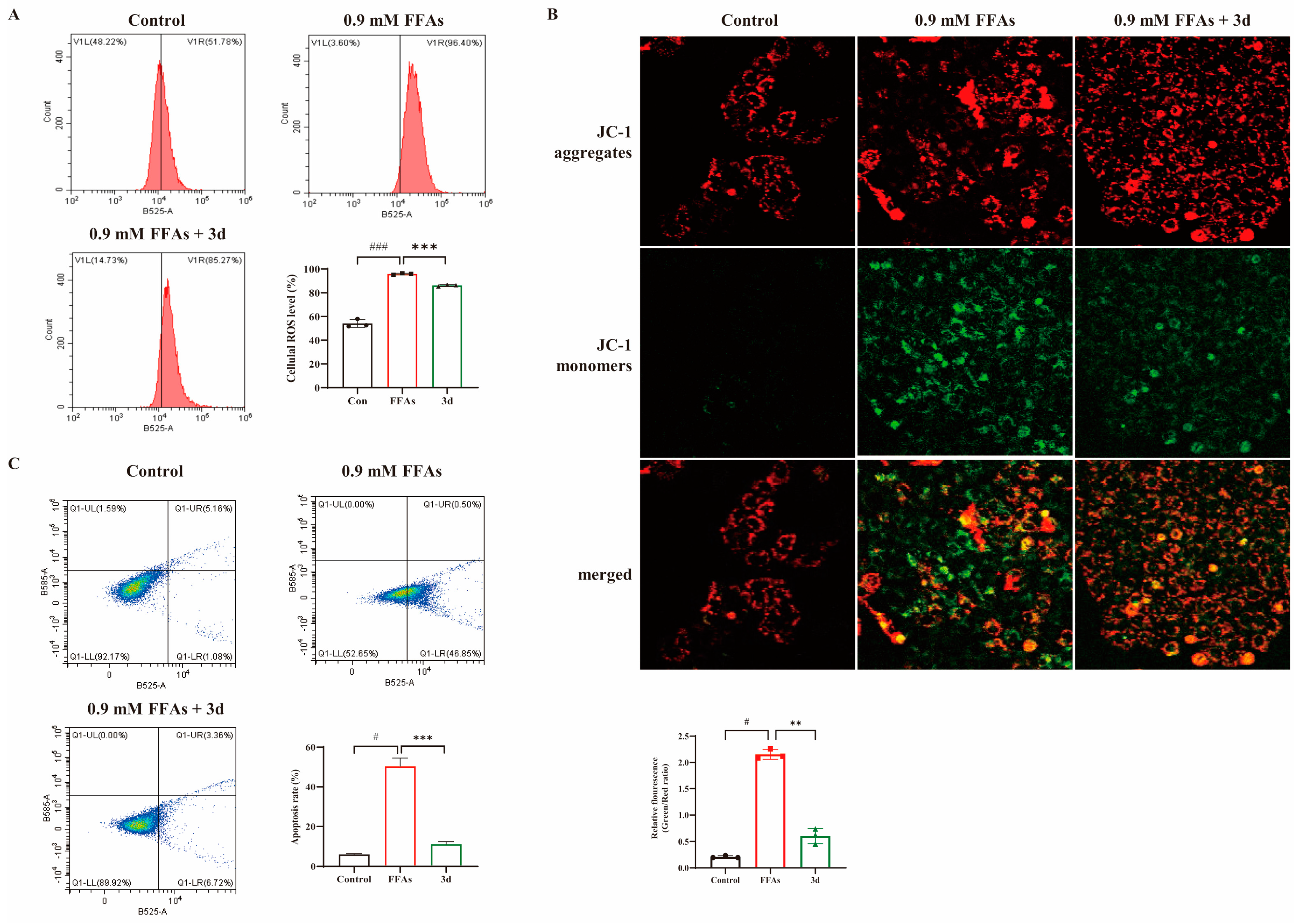
3.2. D Maintained Mitochondrial Function and Attenuated Apoptosis In Vitro
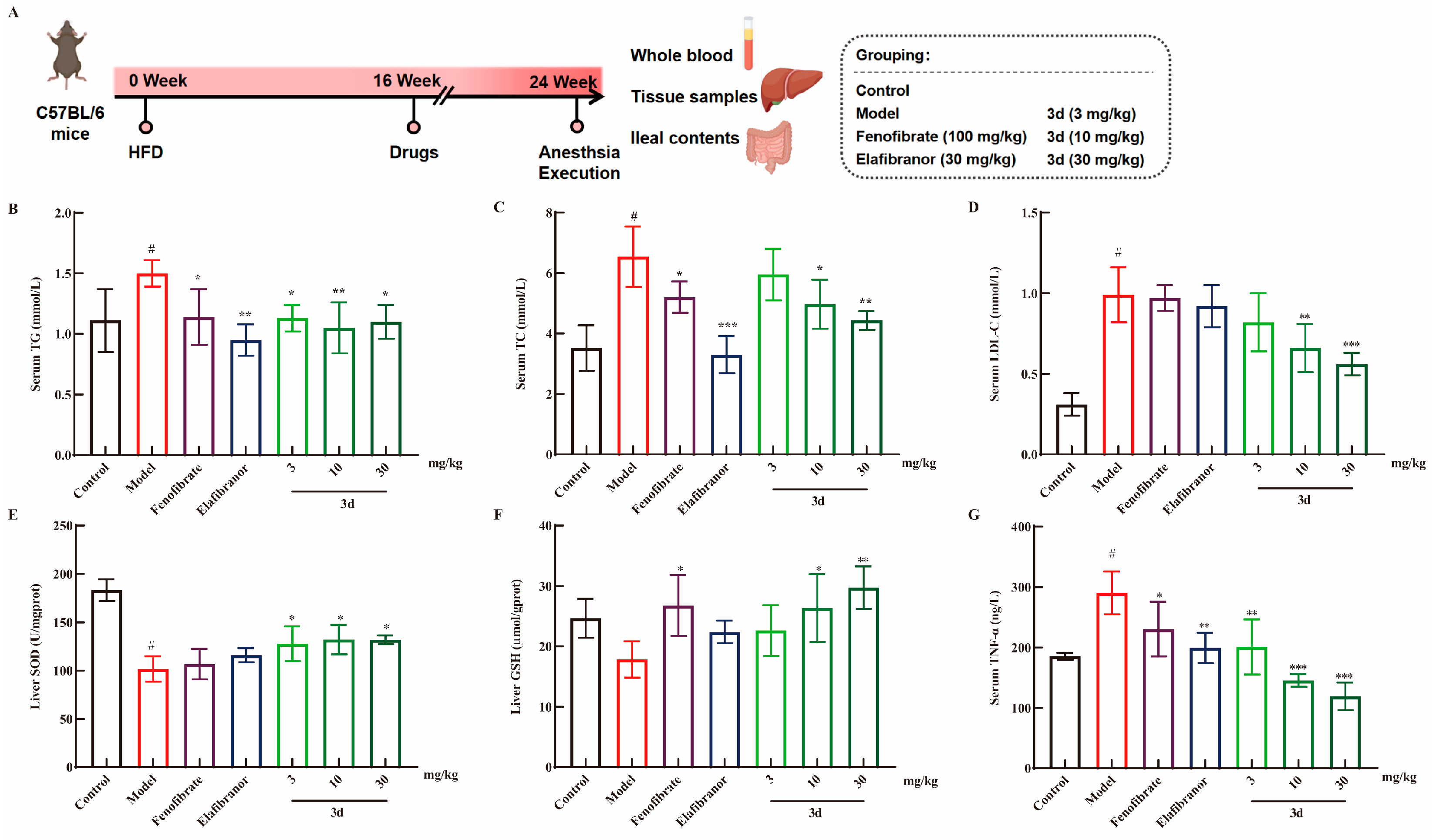
3.3. D Effectively Inhibited HFD-Induced MASH Model in Mice
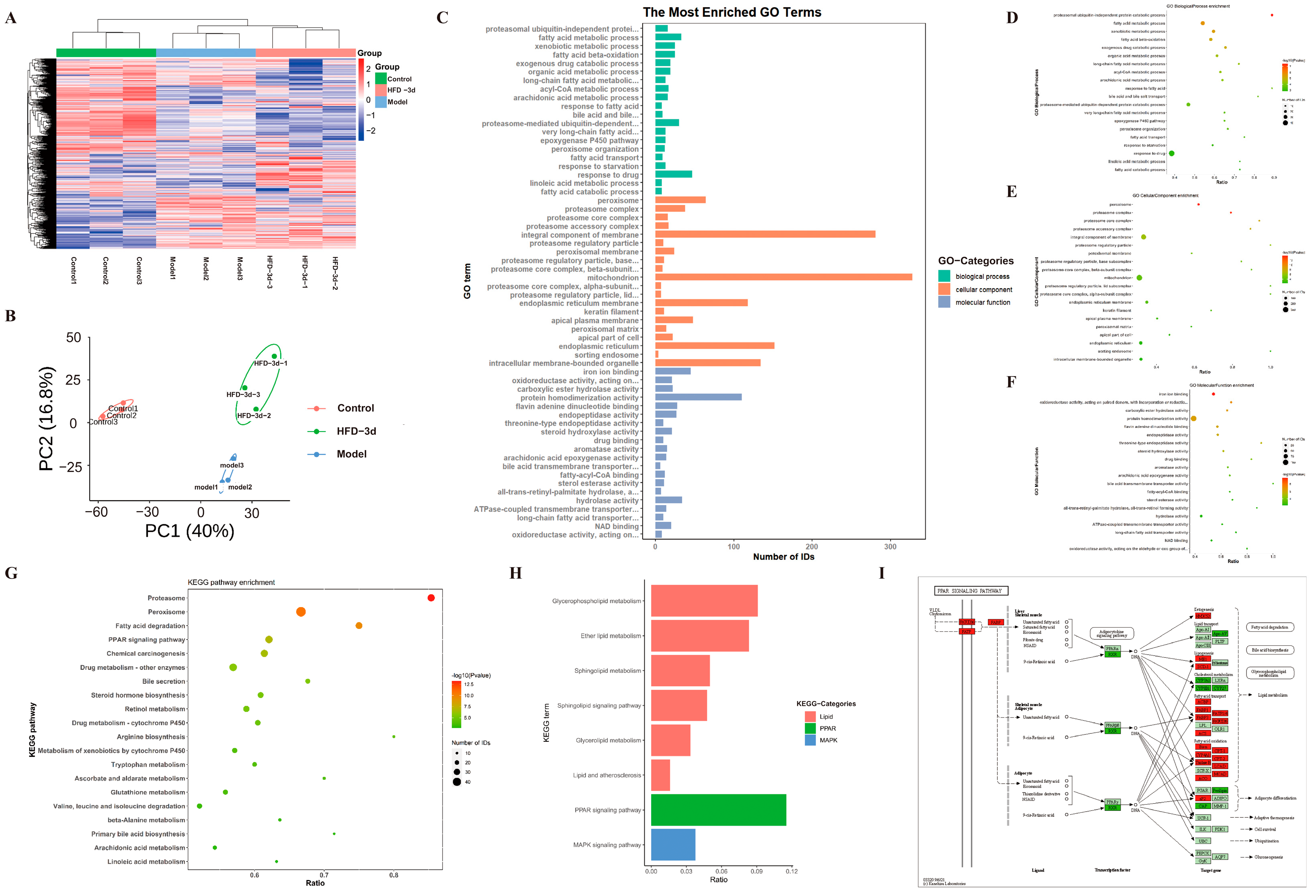
3.4. D Regulated Protein Expression in MASH Mice
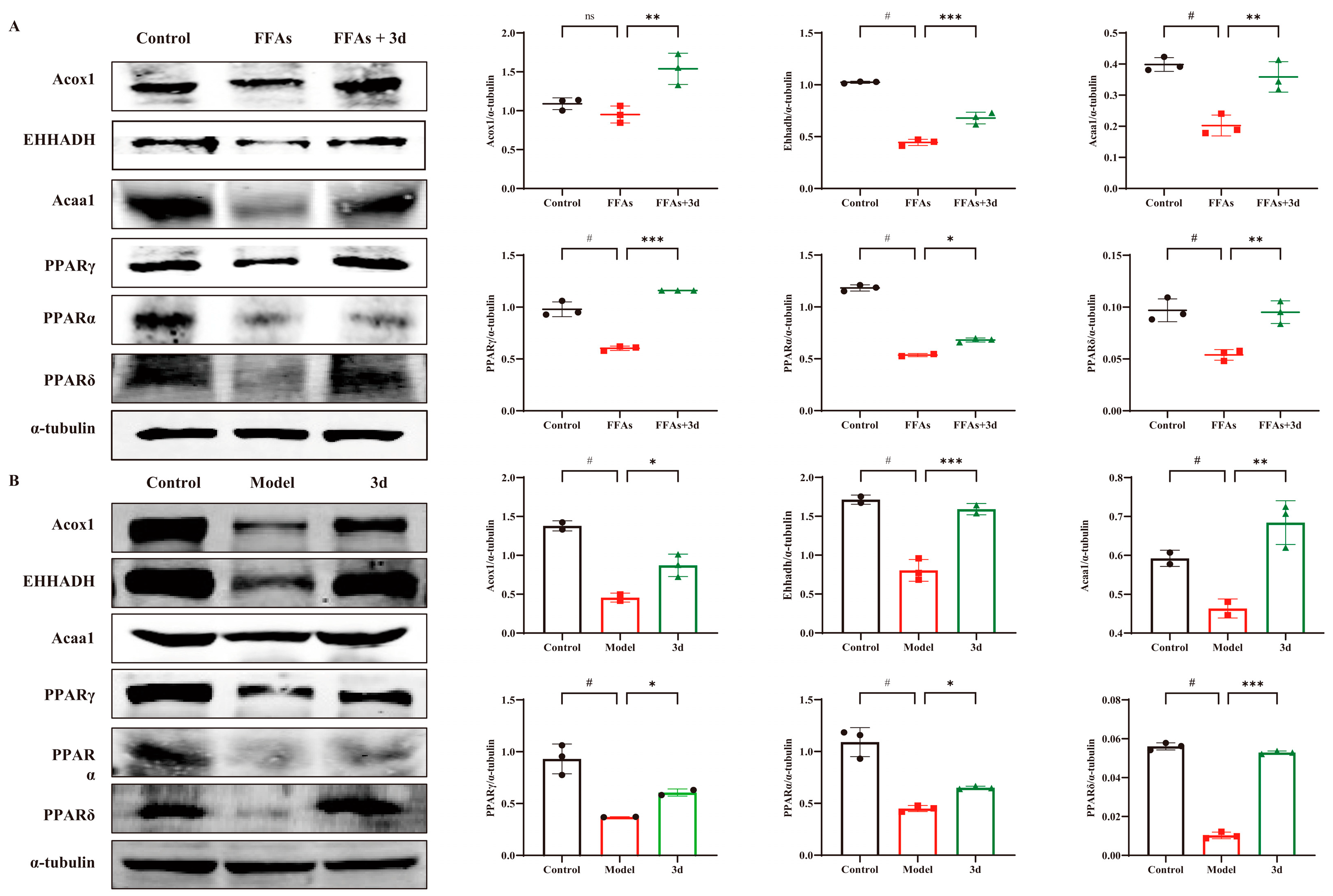
3.5. D Regulated PPAR Pathway to Inhibit Progression of MASH
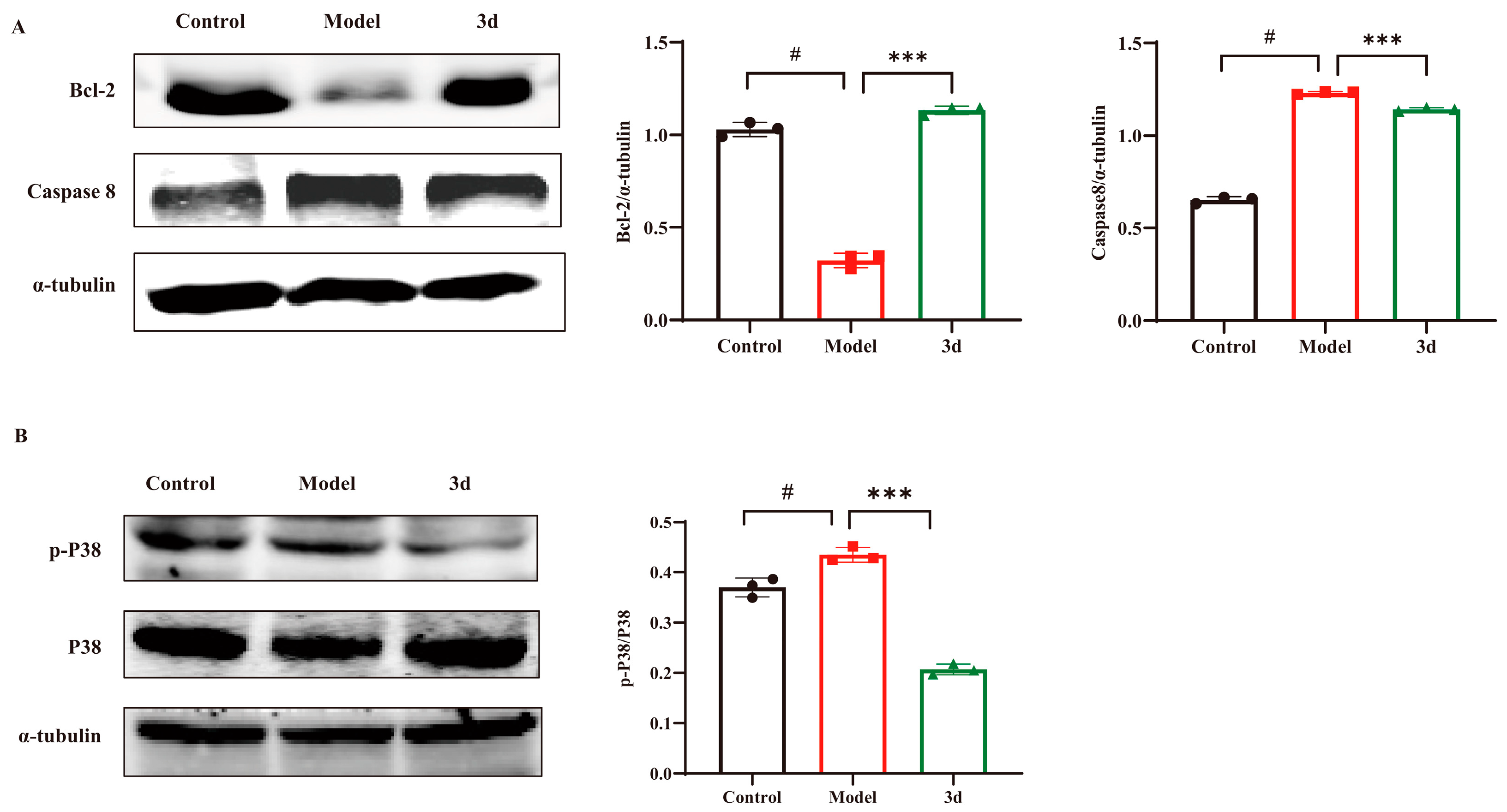
3.6. D Inhibited Apoptosis via the P38 Inflammatory Pathway
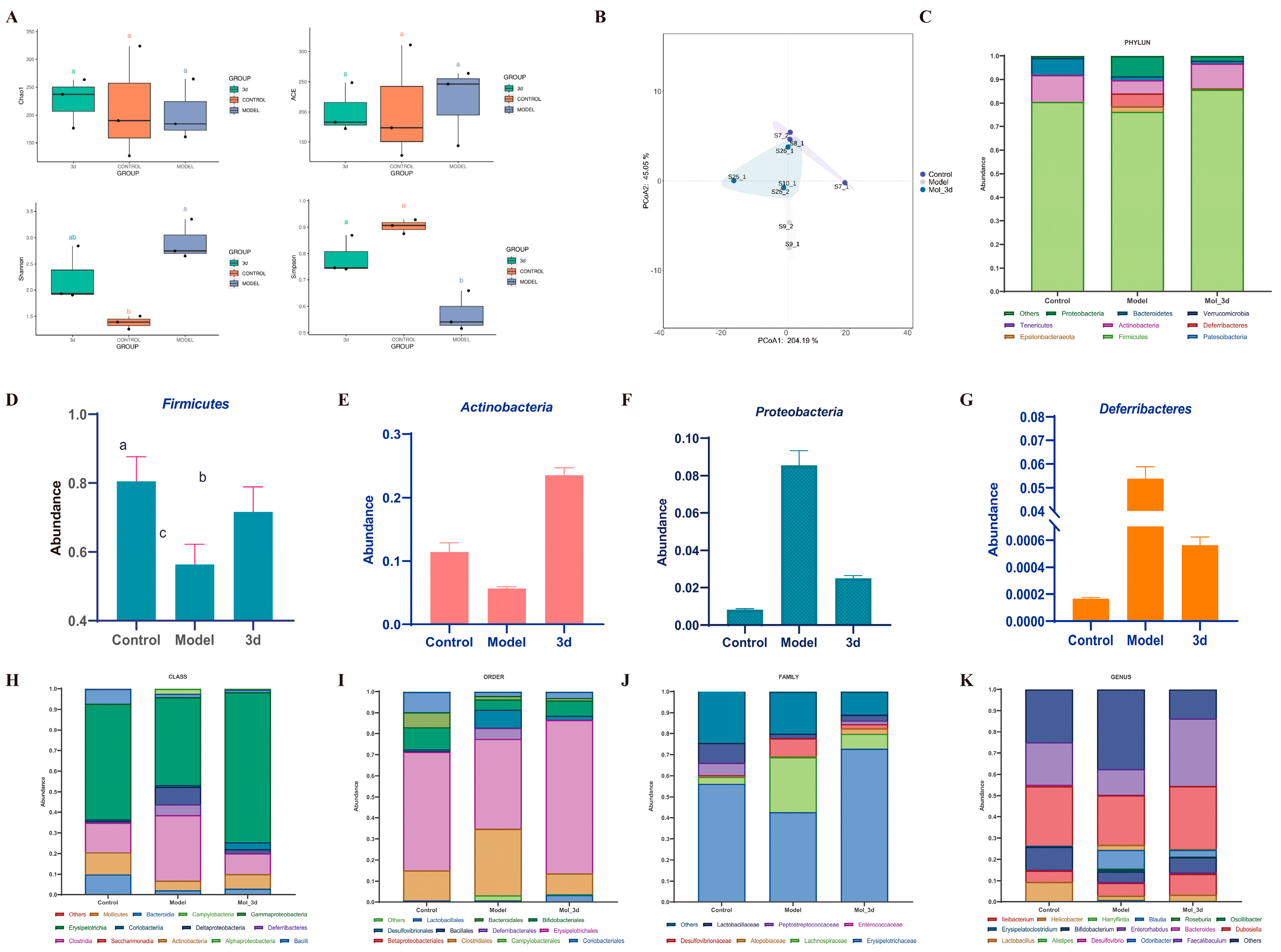
3.7. D Improves MASH by Modulating Gut Microbiota
3.8. D Has Good Pharmacokinetic Properties
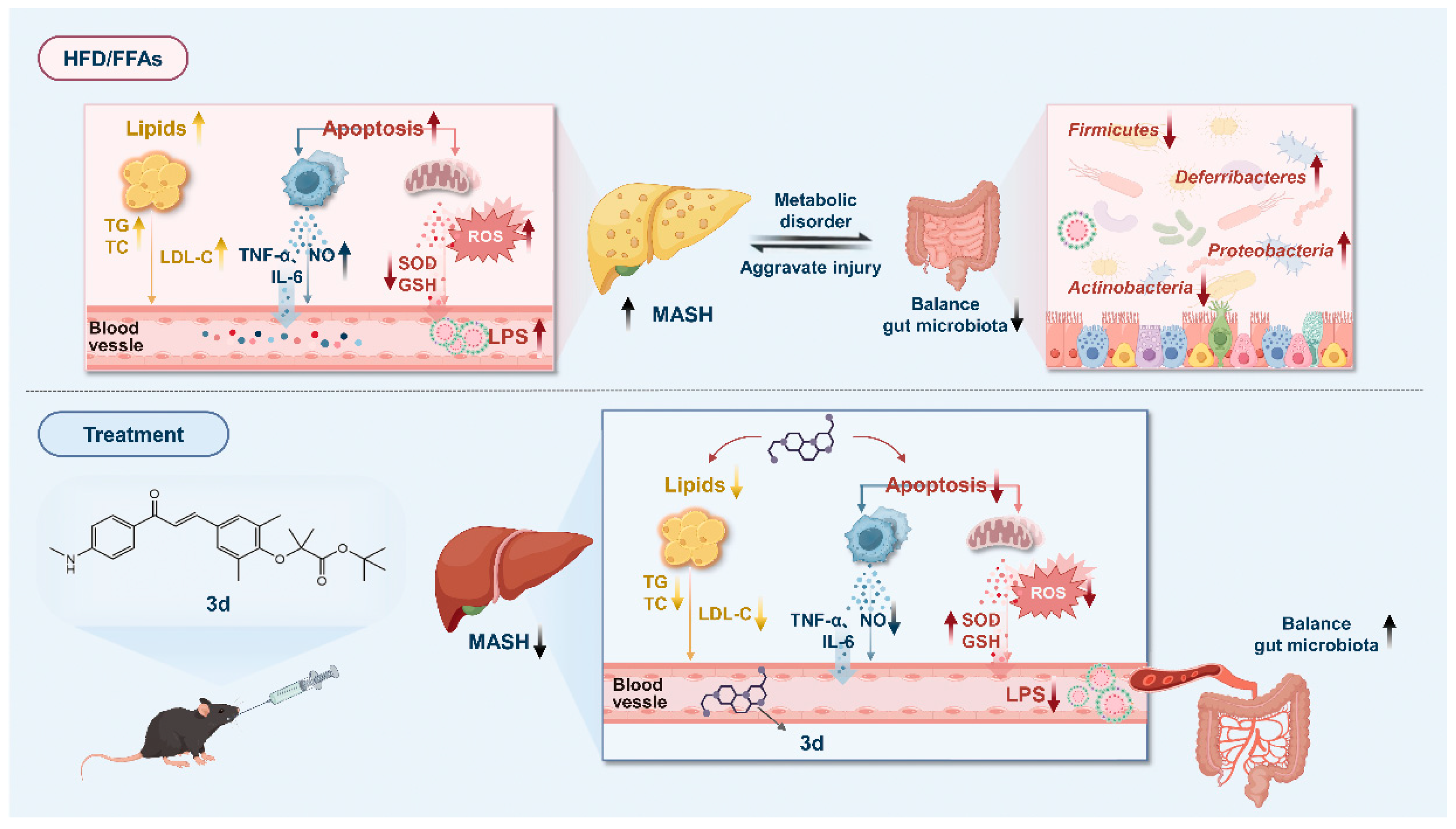
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Van Meeteren, M.J.W.; Drenth, J.P.H.; Tjwa, E. Elafibranor: A potential drug for the treatment of nonalcoholic steatohepatitis (NASH). Expert. Opin. Investig. Drugs 2020, 29, 117–123. [Google Scholar] [CrossRef]
- Loomba, R.; Friedman, S.L.; Shulman, G.I. Mechanisms and disease consequences of nonalcoholic fatty liver disease. Cell 2021, 184, 2537–2564. [Google Scholar] [CrossRef] [PubMed]
- Powell, E.E.; Wong, V.W.S.; Rinella, M. Non-alcoholic fatty liver disease. Lancet 2021, 397, 2212–2224. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Tan, H.B.; Wan, J.; Zeng, Y.; Wang, J.C.; Wang, H.C.; Lu, X.J. PPAR-? signaling in nonalcoholic fatty liver disease: Pathogenesis and therapeutic targets. Pharmacol. Ther. 2023, 245, 108391. [Google Scholar] [CrossRef] [PubMed]
- Bervejillo, M.L.; Ferreira, A.M. Understanding Peroxisome Proliferator-Activated Receptors: From the Structure to the Regulatory Actions on Metabolism. Adv. Exp. Med. Biol. 2019, 1127, 39–57. [Google Scholar]
