
Transcriptomic and Lipidomic Analysis Reveals Complex Regulation Mechanisms Underlying Rice Roots’ Response to Salt Stress

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Phenotypic Measurement
2.3. Analysis of Physiological and Biochemical Indices
2.4. RNA Sequencing and Analysis
2.5. RNA Extraction, Reverse Transcription, and Real-Time Quantitative PCR
2.6. Lipid Extraction and UHPLC-MS/MS Analysis
2.7. Data Analysis
3. Results
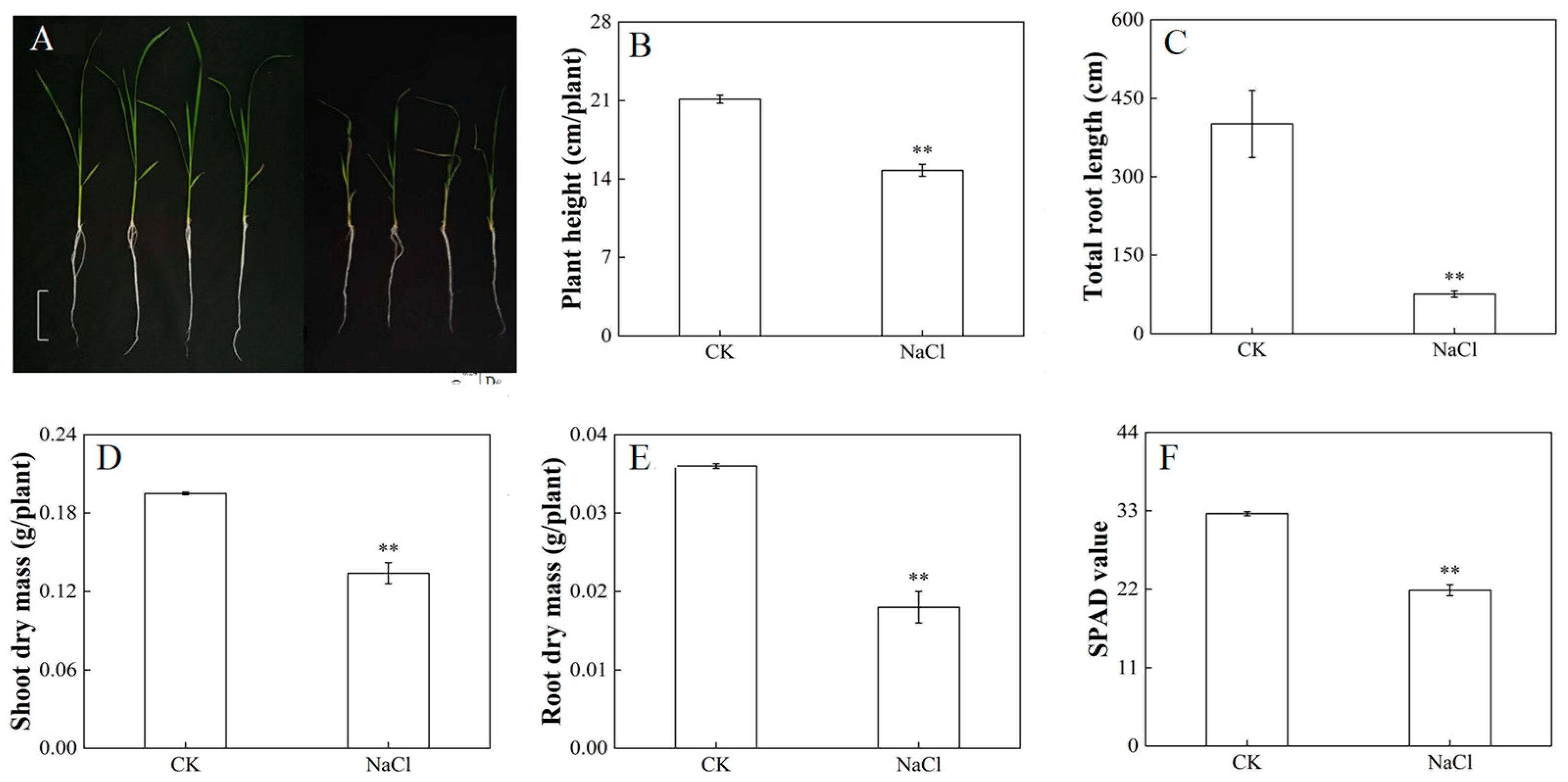
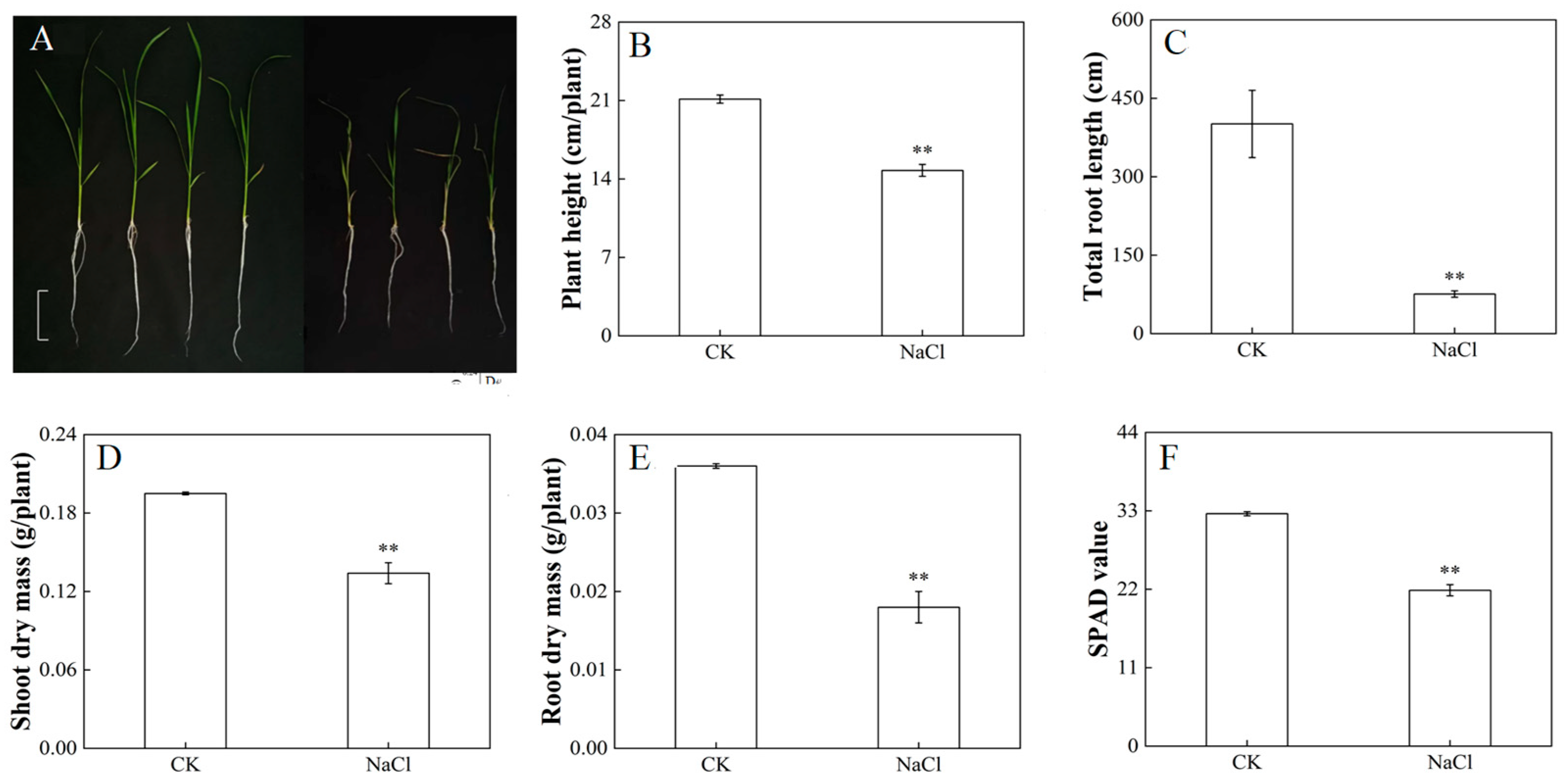
3.1. Response of Growth and Photosynthetic Properties of Rice under Salt Stress
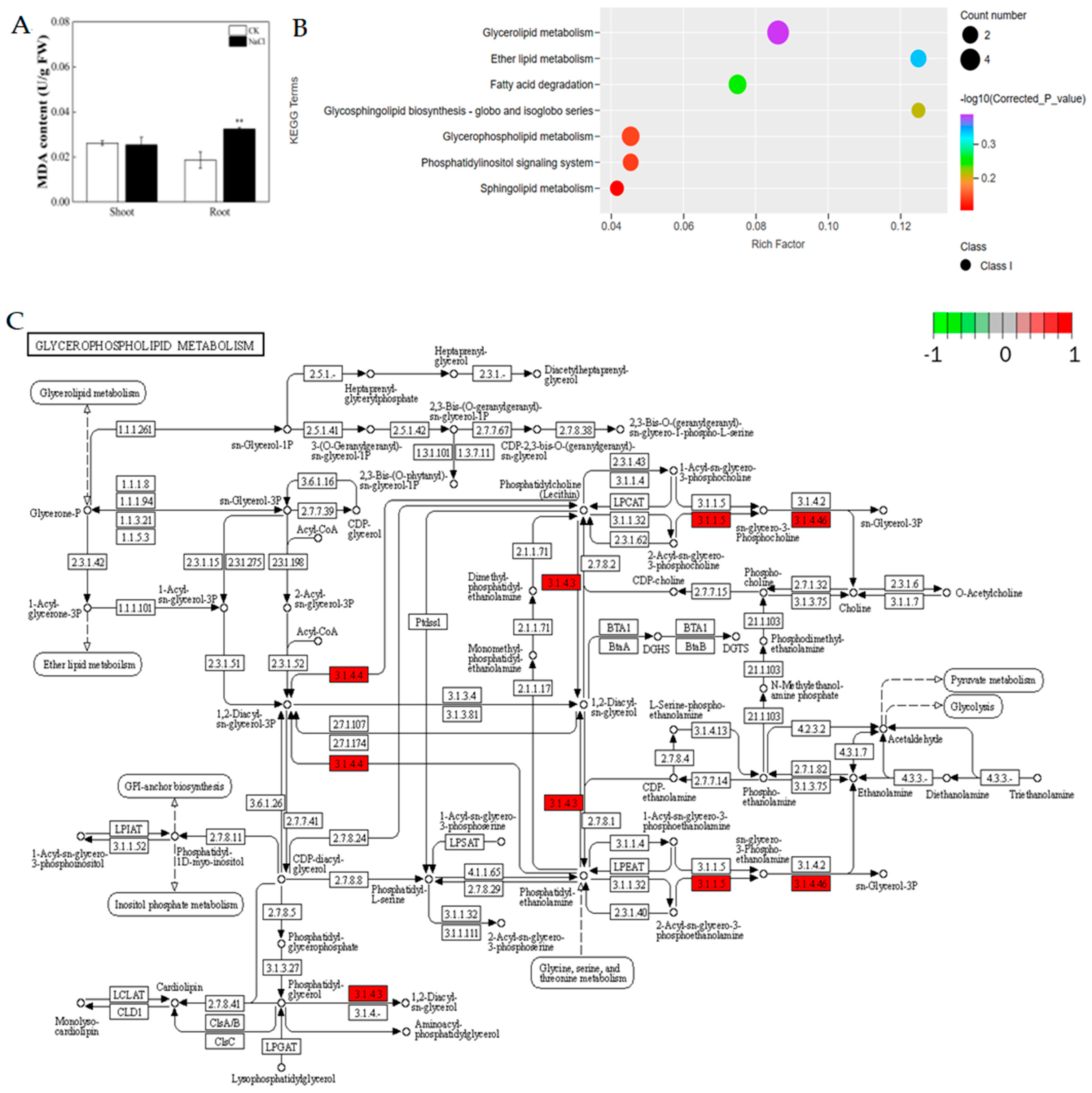
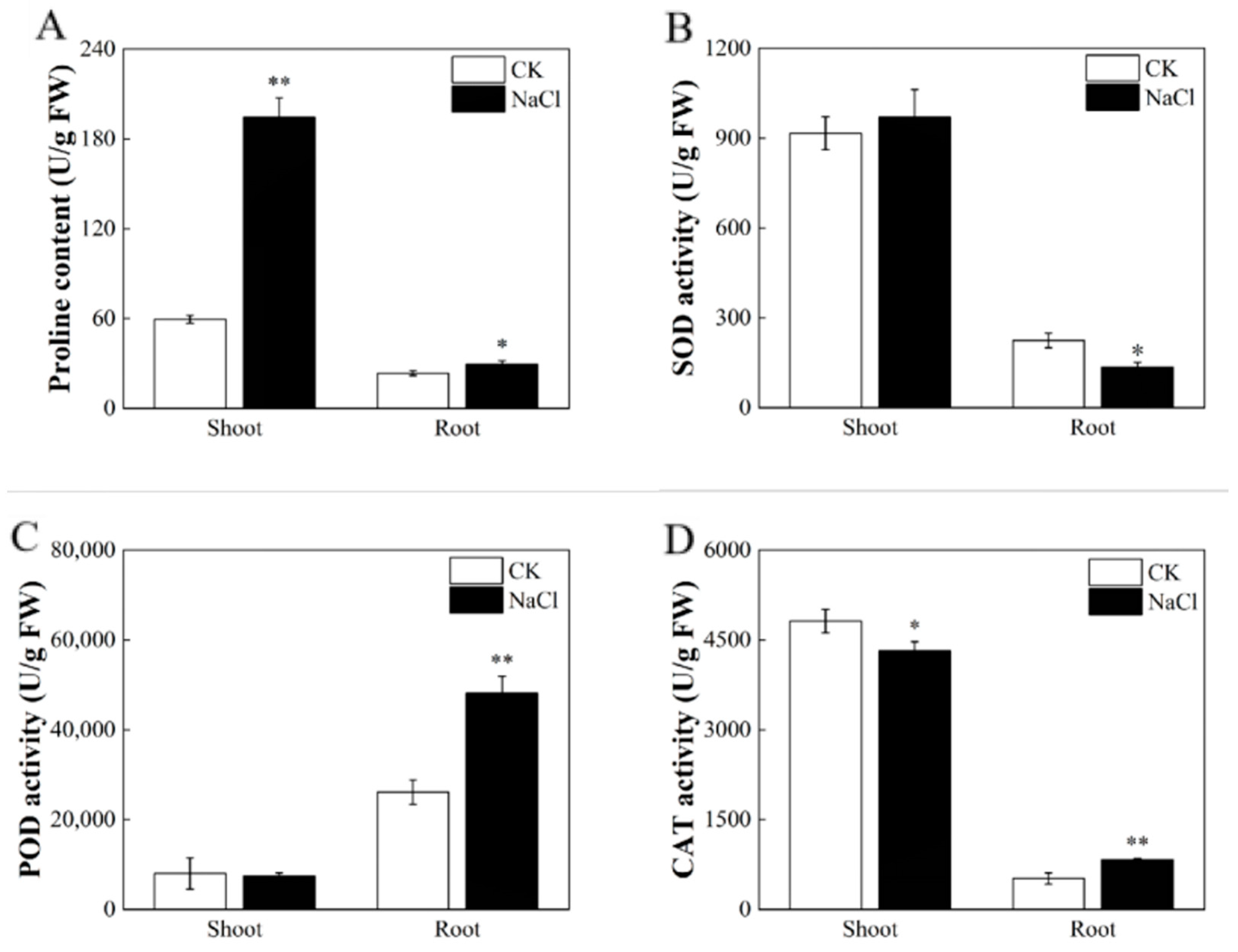
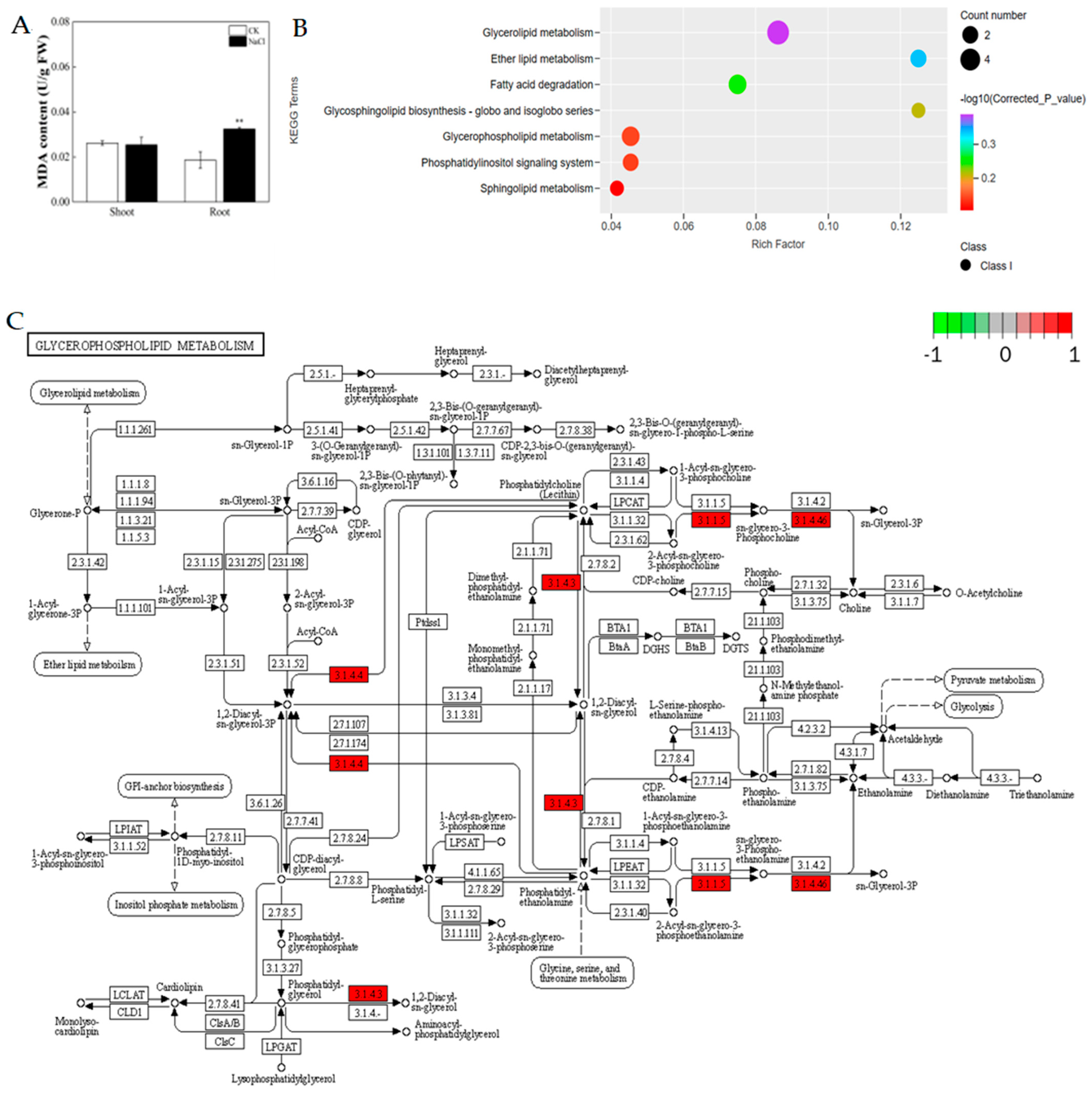
3.2. Response of the Physiological Properties of Rice under Salt Stress
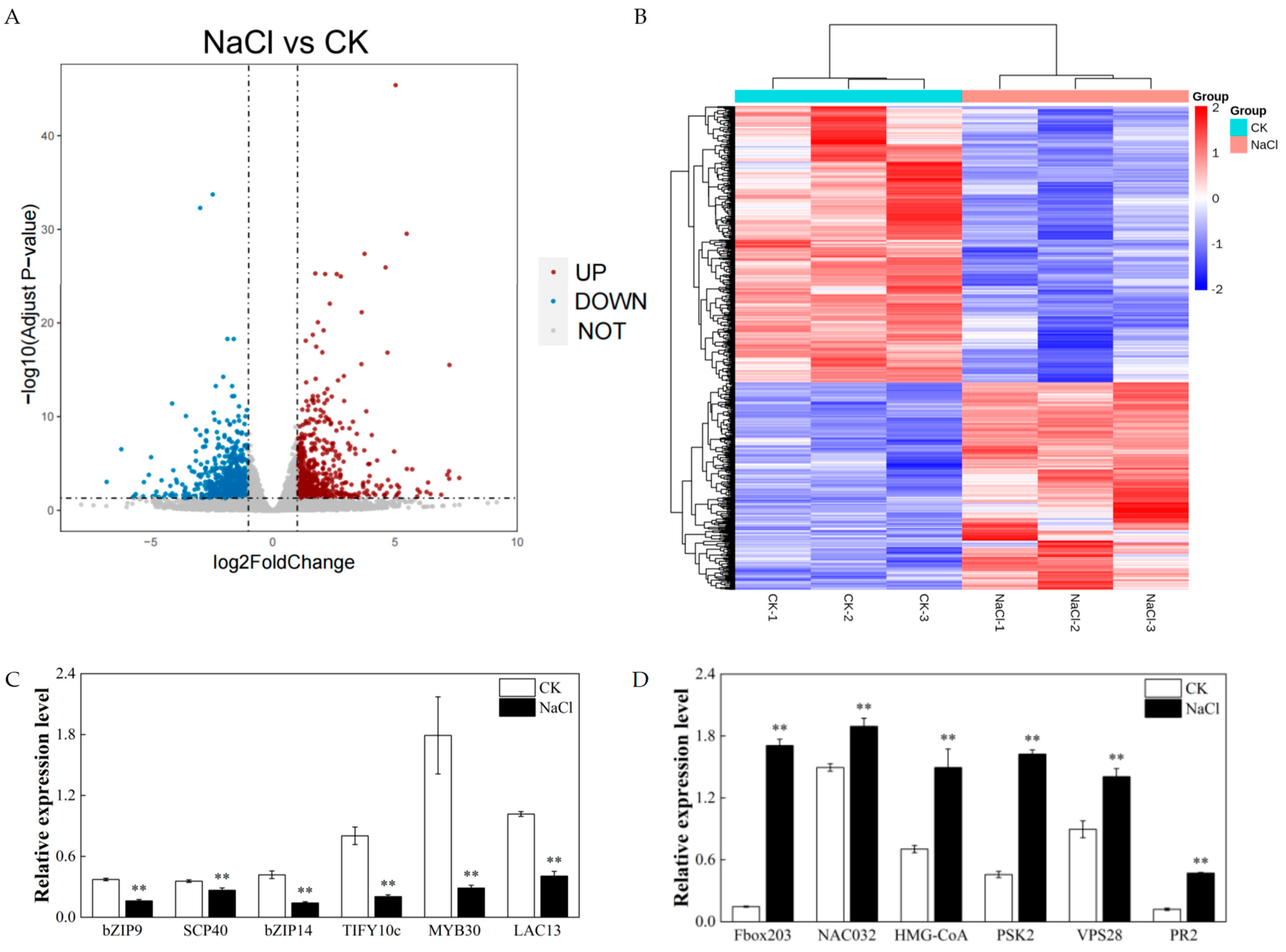
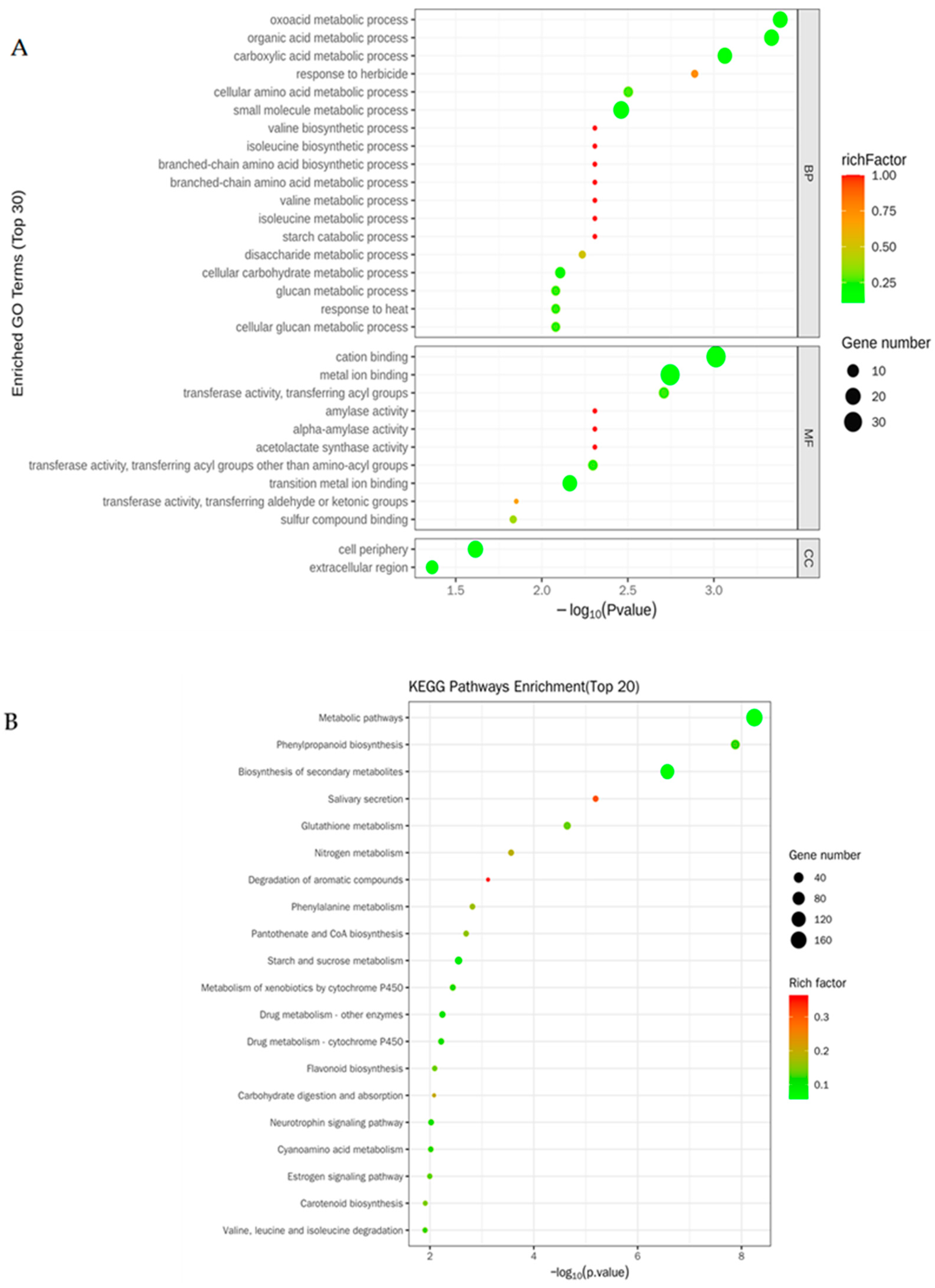
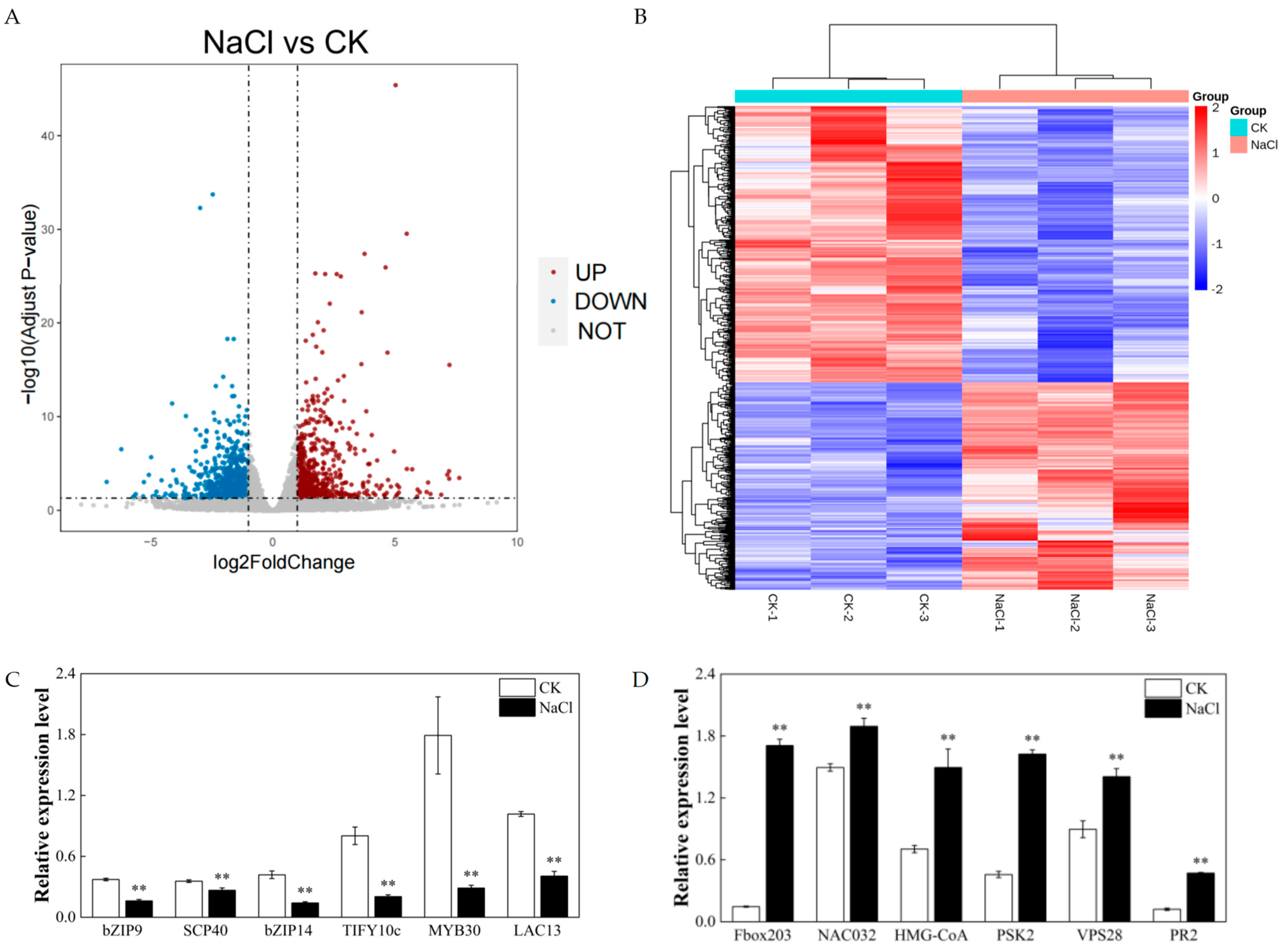
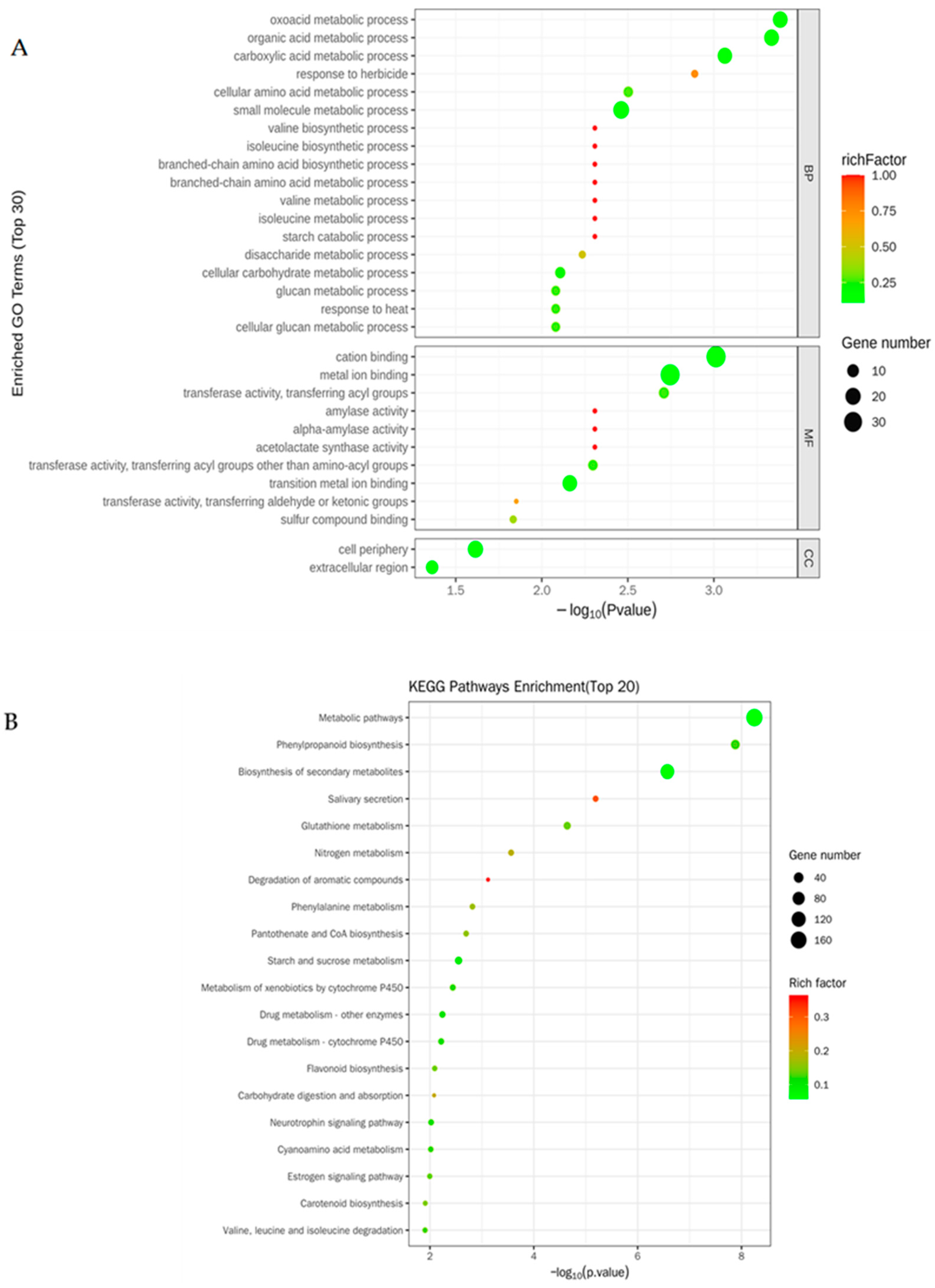
3.3. Changes in the Transcriptomes in Rice Roots Resulting from NaCl Stress
3.4. Expression of Key Genes Associated with Lipids under Salt Stress
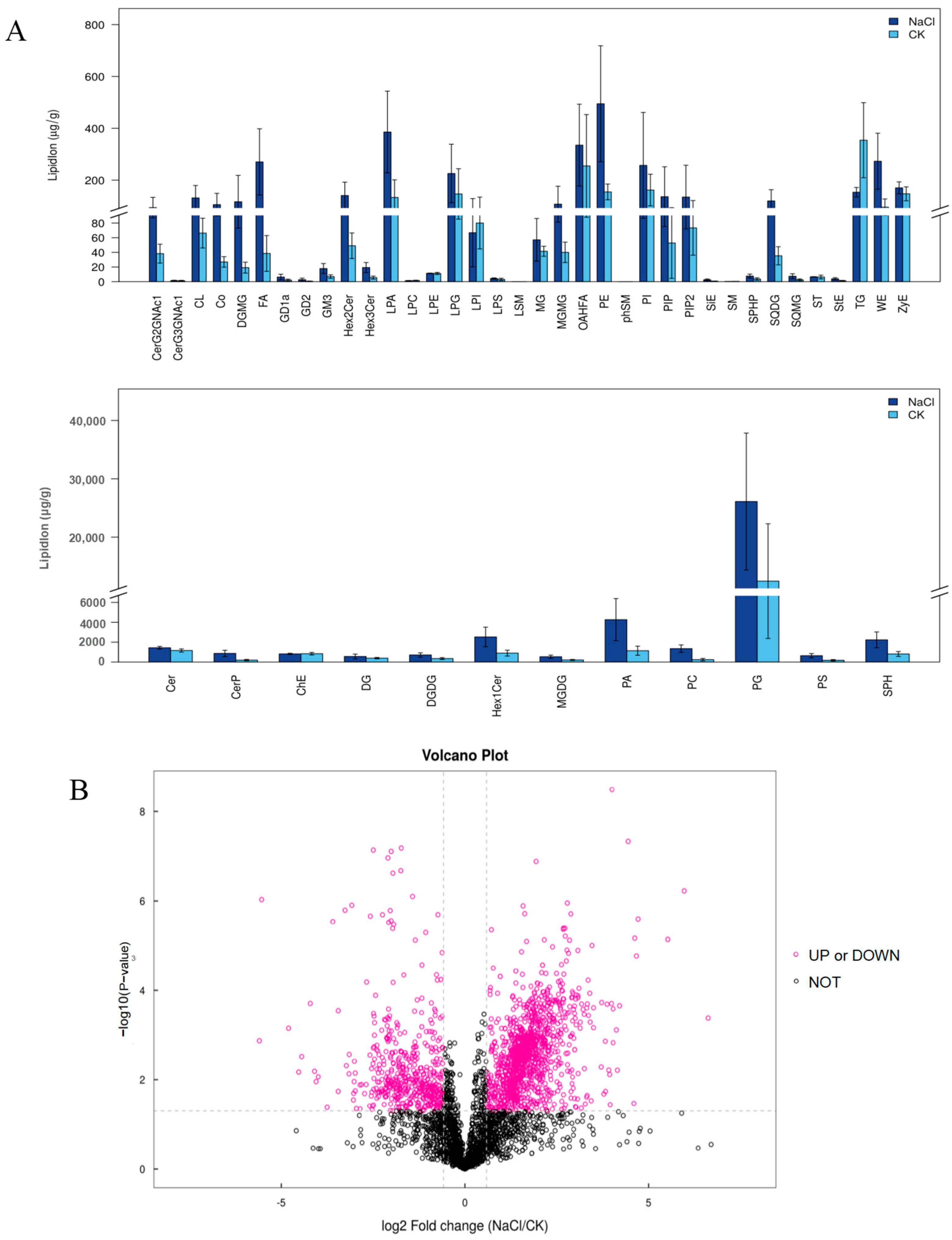
3.5. Response of the Lipid Composition of Rice under Salt Stress
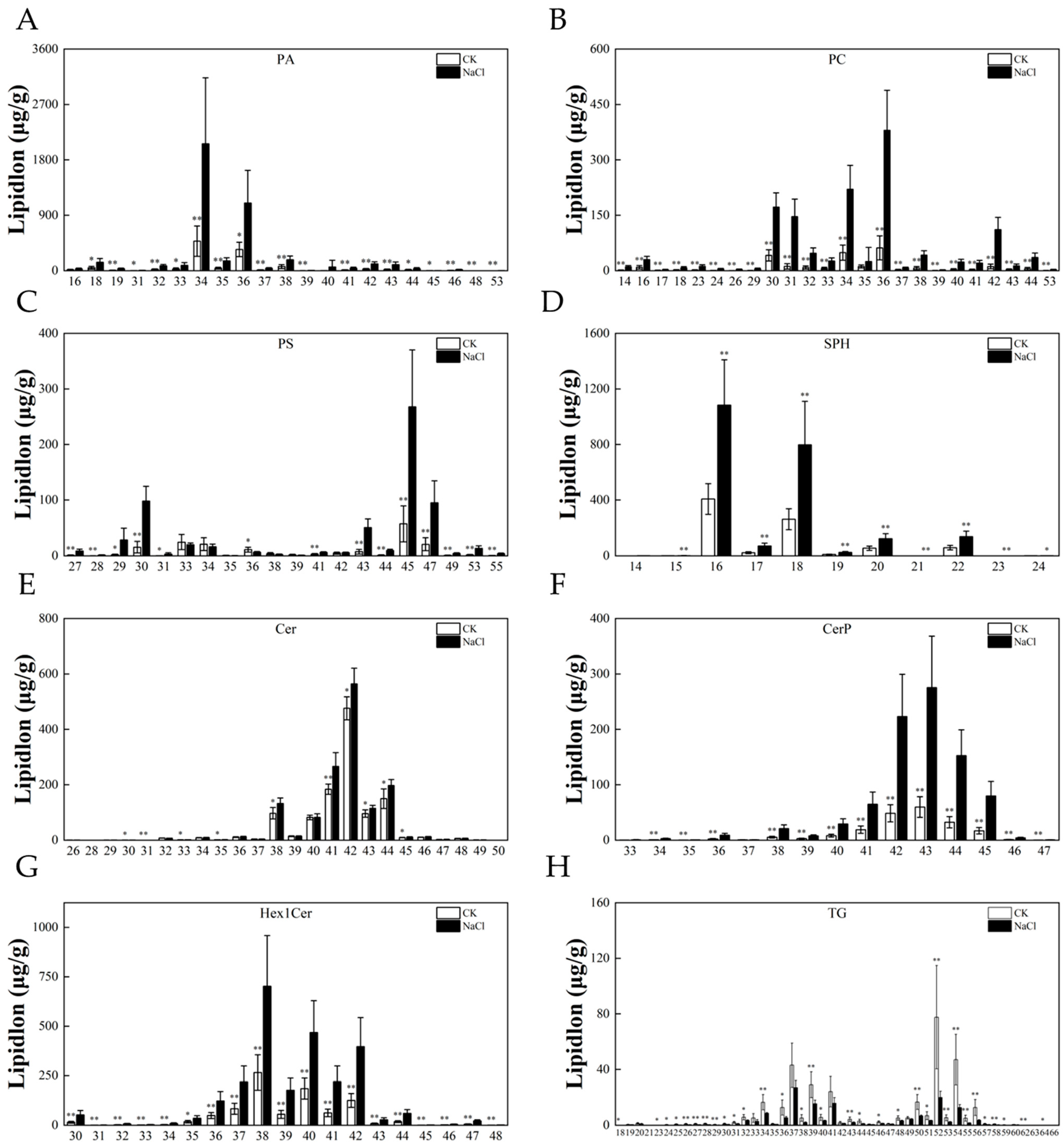
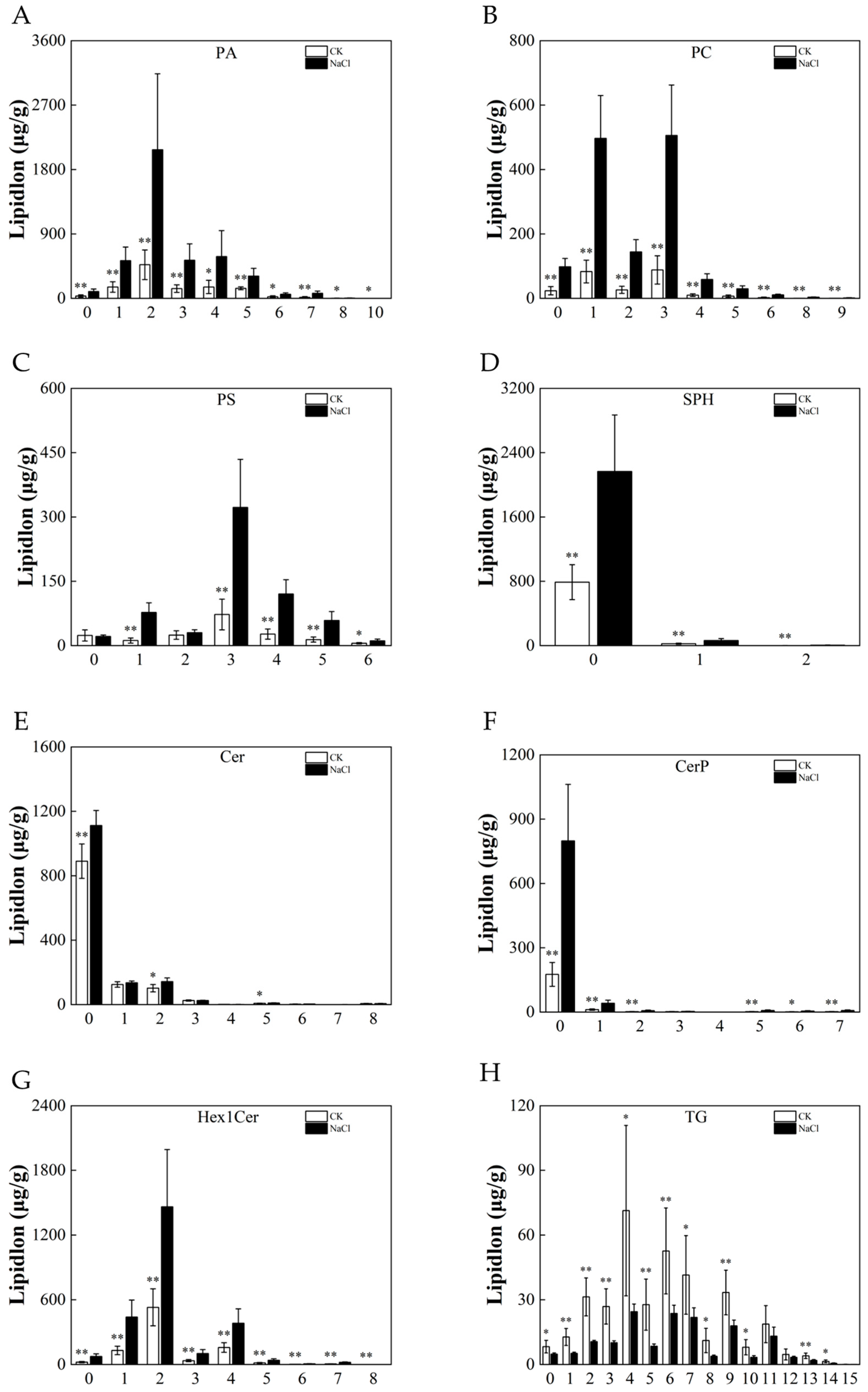
3.6. Response of the Lipid Chain Length and Chain Saturation under Salt Stress
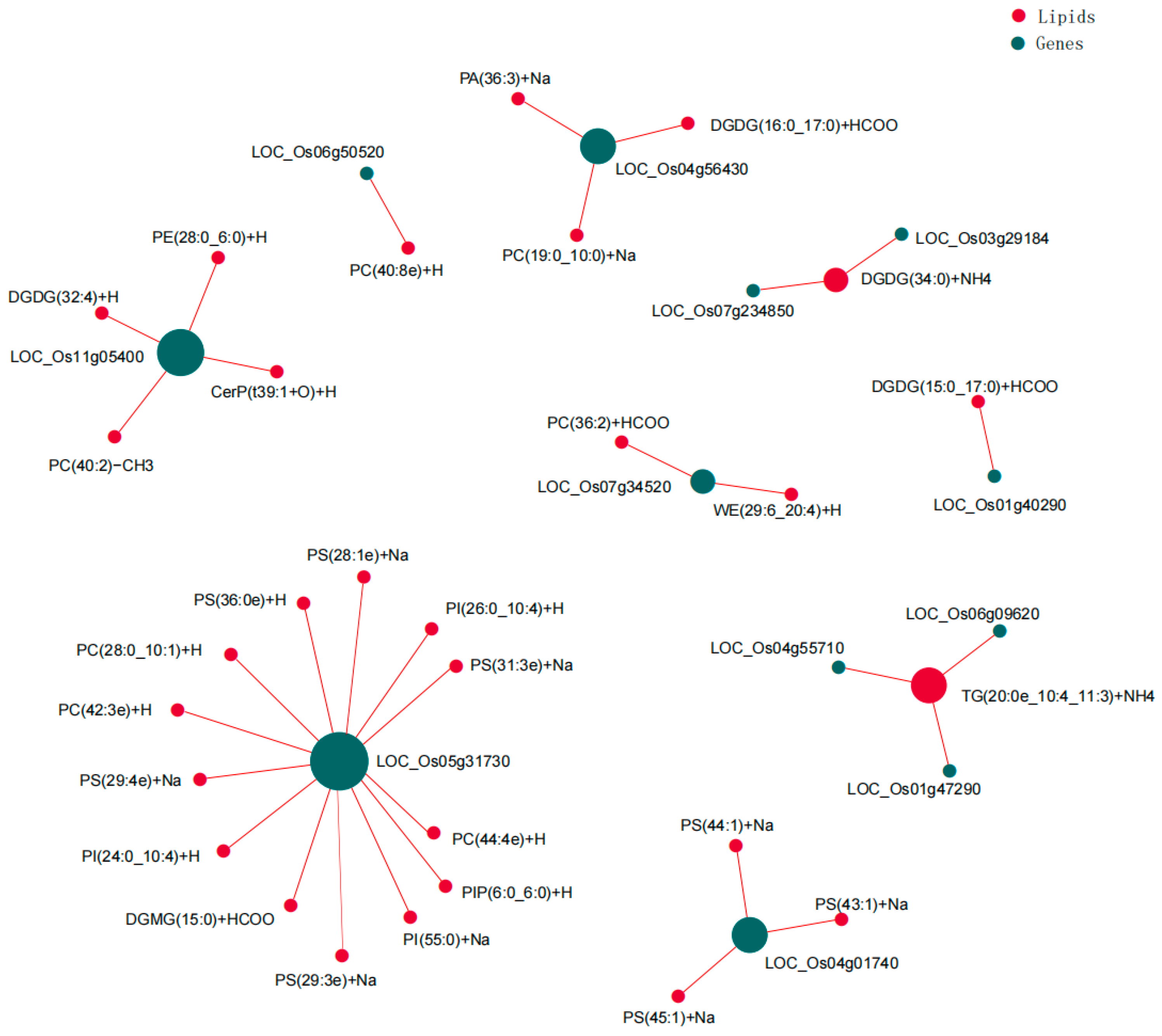
3.7. Multi-Omics Combined Analysis of Rice Seedlings’ Roots in Response to Salt Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lou, W.; Wu, L.; Chen, H.; Ji, Z.; Sun, Y. Assessment of Rice Yield Loss Due to Torrential Rain: A Case Study of Yuhang County, Zhejiang Province, China. Nat. Hazards 2012, 60, 311–320. [Google Scholar] [CrossRef]
