
A New Advancement in Germination Biotechnology of Purple Creole Corn: Bioactive Compounds and In Situ Enzyme Activity for Water-Soluble Extract and Pan Bread

 , , ,
, , , 

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Experimental Desing
2.2.2. Germination Process
2.2.3. Radicle Length and Seed Vigor
2.2.4. Analysis of Flours from Germinated and Non-Germinated Seeds
Total Soluble Phenolic Compounds (TSPCs)
γ-Aminobutyric Acid (GABA)
2.2.5. Obtaining Water-Soluble Extracts
2.2.6. Characterization of Water-Soluble Extracts
Soluble Protein
Soluble Solids
Reducing Sugars
2.2.7. Numerical Optimization and Mathematical Models’ Validation for Germination Process and Water-Soluble Extracts
Antioxidant Capacity
2.2.8. Breadmaking Process
2.2.9. Dough Volume Increase
2.2.10. Technological Aspects of the Bread
Specific Volume
Crumb Instrumental Color
Instrumental Texture
2.2.11. Numerical Optimization of Breadmaking
2.2.12. Statistical Analysis
3. Results and Discussion
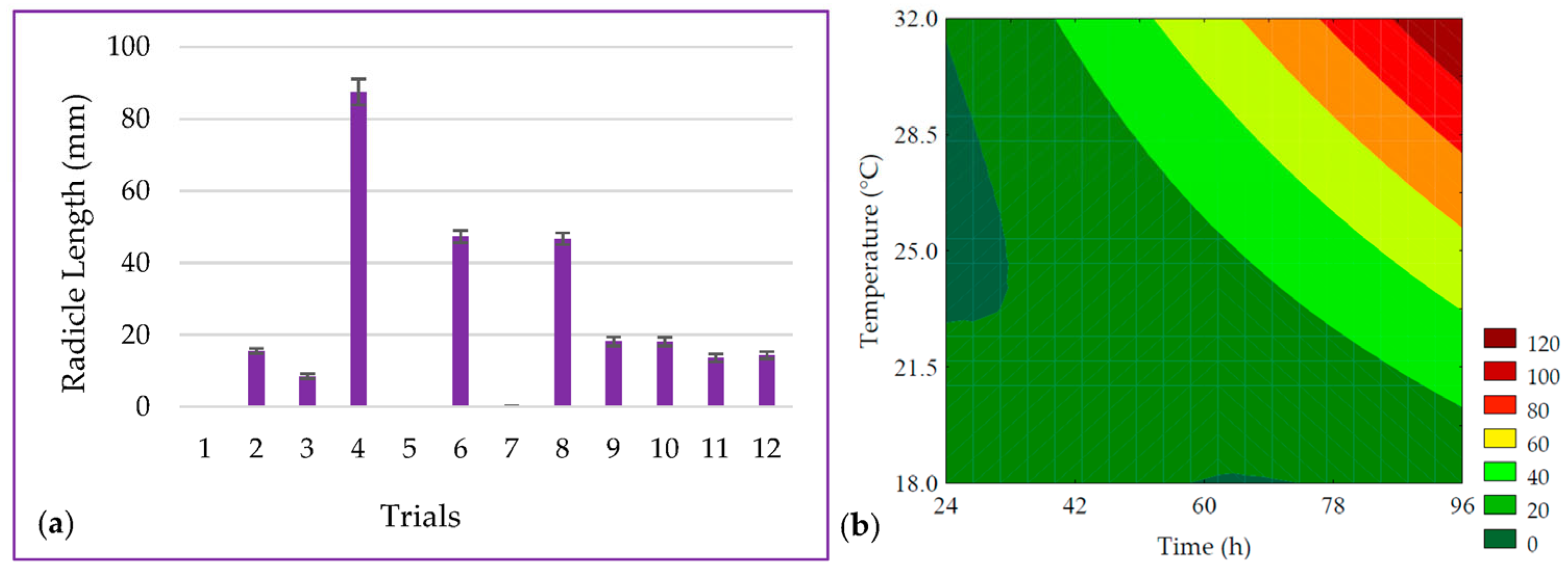
3.1. Radicle Lenght and Seed Vigor
3.2. Analysis of Germinated and Non-Germinated Corn Flours
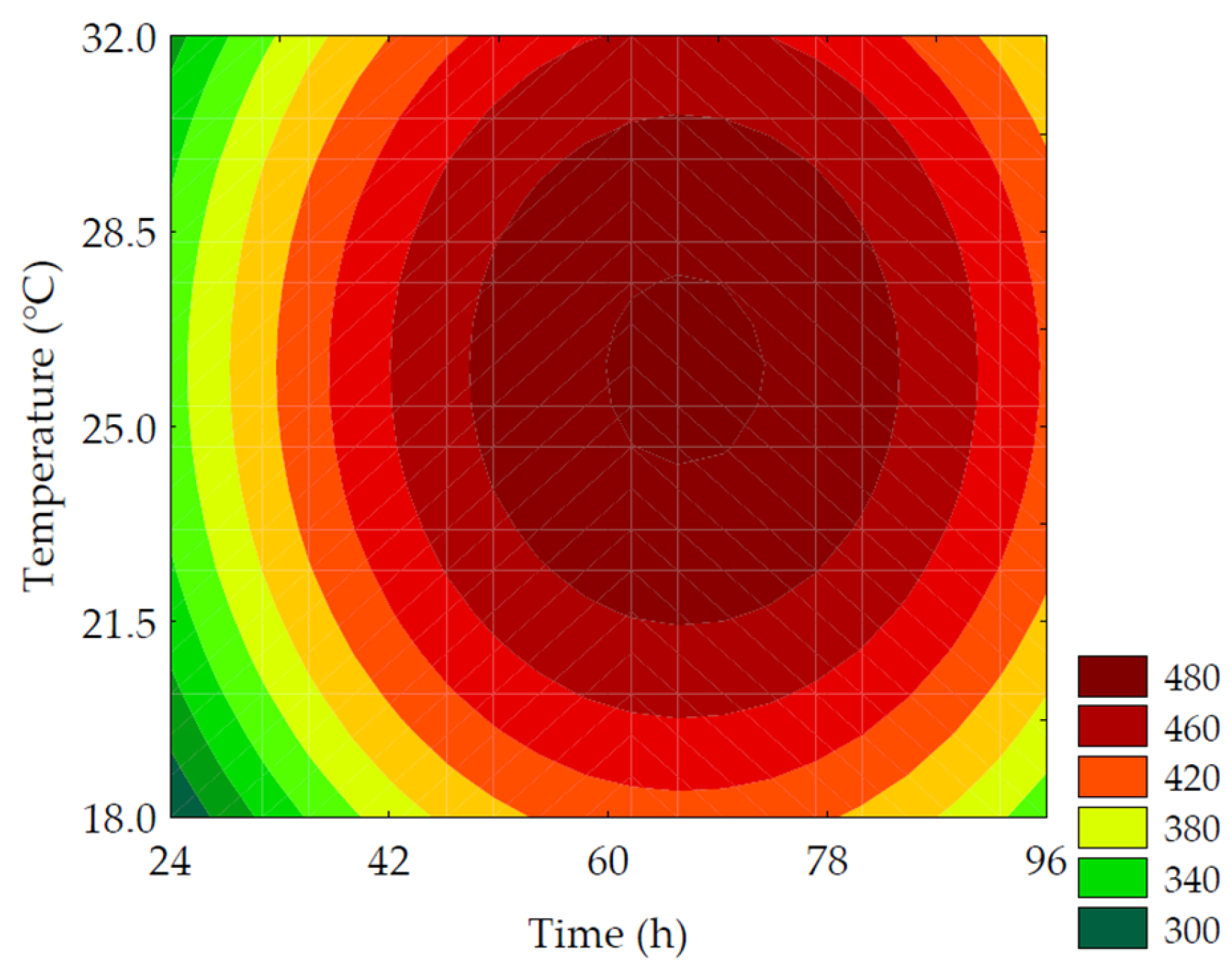
3.2.1. Total Soluble Phenolic Compounds
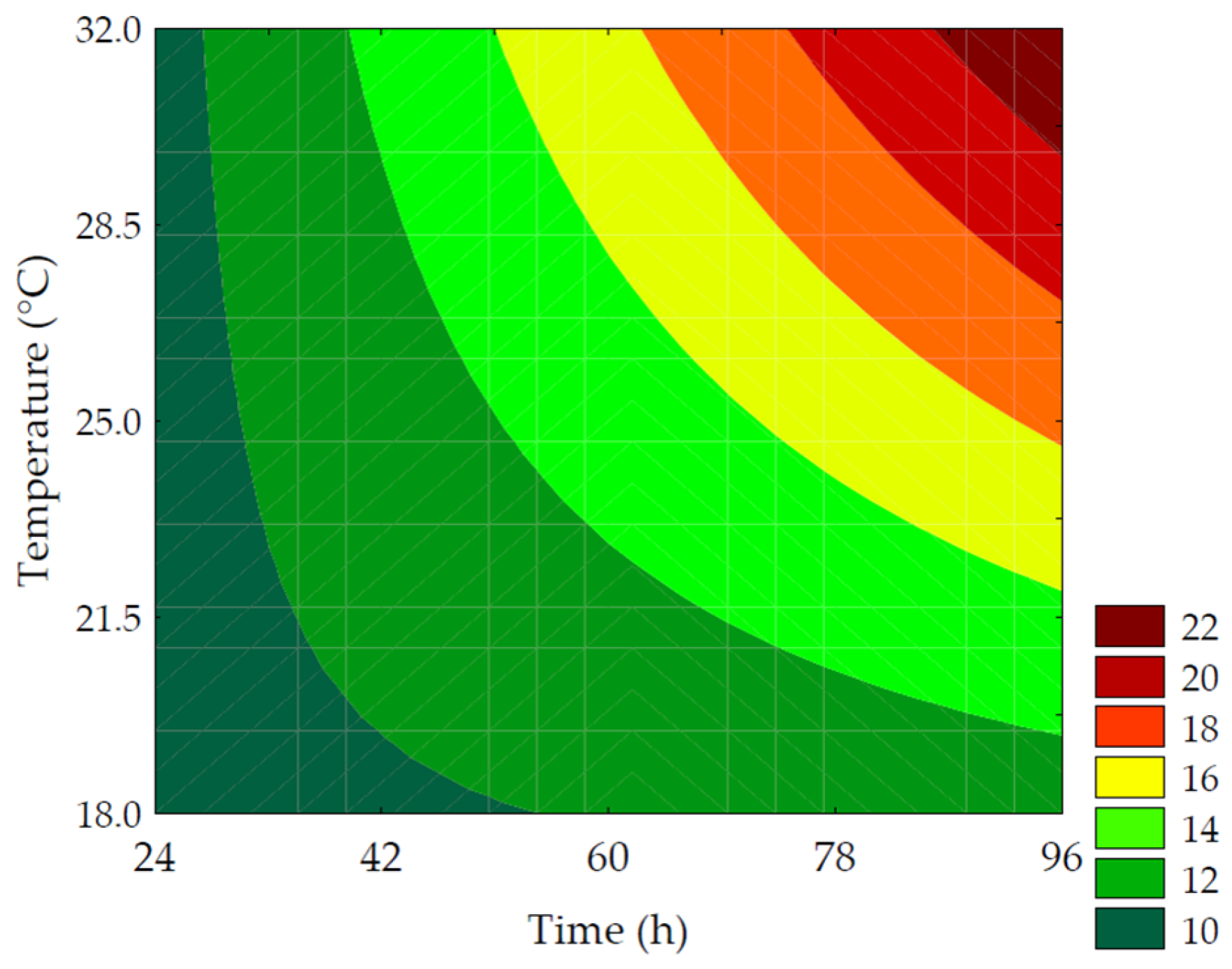
3.2.2. γ-Aminobutyric Acid (GABA)
3.3. Analysis of Water-Soluble Extracts
3.3.1. Soluble Protein
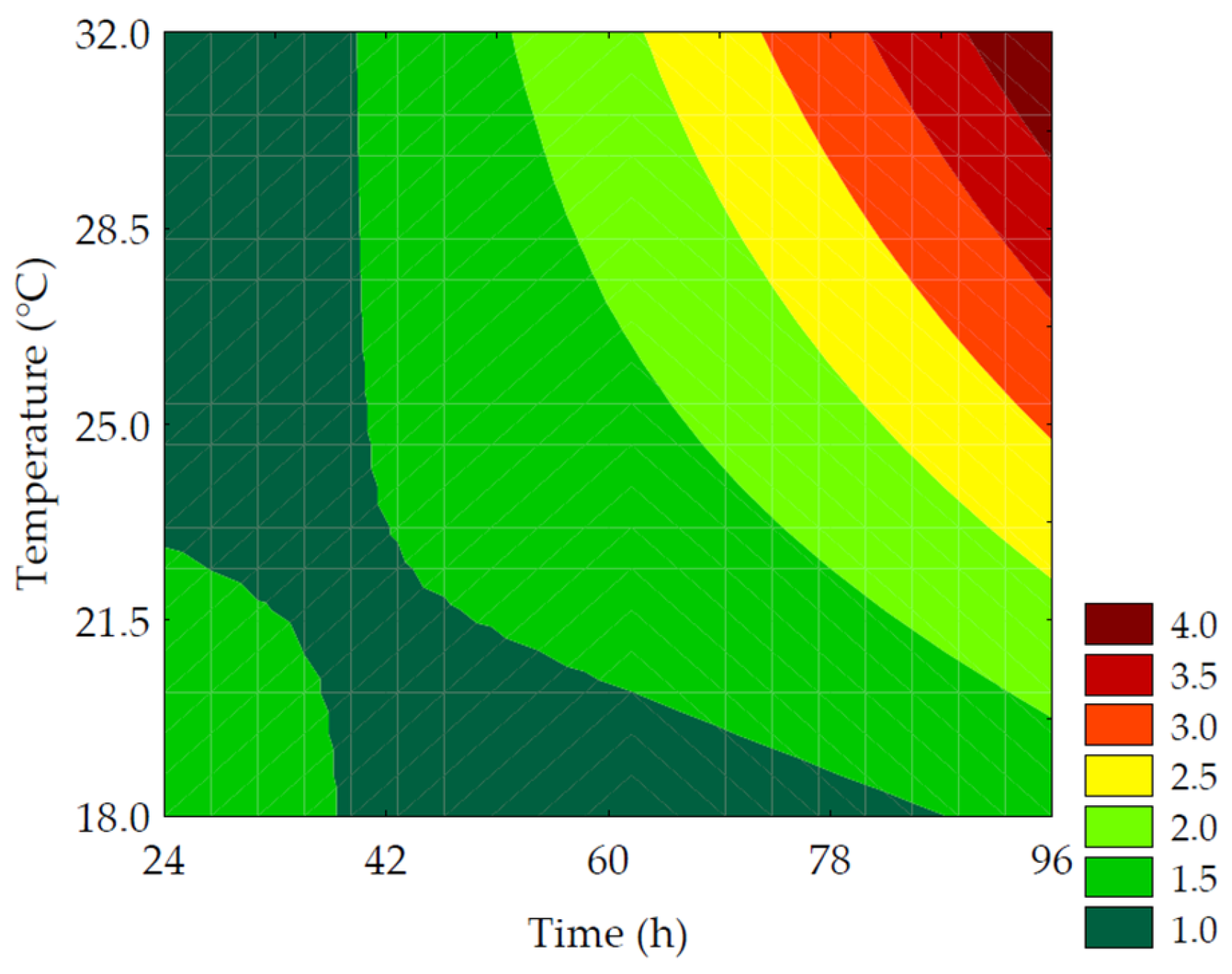
3.3.2. Soluble Solids
3.3.3. Reducing Sugar
3.4. Numerical Optimization and Validation of Mathematical Models
Antioxidant Capacity
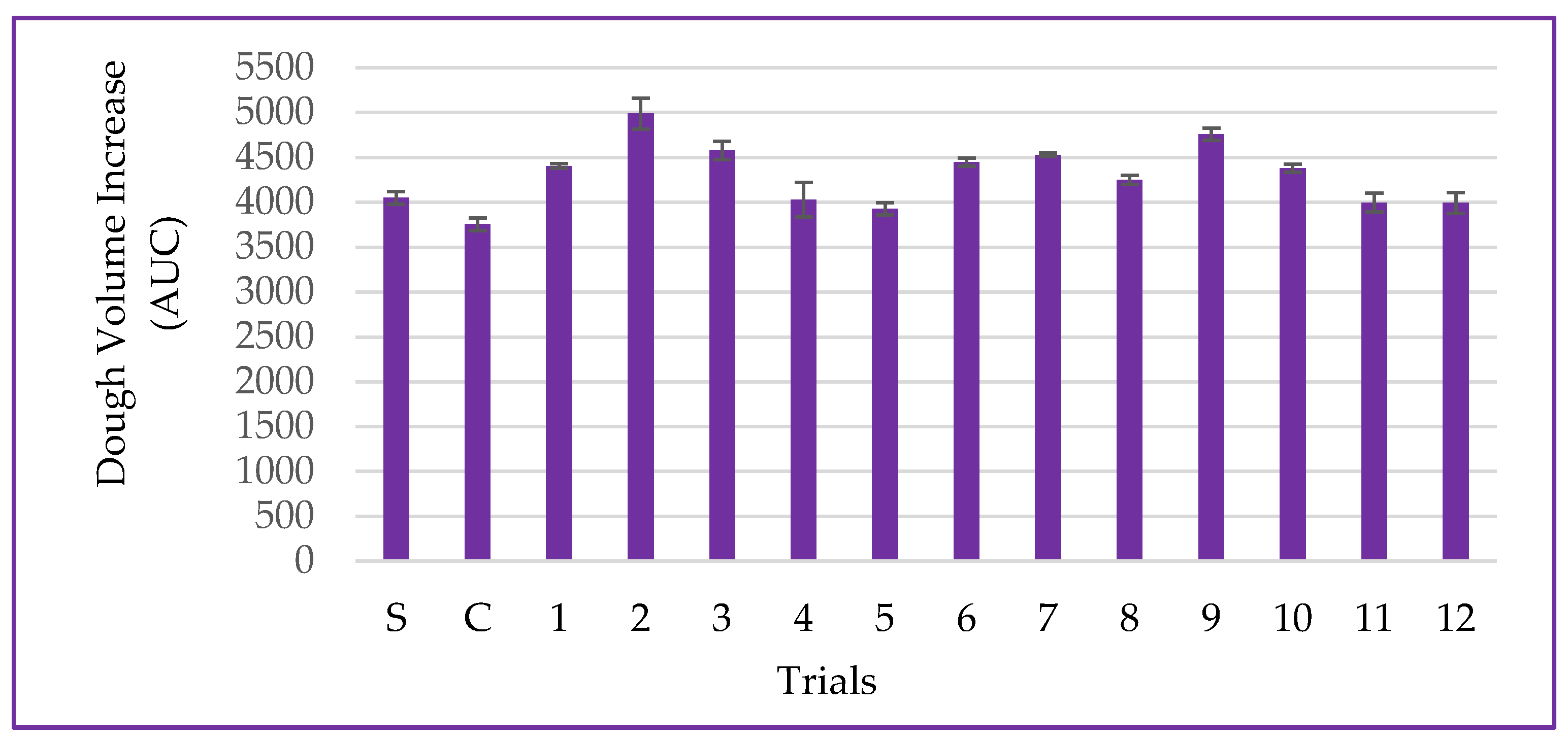
3.5. Dough Volume Increase
3.6. Pan Bread Characterization
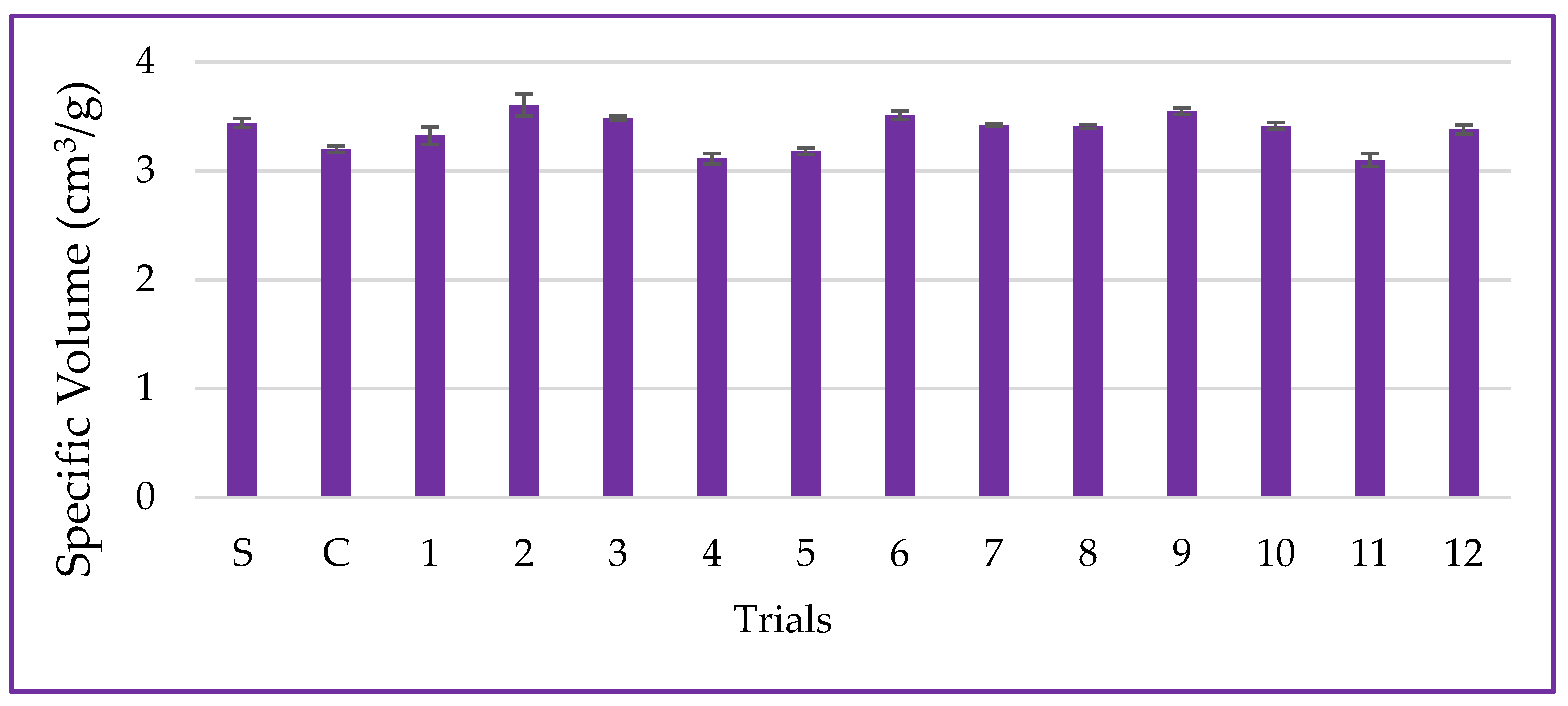
3.6.1. Specific Volume
3.6.2. Instrumental Color
3.6.3. Instrumental Texture
3.7. Numerical Optimization
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laskowski, W.; Górska-Warsewicz, H.; Rejman, K.; Czeczotko, M.; Zwolińska, J. How Important are Cereals and Cereal Products in the Average Polish Diet? Nutrients 2019, 11, 679. [Google Scholar] [CrossRef] [PubMed]
- Hadidi, M.; Garcia, S.R.; Ziogkas, D.; McClements, D.J.; Moreno, A. Cereal bran proteins: Recent advances in extraction, properties, and applications. Crit. Rev. Food Sci. Nutr. 2023, 1–25. [Google Scholar] [CrossRef]
- Paucar-Menacho, L.M.; Castillo-Martínez, W.E.; Simpalo-Lopez, W.D.; Verona-Ruiz, A.; Lavado-Cruz, A.; Martínez-Villaluenga, C.; Peñas, E.; Frias, J.; Schmiele, M. Performance of thermoplastic extrusion, germination, fermentation, and hydrolysis techniques on phenolic compounds in cereals and pseudocereals. Foods 2022, 11, 1957. [Google Scholar] [CrossRef] [PubMed]
- Andressa, I.; Nascimento, G.K.S.; Santos, T.M.; Oliveira, J.R.; Benassi, V.M.; Schmiele, M. Physicochemical changes induced by the sprouting process of purple pericap corn kernels. Braz. J. Dev. 2023, 9, 8051–8070. [Google Scholar] [CrossRef]
- Carrera-Castaño, G.; Calleja-Cabrera, J.; Pernas, M.; Gómez, L.; Oñate-Sánchez, L. An Updated Overview on the Regulation of Seed Germination. Plants 2020, 9, 703. [Google Scholar] [CrossRef]
