
Longitudinal Metabolomics Reveals Metabolic Dysregulation Dynamics in Patients with Severe COVID-19

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
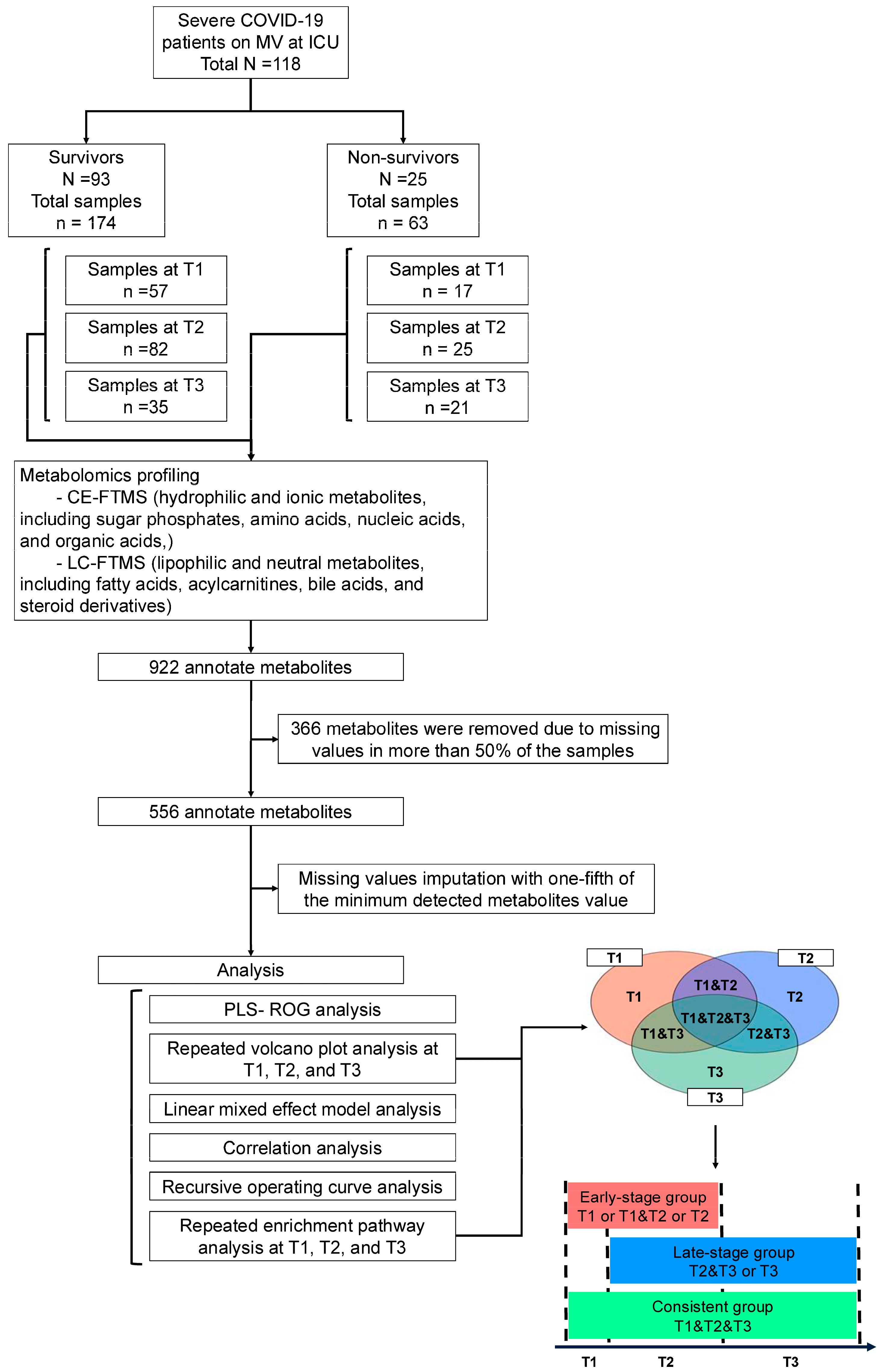
2.1. Study Design and Participants
2.2. Blood Sample Collection
2.3. Metabolome Measurements
2.4. Clinical Parameters and Laboratory Test Measurements
2.5. PCA and PLS-ROG Analysis
2.6. Selection of Differential Metabolites and Enriched Pathways
2.7. Linear Mixed-Effects Model
2.8. Prognostic Ability
2.9. Correlation Analysis
3. Results
3.1. Patient Characteristics
3.2. PLS-ROG Analysis Showed Longitudinal Differences in Metabolic Dysregulation Between Survivors and Non-Survivors
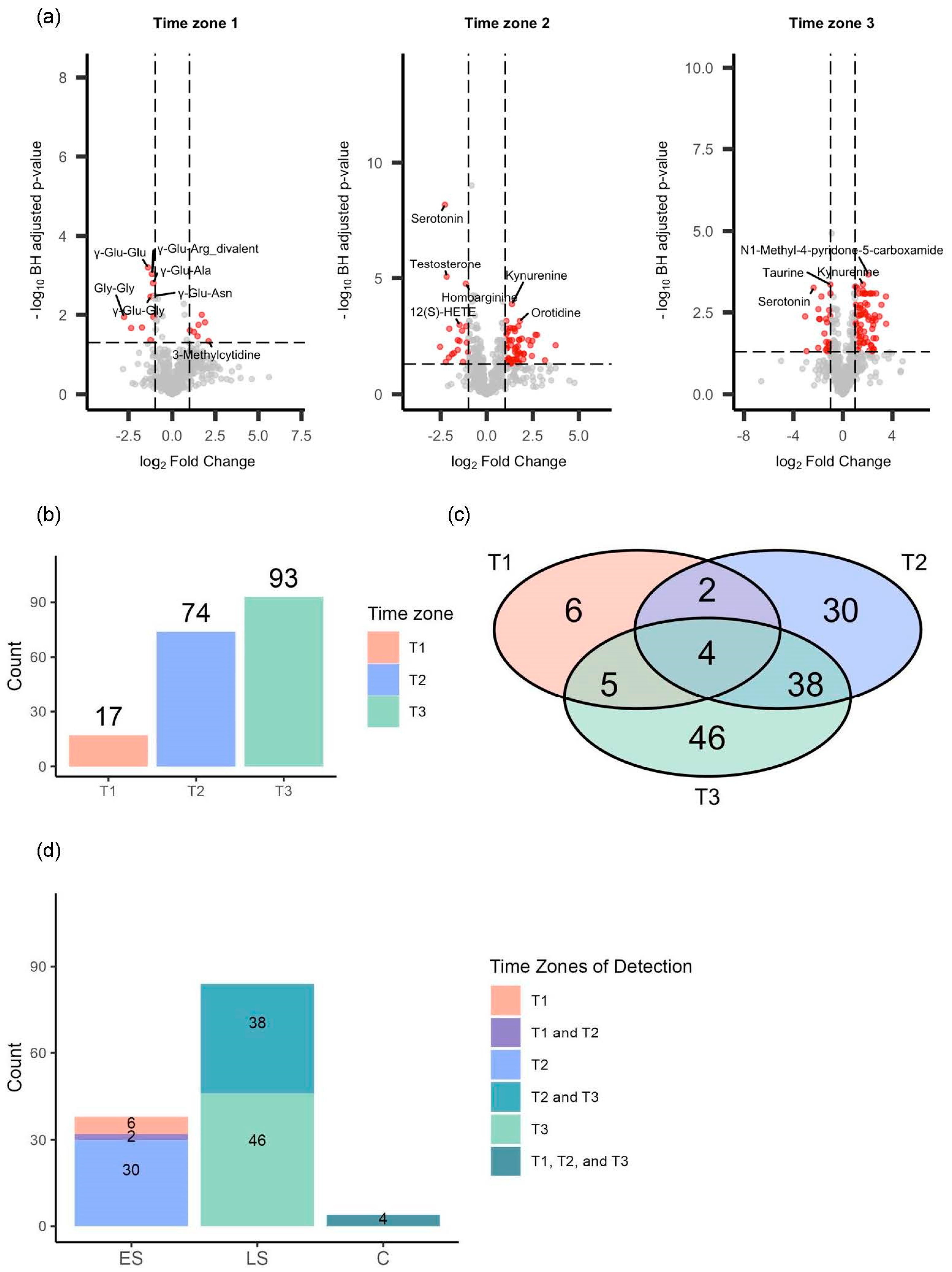
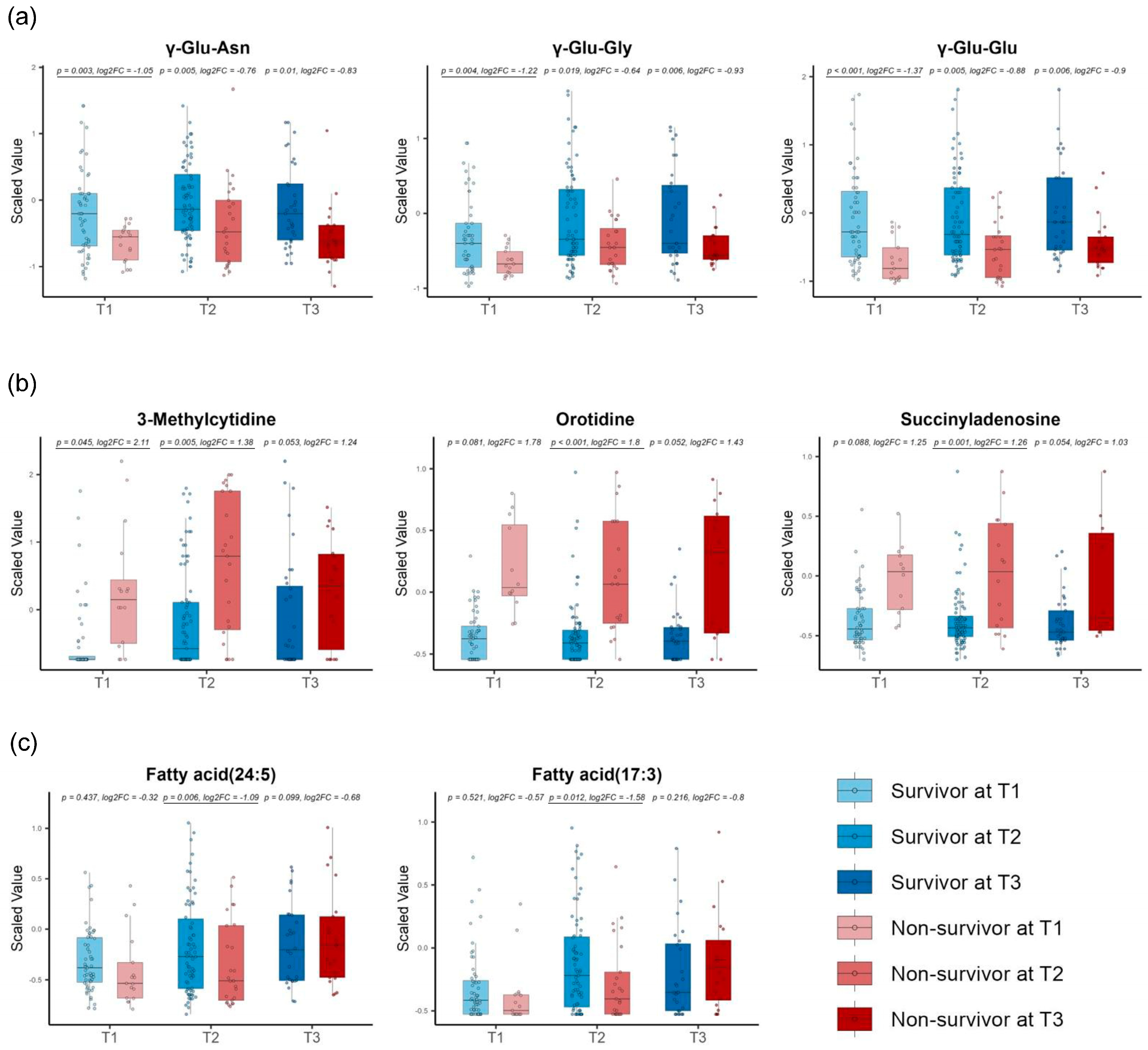
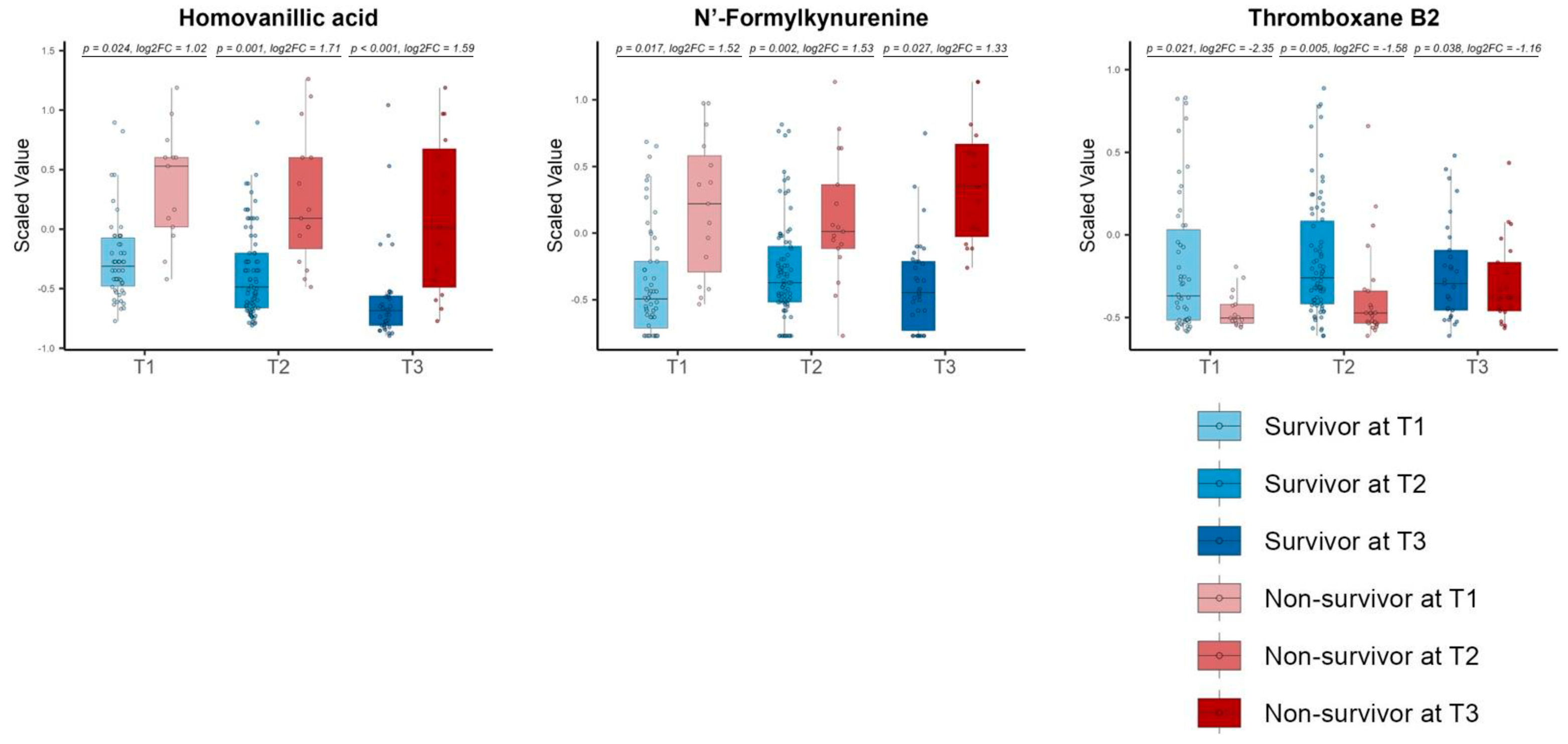
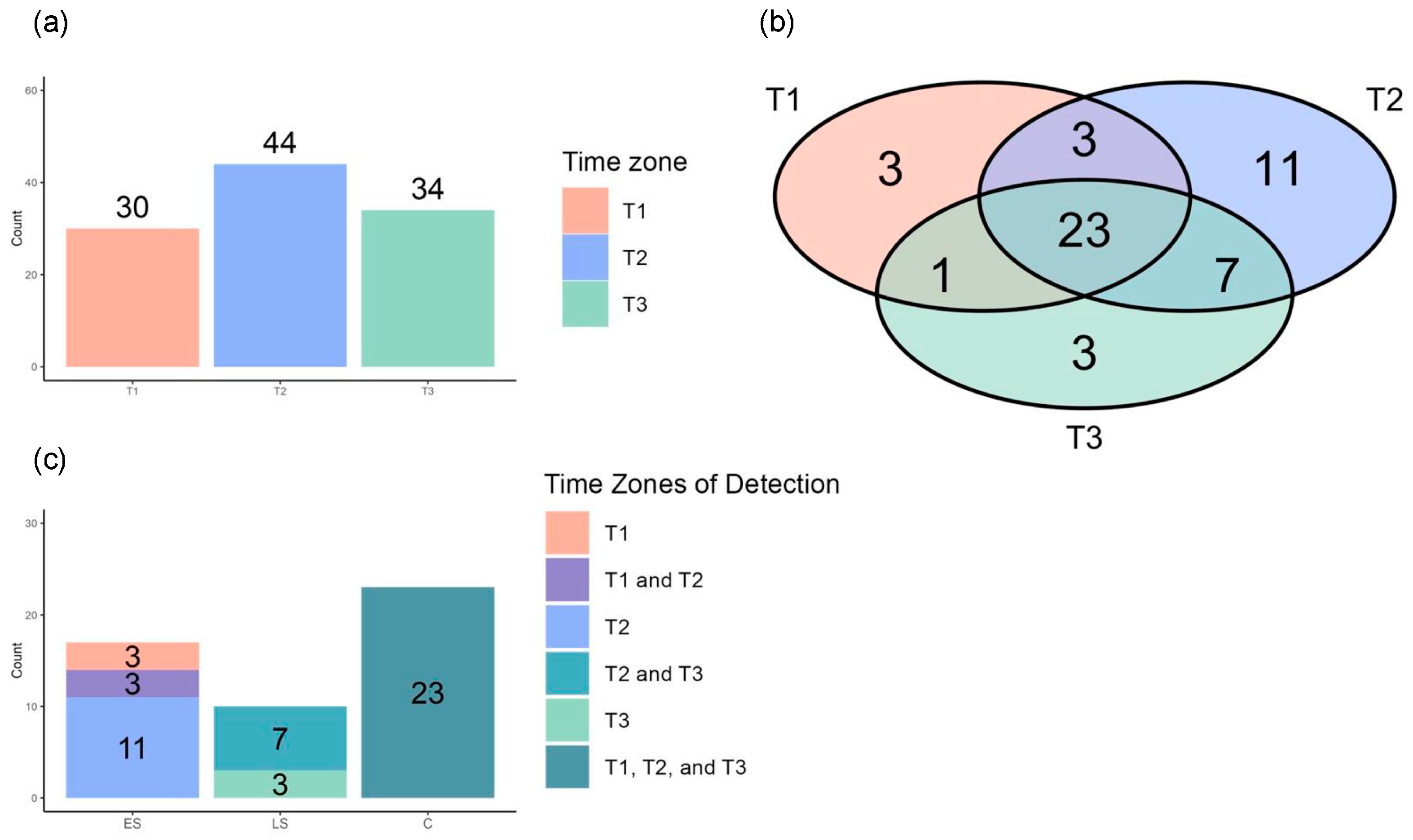
3.3. Repeated Volcano Plot Analysis Identified Differential Metabolites Between Survivors and Non-Survivors at Each Time Zone
3.4. Linear Mixed Models Detected Different Metabolite Trends Between Survivors and Non-Survivors over Time
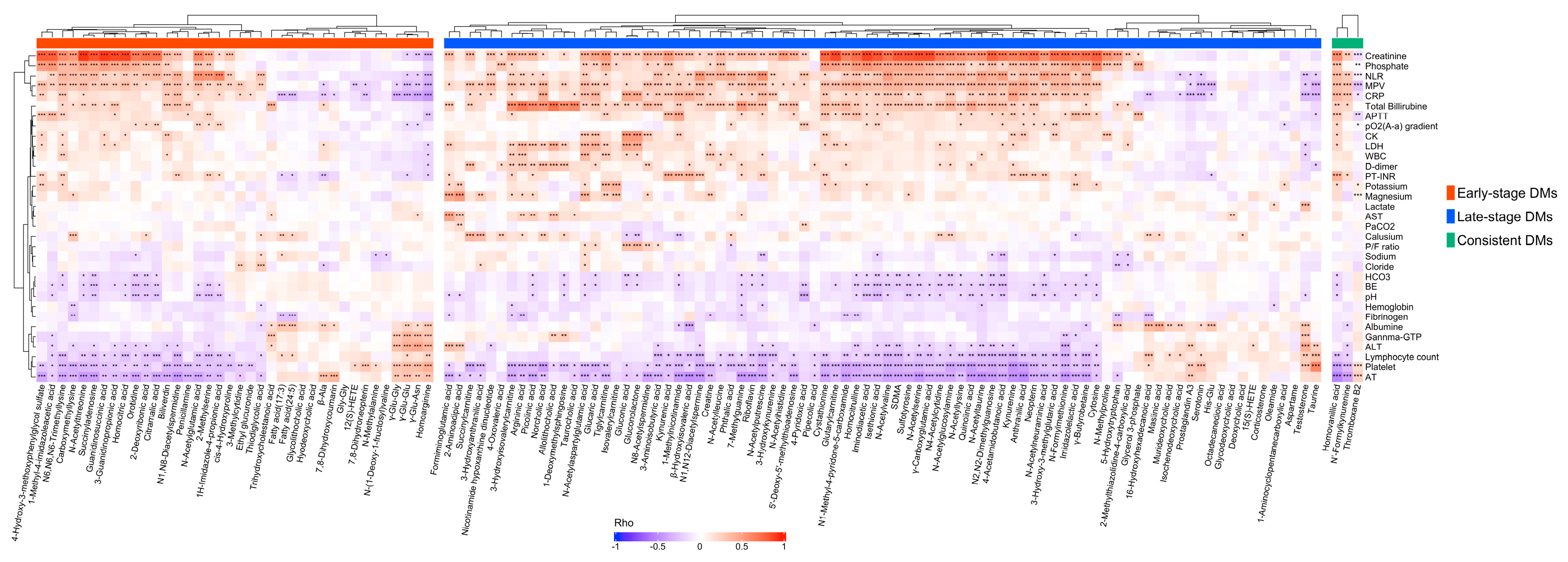
3.5. Correlation Analysis Discovered Correlations of Differential Metabolites with Clinical Laboratory Test Results
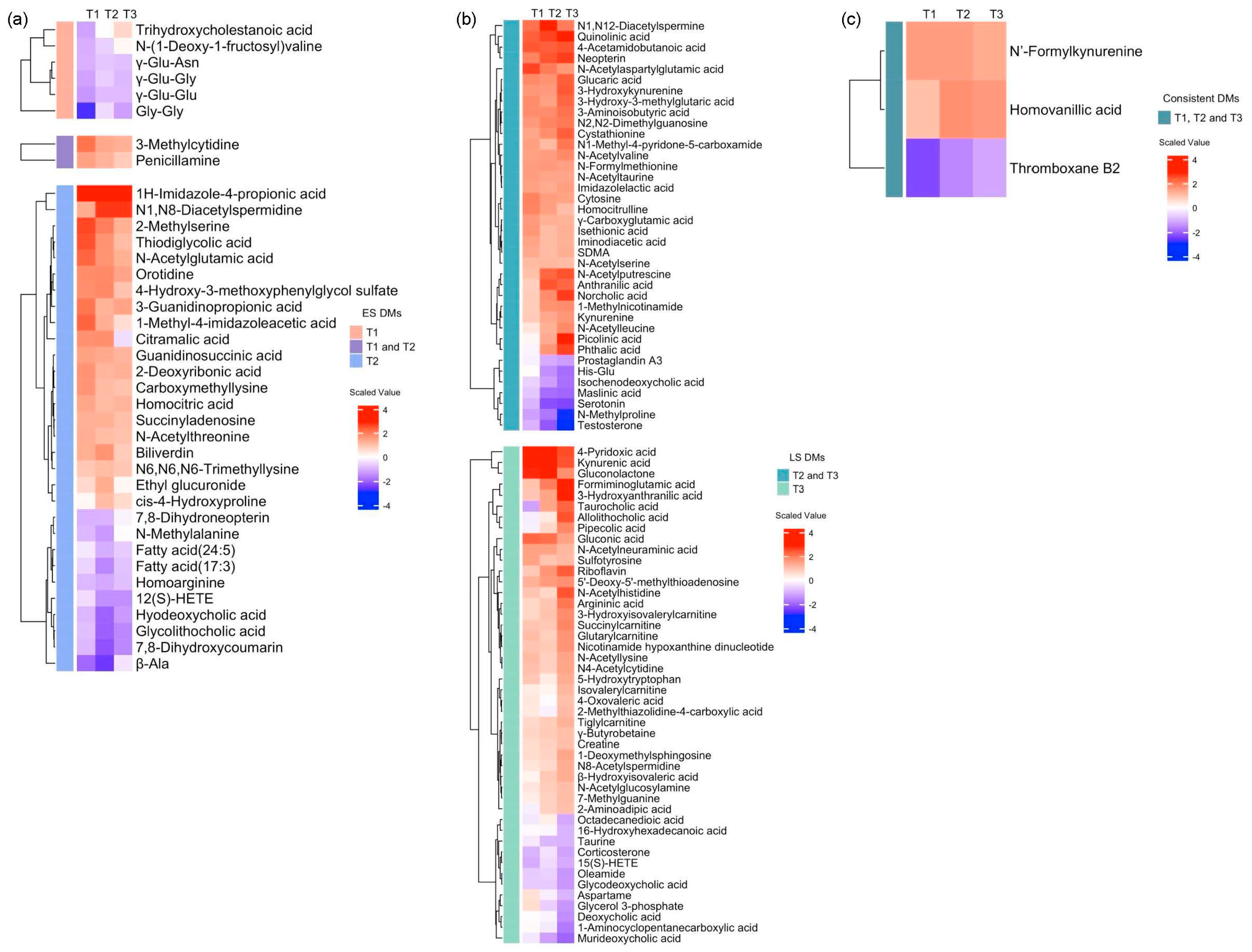
3.6. The Differential Metabolites in the Consistent Group Showed Consistently High Prognostic Relevance
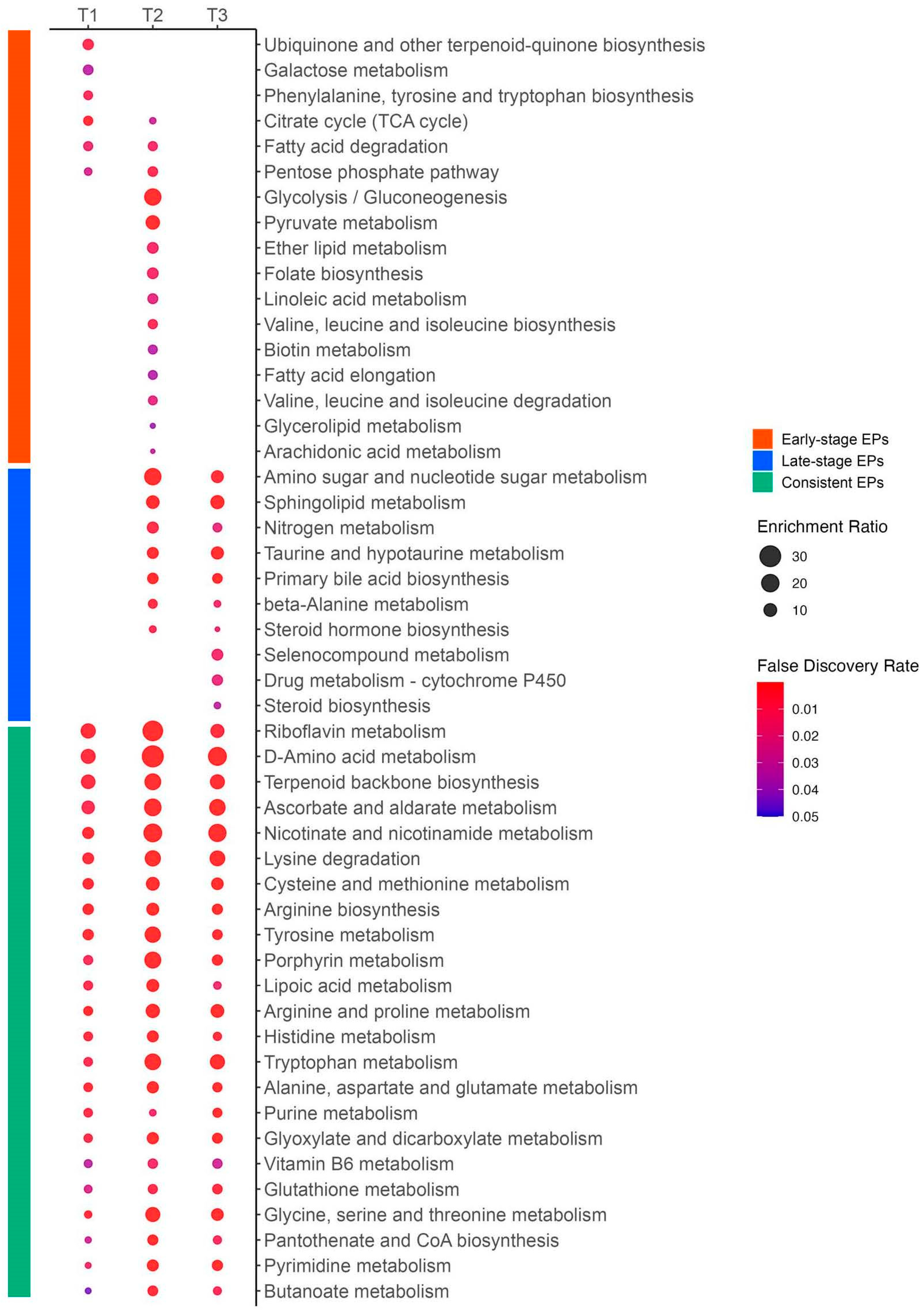
3.7. Repeated Enriched Pathway Analysis Identified Enriched Pathways Between Survivors and Non-Survivors at Each Time Zone
4. Discussion
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andrade Silva, M.; da Silva, A.R.P.A.; do Amaral, M.A.; Fragas, M.G.; Câmara, N.O.S. Metabolic Alterations in SARS-CoV-2 Infection and Its Implication in Kidney Dysfunction. Front. Physiol. 2021, 12, 624698. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Casals, M.; Brito-Zerón, P.; Mariette, X. Systemic and Organ-Specific Immune-Related Manifestations of COVID-19. Nat. Rev. Rheumatol. 2021, 17, 315–332. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.-L. COVID-19: It’s All about Sepsis. Future Microbiol. 2021, 16, 131–133. [Google Scholar] [CrossRef]
- Zaim, S.; Chong, J.H.; Sankaranarayanan, V.; Harky, A. COVID-19 and Multiorgan Response. Curr. Probl. Cardiol. 2020, 45, 100618. [Google Scholar] [CrossRef]
- Termorshuizen, F.; Dongelmans, D.A.; Brinkman, S.; Bakhshi-Raiez, F.; Arbous, M.S.; de Lange, D.W.; van Bussel, B.C.T.; de Keizer, N.F. Dutch COVID-19 Research Consortium Characteristics and Outcome of COVID-19 Patients Admitted to the ICU: A Nationwide Cohort Study on the Comparison between the Consecutive Stages of the COVID-19 Pandemic in the Netherlands, an Update. Ann. Intensive Care 2024, 14, 11. [Google Scholar] [CrossRef] [PubMed]