- Lange, N.F.; Graf, V.; Caussy, C.; Dufour, J.F. PPAR-Targeted Therapies in the Treatment of Non-Alcoholic Fatty Liver Disease in Diabetic Patients. Int. J. Mol. Sci. 2022, 23, 4305. [Google Scholar] [CrossRef]
- Christofides, A.; Konstantinidou, E.; Jani, C.; Boussiotis, V.A. The role of peroxisome proliferator-activated receptors (PPAR) in immune responses. Metab.-Clin. Exp. 2021, 114, 154338. [Google Scholar] [CrossRef]
- Cooreman, M.P.; Vonghia, L.; Francque, S.M. MASLD/MASH and type 2 diabetes: Two sides of the same coin? From single PPAR to pan-PPAR agonists. Diabetes Res. Clin. Pract. 2024, 212, 111688. [Google Scholar] [CrossRef]
- Hernandez-Quiles, M.; Broekema, M.F.; Kalkhoven, E. PPARgamma in Metabolism, Immunity, and Cancer: Unified and Diverse Mechanisms of Action. Front. Endocrinol. 2021, 12, 624112. [Google Scholar] [CrossRef]
- Yu, H.J.; Wang, L.J.; Huang, K.; Guo, Q.F.; Lin, B.Y.; Liu, Y.Y.; Yu, M.; Ma, G.P. PPAR-γ agonist pioglitazone alleviates inflammatory response induced by lipopolysaccharides in osteoblast cells. J. Orthop. Res. 2022, 40, 2471–2479. [Google Scholar] [CrossRef]
- Meng, X.; Wang, L.; Du, Y.C.; Cheng, D.; Zeng, T. PPARβ/δ as a promising molecular drug target for liver diseases: A focused review. Clin. Res. Hepatol. Gastroenterol. 2024, 48, 102343. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.J.; Barroso, E.; Ruart, M.; Peña, L.; Peyman, M.; Aguilar-Recarte, D.; Montori-Grau, M.; Rada, P.; Cugat, C.; Montironi, C.; et al. Elafibranor upregulates the EMT-inducer S100A4 via PPARβ/δ. Biomed. Pharmacother. 2023, 167, 115623. [Google Scholar] [CrossRef]