- Rasera, G.B.; Castro, R.J.S. Grain germination: A systematic review of the biochemical processes involved in the content and profile of phenolic compounds and their antioxidant properties. Braz. J. Natural Sci. 2020, 3, 287–300. [Google Scholar] [CrossRef]
- Lima, C.T.; Santos, T.M.; Neves, N.A.; Lavado-Cruz, A.; Paucar-Menacho, L.M.; Clerici, M.T.P.S.; Meza, S.L.R.; Schmiele, M. New breakfast cereal developed with sprouted whole ryegrass flour: Evaluation of technological and nutritional parameters. Foods 2023, 12, 3902. [Google Scholar] [CrossRef] [PubMed]
- Naumenko, N.; Fatkullin, R.; Popova, N.; Ruskina, A.; Kalinina, I.; Morozov, R.; Avdin, V.V.; Antonova, A.; Vasileva, E. Effect of a combination of ultrasonic germination and fermentation processes on the antioxidant activity and γ-aminobutyric acid content of food ingredients. Fermentation 2023, 9, 246. [Google Scholar] [CrossRef]
- Thirumurugan, D.; Cholarajan, A.; Raja, S.S.S.; Vijayakumar, R. An Introductory Chapter: Secondary Metabolites. In Secondary Metabolites: Sources and Applications; Vijayakumar, R., Raja, S.S.S., Eds.; IntechOpen: London, UK, 2018; pp. 3–21. [Google Scholar]
- Salah, A.; Zhan, M.; Cao, C.; Han, Y.; Ling, L.; Liu, Z.; Li, P.; Ye, M.; Jiang, Y. γ-aminobutyric acid promotes chloroplast ultrastructure, antioxidant capacity, and growth of waterlogged maize seedlings. Sci. Rep. 2019, 9, 484. [Google Scholar] [CrossRef]
- Li, L.; Dou, N.; Zhang, H.; Wu, C. The versatile GABA in plants. Plant Signaling Behav. 2021, 16, 1862565. [Google Scholar] [CrossRef]
- Zhang, L.; Shi, T.; Xie, M.; Yu, M.; Hao, N.; Li, W.; Zhang, B. Effect of ultrasonic induction on the main physiological and biochemical indicators and γ-aminobutyric acid content of maize during germination. Foods 2022, 11, 1358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, S.; Simpson, B.K. Enzymes in food bioprocessing—novel food enzymes, applications, and related techniques. Curr. Opin. Food Sci. 2018, 19, 30–35. [Google Scholar] [CrossRef]
- Rosell, C.M.; Dura, A. Enzymes in bakeries. In Enzymes in Food and Beverage Processing, 1st ed.; Chandrasekaran, M., Ed.; CRC Press: Boca Raton, FL, USA, 2015; pp. 171–204. [Google Scholar]
- Nascimento, G.K.S.; Rocha, L.O.F.; Schmiele, M. Potencial of enhancing agents in replacing chemical additives in bread processing. In The Food Industry: Perceptions, Practices and Future Prospects; Santos, D.T., Castilho-Torres, R.A., Carvalho, G.B.M., Eds.; Nova Science Publishers: New York, NY, USA, 2021; pp. 13–43. [Google Scholar]
- Rodrigues, M.I.; Iemma, A.F. Experiment Planning and Process Optimization: A Sequential Planning Strategy, 3rd ed.; Casa do Pão: Campinas, SP, Brazil, 2014; p. 100. [Google Scholar]
- Nascimento, G.K.S.; Neves, N.A.; Silva, M.S.; Gomes, L.; Lima, C.; Schmiele, M. Use of low-frequency ultrasound and optimization of the extracting solution through white chemistry for total soluble phenolic compounds of purple pericarp creole maize. In Proceedings of the XV Latin American Symposium on Food Science and Nutrition-15 SLACAN–The Food and Nutrition Science Revolution: Feeding the World Sustainably, Campinas, Brazil, 12–14 November 2023. [Google Scholar]