- Solis, C.A.; Yong, M.T.; Vinarao, R.; Jena, K.; Holford, P.; Shabala, L.; Zhou, M.; Shabala, S.; Chen, Z.H. Back to the Wild: On a Quest for Donors toward Salinity Tolerant Rice. Front. Plant Sci. 2020, 11, 323. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Liu, C.; Liu, L.; Tan, Y.; Sheng, X.; Yu, D.; Sun, Z.; Sun, X.; Chen, J.; Yuan, D.; et al. Effect of Salinity Stress on Rice Yield and Grain Quality: A Meta-Analysis. Eur. J. Agron. 2023, 144, 126765. [Google Scholar] [CrossRef]
- Djaman, K.; Mel, V.; Boye, A.; Diop, L.; Manneh, B.; El-Namaky, R.; Koudahe, K.; Futakuchi, K. Rice Genotype and Fertilizer Management for Improving Rice Productivity under Saline Soil Conditions. Paddy Water Environ. 2020, 18, 43–57. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. NaCl-Induced Senescence in Leaves of Rice (Oryza sativa L.) Cultivars Differing in Salinity Resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Khatun, S.; Flowers, T.J. Effects of Salinity on Seed Set in Rice. Plant Cell Environ. 1995, 18, 61–67. [Google Scholar] [CrossRef]
- Khan, M.S.A.; Hamid, A.; M Salahuddin, A.B.; Quasem, A.; Karim, M.A. Effect of Sodium Chloride on Growth, Photosynthesis and Mineral Ions Accumulation of Different Types of Rice (Oryza sativa L.). J. Agron. Crop Sci. 1997, 179, 149–161. [Google Scholar] [CrossRef]
- Zeng, L.; Shannon, M.C. Effects of Salinity on Grain Yield and Yield Components of Rice at Different Seeding Densities. Agron. J. 2000, 92, 418–423. [Google Scholar] [CrossRef]
- Liu, C.; Mao, B.; Yuan, D.; Chu, C.; Duan, M. Salt Tolerance in Rice: Physiological Responses and Molecular Mechanisms. Crop J. 2022, 10, 13–25. [Google Scholar] [CrossRef]
- Kohli, S.K.; Khanna, K.; Bhardwaj, R.; Abd Allah, E.F.; Ahmad, P.; Corpas, F.J. Assessment of Subcellular ROS and NO Metabolism in Higher Plants: Multifunctional Signaling Molecules. Antioxidants 2019, 8, 641. [Google Scholar] [CrossRef]
- Ben Abdallah, S.; Aung, B.; Amyot, L.; Lalin, I.; Lachâal, M.; Karray-Bouraoui, N.; Hannoufa, A. Salt Stress (NaCl) Affects Plant Growth and Branch Pathways of Carotenoid and Flavonoid Biosyntheses in Solanum nigrum. Acta Physiol. Plant 2016, 38, 1–13. [Google Scholar] [CrossRef]