- Carbonell, R.; Urgelés, S.; Rodríguez, A.; Bodí, M.; Martín-Loeches, I.; Solé-Violán, J.; Díaz, E.; Gómez, J.; Trefler, S.; Vallverdú, M.; et al. Mortality Comparison between the First and Second/Third Waves among 3,795 Critical COVID-19 Patients with Pneumonia Admitted to the ICU: A Multicentre Retrospective Cohort Study. Lancet Reg. Health–Eur. 2021, 11, 100243. [Google Scholar] [CrossRef]
- Chen, P.; Wu, M.; He, Y.; Jiang, B.; He, M.-L. Metabolic Alterations upon SARS-CoV-2 Infection and Potential Therapeutic Targets against Coronavirus Infection. Signal Transduct. Target. Ther. 2023, 8, 237. [Google Scholar] [CrossRef]
- Caterino, M.; Costanzo, M.; Fedele, R.; Cevenini, A.; Gelzo, M.; Di Minno, A.; Andolfo, I.; Capasso, M.; Russo, R.; Annunziata, A.; et al. The Serum Metabolome of Moderate and Severe COVID-19 Patients Reflects Possible Liver Alterations Involving Carbon and Nitrogen Metabolism. Int. J. Mol. Sci. 2021, 22, 9548. [Google Scholar] [CrossRef]
- Martínez-Gómez, L.E.; Ibarra-González, I.; Fernández-Lainez, C.; Tusie, T.; Moreno-Macías, H.; Martinez-Armenta, C.; Jimenez-Gutierrez, G.E.; Vázquez-Cárdenas, P.; Vidal-Vázquez, P.; Ramírez-Hinojosa, J.P.; et al. Metabolic Reprogramming in SARS-CoV-2 Infection Impacts the Outcome of COVID-19 Patients. Front. Immunol. 2022, 13, 936106. [Google Scholar] [CrossRef]
- Codo, A.C.; Davanzo, G.G.; de Brito Monteiro, L.; de Souza, G.F.; Muraro, S.P.; Virgilio-da-Silva, J.V.; Prodonoff, J.S.; Carregari, V.C.; de Biagi Junior, C.A.O.; Crunfli, F.; et al. Elevated Glucose Levels Favor SARS-CoV-2 Infection and Monocyte Response through a HIF-1α/Glycolysis-Dependent Axis. Cell Metab. 2020, 32, 437–446.e5. [Google Scholar] [CrossRef]
- Rodriguez, A.E.; Ducker, G.S.; Billingham, L.K.; Martinez, C.A.; Mainolfi, N.; Suri, V.; Friedman, A.; Manfredi, M.G.; Weinberg, S.E.; Rabinowitz, J.D.; et al. Serine Metabolism Supports Macrophage IL-1β Production. Cell Metab. 2019, 29, 1003–1011.e4. [Google Scholar] [CrossRef] [PubMed]
- Karu, N.; Kindt, A.; van Gammeren, A.J.; Ermens, A.A.M.; Harms, A.C.; Portengen, L.; Vermeulen, R.C.H.; Dik, W.A.; Langerak, A.W.; van der Velden, V.H.J.; et al. Severe COVID-19 Is Characterised by Perturbations in Plasma Amines Correlated with Immune Response Markers, and Linked to Inflammation and Oxidative Stress. Metabolites 2022, 12, 618. [Google Scholar] [CrossRef]