- Filho, W.G.V. Bebidas Alcoólicas: Ciência e Tecnologia, 1st ed.; Blucher: São Paulo, Brazil, 2010; pp. 29–30. [Google Scholar]
- AACCI. Approved Methods of AACCI, 11th ed.; Methods 46-13.01, 80-51.01, 10-05.01 and 74-09.01; The American Association of Cereal Chemists International: St. Paul, MN, USA, 2010. [Google Scholar]
- Santos, A.A.; Deoti, J.R.; Müller, G.; Dário, M.G.; Stambuk, B.U.; Alves, S.L.J. Microwell plate-based method for the determination of reducing sugars with the DNS reagent. Braz. J. Food Technol. 2017, 20, 2015113. [Google Scholar] [CrossRef]
- Derringer, G.; Suich, R. Simultaneous optimization of several response variables. J. Qual. Technol. 1980, 12, 214–219. [Google Scholar] [CrossRef]
- Dávalos, A.; Gómez-Cordovés, C.; Bartolomé, B. Extending applicability of the oxygen radical absorbance capacity (ORAC-fluorescein) assay. J. Agric. Food Chem. 2004, 52, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Camponogara, J.A.; Farias, C.A.A.; Moraes, D.P.; Bettio, L.; Citadin, I.; Mallman, C.A.; Schmiele, M.; Ballus, C.A.; Barin, J.S.; Barcia, M.T. Optimization of Microwave Hydrodiffusion and Gravity (MHG) for Pre-treatment of Dehydration and Obtaining a Jaboticaba Extract. Food Bioprocess Technol. 2023, 16, 03202. [Google Scholar] [CrossRef]
- Nascimento, G.K.S.; dos Santos, T.M.; Andressa, I.; Rodrigues, S.M.; da Silva, L.E.P.; Benassi, V.M.; Schmiele, M. Applicability of corn cob flour and the cocktail rich in xylanase produced by the filamentous fungus Fusarium sp. EA 1.3.1 in bread. Braz. J. Dev. 2023, 9, 8031–8050. [Google Scholar] [CrossRef]
- De Oliveira Teotônio, D.; da Costa, B.A.F.; Gomes, P.T.G.; Santos, M.P.; Amaral, E.F.G.; Clerici, M.T.P.S.; Leoro, M.G.V.; Schmiele, M. Fructo-oligosaccharides, hydrolyzed soy protein and yeast (Saccharomyces sp.) extract as potential cryoprotectans in gluten-free frozen dough and bread quality. Res. Soc. Dev. 2021, 10, 44510313556. [Google Scholar] [CrossRef]
- Silva, J.D.R.; Rosa, G.C.; Neves, N.A.; Leoro, M.G.V.; Schmiele, M. Production of sourdough and gluten-free bread with brown rice and carioca and cowpea beans flours: Biochemical, nutritional and structural characteristics. Res. Soc. Dev. 2021, 10, 303101623992. [Google Scholar] [CrossRef]
- Sharanagat, V.S.; Nema, P.K. Bread preparation by partial replacement of wheat by germinated sorghum. Food Sci. Technol. Int. 2023, 29, 13–24. [Google Scholar] [CrossRef]
- Wang, Y.; Peng, Y.; Qiao, X.; Zhuang, Q. Discriminant analysis and comparison of corn seed vigor based on multiband spectrum. Comput. Electron. Agric. 2021, 190, 106444. [Google Scholar] [CrossRef]
- Nieves-Hernández, M.G.; Correa-Piña, B.L.; Esquivel-Fajardo, E.A.; Barrón-García, O.Y.; Gaytán-Martínez, M.; Rodriguez-Garcia, M.E. Study of morphological, structural, pasting, thermal, and vibrational changes in maize and isolated maize starch during germination. J. Cereal Sci. 2023, 111, 103685. [Google Scholar] [CrossRef]