- Jammes, F.; Song, C.; Shin, D.; Munemasa, S.; Takeda, K.; Gu, D.; Cho, D.; Lee, S.; Giordo, R.; Sritubtim, S.; et al. MAP Kinases MPK9 and MPK12 Are Preferentially Expressed in Guard Cells and Positively Regulate ROS-Mediated ABA Signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 20520–20525. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Jia, W.; Zhang, J. AtMKK1 Mediates ABA-Induced CAT1 Expression and H2O2 Production via AtMPK6-Coupled Signaling in Arabidopsis. Plant J. 2008, 54, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Hasanuzzaman, M.; Nahar, K.; Fujita, M. Exogenous Proline and Glycine Betaine Mediated Upregulation of Antioxidant Defense and Glyoxalase Systems Provides Better Protection against Salt-Induced Oxidative Stress in Two Rice (Oryza sativa L.) Varieties. Biomed. Res. Int. 2014, 2014, 757219. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xu, J.; Meng, X.; Fang, X.; Xia, M.; Zhang, J.; Cao, S.; Fan, T. Linker Histone Variant HIS1-3 and WRKY1 Oppositely Regulate Salt Stress Tolerance in Arabidopsis. Plant Physiol. 2022, 189, 1833–1847. [Google Scholar] [CrossRef]
- Liu, S.; Yang, R.; Liu, M.; Zhang, S.; Yan, K.; Yang, G.; Huang, J.; Zheng, C.; Wu, C. PLATZ2 Negatively Regulates Salt Tolerance in Arabidopsis Seedlings by Directly Suppressing the Expression of the CBL4/SOS3 and CBL10/SCaBP8 Genes. J. Exp. Bot. 2020, 71, 5589–5602. [Google Scholar] [CrossRef]
- Fu, L.; Shen, Q.; Kuang, L.; Wu, D.; Zhang, G. Transcriptomic and Alternative Splicing Analyses Reveal Mechanisms of the Difference in Salt Tolerance between Barley and Rice. Environ. Exp. Bot. 2019, 166, 103810. [Google Scholar] [CrossRef]
- Liu, S.; Zheng, L.; Xue, Y.; Zhang, Q.; Wang, L.; Shou, H. Overexpression of OsVP1 and OsNHX1 Increases Tolerance to Drought and Salinity in Rice. J. Plant Biol. 2010, 53, 444–452. [Google Scholar] [CrossRef]