- Boeckmans, J.; Gatzios, A.; Heymans, A.; Rombaut, M.; Rogiers, V.; De Kock, J.; Vanhaecke, T.; Rodrigues, R.M. Transcriptomics Reveals Discordant Lipid Metabolism Effects between In Vitro Models Exposed to Elafibranor and Liver Samples of NAFLD Patients after Bariatric Surgery. Cells 2022, 11, 893. [Google Scholar] [CrossRef]
- Blair, H.A. Elafibranor: First Approval. Drugs 2024, 84, 1143–1148. [Google Scholar] [CrossRef]
- Kowdley, K.V.; Bowlus, C.L.; Levy, C.; Akarca, U.S.; Alvares-da-Silva, M.R.; Andreone, P.; Arrese, M.; Corpechot, C.; Francque, S.M.; Heneghan, M.A.; et al. Efficacy and Safety of Elafibranor in Primary Biliary Cholangitis. N. Engl. J. Med. 2024, 390, 795–805. [Google Scholar] [CrossRef] [PubMed]
- Xiang, C.; Chen, X.; Yao, J.; Yang, N.; Yu, J.; Qiu, Q.; Zhang, S.; Kong, X.; Zhao, L.; Fan, Z.C.; et al. Design, synthesis and anti-NASH effect evaluation of novel GFT505 derivatives in vitro and in vivo. Eur. J. Med. Chem. 2023, 257, 115510. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Shen, G.; Xu, Y.; Liu, H.; Zhang, J.; Li, S.; Li, J.; Zhang, C.; Qi, J.; Wang, L.; et al. Extensive Inter-plant Protein Transfer between Cuscuta Parasites and Their Host Plants. Mol. Plant 2020, 13, 573–585. [Google Scholar] [CrossRef]
- Windriyati, Y.N.; Sumirtapura, Y.C.; Pamudji, J.S. Comparative In Vitro and In Vivo Evaluation of Fenofibric Acid as an Antihyperlipidemic Drug. Turk. J. Pharm. Sci. 2020, 17, 203–210. [Google Scholar] [CrossRef]
- Peng, Y.; Ao, M.Y.; Dong, B.H.; Jiang, Y.X.; Yu, L.Y.; Chen, Z.M.; Hu, C.J.; Xu, R.C. Anti-Inflammatory Effects of Curcumin in the Inflammatory Diseases: Status, Limitations and Countermeasures. Drug Des. Dev. Ther. 2021, 15, 4503–4525. [Google Scholar] [CrossRef]
