

Effects of Long-Term Airport Noise Exposure on Inflammation and Intestinal Flora and Their Metabolites in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Grouping and Airport Noise Exposure
2.2. Noninvasive Blood Pressure Test
2.3. Effect of Noise Exposure on Inflammatory Factor Levels
2.4. Targeted Metabolomics Analysis
2.5. GC-MS/MSanalysis
2.6. Effect of Noise Exposure on LPS, LTA, TMA, and TMAO Levels
2.7. Sequencing and Analysis of 16s rDNA of Intestinal Flora
2.8. Preparation of Graphics and Statistical Analysis
3. Results
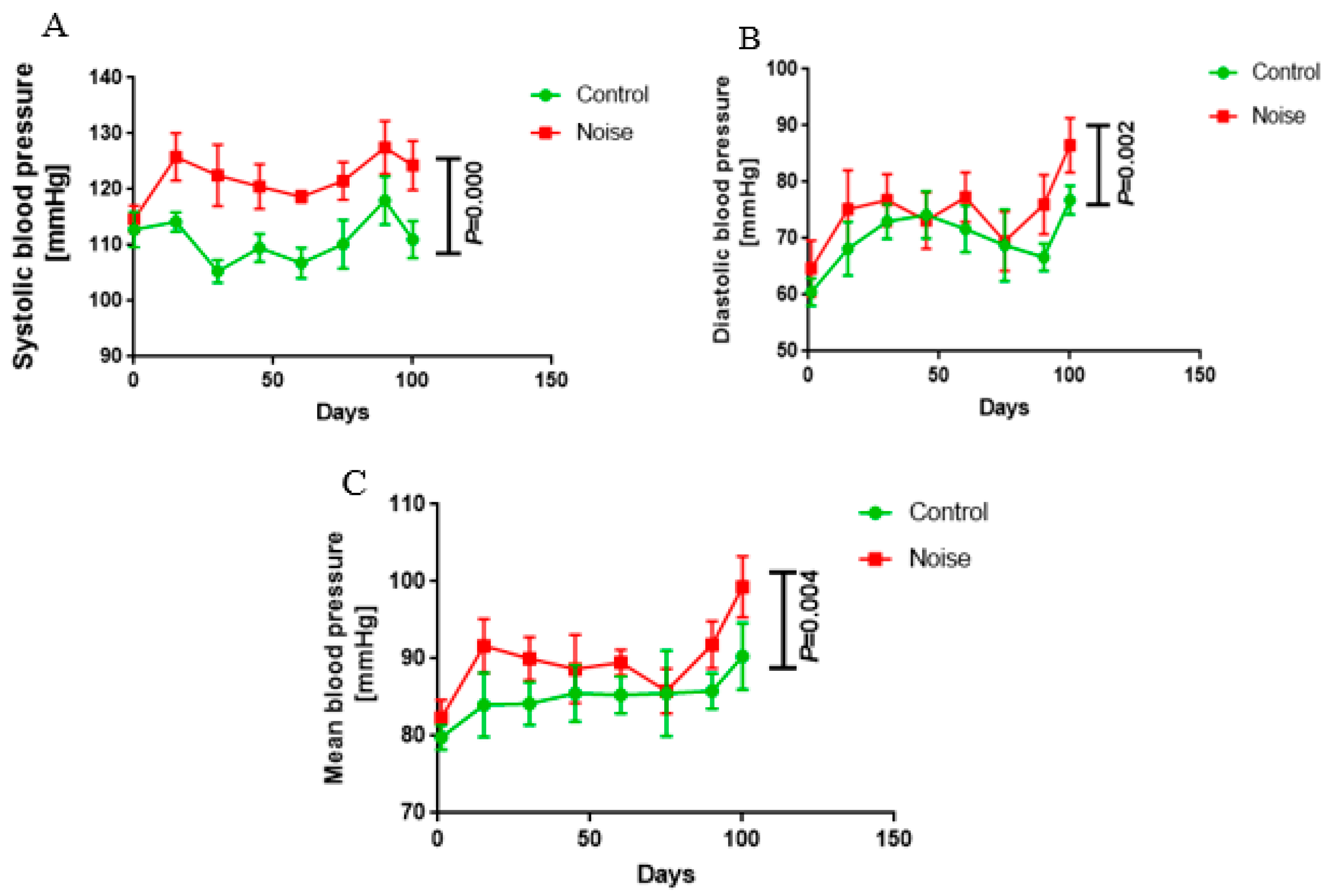
3.1. Effect of Chronic Airport Noise on Blood Pressure in Mice
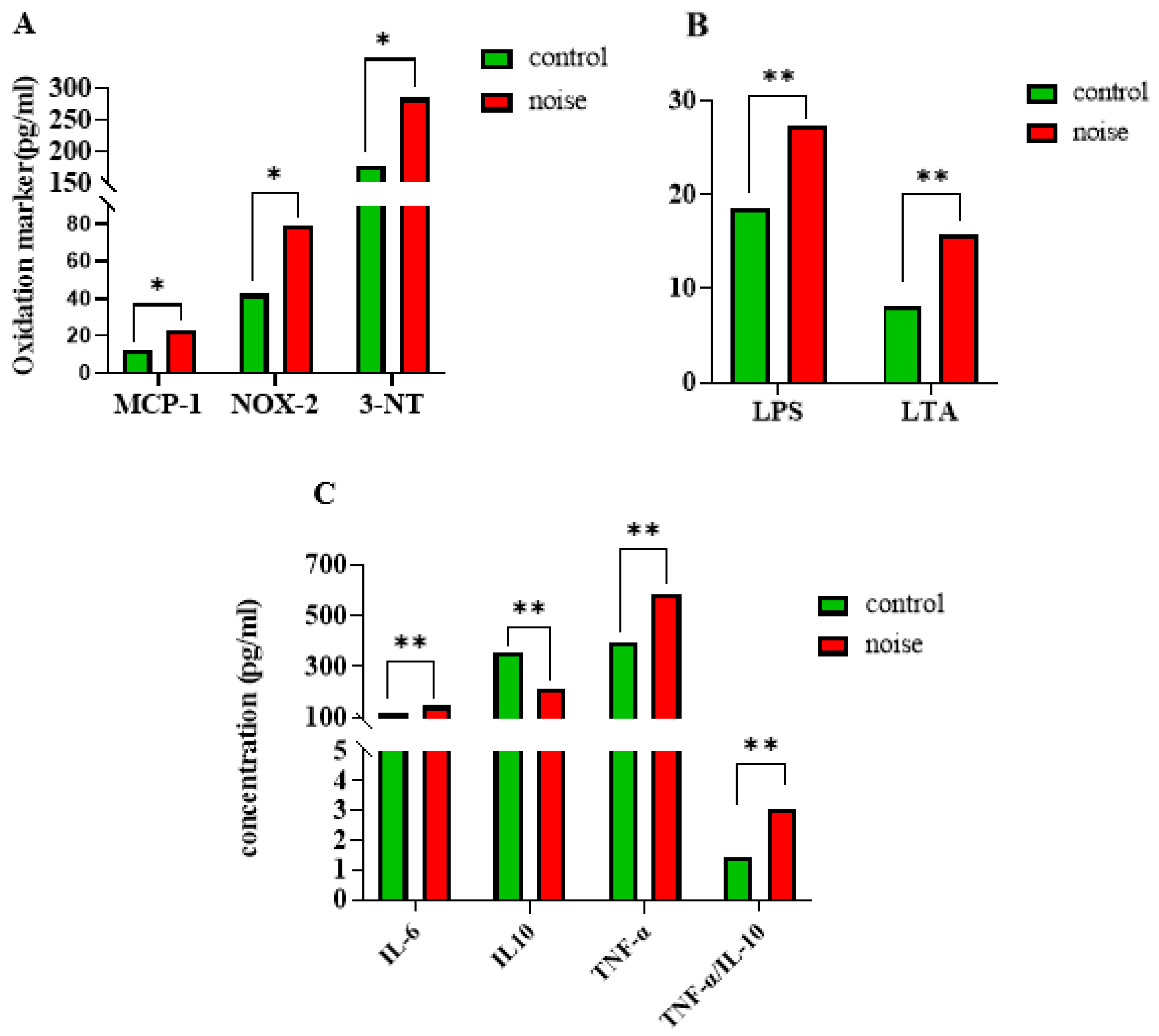
3.2. Changes in Oxidative Stress, Inflammatory Markers, and LPS and LTA Levels in Mice After Chronic Airport Noise Exposure
3.3. Effects of Chronic Airport Noise Exposure on Colon Tissue Structure Changes in Mice
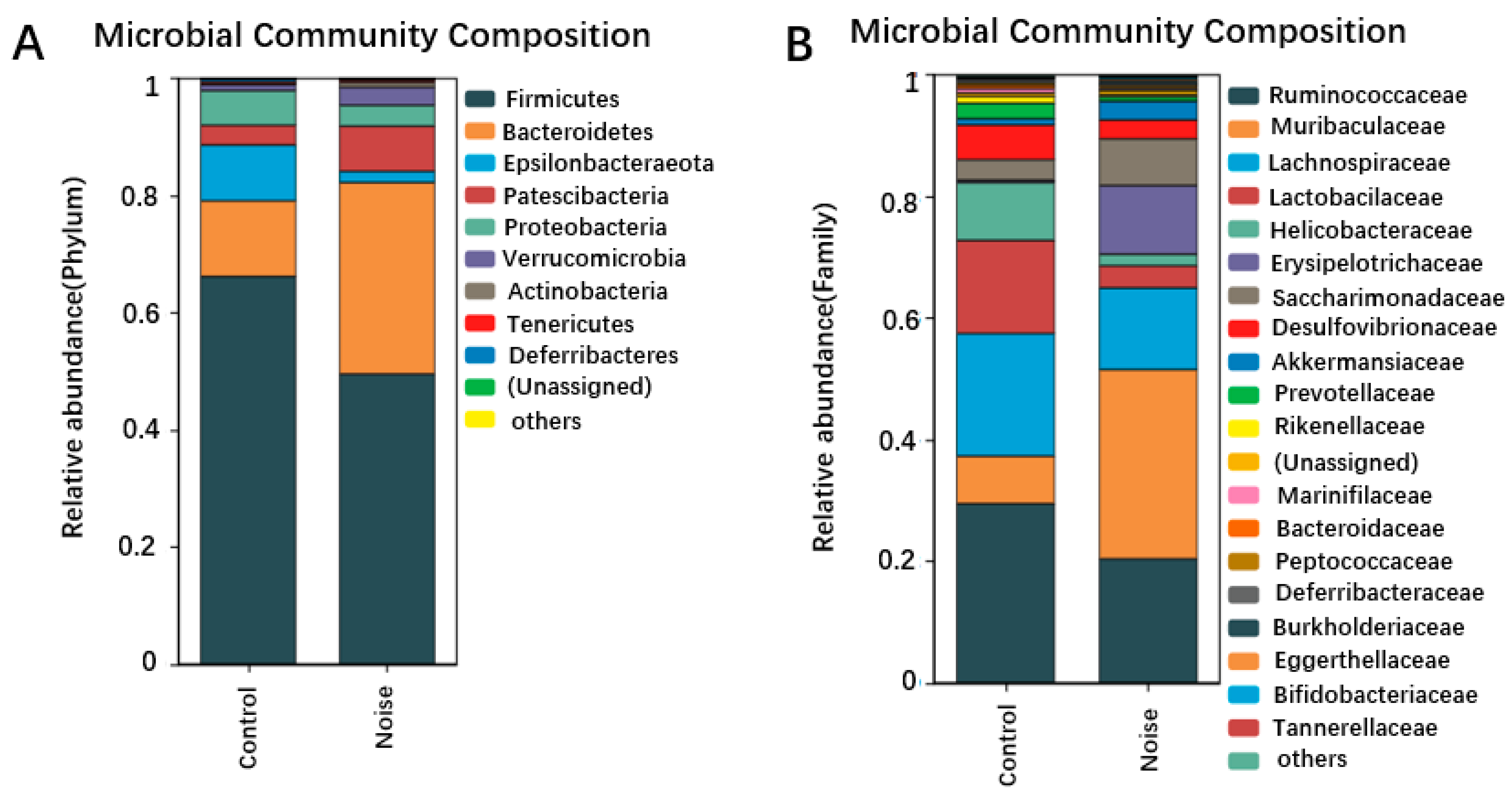
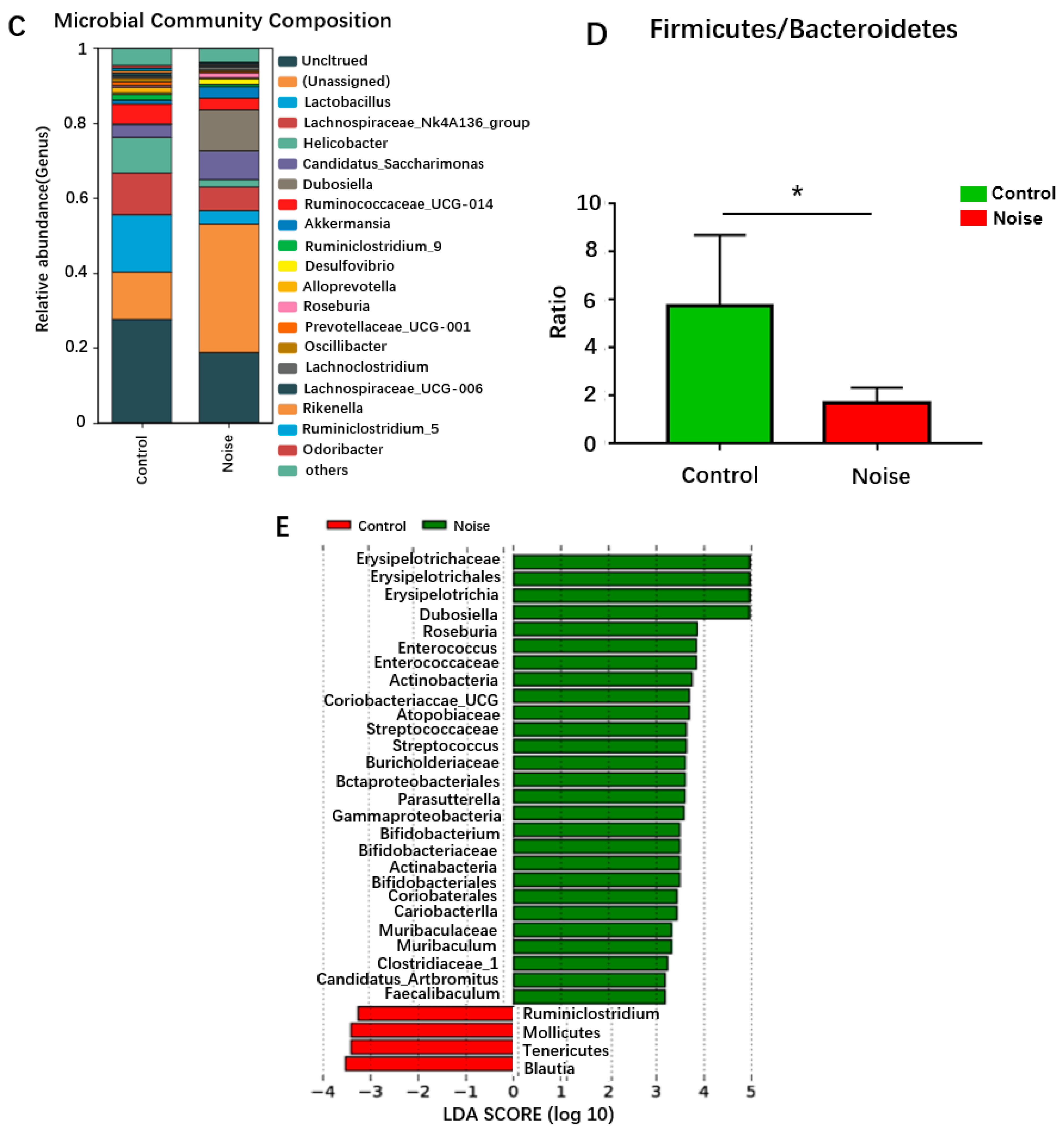
3.4. Effects of Chronic Airport Noise Exposure on Intestinal Flora Structure Changes in Mice
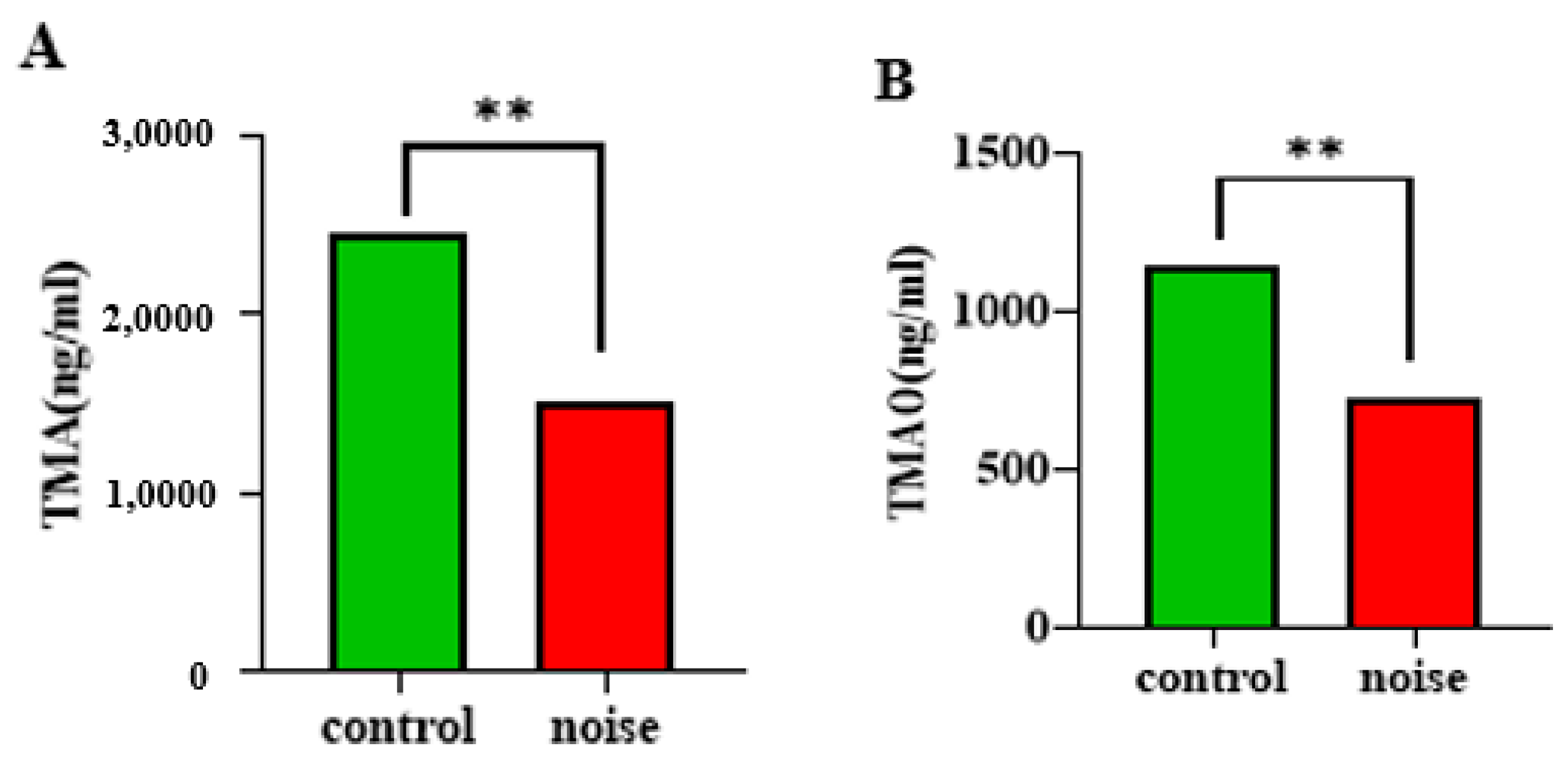
3.5. Effects of Chronic Airport Noise Exposure on Levels of Trimethylamine and Trimethylamine Oxide
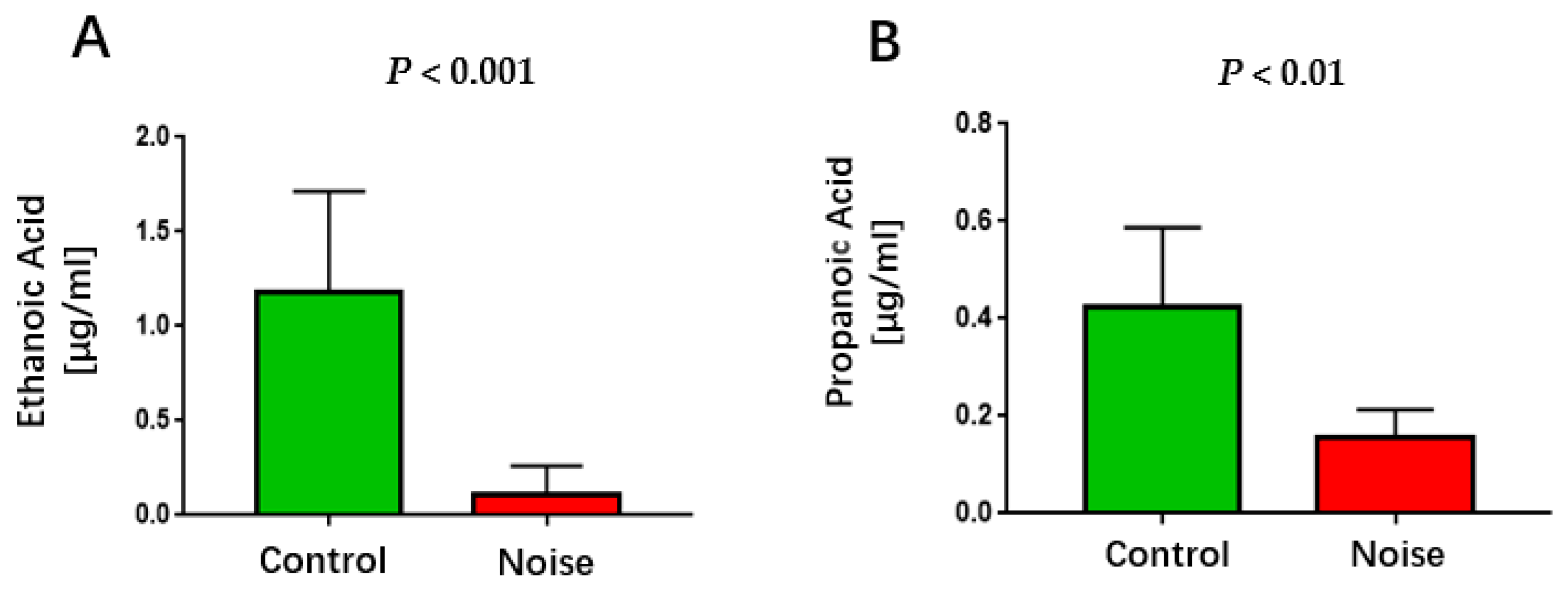
3.6. Effect of Chronic Airport Noise Exposure on Serum Lower Carboxylic Acid Levels
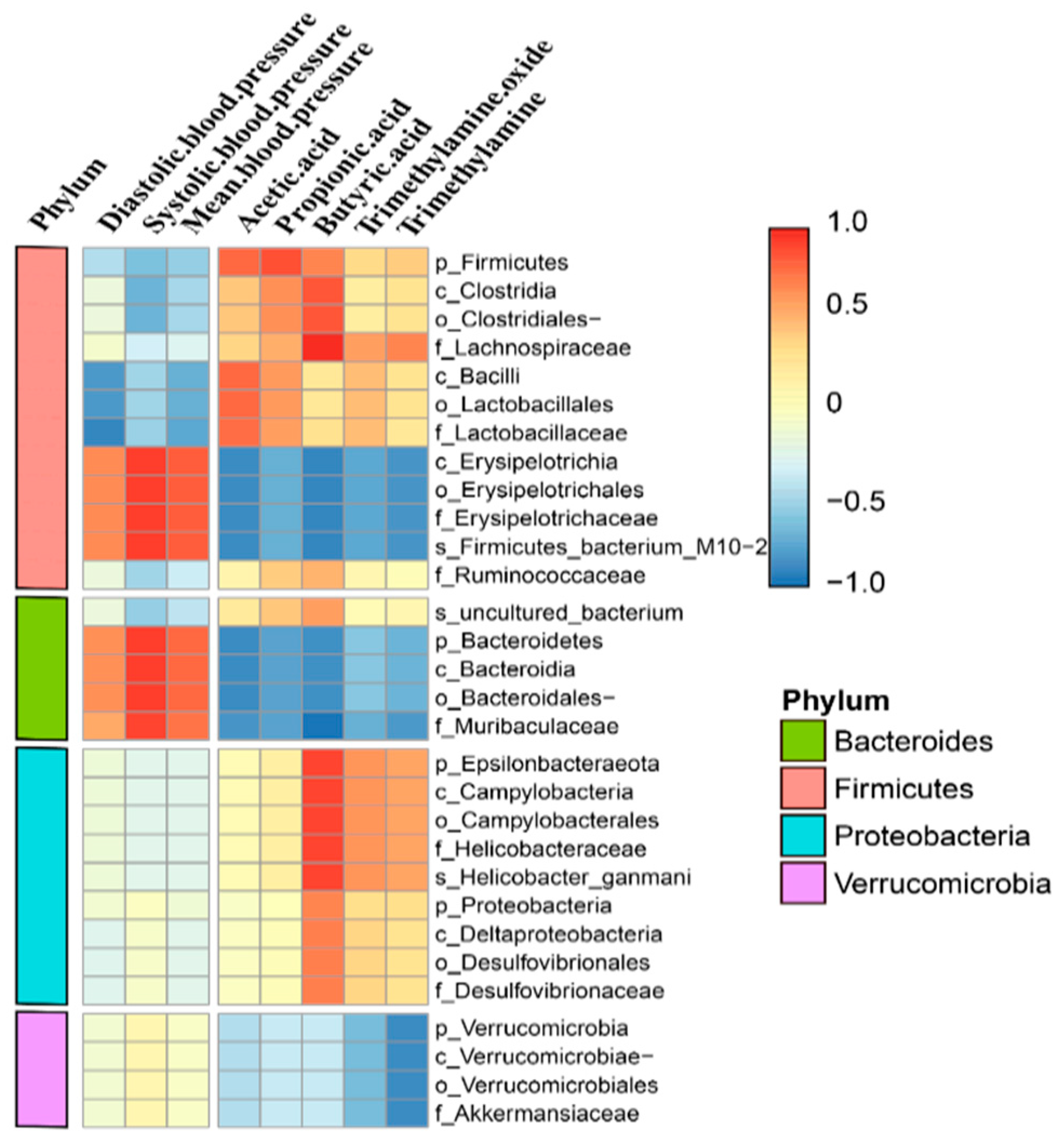
3.7. Correlation Among Microbial Composition, Blood Pressure, and Intestinal Metabolites in Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hänninen, O.; Knol, A.B.; Jantunen, M.; Lim, T.A.; Conrad, A.; Rappolder, M.; Carrer, P.; Fanetti, A.C.; Kim, R.; Buekers, J.; et al. Environmental burden of disease in Europe: Assessing nine risk factors in six countries. Env. Health Perspect. 2014, 122, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Pyko, A.; Roswall, N.; Ögren, M.; Oudin, A.; Rosengren, A.; Eriksson, C.; Segersson, D.; Rizzuto, D.; Andersson, E.M.; Aasvang, G.M.; et al. Long-term exposure to transportation noise and ischemic heart disease: A pooled analysis of nine scandinavian cohorts. Env. Health Perspect. 2023, 131, 17003. [Google Scholar] [CrossRef]
- Thacher, J.D.; Poulsen, A.H.; Raaschou-Nielsen, O.; Hvidtfeldt, U.A.; Brandt, J.; Christensen, J.H.; Khan, J.; Levin, G.; Münzel, T.; Sørensen, M. Exposure to transportation noise and risk for cardiovascular disease in a nationwide cohort study from Denmark. Env. Res. 2022, 211, 113106. [Google Scholar] [CrossRef]