- Fukuda, A.; Nakamura, A.; Hara, N.; Toki, S.; Tanaka, Y. Molecular and Functional Analyses of Rice NHX-Type Na+/H+ Antiporter Genes. Planta 2011, 233, 175–188. [Google Scholar] [CrossRef]
- Lv, S.; Tai, F.; Guo, J.; Jiang, P.; Lin, K.; Wang, D.; Zhang, X.; Li, Y. Phosphatidylserine Synthase from Salicornia Europaea Is Involved in Plant Salt Tolerance by Regulating Plasma Membrane Stability. Plant Cell Physiol. 2021, 62, 66–79. [Google Scholar] [CrossRef]
- Mansour, M.M.F. Plasma Membrane Permeability as an Indicator of Salt Tolerance in Plants. Biol. Plant 2013, 57, 1–10. [Google Scholar] [CrossRef]
- Li, M.; Yu, A.; Sun, Y.; Hu, Q.; Kang, J.; Chen, L.; Zhu, X.; Yang, Q.; Long, R. Lipid Composition Remodeling and Storage Lipid Conversion Play a Critical Role in Salt Tolerance in Alfalfa (Medicago sativa L.) Leaves. Environ. Exp. Bot. 2023, 205, 105144. [Google Scholar] [CrossRef]
- Yu, D.; Boughton, B.A.; Hill, C.B.; Feussner, I.; Roessner, U.; Rupasinghe, T.W.T. Insights Into Oxidized Lipid Modification in Barley Roots as an Adaptation Mechanism to Salinity Stress. Front. Plant Sci. 2020, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Sarabia, L.D.; Boughton, B.A.; Rupasinghe, T.; Callahan, D.L.; Hill, C.B.; Roessner, U. Comparative Spatial Lipidomics Analysis Reveals Cellular Lipid Remodelling in Different Developmental Zones of Barley Roots in Response to Salinity. Plant Cell Environ. 2020, 43, 327–343. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Liu, L.; Barkla, B.J. Membrane Lipid Remodeling in Response to Salinity. Int. J. Mol. Sci. 2019, 20, 4264. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Song, T.; Wallrad, L.; Kudla, J.; Wang, X.; Zhang, W. Tissue-Specific Accumulation of PH-Sensing Phosphatidic Acid Determines Plant Stress Tolerance. Nat. Plants 2019, 5, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Nie, J.; Cao, C.; Jin, Y.; Yan, M.; Wang, F.; Liu, J.; Xiao, Y.; Liang, Y.; Zhang, W. Phosphatidic Acid Mediates Salt Stress Response by Regulation of MPK6 in Arabidopsis thaliana. New Phytol. 