- Maeda, R.; Seki, N.; Uwamino, Y.; Wakui, M.; Nakagama, Y.; Kido, Y.; Sasai, M.; Taira, S.; Toriu, N.; Yamamoto, M.; et al. Amino Acid Catabolite Markers for Early Prognostication of Pneumonia in Patients with COVID-19. Nat. Commun. 2023, 14, 8469. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, F.; Li, Z.; Remes-Lenicov, F.; Dávola, M.E.; Elizalde, M.; Paletta, A.; Ashkar, A.A.; Mossman, K.L.; Dugour, A.V.; Figueroa, J.M.; et al. AHR Signaling Is Induced by Infection with Coronaviruses. Nat. Commun. 2021, 12, 5148. [Google Scholar] [CrossRef] [PubMed]
- Mpekoulis, G.; Kalliampakou, K.I.; Milona, R.S.; Lagou, D.; Ioannidis, A.; Jahaj, E.; Chasapis, C.T.; Kefallinos, D.; Karakasiliotis, I.; Kotanidou, A.; et al. Significance of Catecholamine Biosynthetic/Metabolic Pathway in SARS-CoV-2 Infection and COVID-19 Severity. Cells 2022, 12, 12. [Google Scholar] [CrossRef]
- Limanaqi, F.; Zecchini, S.; Dino, B.; Strizzi, S.; Cappelletti, G.; Utyro, O.; Vanetti, C.; Garziano, M.; Saulle, I.; Clerici, M.; et al. Dopamine Reduces SARS-CoV-2 Replication In Vitro through Downregulation of D2 Receptors and Upregulation of Type-I Interferons. Cells 2022, 11, 1691. [Google Scholar] [CrossRef]
- Jud, P.; Gressenberger, P.; Muster, V.; Avian, A.; Meinitzer, A.; Strohmaier, H.; Sourij, H.; Raggam, R.B.; Stradner, M.H.; Demel, U.; et al. Evaluation of Endothelial Dysfunction and Inflammatory Vasculopathy After SARS-CoV-2 Infection-A Cross-Sectional Study. Front. Cardiovasc. Med. 2021, 8, 750887. [Google Scholar] [CrossRef] [PubMed]
- Glassman, I.; Le, N.; Mirhosseini, M.; Alcantara, C.A.; Asif, A.; Goulding, A.; Muneer, S.; Singh, M.; Robison, J.; Guilford, F.; et al. The Role of Glutathione in Prevention of COVID-19 Immunothrombosis: A Review. Front. Biosci. 2023, 28, 59. [Google Scholar] [CrossRef] [PubMed]
- Žarković, N.; Łuczaj, W.; Jarocka-Karpowicz, I.; Orehovec, B.; Baršić, B.; Tarle, M.; Kmet, M.; Lukšić, I.; Biernacki, M.; Skrzydlewska, E. Diversified Effects of COVID-19 as a Consequence of the Differential Metabolism of Phospholipids and Lipid Peroxidation Evaluated in the Plasma of Survivors and Deceased Patients upon Admission to the Hospital. Int. J. Mol. Sci. 2022, 23, 11810. [Google Scholar] [CrossRef]
- Conti, V.; Corbi, G.; Sabbatino, F.; De Pascale, D.; Sellitto, C.; Stefanelli, B.; Bertini, N.; De Simone, M.; Liguori, L.; Di Paola, I.; et al. Long COVID: Clinical Framing, Biomarkers, and Therapeutic Approaches. J. Pers. Med. 2023, 13, 334. [Google Scholar] [CrossRef]