- Ba̧czalska, J.; Wojciechowska, W.; Rojek, M.; Hahad, O.; Daiber, A.; Münzel, T.; Rajzer, M. Cardiovascular consequences of aircraft noise exposure. Front. Public. Health 2022, 10, 1058423. [Google Scholar] [CrossRef]
- Saucy, A.; Schäffer, B.; Tangermann, L.; Vienneau, D.; Wunderli, J.M.; Röösli, M. Does night-time aircraft noise trigger mortality? A case-crossover study on 24 886 cardiovascular deaths. Eur. Heart J. 2021, 42, 835–843. [Google Scholar] [CrossRef]
- Basner, M.; Witte, M.; McGuire, S. Aircraft noise effects on sleep-results of a pilot study near philadelphia international airport. Int. J. Env. Res. Public. Health 2019, 16, 3178. [Google Scholar] [CrossRef]
- Eckrich, J.; Frenis, K.; Rodriguez-Blanco, G.; Ruan, Y.; Jiang, S.; Bayo Jimenez, M.T.; Kuntic, M.; Oelze, M.; Hahad, O.; Li, H.; et al. Aircraft noise exposure drives the activation of white blood cells and induces microvascular dysfunction in mice. Redox Biol. 2021, 46, 102063. [Google Scholar] [CrossRef]
- Kröller-Schön, S.; Daiber, A.; Steven, S.; Oelze, M.; Frenis, K.; Kalinovic, S.; Heimann, A.; Schmidt, F.P.; Pinto, A.; Kvandova, M.; et al. Crucial role for Nox2 and sleep deprivation in aircraft noise-induced vascular and cerebral oxidative stress, inflammation, and gene regulation. Eur. Heart J. 2018, 39, 3528–3539. [Google Scholar] [CrossRef]
- Huang, D.; Song, X.; Cui, Q.; Tian, J.; Wang, Q.; Yang, K. Is there an association between aircraft noise exposure and the incidence of hypertension? A meta-analysis of 16784 participants. Noise Health 2015, 17, 93–97. [Google Scholar] [CrossRef]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar]
- Kim, S.; Goel, R.; Kumar, A.; Qi, Y.; Lobaton, G.; Hosaka, K.; Mohammed, M.; Handberg, E.M.; Richards, E.M.; Pepine, C.J.; et al. Imbalance of gut microbiome and intestinal epithelial barrier dysfunction in patients with high blood pressure. Clin. Sci. 2018, 132, 701–718. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Gu, Y.; Li, X.; Yang, W.; Jia, L.; Chen, C.; Han, X.; Huang, Y.; Zhao, L.; Li, P.; et al. Alterations of the gut microbiome in hypertension. Front. Cell Infect. Microbiol. 2017, 7, 381. [Google Scholar] [CrossRef]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut dysbiosis is linked to hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef]
- Bartolomaeus, H.; Balogh, A.; Yakoub, M.; Homann, S.; Markó, L.; Höges, S.; Tsvetkov, D.; Krannich, A.; Wundersitz, S.; Avery, E.G.; et al. Short-chain fatty acid propionate protects from hypertensive cardiovascular damage. Circulation 2019, 139, 1407–1421. [Google Scholar] [CrossRef]