2010, 188, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shen, L.; Han, X.; He, G.; Fan, W.; Li, Y.; Yang, S.; Zhang, Z.; Yang, Y.; Jin, W.; et al. Phosphatidic Acid–Regulated SOS2 Controls Sodium and Potassium Homeostasis in Arabidopsis under Salt Stress. EMBO J. 2023, 42, e112401. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Liu, X.; Gao, H.; Zhang, B.; Peng, F.; Xiao, Y. Phosphatidylcholine Enhances Homeostasis in Peach Seedling Cell Membrane and Increases Its Salt Stress Tolerance by Phosphatidic Acid. Int. J. Mol. Sci. 2022, 23, 2585. [Google Scholar] [CrossRef]
- Qiao, K.; Wang, M.; Takano, T.; Liu, S. Overexpression of Acyl-Coa-Binding Protein 1 (ChACBP1) from Saline-Alkali-Tolerant Chlorella Sp. Enhances Stress Tolerance in Arabidopsis. Front. Plant Sci. 2018, 9, 1772. [Google Scholar] [CrossRef]
- Yu, Y.; Kou, M.; Gao, Z.; Liu, Y.; Xuan, Y.; Liu, Y.; Tang, Z.; Cao, Q.; Li, Z.; Sun, J. Involvement of Phosphatidylserine and Triacylglycerol in the Response of Sweet Potato Leaves to Salt Stress. Front. Plant Sci. 2019, 10, 1086. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Yang, Y. Phospholipids in Salt Stress Response. Plants 2021, 10, 2204. [Google Scholar] [CrossRef]
- Zhang, H.; Zhai, J.; Mo, J.; Li, D.; Song, F. Overexpression of Rice Sphingosine-1-Phoshpate Lyase Gene OsSPL1 in Transgenic Tobacco Reduces Salt and Oxidative Stress Tolerance. J. Integr. Plant Biol. 2012, 54, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.X.; Li, J.; Liu, Z.; Yin, J.; Chang, Z.Y.; Rong, C.; Wu, J.L.; Bi, F.C.; Yao, N. The Arabidopsis Ceramidase AtACER Functions in Disease Resistance and Salt Tolerance. Plant J. 2015, 81, 767–780. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Li, X.; Luo, M. Detailed Sphingolipid Profile Responded to Salt Stress in Cotton Root and the GhIPCS1 Is Involved in the Regulation of Plant Salt Tolerance. Plant Sci. 2022, 316, 111174. [Google Scholar] [CrossRef]