- Li, R.B.; Xin, T.; Li, D.D.; Wang, C.B.; Zhu, H.; Zhou, H. Therapeutic effect of Sirtuin 3 on ameliorating nonalcoholic fatty liver disease: The role of the ERK-CREB pathway and Bnip3-mediated mitophagy. Redox Biol. 2018, 18, 229–243. [Google Scholar] [CrossRef]
- Zhou, H.; Li, D.D.; Zhu, P.J.; Ma, Q.; Toan, S.; Wang, J.; Hu, S.Y.; Chen, Y.D.; Zhang, Y.M. Inhibitory effect of melatonin on necroptosis via repressing the Ripk3-PGAM5-CypD-mPTP pathway attenuates cardiac microvascular ischemia-reperfusion injury. J. Pineal Res. 2018, 65, 12503. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Geng, T.H.; Zou, Q.H.; Yang, N.R.; Zhao, W.Y.; Li, Y.T.; Tan, X.T.; Yuan, T.; Liu, X.B.; Liu, Z.G. Lycopene prevents lipid accumulation in hepatocytes by stimulating PPARα and improving mitochondrial function. J. Funct. Foods 2020, 67, 103857. [Google Scholar] [CrossRef]
- Lee, E.; Korf, H.; Vidal-Puig, A. An adipocentric perspective on the development and progression of non-alcoholic fatty liver disease. J. Hepatol. 2023, 78, 1048–1062. [Google Scholar] [CrossRef]
- Dong, L.Z.; Qin, C.; Li, Y.; Wu, Z.F.; Liu, L.L. Oat phenolic compounds regulate metabolic syndrome in high fat diet-fed mice via gut microbiota. Food Biosci. 2022, 50, 101946. [Google Scholar] [CrossRef]
- Bittner, V.A.; Szarek, M.; Aylward, P.; Bhatt, D.L.; Diaz, R.; Edelberg, J.M.; Fras, Z.; Goodman, S.G.; Halvorsen, S.; Hanotin, C.; et al. Effect of Alirocumab on Lipoprotein(a) and Cardiovascular Risk After Acute Coronary Syndrome. J. Am. Coll. Cardiol. 2020, 75, 133–144. [Google Scholar] [CrossRef]
- Benedé-Ubieto, R.; Cubero, F.J.; Nevzorova, Y.A. Breaking the barriers: The role of gut homeostasis in Metabolic-Associated Steatotic Liver Disease (MASLD). Gut Microbes 2024, 16, 2331460. [Google Scholar] [CrossRef] [PubMed]