- Roberts, I.; Wright Muelas, M.; Taylor, J.M.; Davison, A.S.; Xu, Y.; Grixti, J.M.; Gotts, N.; Sorokin, A.; Goodacre, R.; Kell, D.B. Untargeted Metabolomics of COVID-19 Patient Serum Reveals Potential Prognostic Markers of Both Severity and Outcome. Metabolomics 2021, 18, 6. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, A.S.; Mukhtar, N.; Aljomaiah, A.; Aljamei, H.; Bakhsh, A.; Alsudani, N.; Elsayed, T.; Alrashidi, N.; Fadel, R.; Alqahtani, E.; et al. The Impact of COVID-19 Viral Infection on the Hypothalamic-Pituitary-Adrenal Axis. Endocr. Pract. 2021, 27, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Freire Santana, M.; Borba, M.G.S.; Baía-da-Silva, D.C.; Val, F.; Alexandre, M.A.A.; Brito-Sousa, J.D.; Melo, G.C.; Queiroga, M.V.O.; Leão Farias, M.E.; Camilo, C.C.; et al. Case Report: Adrenal Pathology Findings in Severe COVID-19: An Autopsy Study. Am. J. Trop. Med. Hyg. 2020, 103, 1604–1607. [Google Scholar] [CrossRef]
- Sindelar, M.; Stancliffe, E.; Schwaiger-Haber, M.; Anbukumar, D.S.; Adkins-Travis, K.; Goss, C.W.; O’Halloran, J.A.; Mudd, P.A.; Liu, W.-C.; Albrecht, R.A.; et al. Longitudinal Metabolomics of Human Plasma Reveals Prognostic Markers of COVID-19 Disease Severity. Cell Rep. Med. 2021, 2, 100369. [Google Scholar] [CrossRef]
- Li, T.; Ning, N.; Li, B.; Luo, D.; Qin, E.; Yu, W.; Wang, J.; Yang, G.; Nan, N.; He, Z.; et al. Longitudinal Metabolomics Reveals Ornithine Cycle Dysregulation Correlates With Inflammation and Coagulation in COVID-19 Severe Patients. Front. Microbiol. 2021, 12, 723818. [Google Scholar] [CrossRef]
- Chatelaine, H.A.S.; Chen, Y.; Braisted, J.; Chu, S.H.; Chen, Q.; Stav, M.; Begum, S.; Diray-Arce, J.; Sanjak, J.; Huang, M.; et al. Nucleotide, Phospholipid, and Kynurenine Metabolites Are Robustly Associated with COVID-19 Severity and Time of Plasma Sample Collection in a Prospective Cohort Study. Int. J. Mol. Sci. 2023, 25, 346. [Google Scholar] [CrossRef]
- Buyukozkan, M.; Alvarez-Mulett, S.; Racanelli, A.C.; Schmidt, F.; Batra, R.; Hoffman, K.L.; Sarwath, H.; Engelke, R.; Gomez-Escobar, L.; Simmons, W.; et al. Integrative Metabolomic and Proteomic Signatures Define Clinical Outcomes in Severe COVID-19. iScience 2022, 25, 104612. [Google Scholar] [CrossRef] [PubMed]
- Mira, J.C.; Gentile, L.F.; Mathias, B.J.; Efron, P.A.; Brakenridge, S.C.; Mohr, A.M.; Moore, F.A.; Moldawer, L.L. Sepsis Pathophysiology, Chronic Critical Illness, and Persistent Inflammation-Immunosuppression and Catabolism Syndrome. Crit. Care Med. 2017, 45, 253–262. [Google Scholar] [CrossRef]
- Sasaki, K.; Sagawa, H.; Suzuki, M.; Yamamoto, H.; Tomita, M.; Soga, T.; Ohashi, Y. Metabolomics Platform with Capillary Electrophoresis Coupled with High-Resolution Mass Spectrometry for Plasma Analysis. Anal. Chem. 2019, 91, 1295–1301. [Google Scholar] [CrossRef]