- Gu, J.; Hu, C.; Jia, X.; Ren, Y.; Su, D.; He, J. Physiological and Biochemical Bases of Spermidine-Induced Alleviation of Cadmium and Lead Combined Stress in Rice. Plant Physiol. Biochem. 2022, 189, 104–114. [Google Scholar] [CrossRef]
- Yan, F.; Wei, H.; Ding, Y.; Li, W.; Liu, Z.; Chen, L.; Tang, S.; Ding, C.; Jiang, Y.; Li, G. Melatonin Regulates Antioxidant Strategy in Response to Continuous Salt Stress in Rice Seedlings. Plant Physiol. Biochem. 2021, 165, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, K.; Wu, C.; Zhao, Y.; Yin, X.; Zhang, B.; Grierson, D.; Chen, K.; Xu, C. Effect of Ethylene on Cell Wall and Lipid Metabolism during Alleviation of Postharvest Chilling Injury in Peach. Cells 2019, 8, 1612. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Zhang, J.; Zhong, C.; Zhu, L.; Cao, X.; Yu, S.; Allen Bohr, J.; Hu, J.; Jin, Q. Effects of Salt Stress on Rice Growth, Development Characteristics, and the Regulating Ways: A Review. J. Integr. Agric. 2017, 16, 2357–2374. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Yousuf, P.Y.; Ahmad, A.; Ganie, A.H.; Sareer, O.; Krishnapriya, V.; Aref, I.M.; Iqbal, M. Antioxidant Response and Proteomic Modulations in Indian Mustard Grown under Salt Stress. Plant Growth Regul. 2017, 81, 31–50. [Google Scholar] [CrossRef]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant Salt-Tolerance Mechanism: A Review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Chan, Z. Ros Regulation during Abiotic Stress Responses in Crop Plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [PubMed]