- Zazueta, A.; Valenzuela-Perez, L.; Ortiz-Lopez, N.; Pinto-Leon, A.; Torres, V.; Guinez, D.; Aliaga, N.; Merino, P.; Sandoval, A.; Covarrubias, N.; et al. Alteration of Gut Microbiota Composition in the Progression of Liver Damage in Patients with Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD). Int. J. Mol. Sci. 2024, 25, 4387. [Google Scholar] [CrossRef]
- Paul, B.; Lewinska, M.; Andersen, J.B. Lipid alterations in chronic liver disease and liver cancer. JHEP Rep. 2022, 4, 100479. [Google Scholar] [CrossRef]
- Ratziu, V.; Harrison, S.A.; Francque, S.; Bedossa, P.; Lehert, P.; Serfaty, L.; Romero-Gomez, M.; Boursier, J.; Abdelmalek, M.; Caldwell, S.; et al. Elafibranor, an Agonist of the Peroxisome Proliferator-Activated Receptor-α and -δ, Induces Resolution of Nonalcoholic Steatohepatitis Without Fibrosis Worsening. Gastroenterol. 2016, 150, 1147–1159.e1145. [Google Scholar] [CrossRef]
- Kotsiliti, E. Elafibranor in primary biliary cholangitis. Nat. Rev. Gastroenterol. Hepatol. 2024, 21, 2. [Google Scholar] [CrossRef]
- Yang, W.; Ling, X.; He, S.J.; Cui, H.A.; Yang, Z.Y.; An, H.H.; Wang, L.H.; Zou, P.; Chen, Q.; Liu, J.Y.; et al. PPARa/ACOX1 as a novel target for hepatic lipid metabolism disorders induced by per- and polyfluoroalkyl substances: An integrated approach. Environ. Int. 2023, 178, 108138. [Google Scholar] [CrossRef]
- He, A.Y.; Dean, J.M.; Lu, D.L.; Chen, Y.L.; Lodhi, I.J. Hepatic peroxisomal β-oxidation suppresses lipophagy via RPTOR acetylation and MTOR activation. Autophagy 2020, 16, 1727–1728. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.F.; Liu, C.; Zhao, M.Y.; Zhang, Q.R.; Lu, Y.; Liu, P.; Yang, H.; Yang, J.L.; Chen, X.X.; Yao, Y.Q. The pharmacodynamic and differential gene expression analysis of PPAR α/δ agonist GFT505 in CDAHFD-induced NASH model. PLoS ONE 2020, 15, 0243911. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.H.; Wu, H.R.; Wang, Z.Y.; Wu, J.S.; Ying, S.B.; Huang, M.J.; Li, Y.M. Integrated expression profiles of mRNA and miRNA in a gerbil model of fatty liver fibrosis treated with exenatide. Clin. Res. Hepatol. Gastroenterol. 2021, 45, 101312. [Google Scholar] [CrossRef]
- Zhang, X.; Fan, L.N.; Wu, J.F.; Xu, H.Z.; Leung, W.Y.; Fu, K.L.; Wu, J.T.; Liu, K.; Man, W.; Yang, X.Y.; et al. Macrophage p38α promotes nutritional steatohepatitis through M1 polarization. J. Hepatol. 2019, 71, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Jung, K.; Lee, J.; Kang, H.J.; Lee, J.Y.; Kim, J.; Ham, D.; Cho, J.; Eom, D.W.; Kang, K.S. Role of 11β-hydroxysteroid dehydrogenase type 1 inhibition in the antiobesity effect of J2H-1702 on adipocytes and a high-fat diet-induced NASH model. Eur. J. Pharmacol. 2025, 989, 177272. [Google Scholar] [CrossRef]
- Caligiuri, A.; Gentilini, A.; Pastore, M.; Gitto, S.; Marra, F. Cellular and Molecular Mechanisms Underlying Liver Fibrosis Regression. Cells 2021, 10, 2759. [Google Scholar] [CrossRef]
- Lai, M.Y.; Li, N.; Li, Q.C.; Ge, X.D.; Huang, Z.F.; Chen, F.J.; Liu, B.; Zeng, F. Pueraria lobata Resistant Starch Regulating Lipid Metabolism and Gut Microbiota in High-Fat Diet Mice. Starch-Starke 2024, 76, 2300123. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, N.; Xiao, Y.Y.; Yuan, K.F.; Wang, Z. The role of gut microbiota in liver regeneration. Front. Immunol. 2022, 13, 1003376. [Google Scholar] [CrossRef]