- Sun, S.; Lulla, A.; Sioda, M.; Winglee, K.; Wu, M.C.; Jacobs, D.R., Jr.; Shikany, J.M.; Lloyd-Jones, D.M.; Launer, L.J.; Fodor, A.A.; et al. Gut microbiota composition and blood pressure. Hypertension 2019, 73, 998–1006. [Google Scholar] [CrossRef]
- Cui, B.; Gai, Z.; She, X.; Wang, R.; Xi, Z. Effects of chronic noise on glucose metabolism and gut microbiota-host inflammatory homeostasis in rats. Sci. Rep. 2016, 6, 36693. [Google Scholar] [CrossRef] [PubMed]
- Hoyles, L.; Pontifex, M.G.; Rodriguez-Ramiro, I.; Anis-Alavi, M.A.; Jelane, K.S.; Snelling, T.; Solito, E.; Fonseca, S.; Carvalho, A.L.; Carding, S.R.; et al. Regulation of blood-brain barrier integrity by microbiome-associated methylamines and cognition by trimethylamine N-oxide. Microbiome 2021, 9, 235. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Massy, Z.A.; Stenvinkel, P.; Chesnaye, N.C.; Larabi, I.A.; Alvarez, J.C.; Caskey, F.J.; Torino, C.; Porto, G.; Szymczak, M.; et al. The association between TMAO, CMPF, and clinical outcomes in advanced chronic kidney disease: Results from the European quality (EQUAL) study. Am. J. Clin. Nutr. 2022, 116, 1842–1851. [Google Scholar] [CrossRef]
- Bjørnestad, E.; Dhar, I.; Svingen, G.F.T.; Pedersen, E.R.; Ørn, S.; Svenningsson, M.M.; Tell, G.S.; Ueland, P.M.; Sulo, G.; Laaksonen, R.; et al. Circulating trimethylamine N-oxide levels do not predict 10-year survival in patients with or without coronary heart disease. J. Intern. Med. 2022, 292, 915–924. [Google Scholar] [CrossRef]
- Svingen, G.F.T.; Zuo, H.; Ueland, P.M.; Seifert, R.; Løland, K.H.; Pedersen, E.R.; Schuster, P.M.; Karlsson, T.; Tell, G.S.; Schartum-Hansen, H.; et al. Increased plasma trimethylamine-N-oxide is associated with incident atrial fibrillation. Int. J. Cardiol. 2018, 267, 100–106. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, S.H.; Chen, Q.C.; Liao, X.M. Moderate noise induced cognition impairment of mice and its underlying mechanisms. Physiol. Behav. 2011, 104, 981–988. [Google Scholar] [CrossRef]
- Cui, B.; Li, K.; Gai, Z.; She, X.; Zhang, N.; Xu, C.; Chen, X.; An, G.; Ma, Q.; Wang, R. Chronic noise exposure acts cumulatively to exacerbate alzheimer’s disease-like amyloid-β pathology and neuroinflammation in the rat hippocampus. Sci. Rep. 2015, 5, 12943. [Google Scholar] [CrossRef]