- Bai, A.; Lu, X. Effects of Calcium and Calcium Effectors on Antioxidant System and Osmotic Adjustment Substances Content of Sour Jujube (Ziziphus jujuba Var. spinosa) Seedlings under NaCl Stress. Plant Physiol. J. 2020, 56, 1910–1920. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, J.; Yan, B.; Wei, Y.; Ge, S.; Li, J.; Han, Y.; Li, Z.; Zhao, C.; Xu, J. The Adjustment of Membrane Lipid Metabolism Pathways in Maize Roots Under Saline–Alkaline Stress. Front. Plant Sci. 2021, 12, 635327. [Google Scholar] [CrossRef] [PubMed]
- Testerink, C.; Munnik, T. Phosphatidic Acid: A Multifunctional Stress Signaling Lipid in Plants. Trends Plant Sci. 2005, 10, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Pan, X.; Welti, R.; Wang, X. Phospholipase Dα3 Is Involved in the Hyperosmotic Response in Arabidopsis. Plant Cell 2008, 20, 803–816. [Google Scholar] [CrossRef]
- Arisz, S.A.; Testerink, C.; Munnik, T. Plant PA Signaling via Diacylglycerol Kinase. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2009, 1791, 869–875. [Google Scholar] [CrossRef]
- Liu, C.; Yin, H.; Gao, P.; Hu, X.; Yang, J.; Liu, Z.; Fu, X.; Luo, D. Phosphatidylserine Synthase1 Is Required for Inflorescence Meristem and Organ Development in Arabidopsis. J. Integr. Plant Biol. 2013, 55, 682–695. [Google Scholar] [CrossRef]
- Ma, J.; Cheng, Z.; Chen, J.; Shen, J.; Zhang, B.; Ren, Y.; Ding, Y.; Zhou, Y.; Zhang, H.; Zhou, K.; et al. Phosphatidylserine Synthase Controls Cell Elongation Especially in the Uppermost Internode in Rice by Regulation of Exocytosis. PLoS ONE 2016, 11, e0153119. [Google Scholar] [CrossRef] [PubMed]
- Chapman, K.D.; Ohlrogge, J.B. Compartmentation of Triacylglycerol Accumulation in Plants. J. Biol. Chem. 2012, 287, 2288–2294. [Google Scholar] [CrossRef] [PubMed]
- Chapman, K.D.; Aziz, M.; Dyer, J.M.; Mullen, R.T. Mechanisms of Lipid Droplet Biogenesis. Biochem. J. 2019, 476, 1929–1942. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Shanklin, J. Triacylglycerol Metabolism, Function, and Accumulation in Plant Vegetative Tissues. Annu. Rev. Plant Biol. 2016, 67, 179–206. [Google Scholar] [CrossRef] [PubMed]