- Song, Z.W.; Cai, Y.Y.; Lao, X.Z.; Wang, X.; Lin, X.X.; Cui, Y.Y.; Kalavagunta, P.K.; Liao, J.; Jin, L.; Shang, J.; et al. Taxonomic profiling and populational patterns of bacterial bile salt hydrolase (BSH) genes based on worldwide human gut microbiome. Microbiome 2019, 7, 9. [Google Scholar] [CrossRef]
- Chen, J.Z.; Thomsen, M.; Vitetta, L. Interaction of gut microbiota with dysregulation of bile acids in the pathogenesis of nonalcoholic fatty liver disease and potential therapeutic implications of probiotics. J. Cell. Biochem. 2019, 120, 2713–2720. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.L.; Luo, P.; Liu, J.; Wang, J.J.; Chen, Y. Dihydromyricetin ameliorated nonalcoholic steatohepatitis in mice by regulating the composition of serous lipids, bile acids and ileal microflora. Lipids Health Dis. 2023, 22, 112. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Z.; Vitetta, L. The Role of Butyrate in Attenuating Pathobiont-Induced Hyperinflammation. Immune Netw. 2020, 20, 15. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Unit | Value | Parameters | Unit | Value |
|---|---|---|---|---|---|
| AUC(0-t) | ng/L*min | 30,236 | Zeta | 1/min | 0.003 |
| AUC(0-∞) | ng/L*min | 34,864 | Zeta | 134 | |
| AUMC(0-t) | 7,351,242 | Cz | ng/L | 15 | |
| AUMC(0-∞) | 12,023,223 | T1/2 | min | 204 | |
| MRT(0-t) | min | 243 | Tmax | min | 120 |
| MRT(0-∞) | min | 344 | VZ/F | L/kg | 177,980 |
| VRT(0-t) | min2 | 31,533 | CLZ/F | L/min/kg | 602 |
| VRT(0-∞) | min2 | 107,286 | Cmax | ng/L | 113 |
| Parameters | Unit | Value | Parameters | Unit | Value |
|---|---|---|---|---|---|
| AUC(0-t) | ng/L*min | 108,480 | Zeta | 1/min | 0.004 |
| AUC(0-∞) | ng/L*min | 136,923 | Zeta | 123 | |
| AUMC(0-t) | 11,816,712 | Cz | ng/L | 107 | |
| AUMC(0-∞) | 29,781,860 | T1/2 | min | 182 | |