- Jafari, Z.; Kolb, B.E.; Mohajerani, M.H. Chronic traffic noise stress accelerates brain impairment and cognitive decline in mice. Exp. Neurol. 2018, 308, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mac Domhnaill, C.; Douglas, O.; Lyons, S.; Murphy, E.; Nolan, A. Road traffic noise and cognitive function in older adults: A cross-sectional investigation of the irish longitudinal study on ageing. BMC Public. Health 2021, 21, 1814. [Google Scholar]
- Wang, S.; Yu, Y.; Feng, Y.; Zou, F.; Zhang, X.; Huang, J.; Zhang, Y.; Zheng, X.; Huang, X.F.; Zhu, Y.; et al. Protective effect of the orientin on noise-induced cognitive impairments in mice. Behav. Brain Res. 2016, 296, 290–300. [Google Scholar] [CrossRef]
- Molitor, M.; Bayo-Jimenez, M.T.; Hahad, O.; Witzler, C.; Finger, S.; Garlapati, V.S.; Rajlic, S.; Knopp, T.; Bieler, T.K.; Aluia, M.; et al. Aircraft noise exposure induces pro-inflammatory vascular conditioning and amplifies vascular dysfunction and impairment of cardiac function after myocardial infarction. Cardiovasc. Res. 2023, 119, 1416–1426. [Google Scholar] [CrossRef] [PubMed]
- Steven, S.; Frenis, K.; Kalinovic, S.; Kvandova, M.; Oelze, M.; Helmstädter, J.; Hahad, O.; Filippou, K.; Kus, K.; Trevisan, C.; et al. Exacerbation of adverse cardiovascular effects of aircraft noise in an animal model of arterial hypertension. Redox Biol. 2020, 34, 101515. [Google Scholar] [CrossRef]
- Kvandová, M.; Rajlic, S.; Stamm, P.; Schmal, I.; Mihaliková, D.; Kuntic, M.; Bayo Jimenez, M.T.; Hahad, O.; Kollárová, M.; Ubbens, H.; et al. Mitigation of aircraft noise-induced vascular dysfunction and oxidative stress by exercise, fasting, and pharmacological α1AMPK activation: Molecular proof of a protective key role of endothelial α1AMPK against environmental noise exposure. Eur. J. Prev. Cardiol. 2023, 30, 1554–1568. [Google Scholar] [CrossRef]
- Münzel, T.; Sørensen, M.; Daiber, A. Transportation noise pollution and cardiovascular disease. Nat. Rev. Cardiol. 2021, 18, 619–636. [Google Scholar] [CrossRef]
- Münzel, T.; Kröller-Schön, S.; Oelze, M.; Gori, T.; Schmidt, F.P.; Steven, S.; Hahad, O.; Röösli, M.; Wunderli, J.M.; Daiber, A.; et al. Adverse cardiovascular effects of traffic noise with a focus on nighttime noise and the new who noise guidelines. Annu. Rev. Public. Health 2020, 41, 309–328. [Google Scholar] [CrossRef]
- Thiesse, L.; Rudzik, F.; Spiegel, K.; Leproult, R.; Pieren, R.; Wunderli, J.M.; Foraster, M.; Héritier, H.; Eze, I.C.; Meyer, M.; et al. Adverse impact of nocturnal transportation noise on glucose regulation in healthy young adults: Effect of different noise scenarios. Environ. Int. 2018, 121, 1011–1023. [Google Scholar] [CrossRef] [PubMed]