- Abida, H.; Dolch, L.J.; Meï, C.; Villanova, V.; Conte, M.; Block, M.A.; Finazzi, G.; Bastien, O.; Tirichine, L.; Bowler, C.; et al. Membrane Glycerolipid Remodeling Triggered by Nitrogen and Phosphorus Starvation in Phaeodactylum tricornutum. Plant Physiol. 2015, 167, 118–136. [Google Scholar] [CrossRef]
- Kim, H.U.; Lee, K.R.; Jung, S.J.; Shin, H.A.; Go, Y.S.; Suh, M.C.; Kim, J.B. Senescence-Inducible LEC2 Enhances Triacylglycerol Accumulation in Leaves without Negatively Affecting Plant Growth. Plant Biotechnol. J. 2015, 13, 1346–1359. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, Y.; Zhou, C.; Feng, N.; Zheng, D.; Shen, X.; Rao, G.; Huang, Y.; Cai, W.; Liu, Y.; Zhang, R. Transcriptomic and Lipidomic Analysis Reveals Complex Regulation Mechanisms Underlying Rice Roots’ Response to Salt Stress. Metabolites 2024, 14, 244. https://doi.org/10.3390/metabo14040244
Xue Y, Zhou C, Feng N, Zheng D, Shen X, Rao G, Huang Y, Cai W, Liu Y, Zhang R. Transcriptomic and Lipidomic Analysis Reveals Complex Regulation Mechanisms Underlying Rice Roots’ Response to Salt Stress. Metabolites. 2024; 14(4):244. https://doi.org/10.3390/metabo14040244
Chicago/Turabian StyleXue, Yingbin, Chenyu Zhou, Naijie Feng, Dianfeng Zheng, Xuefeng Shen, Gangshun Rao, Yongxiang Huang, Wangxiao Cai, Ying Liu, and Rui Zhang. 2024. "Transcriptomic and Lipidomic Analysis Reveals Complex Regulation Mechanisms Underlying Rice Roots’ Response to Salt Stress" Metabolites 14, no. 4: 244. https://doi.org/10.3390/metabo14040244
APA StyleXue, Y., Zhou, C., Feng, N., Zheng, D., Shen, X., Rao, G., Huang, Y., Cai, W., Liu, Y., & Zhang, R. (2024). Transcriptomic and Lipidomic Analysis Reveals Complex Regulation Mechanisms Underlying Rice Roots’ Response to Salt Stress. Metabolites, 14(4), 244. https://doi.org/10.3390/metabo14040244