- Reed, R.C.; Bradford, K.J.; Khanday, I. Seed germination and vigor: Ensuring crop sustainability in a changing climate. Heredity 2022, 128, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Chavarín-Martínez, C.D.; Gutiérrez-Dorado, R.; Perales-Sánchez, J.X.K.; Cuevas-Rodríguez, E.O.; Milán-Carrillo, J.; Reyes-Moreno, C. Germination in optimal conditions as effective strategy to improve nutritional and nutraceutical value of underutilized mexican blue maize seeds. Plant Food Hum. Nutr. 2019, 74, 192–199. [Google Scholar] [CrossRef]
- Costa, C.A.R.; Machado, G.G.L.; Rodrigues, L.J.; Barros, H.E.A.; Natarelli, C.V.L.; Vilas Boas, E.V.B. Phenolic compounds profile and antioxidant activity of purple passion fruit’s pulp, peel and seed at different maturation stages. Sci. Hortic. 2023, 321, 112244. [Google Scholar] [CrossRef]
- Paucar-Menacho, L.M.; Schmiele, M.; Lavado-Cruz, A.A.; Verona-Ruiz, A.L.; Mollá, C.; Peñas, E.; Frias, J.; Simpalo-Lopez, W.D.; Castillo-Martínez, W.E.; Martínez-Villaluenga, C. Andean sprouted pseudocereals to produce healthier extrudates: Impact in nutritional and physicochemical properties. Foods 2022, 11, 3259. [Google Scholar] [CrossRef]
- Ukpong, E.S.; Onyeka, E.U.; Oladeji, B.S. Bioactive compounds, nutrients and pasting properties of parboiled milled rice, brown rice and germinated brown rice of selected cultivars and the effects of germination durations. Food Chem. Adv. 2023, 2, 100234. [Google Scholar] [CrossRef]
- Millar, K.A.; Barry-Ryan, C.; Burke, R.; McCarthy, S.; Gallagher, E. Dough properties and baking characteristics of white bread, as affected by addition of raw, germinated and toasted pea flour. Innovative Food Sci. Emerg. Technol. 2019, 56, 102189. [Google Scholar] [CrossRef]
- Yang, T.; Wu, X.; Wang, W.; Wu, Y. Regulation of seed storage protein synthesis in monocot and dicot plants: A comparative review. Mol. Plant 2023, 16, 145–167. [Google Scholar] [CrossRef]
- del R. M. Concha, D.; Martínez, J.E.B.; Velázquez, T.G.G.; Martínez, C.J.; Ruiz, J.C.R. Impact of germination time on protein solubility and anti-inflammatory properties of Pisum sativum L. grains. Food Chem. X 2022, 13, 100219. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, C.; Qu, M.; Pan, K.; Ouyang, K.; Song, X.; Zhao, X. Characterization of a recombinant zein-degrading protease from Zea mays by Pichia pastoris and its effects on enzymatic hydrolysis of corn starch. Int. J. Biol. Macromol. 2020, 164, 3287–3293. [Google Scholar] [CrossRef] [PubMed]
- Baranzelli, J.; Kringel, D.H.; Colussi, R.; Paiva, F.F.; Aranha, B.C.; Miranda, M.Z.; Zavareze, E.R.; Dias, A.R.G. Changes in enzymatic activity, technological quality and gamma-aminobutyric acid (GABA) content of wheat flour as affected by germination. Food Sci. Technol. 2018, 90, 483–490. [Google Scholar] [CrossRef]