- Münzel, T.; Daiber, A.; Steven, S.; Tran, L.P.; Ullmann, E.; Kossmann, S.; Schmidt, F.P.; Oelze, M.; Xia, N.; Li, H.; et al. Effects of noise on vascular function, oxidative stress, and inflammation: Mechanistic insight from studies in mice. Eur. Heart J. 2017, 38, 2838–2849. [Google Scholar] [CrossRef] [PubMed]
- Kempen, E.V.; Casas, M.; Pershagen, G.; Foraster, M. WHO environmental noise guidelines for the european region: A systematic review on environmental noise and cardiovascular and metabolic effects: A summary. Int. J. Environ. Res. Public. Health 2018, 15, 379. [Google Scholar] [CrossRef]
- Galla, S.; Chakraborty, S.; Mell, B.; Vijay-Kumar, M.; Joe, B. Microbiotal-host interactions and hypertension. Physiology 2017, 32, 224–233. [Google Scholar] [CrossRef]
- Kaye, D.M.; Shihata, W.A.; Jama, H.A.; Tsyganov, K.; Ziemann, M.; Kiriazis, H.; Horlock, D.; Vijay, A.; Giam, B.; Vinh, A.; et al. Deficiency of prebiotic fiber and insufficient signaling through gut metabolite-sensing receptors leads to cardiovascular disease. Circulation 2020, 141, 1393–1403. [Google Scholar] [CrossRef]
- Karbach, S.H.; Schönfelder, T.; Brandão, I.; Wilms, E.; Hörmann, N.; Jäckel, S.; Schüler, R.; Finger, S.; Knorr, M.; Lagrange, J.; et al. Gut microbiota promote angiotensin II-induced arterial hypertension and vascular dysfunction. J. Am. Heart Assoc. 2016, 5, e003698. [Google Scholar] [CrossRef] [PubMed]
- He, X.; McLean, J.S.; Edlund, A.; Yooseph, S.; Hall, A.P.; Liu, S.Y.; Dorrestein, P.C.; Esquenazi, E.; Hunter, R.C.; Cheng, G.; et al. Cultivation of a human-associated TM7 phylotype reveals a reduced genome and epibiotic parasitic lifestyle. Proc. Natl. Acad. Sci. USA 2015, 112, 244–249. [Google Scholar] [CrossRef]
- Chen, W.; Liu, F.; Ling, Z.; Tong, X.; Xiang, C. Human intestinal lumen and mucosa-associated microbiota in patients with colorectal cancer. PLoS ONE 2012, 7, e39743. [Google Scholar] [CrossRef]
- Dinh, D.M.; Volpe, G.E.; Duffalo, C.; Bhalchandra, S.; Tai, A.K.; Kane, A.V.; Wanke, C.A.; Ward, H.D. Intestinal microbiota, microbial translocation, and systemic inflammation in chronic HIV infection. J. Infect. Dis. 2015, 211, 19–27. [Google Scholar] [CrossRef]
- Schaubeck, M.; Clavel, T.; Calasan, J.; Lagkouvardos, I.; Haange, S.B.; Jehmlich, N.; Basic, M.; Dupont, A.; Hornef, M.; von Bergen, M.; et al. Dysbiotic gut microbiota causes transmissible Crohn’s disease-like ileitis independent of failure in antimicrobial defence. Gut 2016, 65, 225–237. [Google Scholar] [CrossRef]