| MRT(0-t) | min | 108 | Tmax | min | 5 |
| MRT(0-∞) | min | 217 | Vz | L/kg | 40,444 |
| VRT(0-t) | min2 | 10,194 | CLZ | L/min/kg | 153 |
| VRT(0-∞) | min2 | 66,630 | Cmax | ng/L | 1092 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Yu, J.; Bai, S.; Li, S.; Qiu, Q.; Kong, X.; Xiang, C.; Liu, Z.; Yu, P.; Teng, Y. Compound 3d Attenuates Metabolic Dysfunction-Associated Steatohepatitis via Peroxisome Proliferator-Activated Receptor Pathway Activation and Inhibition of Inflammatory and Apoptotic Signaling. Metabolites 2025, 15, 296. https://doi.org/10.3390/metabo15050296
Zhang S, Yu J, Bai S, Li S, Qiu Q, Kong X, Xiang C, Liu Z, Yu P, Teng Y. Compound 3d Attenuates Metabolic Dysfunction-Associated Steatohepatitis via Peroxisome Proliferator-Activated Receptor Pathway Activation and Inhibition of Inflammatory and Apoptotic Signaling. Metabolites. 2025; 15(5):296. https://doi.org/10.3390/metabo15050296
Chicago/Turabian StyleZhang, Shouqing, Jiajia Yu, Sule Bai, Shuhan Li, Quanyuan Qiu, Xiangshun Kong, Cen Xiang, Zhen Liu, Peng Yu, and Yuou Teng. 2025. "Compound 3d Attenuates Metabolic Dysfunction-Associated Steatohepatitis via Peroxisome Proliferator-Activated Receptor Pathway Activation and Inhibition of Inflammatory and Apoptotic Signaling" Metabolites 15, no. 5: 296. https://doi.org/10.3390/metabo15050296
APA StyleZhang, S., Yu, J., Bai, S., Li, S., Qiu, Q., Kong, X., Xiang, C., Liu, Z., Yu, P., & Teng, Y. (2025). Compound 3d Attenuates Metabolic Dysfunction-Associated Steatohepatitis via Peroxisome Proliferator-Activated Receptor Pathway Activation and Inhibition of Inflammatory and Apoptotic Signaling. Metabolites, 15(5), 296. https://doi.org/10.3390/metabo15050296