- Almeida, F.S.; Silva, C.A.A.; Lima, S.M.; Suarez, Y.S.; Cunha Andrade, L.H.C. Use of Fourier transform infrared spectroscopy to monitor sugars in the beer mashing process. Food Chem. 2018, 263, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Tao, Y.; Han, Y.; Ding, Y.; Jian, X.; Li, D. Preparation of germinated brown rice with high γ-aminobutyric acid content and short root by magnetic field treatment. J. Cereal Sci. 2023, 112, 103720. [Google Scholar] [CrossRef]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.B. Fermentation and germination improve nutritional value of cereals and legumes through activation of endogenous enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef] [PubMed]
- Paucar-Menacho, L.M.; Martínez-Villaluenga, C.; Dueñas, M.; Frias, J.; Peñas, E. Optimization of germination time and temperature to maximize the content of bioactive compounds and the antioxidant activity of purple corn (Zea mays L.) by response surface methodology. Food Sci.Technol. 2017, 76, 236–244. [Google Scholar] [CrossRef]
- Xiang, J.; Yuan, Y.; Du, L.; Zhang, Y.; Li, C.; Beta, T. Modification on phenolic profiles and enhancement of antioxidant activity of proso millets during germination. Food Chem X 2023, 18, 100628. [Google Scholar] [CrossRef]
- Paucar-Menacho, L.M.; Peñas, E.; Dueñas, M.; Frias, J.; Martínez-Villaluenga, C. Optimizing germination conditions to enhance the accumulation of bioactive compounds and the antioxidant activity of kiwicha (Amaranthus caudatus) using response surface methodology. Food Sci. Technol. 2017, 76, 245–252. [Google Scholar] [CrossRef]
- Ding, J.; Hu, H.; Yang, J.; Wu, T.; Sun, X.; Fang, Y.; Huang, Q. Mechanistic study of the impact of germinated brown rice flour on gluten network formation, dough properties and bread quality. Innovative Food Sci. Emerg. Technol. 2023, 83, 103217. [Google Scholar] [CrossRef]
- Monterde, V.; Janssen, F.; Wouters, A.G.B. Understanding the impact of water-extractable cereal flour constituents on network formation in gluten-starch dough. J. Cereal Sci. 2023, 114, 103797. [Google Scholar] [CrossRef]
- de Souza, E.C.; Cordeiro, D.A.; Silva, B.S.; de Andrade Neves, N.; Schmiele, M. Development of muffin with the incorporation of olive pomace flour, extra virgin olive oil and hydrolyzed soy protein. Res. Soc. Dev. 2022, 11, 58511226012. [Google Scholar] [CrossRef]
- Cauduro, T.; D’Almeida, C.T.S.; Biduski, B.; Santos, A.; Santos, M.C.B.; Lima, L.R.S.; Cameron, L.C.; Bertolin, T.E.; Ferreira, M.S.L.; Gutkoski, L.C. Whole wheat flour replaced by sprouted wheat improves phenolic compounds profile, rheological and bread-making properties. J. Cereal Sci. 2023, 114, 103778. [Google Scholar] [CrossRef]
- Neves, N.A.; Gomes, P.T.G.; Carmo, E.M.R.; Silva, B.S.; Amaral, T.N.; Schmiele, M. Sourdough and jaboticaba (Plinia cauliflora) for improvement on pan bread characteristics. Res. Soc. Dev. 2020, 11, 90691110552. [Google Scholar] [CrossRef]
- Mahajan, S.; Bhaskar, N.; Kaur, R.K.; Jain, A. A comparison of oral health status in diabetic and non-diabetic patients receiving hemodialysis-A systematic review and meta-analysis. Diabetes Metab. Synd. Clin. Res. Rev. 2021, 15, 102256. [Google Scholar] [CrossRef]