- Qiu, X.; Macchietto, M.G.; Liu, X.; Lu, Y.; Ma, Y.; Guo, H.; Saqui-Salces, M.; Bernlohr, D.A.; Chen, C.; Shen, S.; et al. Identification of gut microbiota and microbial metabolites regulated by an antimicrobial peptide lipocalin 2 in high fat diet-induced obesity. Int. J. Obes. 2021, 45, 143–154. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Warrier, M. Trimethylamine N-oxide, the microbiome, and heart and kidney disease. Annu. Rev. Nutr. 2017, 37, 157–181. [Google Scholar] [CrossRef]
- Bu, J.; Wang, Z. Cross–talk between gut microbiota and heart via the routes of metabolite and immunity. Gastroenterol. Res. Pr. 2018, 2018, 6458094. [Google Scholar] [CrossRef]
- Hsu, C.N.; Lu, P.C.; Hou, C.Y.; Tain, Y.L. Blood pressure abnormalities associated with gut microbiota-derived short chain fatty acids in children with congenital anomalies of the kidney and urinary tract. J. Clin. Med. 2019, 8, 1090. [Google Scholar] [CrossRef]
- Li, N.; Dai, Z.; Wang, Z.; Deng, Z.; Zhang, J.; Pu, J.; Cao, W.; Pan, T.; Zhou, Y.; Yang, Z.; et al. Gut microbiota dysbiosis contributes to the development of chronic obstructive pulmonary disease. Respir. Res. 2021, 22, 274. [Google Scholar]
- Zhang, W.Q.; Wang, Y.J.; Zhang, A.; Ding, Y.J.; Zhang, X.N.; Jia, Q.J.; Zhu, Y.P.; Li, Y.Y.; Lv, S.C.; Zhang, J.P. TMA/TMAO in hypertension: Novel horizons and potential therapies. J. Cardiovasc. Transl. Res. 2021, 14, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Jiang, H.; Zhuo, C.; Jiang, W. Trimethylamine/trimethylamine-N-oxide as a key between diet and cardiovascular diseases. Cardiovasc. Toxicol. 2021, 21, 593–604. [Google Scholar] [CrossRef]
- Xu, J.; Cheng, A.; Song, B.; Zhao, M.; Xue, J.; Wang, A.; Dai, L.; Jing, J.; Meng, X.; Li, H.; et al. Trimethylamine N-oxide and stroke recurrence depends on ischemic stroke subtypes. Stroke 2022, 53, 1207–1215. [Google Scholar] [CrossRef]
- Tang, W.H.W.; Lemaitre, R.N.; Jensen, P.N.; Wang, M.; Wang, Z.; Li, X.S.; Nemet, I.; Lee, Y.; Lai, H.T.M.; de Oliveira Otto, M.C.; et al. Trimethylamine N-oxide and related gut microbe-derived metabolites and incident heart failure development in community-based populations. Circ. Heart Fail. 2024, 17, e011569. [Google Scholar] [CrossRef]
- Nogal, A.; Valdes, A.M.; Menni, C. The role of short-chain fatty acids in the interplay between gut microbiota and diet in cardio-metabolic health. Gut Microbes 2021, 13, 1–24. [Google Scholar] [CrossRef]
- Birkeland, E.; Gharagozlian, S.; Valeur, J.; Aas, A.M. Short-chain fatty acids as a link between diet and cardiometabolic risk: A narrative review. Lipids Health Dis. 2023, 22, 40. [Google Scholar] [PubMed]