- Sugimoto, M.; Wong, D.T.; Hirayama, A.; Soga, T.; Tomita, M. Capillary Electrophoresis Mass Spectrometry-Based Saliva Metabolomics Identified Oral, Breast and Pancreatic Cancer-Specific Profiles. Metabolomics 2010, 6, 78–95. [Google Scholar] [CrossRef]
- Ohashi, Y.; Hirayama, A.; Ishikawa, T.; Nakamura, S.; Shimizu, K.; Ueno, Y.; Tomita, M.; Soga, T. Depiction of Metabolome Changes in Histidine-Starved Escherichia Coli by CE-TOFMS. Mol. Biosyst. 2008, 4, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Ooga, T.; Sato, H.; Nagashima, A.; Sasaki, K.; Tomita, M.; Soga, T.; Ohashi, Y. Metabolomic Anatomy of an Animal Model Revealing Homeostatic Imbalances in Dyslipidaemia. Mol. Biosyst. 2011, 7, 1217–1223. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H. PLS-ROG: Partial Least Squares with Rank Order of Groups. J. Chemom. 2017, 31, e2883. [Google Scholar] [CrossRef]
- Ogawa, A.; Nagiri, C.; Shihoya, W.; Inoue, A.; Kawakami, K.; Hiratsuka, S.; Aoki, J.; Ito, Y.; Suzuki, T.; Suzuki, T.; et al. N6-Methyladenosine (M6A) Is an Endogenous A3 Adenosine Receptor Ligand. Mol. Cell 2021, 81, 659–674.e7. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Hua, X.; Li, Q.; Zhou, Q.; Chen, J. M6A Regulator-Mediated Methylation Modification Patterns and Characteristics of Immunity in Blood Leukocytes of COVID-19 Patients. Front. Immunol. 2021, 12, 774776. [Google Scholar] [CrossRef]
- Qing, X.; Chen, Q.; Wang, K. M6A Regulator-Mediated Methylation Modification Patterns and Characteristics in COVID-19 Patients. Front. Public Health 2022, 10, 914193. [Google Scholar] [CrossRef]
- Faraj, R.; Liang, Y.; Feng, A.; Wu, J.; Black, S.M.; Wang, T. Exploring M6A-RNA Methylation as a Potential Therapeutic Strategy for Acute Lung Injury and Acute Respiratory Distress Syndrome. Pulm. Circ. 2023, 13, e12230. [Google Scholar] [CrossRef]
- Qian, W.; Zhou, J.; Shou, S. Exploration of M6A Methylation Regulators as Epigenetic Targets for Immunotherapy in Advanced Sepsis. BMC Bioinform. 2023, 24, 257. [Google Scholar] [CrossRef]
- Krishnan, S.; Nordqvist, H.; Ambikan, A.T.; Gupta, S.; Sperk, M.; Svensson-Akusjärvi, S.; Mikaeloff, F.; Benfeitas, R.; Saccon, E.; Ponnan, S.M.; et al. Metabolic Perturbation Associated With COVID-19 Disease Severity and SARS-CoV-2 Replication. Mol. Cell. Proteom. 2021, 20, 100159. [Google Scholar] [CrossRef]
- Chadda, K.R.; Blakey, E.E.; Davies, T.W.; Puthucheary, Z. Risk Factors, Biomarkers, and Mechanisms for Persistent Inflammation, Immunosuppression, and Catabolism Syndrome (PICS): A Systematic Review and Meta-Analysis. Br. J. Anaesth. 2024, 133, 538–549. [Google Scholar] [CrossRef]