- Mokrzycki, W.; Tatol, M. Color difference Delta E–A survey. Mach. Graph. Vis. 2011, 20, 383–411. [Google Scholar]
- Liu, P.; Ma, L.; Duan, W.; Gao, W.; Fang, Y.; Guo, L.; Yuan, C.; Wu, Z.; Cui, B. Maltogenic amylase: Its structure, molecular modification, and effects on starch and starch-based products. Carbohydr. Polym. 2023, 319, 121183. [Google Scholar] [CrossRef]
- Al-Ansi, W.; Zhang, Y.; Alkawry, T.A.A.; Al-Adeeb, A.; Mahdi, A.A.; Al-Maqtari, Q.A.; Ahmed, A.; Mushtaq, B.S.; Fan, M.; Li, Y.; et al. Influence of germination on bread-making behaviors, functional and shelf-life properties, and overall quality of highland barley bread. Food Sci. Technol. 2022, 159, 113200. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trials | Encoded Levels | Real Levels | ||
|---|---|---|---|---|
| x1 | x2 | X1 (h) | X2 (°C) | |
| 1 | −1 | −1 | 34 | 20 |
| 2 | 1 | −1 | 86 | 20 |
| 3 | −1 | 1 | 34 | 30 |
| 4 | 1 | 1 | 86 | 30 |
| 5 | −α | 0 | 24 | 25 |
| 6 | α | 0 | 96 | 25 |
| 7 | 0 | −α | 60 | 18 |
| 8 | 0 | α | 60 | 32 |
| 9 | 0 | 0 | 60 | 25 |
| 10 | 0 | 0 | 60 | 25 |
| 11 | 0 | 0 | 60 | 25 |
| 12 | 0 | 0 | 60 | 25 |
| Trials | Total Soluble Phenolic Compounds (mg GAE·100 g−1 of Flour, d.b.) | γ-Aminobutyric Acid (mg·100 g−1 of Flour, d.b.) |
|---|---|---|
| Control | 348.05 ± 31.02 | 43.41 ± 0.30 |
| 1 | 385.29 ± 46.91 | 43.38 ± 0.19 |
| 2 | 407.13 ± 43.41 | 48.56 ± 1.27 |
| 3 | 386.60 ± 37.60 | 47.56 ± 0.06 |
| 4 | 436.48 ± 45.95 | 61.13 ± 2.69 |
| 5 | 342.36 ± 49.10 | 44.89 ± 1.50 |
| 6 | 424.58 ± 25.16 | 58.32 ± 2.40 |
| 7 | 404.15 ± 29.50 | 43.89 ± 0.39 |
| 8 | 445.62 ± 41.31 | 54.52 ± 3.50 |
| 9 | 476.49 ± 63.72 | 52.21 ± 2.12 |
| 10 | 479.34 ± 73.29 | 54.97 ± 4.71 |
| 11 | 497.12 ± 80.59 | 52.48 ± 3.87 |
| 12 | 464.40 ± 69.30 | 54.12 ± 0.41 |
| Trials | Soluble Protein (%) | Soluble Solids (°Brix) | Reducing Sugars (g of Glucose·100−1 g of Extract) |
|---|---|---|---|
| Control | 8.43 ± 0.11 | 0.8 ± < 0.01 | 3.28 ± 0.33 |
| 1 | 9.31 ± 0.13 | 1.0 ± < 0.01 | 3.74 ± 0.18 |
| 2 | 11.96 ± 0.34 | 1.2 ± < 0.01 | 6.37 ± 0.44 |
| 3 | 10.04 ± 0.08 | 1.0 ± < 0.01 | 4.01 ± 0.10 |
| 4 | 17.23 ± 0.48 | 3.0 ± < 0.01 | 10.46 ± 0.75 |
| 5 | 9.89 ± 0.10 | 0.8 ± < 0.01 | 4.54 ± 0.33 |
| 6 | 17.58 ± 1.11 | 2.6 ± < 0.01 | 10.77 ± 1.48 |
| 7 | 10.09 ± 0.21 | 1.0 ± < 0.01 | 4.87 ± 0.12 |
| 8 | 16.42 ± 0.16 | 1.8 ± < 0.01 | 10.09 ± 0.60 |
| 9 | 13.09 ± 0.04 | 1.2 ± < 0.01 | 7.98 ± 0.46 |
| 10 | 11.83 ± 0.28 | 1.4 ± < 0.01 | 7.29 ± 0.39 |
| 11 | 12.79 ± 0.22 | 1.2 ± < 0.01 | 8.88 ± 0.70 |
| 12 | 13.37 ± 0.01 | 1.4 ± < 0.01 | 8.37 ± 0.54 |
| Independent Variables | |||||
| Parameters | Goal | Importance | Solution | ||
| Coded Value | Real Value | ||||
| Time (h) | In range | 3 | 0.99 | 85.3 | |
| Temperature (°C) | In range | 3 | 1.10 | 30.46 | |
| Dependent Variables | |||||
| Parameter | Goal | Importance | Solution | Optimal Point Values | Relative Deviation (%) |
| Radicle Length (mm) | Maximize | 5 | 83.63 | 72.39 ± 3.94 | 15.54 |
| Seed Vigor (%) | Maximize | 5 | 75.87 | 71.33 ± 0.94 | 6.36 |