- Salamone, D.; Rivellese, A.A.; Vetrani, C. The relationship between gut microbiota, short-chain fatty acids and type 2 diabetes mellitus: The possible role of dietary fibre. Acta Diabetol. 2021, 58, 1131–1138. [Google Scholar] [CrossRef]
- He, J.; Zhang, P.; Shen, L.; Niu, L.; Tan, Y.; Chen, L.; Zhao, Y.; Bai, L.; Hao, X.; Li, X.; et al. Short-chain fatty acids and their association with signalling pathways in inflammation, glucose and lipid metabolism. Int. J. Mol. Sci. 2020, 21, 6356. [Google Scholar] [CrossRef]
- Campos-Perez, W.; Martinez-Lopez, E. Effects of short chain fatty acids on metabolic and inflammatory processes in human health. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 158900. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, M.; Wang, H.; Li, N.; Lu, Z.; Li, L.; Hui, S.; Xu, H. Gut microbiota and short chain fatty acids partially mediate the beneficial effects of inulin on metabolic disorders in obese ob/ob mice. J. Food Biochem. 2022, 46, e14063. [Google Scholar] [CrossRef]
- Huart, J.; Leenders, J.; Taminiau, B.; Descy, J.; Saint-Remy, A.; Daube, G.; Krzesinski, J.M.; Melin, P.; de Tullio, P.; Jouret, F. Gut microbiota and fecal levels of short-chain fatty acids differ upon 24-hour blood pressure levels in men. Hypertension 2019, 74, 1005–1013. [Google Scholar] [CrossRef]
- Chen, L.; He, F.J.; Dong, Y.; Huang, Y.; Wang, C.; Harshfield, G.A.; Zhu, H. Modest sodium reduction increases circulating short-chain fatty acids in untreated hypertensives: A randomized, double-blind, placebo-controlled trial. Hypertension 2020, 76, 73–79. [Google Scholar] [CrossRef]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef]
- Robles-Vera, I.; Toral, M.; de la Visitación, N.; Sánchez, M.; Gómez-Guzmán, M.; Romero, M.; Yang, T.; Izquierdo-Garcia, J.L.; Jiménez, R.; Ruiz-Cabello, J.; et al. Probiotics prevent dysbiosis and the rise in blood pressure in genetic hypertension: Role of short-chain fatty acids. Mol. Nutr. Food Res. 2020, 64, e1900616. [Google Scholar] [CrossRef]
- Vinolo, M.A.; Rodrigues, H.G.; Hatanaka, E.; Sato, F.T.; Sampaio, S.C.; Curi, R. Suppressive effect of short-chain fatty acids on production of proinflammatory mediators by neutrophils. J. Nutr. Biochem. 2011, 22, 849–855. [Google Scholar] [CrossRef]
- Cai, L.; Zhan, M.; Li, Q.; Li, D.; Xu, Q. DNA methyltransferase DNMT1 inhibits lipopolysaccharide-induced inflammatory response in human dental pulp cells involving the methylation changes of IL-6 and TRAF6. Mol. Med. Rep. 2020, 21, 959–968. [Google Scholar] [CrossRef]