- Leisman, D.E.; Mehta, A.; Thompson, B.T.; Charland, N.C.; Gonye, A.L.K.; Gushterova, I.; Kays, K.R.; Khanna, H.K.; LaSalle, T.J.; Lavin-Parsons, K.M.; et al. Alveolar, Endothelial, and Organ Injury Marker Dynamics in Severe COVID-19. Am. J. Respir. Crit. Care Med. 2022, 205, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Iwegbulem, O.; Wang, J.; Pfirrmann, R.W.; Redmond, H.P. The Role of Taurine Derivatives in the Putative Therapy of COVID-19-Induced Inflammation. Ir. J. Med. Sci. 2022, 191, 485–486. [Google Scholar] [CrossRef]
- Rubio-Casillas, A.; Gupta, R.C.; Redwan, E.M.; Uversky, V.N.; Badierah, R.A. Early Taurine Administration as a Means for Halting the Cytokine Storm Progression in COVID-19 Patients. Explor. Med. 2022, 3, 234. [Google Scholar] [CrossRef]
- van Eijk, L.E.; Offringa, A.K.; Bernal, M.-E.; Bourgonje, A.R.; van Goor, H.; Hillebrands, J.-L. The Disease-Modifying Role of Taurine and Its Therapeutic Potential in Coronavirus Disease 2019 (COVID-19). Adv. Exp. Med. Biol. 2022, 1370, 3–21. [Google Scholar]
- Roşca, A.E.; Vlădăreanu, A.-M.; Mirica, R.; Anghel-Timaru, C.-M.; Mititelu, A.; Popescu, B.O.; Căruntu, C.; Voiculescu, S.E.; Gologan, Ş.; Onisâi, M.; et al. Taurine and Its Derivatives: Analysis of the Inhibitory Effect on Platelet Function and Their Antithrombotic Potential. J. Clin. Med. Res. 2022, 11, 666. [Google Scholar] [CrossRef] [PubMed]
- Sezer, S.; Bal, C.; Kalem, A.K.; Kayaaslan, B.; Eser, F.; Hasanoglu, İ.; Akıncı, E.; Güner, R.; Erel, Ö.; Yılmaz, G. COVID-19 Patients with Altered Steroid Hormone Levels Are More Likely to Have Higher Disease Severity. Endocrine 2022, 78, 373–379. [Google Scholar] [CrossRef]
- Richard, V.R.; Gaither, C.; Popp, R.; Chaplygina, D.; Brzhozovskiy, A.; Kononikhin, A.; Mohammed, Y.; Zahedi, R.P.; Nikolaev, E.N.; Borchers, C.H. Early Prediction of COVID-19 Patient Survival by Targeted Plasma Multi-Omics and Machine Learning. Mol. Cell. Proteom. 2022, 21, 100277. [Google Scholar] [CrossRef] [PubMed]
- Cappelletti, G.; Carsana, E.V.; Lunghi, G.; Breviario, S.; Vanetti, C.; Di Fonzo, A.B.; Frattini, E.; Magni, M.; Zecchini, S.; Clerici, M.; et al. SARS-CoV-2 Hampers Dopamine Production in IPSC-Derived Dopaminergic Neurons. Exp. Mol. Pathol. 2023, 134, 104874. [Google Scholar] [CrossRef]
- Rasmi, Y.; Shokati, A.; Hatamkhani, S.; Farnamian, Y.; Naderi, R.; Jalali, L. Assessment of the Relationship between the Dopaminergic Pathway and Severe Acute Respiratory Syndrome Coronavirus 2 Infection, with Related Neuropathological Features, and Potential Therapeutic Approaches in COVID-19 Infection. Rev. Med. Virol. 2024, 34, e2506. [Google Scholar] [CrossRef]