| TSPC (mg GAE·100 g−1 of flour, d.b.) | Maximize | 5 | 435.15 | 395.98 ± 31.02 | 9.89 |
| GABA (mg·100 g−1 of flour, d.b.) | Maximize | 5 | 61.15 | 54.83 ± 2.07 | 10.73 |
| Soluble Protein (%) | Maximize | 5 | 18.87 | 19.24 ± 0.68 | 1.88 |
| Soluble Solids (%) | Maximize | 5 | 2.99 | 2.93 ± 0.25 | 2.01 |
| Reducing Sugars (%, in glucose) | Maximize | 5 | 12.13 | 14.02 ± 0.33 | 13.46 |
| Desirability 90.42% | |||||
| Trials | L* | a* | b* | ΔE | WI |
|---|---|---|---|---|---|
| Standard | 79.23 ± < 0.01 | 0.08 ± < 0.01 | 21.16 ± 0.09 | - | 70.34 ± 0.06 |
| Control | 77.18 ± 0.33 | 0.59 ± < 0.01 | 20.33 ± 0.27 | 1.15 ± 0.40 | 69.43 ± 0.08 |
| 1 | 75.25 ± 0.20 | 0.52 ± 0.02 | 20.27 ± 0.08 | 3.00 ± 0.19 | 68.00 ± 0.19 |
| 2 | 75.78 ± 0.02 | 0.58 ± 0.01 | 20.40 ± 0.19 | 2.47 ± 0.05 | 68.33 ± 0.11 |
| 3 | 76.22 ± 0.21 | 0.44 ± 0.03 | 20.39 ± 0.04 | 2.03 ± 0.21 | 68.67 ± 0.67 |
| 4 | 77.71 ± 0.30 | 0.65 ± 0.02 | 19.71 ± 0.21 | 1.24 ± 0.15 | 70.24 ± 0.35 |
| 5 | 76.89 ± 0.98 | 0.42 ± 0.03 | 20.01 ± 0.12 | 1.65 ± 0.74 | 69.42 ± 0.67 |
| 6 | 75.92 ± 0.31 | 0.48 ± 0.02 | 19.61 ± 0.25 | 2.58 ± 0.26 | 68.94 ± 0.34 |
| 7 | 76.38 ± 0.16 | 0.45 ± < 0.01 | 20.27 ± 0.31 | 1.92 ± 0.21 | 68.87 ± 0.21 |
| 8 | 77.28 ± 0.72 | 0.37 ± 0.02 | 19.57 ± 0.10 | 1.62 ± 0.39 | 70.01 ± 0.59 |
| 9 | 75.99 ± 0.23 | 0.54 ± < 0.01 | 20.53 ± 0.05 | 2.25 ± 0.23 | 68.40 ± 0.18 |
| 10 | 75.86 ± 0.15 | 0.34 ± 0.01 | 20.02 ± 0.23 | 2.48 ± 0.09 | 68.64 ± 0.26 |
| 11 | 76.26 ± 0.28 | 0.31 ± 0.02 | 19.67 ± 0.18 | 2.24 ± 0.24 | 69.17 ± 0.26 |
| 12 | 76.15 ± 0.18 | 0.37 ± 0.03 | 20.02 ± 0.29 | 2.21 ± 0.06 | 68.86 ± 0.32 |
| Trials | Firmness (N) | Hardness (N) | Springiness (%) | Cohesiveness (%) | Gumminess (N) | Chewiness (N) | Resilience (%) |
|---|---|---|---|---|---|---|---|
| Standard | 7.18 ± 0.39 | 9.29 ± 0.57 | 89.83 ± 0.44 | 56.20 ± 0.84 | 5.22 ± 0,34 | 4.69 ± 0.31 | 24.46 ± 0.77 |
| Control | 9.73 ± 0.88 | 11.04 ± 1.03 | 87.62 ± 1.38 | 53.54 ± 1.31 | 5.90 ± 0.45 | 5.17 ± 0.41 | 22.18 ± 0.99 |
| 1 | 8.63 ± 0.50 | 9.73 ± 0.58 | 88.29 ± 0.78 | 56.66 ± 0.70 | 5.51 ± 0.31 | 4.87 ± 0.29 | 24.64 ± 0.57 |
| 2 | 5.45 ± 0.55 | 7.47 ± 0.58 | 86.97 ± 0.82 | 55.45 ± 1.96 | 4.14 ± 0.32 | 3.60 ± 0.29 | 22.55 ± 1.20 |
| 3 | 5.55 ± 0.51 | 7.45 ± 0.68 | 90.45 ± 0.78 | 58.51 ± 0.71 | 4.36 ± 0.37 | 3.94 ± 0.32 | 25.54 ± 0.61 |
| 4 | 7.05 ± 0.52 | 9.04 ± 0.65 | 78.50 ± 3.58 | 46.15 ± 0.72 | 4.17 ± 0.31 | 3.27 ± 0.28 | 13.35 ± 0.31 |
| 5 | 9.13 ± 0.40 | 10.39 ± 0.45 | 87.66 ± 0.95 | 53.67 ± 1.03 | 5.58 ± 0.30 | 4.89 ± 0.24 | 22.46 ± 0.90 |
| 6 | 4.60 ± 0.27 | 6.19 ± 0.40 | 78.35 ± 0.83 | 51.31 ± 1.12 | 3.18 ± 0.25 | 2.49 ± 0.20 | 16.81 ± 0.70 |
| 7 | 7.49 ± 0.52 | 8.37 ± 0.54 | 89.54 ± 1.01 | 59.33 ± 1.12 | 4.97 ± 0.39 | 4.45 ± 0.35 | 26.61 ± 0.90 |
| 8 | 4.90 ± 0.27 | 6.66 ± 0.38 | 80.50 ± 0.88 | 53.91 ± 1.05 | 3.59 ± 0.20 | 2.89 ± 0.18 | 19.15 ± 0.31 |
| 9 | 5.31 ± 0.36 | 7.19 ± 0.45 | 86.92 ± 0.84 | 56.90 ± 1.30 | 4.09 ± 0.22 | 3.55 ± 0.21 | 23.26 ± 0.77 |
| 10 | 6.75 ± 0.38 | 7.53 ± 0.46 | 85.67 ± 0.81 | 56.37 ± 1.66 | 4.24 ± 0.25 | 3.63 ± 0.21 | 22.62 ± 0.66 |