- Li, T.; Garcia-Gomez, A.; Morante-Palacios, O.; Ciudad, L.; Özkaramehmet, S.; Van Dijck, E.; Rodríguez-Ubreva, J.; Vaquero, A.; Ballestar, E. SIRT1/2 orchestrate acquisition of DNA methylation and loss of histone H3 activating marks to prevent premature activation of inflammatory genes in macrophages. Nucleic Acids Res. 2020, 48, 665–681. [Google Scholar] [CrossRef] [PubMed]
- Pierre, W.C.; Legault, L.M.; Londono, I.; McGraw, S.; Lodygensky, G.A. Alteration of the brain methylation landscape following postnatal inflammatory injury in rat pups. Faseb J. 2020, 34, 432–445. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wu, Y.; Sun, Y.; Dong, X.; Wang, Z.; Zhang, Z.; Xiao, Y.; Dong, G. Bacterial lipopolysaccharide induced alterations of genome-wide dna methylation and promoter methylation of lactation-related genes in bovine mammary epithelial cells. Toxins 2019, 11, 298. [Google Scholar] [CrossRef] [PubMed]
- Roswall, N.; Thacher, J.D.; Ögren, M.; Pyko, A.; Åkesson, A.; Oudin, A.; Tjønneland, A.; Rosengren, A.; Poulsen, A.H.; Eriksson, C.; et al. Long-term exposure to traffic noise and risk of incident colon cancer: A pooled study of eleven Nordic cohorts. Environ. Res. 2023, 224, 115454. [Google Scholar] [CrossRef]
- Cho, I.; Kim, J.; Jung, S.; Kim, S.Y.; Kim, E.J.; Choo, S.; Kam, E.H.; Koo, B.N. The impact of persistent noise exposure under inflammatory conditions. Healthcare 2023, 11, 2067. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Wei, L.; Xia, Y.; Wang, J.; Bai, Y.; Xia, Y. Effects of Long-Term Airport Noise Exposure on Inflammation and Intestinal Flora and Their Metabolites in Mice. Metabolites 2025, 15, 251. https://doi.org/10.3390/metabo15040251
Yang J, Wei L, Xia Y, Wang J, Bai Y, Xia Y. Effects of Long-Term Airport Noise Exposure on Inflammation and Intestinal Flora and Their Metabolites in Mice. Metabolites. 2025; 15(4):251. https://doi.org/10.3390/metabo15040251
Chicago/Turabian StyleYang, Jian, Longwei Wei, Yuan Xia, Junyi Wang, Yan Bai, and Yun Xia. 2025. "Effects of Long-Term Airport Noise Exposure on Inflammation and Intestinal Flora and Their Metabolites in Mice" Metabolites 15, no. 4: 251. https://doi.org/10.3390/metabo15040251
APA StyleYang, J., Wei, L., Xia, Y., Wang, J., Bai, Y., & Xia, Y. (2025). Effects of Long-Term Airport Noise Exposure on Inflammation and Intestinal Flora and Their Metabolites in Mice. Metabolites, 15(4), 251. https://doi.org/10.3390/metabo15040251