- Yang, L.; Kim, T.W.; Han, Y.; Nair, M.S.; Harschnitz, O.; Zhu, J.; Wang, P.; Koo, S.Y.; Lacko, L.A.; Chandar, V.; et al. SARS-CoV-2 Infection Causes Dopaminergic Neuron Senescence. Cell Stem Cell 2024, 31, 196–211.e6. [Google Scholar] [CrossRef]
- Garcia, C.; Au Duong, J.; Poëtte, M.; Ribes, A.; Payre, B.; Mémier, V.; Sié, P.; Minville, V.; Voisin, S.; Payrastre, B.; et al. Platelet Activation and Partial Desensitization Are Associated with Viral Xenophagy in Patients with Severe COVID-19. Blood Adv. 2022, 6, 3884–3898. [Google Scholar] [CrossRef] [PubMed]
- De Simone, M.; Choucha, A.; Ciaglia, E.; Conti, V.; Pecoraro, G.; Santurro, A.; Puca, A.A.; Cascella, M.; Iaconetta, G. Discogenic Low Back Pain: Anatomic and Pathophysiologic Characterization, Clinical Evaluation, Biomarkers, AI, and Treatment Options. J. Clin. Med. 2024, 13, 5915. [Google Scholar] [CrossRef] [PubMed]
- De Simone, M.; De Feo, R.; Choucha, A.; Ciaglia, E.; Fezeu, F. Enhancing Sleep Quality: Assessing the Efficacy of a Fixed Combination of Linden, Hawthorn, Vitamin B1, and Melatonin. Med. Sci. 2023, 12, 2. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, E.; Banoei, M.M.; Kaur, J.; Lee, C.H.; Winston, B.W. Metabolomic Insights into COVID-19 Severity: A Scoping Review. Metabolites 2024, 14, 617. [Google Scholar] [CrossRef]
- Almulla, A.F.; Supasitthumrong, T.; Tunvirachaisakul, C.; Algon, A.A.A.; Al-Hakeim, H.K.; Maes, M. The Tryptophan Catabolite or Kynurenine Pathway in COVID-19 and Critical COVID-19: A Systematic Review and Meta-Analysis. BMC Infect. Dis. 2022, 22, 615. [Google Scholar] [CrossRef]
- Herrera-Van Oostdam, A.S.; Castañeda-Delgado, J.E.; Oropeza-Valdez, J.J.; Borrego, J.C.; Monárrez-Espino, J.; Zheng, J.; Mandal, R.; Zhang, L.; Soto-Guzmán, E.; Fernández-Ruiz, J.C.; et al. Immunometabolic Signatures Predict Risk of Progression to Sepsis in COVID-19. PLoS ONE 2021, 16, e0256784. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Overall n = 118 | Survivor n = 93 | Non-Survivor n = 25 |
|---|---|---|---|
| Age (years) | 61 (53, 73) | 58 (52, 71) | 71 (63, 78) |
| Female | 22 (19%) | 17 (18%) | 5 (20%) |
| BMI (kg/m2) | 24.6 (22.5,28.2) | 24.7 (22.5,28.2) | 24.0 (23.2,27.6) |
| SARS-CoV-2 Delta variant | 25 (21%) | 20 (22%) | 5 (20%) |
| Onset to intubation interval (days) | 9.1 (6.6, 11.6) | 9.6 (7.6, 12.6) | 7.6 (5.6, 10.6) |
| Comorbidity | |||
| Hypertension | 63 (53%) | 47 (51%) | 16 (64%) |
| Diabetes mellitus | 39 (33%) | 27 (29%) | 12 (48%) |
| Ischemic heart disease | 9 (8%) | 3 (3%) | 6 (24%) |
| Congestive heart failure | 6 (5%) | 2 (2%) | 4 (16%) |
| Cerebrovascular disease | 10 (8%) | 8 (9%) | 2 (8%) |
| Hemodialysis | 4 (3%) | 1 (1%) | 3 (12%) |
| Dyslipidemia | 27 (23%) | 21 (23%) | 6 (24%) |