| 11 | 8.17 ± 0.49 | 9.15 ± 0.59 | 84.94 ± 0.63 | 55.32 ± 1.32 | 5.05 ± 0.26 | 4.29 ± 0.22 | 22.39 ± 0.95 |
| 12 | 6.01 ± 0.48 | 8.01 ± 0.60 | 86.42 ± 0.70 | 55.05 ± 0.93 | 4.41 ± 0.33 | 3.81 ± 0.28 | 22.17 ± 0.81 |
| Dependent Variable | Predictive Mathematical Model | R2 (%) | p-Value | Fcalc/Ftab |
|---|---|---|---|---|
| Springiness (%) | 86.01 − 3.31x1 − 2.39x2 − 0.99x12 − 2.66x1x2 | 91.33 | 0.001 | 6.22 |
| Cohesiveness (%) | 56.12 − 2.12x1 − 1.89x2 − 1.86x12 − 2.79x1x2 | 88.08 | 0.002 | 4.37 |
| Gumminess (N) | 4.37 − 0.32x1 − 0.38x2 + 0.30x1x2 | 85.78 | 0.001 | 5.51 |
| Chewiness (N) | 3.75 − 0.67x1 − 0.43x2 + 0.15x1x2 | 88.84 | <0.001 | 8.24 |
| Resilience (%) | 22.77 − 2.79x1 − 2.36x2 − 1.47x12 − 2.53x1x2 | 95.67 | <0.001 | 13.06 |
| Independent Variables | ||||
| Parameters | Goal | Importance | Coded Value | Real Value |
| Time (h) | In range | 3 | 1.41 | 96.0 |
| Temperature (°C) | In range | 3 | 0.44 | 29.34 |
| Dependent Variables | ||||
| Parameters | Goal | Importance | Solution | |
| Springiness (%) | Minimize | 5 | 76.70 | |
| Cohesiveness (%) | Minimize | 5 | 46.91 | |
| Gumminess (N) | Minimize | 5 | 3.52 | |
| Chewiness (N) | Minimize | 5 | 2.72 | |
| Resilience (%) | Is in range | 3 | 13.35 | |
| Desirability | 92.60% | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nascimento, G.K.S.d.; Silva, M.S.; Andressa, I.; Fagundes, M.B.; Vendruscolo, R.G.; Oliveira, J.R.; Barcia, M.T.; Benassi, V.M.; Neves, N.d.A.; Lima, C.T.; et al. A New Advancement in Germination Biotechnology of Purple Creole Corn: Bioactive Compounds and In Situ Enzyme Activity for Water-Soluble Extract and Pan Bread. Metabolites 2024, 14, 35. https://doi.org/10.3390/metabo14010035
Nascimento GKSd, Silva MS, Andressa I, Fagundes MB, Vendruscolo RG, Oliveira JR, Barcia MT, Benassi VM, Neves NdA, Lima CT, et al. A New Advancement in Germination Biotechnology of Purple Creole Corn: Bioactive Compounds and In Situ Enzyme Activity for Water-Soluble Extract and Pan Bread. Metabolites. 2024; 14(1):35. https://doi.org/10.3390/metabo14010035
Chicago/Turabian StyleNascimento, Glauce Kelly Silva do, Michelle Santos Silva, Irene Andressa, Mariane Bittencourt Fagundes, Raquel Guidetti Vendruscolo, Josimar Rodrigues Oliveira, Milene Teixeira Barcia, Vivian Machado Benassi, Nathália de Andrade Neves, Cristiane Teles Lima, and et al. 2024. "A New Advancement in Germination Biotechnology of Purple Creole Corn: Bioactive Compounds and In Situ Enzyme Activity for Water-Soluble Extract and Pan Bread" Metabolites 14, no. 1: 35. https://doi.org/10.3390/metabo14010035
APA StyleNascimento, G. K. S. d., Silva, M. S., Andressa, I., Fagundes, M. B., Vendruscolo, R. G., Oliveira, J. R., Barcia, M. T., Benassi, V. M., Neves, N. d. A., Lima, C. T., & Schmiele, M. (2024). A New Advancement in Germination Biotechnology of Purple Creole Corn: Bioactive Compounds and In Situ Enzyme Activity for Water-Soluble Extract and Pan Bread. Metabolites, 14(1), 35. https://doi.org/10.3390/metabo14010035