| Asthma | 7 (6%) | 6 (6%) | 1 (4%) |
| COPD | 5 (4%) | 4(4%) | 1 (4%) |
| Interstitial pneumonia | 2 (2%) | 0 (0%) | 2 (8%) |
| Lymphoma | 1 (1%) | 1 (1%) | 0 (0%) |
| Cancer | 6 (5%) | 3 (3%) | 3 (12%) |
| HIV | 1 (1%) | 1 (1%) | 0 (0%) |
| Immunocompromised | 8 (7%) | 4 (4%) | 4 (16%) |
| Severity | |||
| APACHE II | 15 (10, 20) | 14 (10, 20) | 19 (15, 23) |
| SOFA score | 6 (4, 8) | 5 (3, 7) | 8 (6, 10) |
| Treatment | |||
| NO therapy | 56 (47%) | 36 (39%) | 20 (80%) |
| Prone positioning | 71 (60%) | 50 (54%) | 21 (84%) |
| Steroid | 99 (84%) | 76 (82%) | 23 (92%) |
| Favipiravir | 43 (36%) | 33 (35%) | 10 (40%) |
| Remdesivir | 84 (71%) | 72 (77%) | 12 (48%) |
| Tocilizumab | 23 (19%) | 20 (22%) | 3 (12%) |
| Baricitinib | 21 (18%) | 18 (19%) | 3 (12%) |
| mPSL pulse | 32 (27%) | 23 (25%) | 9 (36%) |
| ECMO | 14 (12%) | 7 (8%) | 7 (28%) |
| CRRT | 23 (19%) | 5 (5%) | 18 (72%) |
| Complication | |||
| Arrythmia | 24 (20%) | 14 (15%) | 10 (40%) |
| DVT or PE | 6 (5%) | 3 (3%) | 3 (12%) |
| Mediastinal emphysema | 4 (3%) | 1 (1%) | 3 (12%) |
| Pneumothorax | 6 (5%) | 3 (3%) | 3 (12%) |
| VAP | 40 (34%) | 25 (27%) | 15 (60%) |
| BSI | 33 (28%) | 18 (19%) | 15 (60%) |
| Clinical Outcome | |||
| Intubation to ICU discharge interval (days) | 12 (7, 21) | 10 (7, 15) | 27 (19, 40) |
| Discharge over 14 days | 43 (36%) | 24 (26%) | 19 (76%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uchimido, R.; Kami, K.; Yamamoto, H.; Yokoe, R.; Tsuchiya, I.; Nukui, Y.; Goto, Y.; Hanafusa, M.; Fujiwara, T.; Wakabayashi, K. Longitudinal Metabolomics Reveals Metabolic Dysregulation Dynamics in Patients with Severe COVID-19. Metabolites 2024, 14, 656. https://doi.org/10.3390/metabo14120656
Uchimido R, Kami K, Yamamoto H, Yokoe R, Tsuchiya I, Nukui Y, Goto Y, Hanafusa M, Fujiwara T, Wakabayashi K. Longitudinal Metabolomics Reveals Metabolic Dysregulation Dynamics in Patients with Severe COVID-19. Metabolites. 2024; 14(12):656. https://doi.org/10.3390/metabo14120656
Chicago/Turabian StyleUchimido, Ryo, Kenjiro Kami, Hiroyuki Yamamoto, Ryo Yokoe, Issei Tsuchiya, Yoko Nukui, Yuki Goto, Mariko Hanafusa, Takeo Fujiwara, and Kenji Wakabayashi. 2024. "Longitudinal Metabolomics Reveals Metabolic Dysregulation Dynamics in Patients with Severe COVID-19" Metabolites 14, no. 12: 656. https://doi.org/10.3390/metabo14120656
APA StyleUchimido, R., Kami, K., Yamamoto, H., Yokoe, R., Tsuchiya, I., Nukui, Y., Goto, Y., Hanafusa, M., Fujiwara, T., & Wakabayashi, K. (2024). Longitudinal Metabolomics Reveals Metabolic Dysregulation Dynamics in Patients with Severe COVID-19. Metabolites, 14(12), 656. https://doi.org/10.3390/metabo14120656