



The Detection of Primary Sclerosing Cholangitis Using Volatile Metabolites in Fecal Headspace and Exhaled Breath

 , , and
, , and 
Abstract
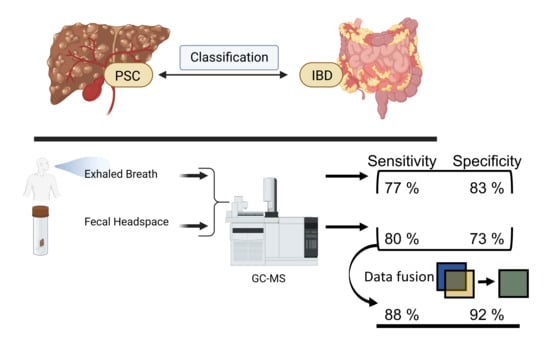
:
1. Introduction
2. Materials and Methods
2.1. Study Design and Population
2.2. Breath and Blood Collection
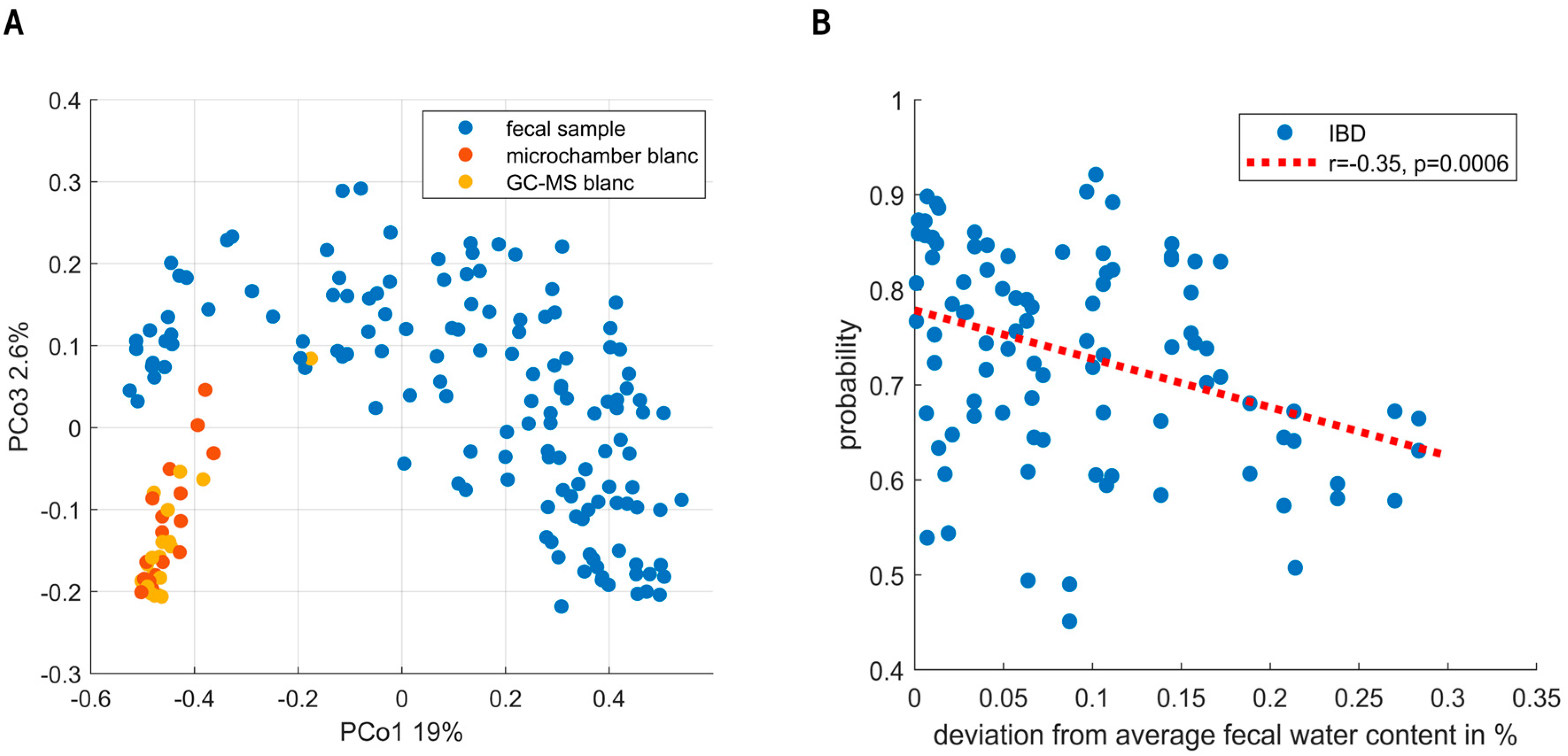
2.3. Fecal Sampling and Analysis
2.4. Statistical Analysis
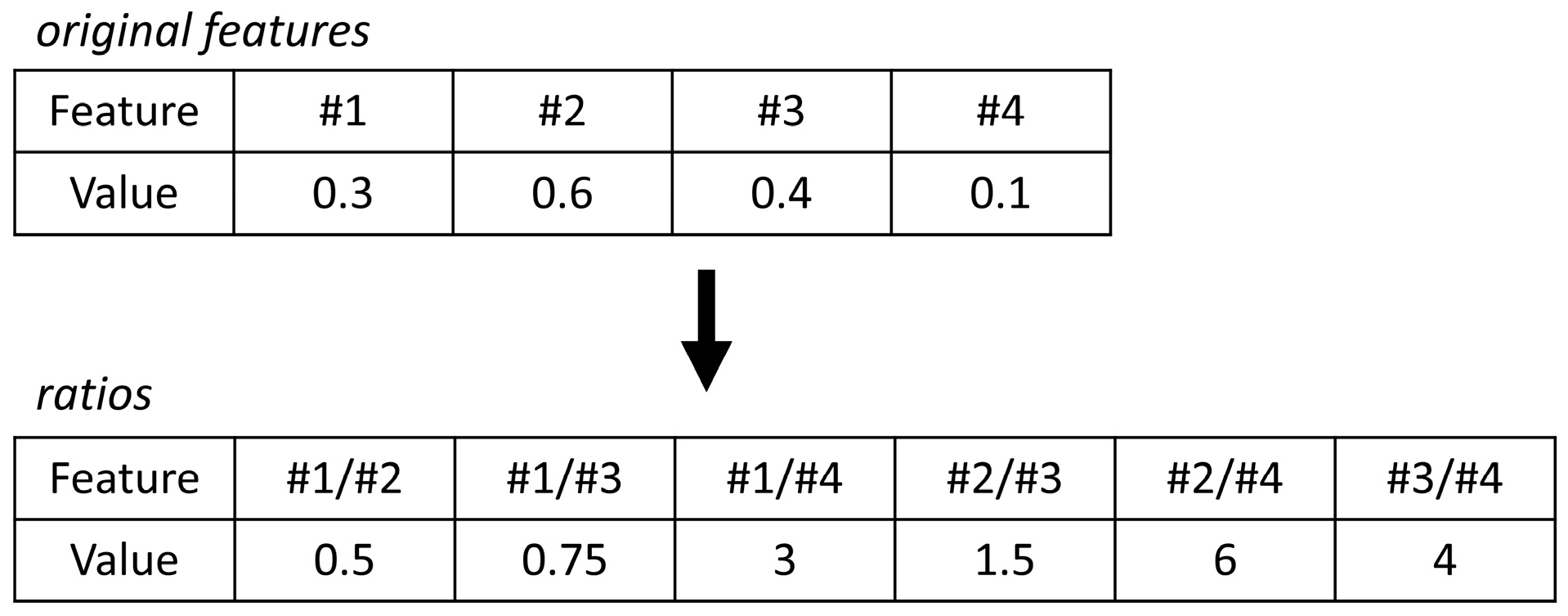
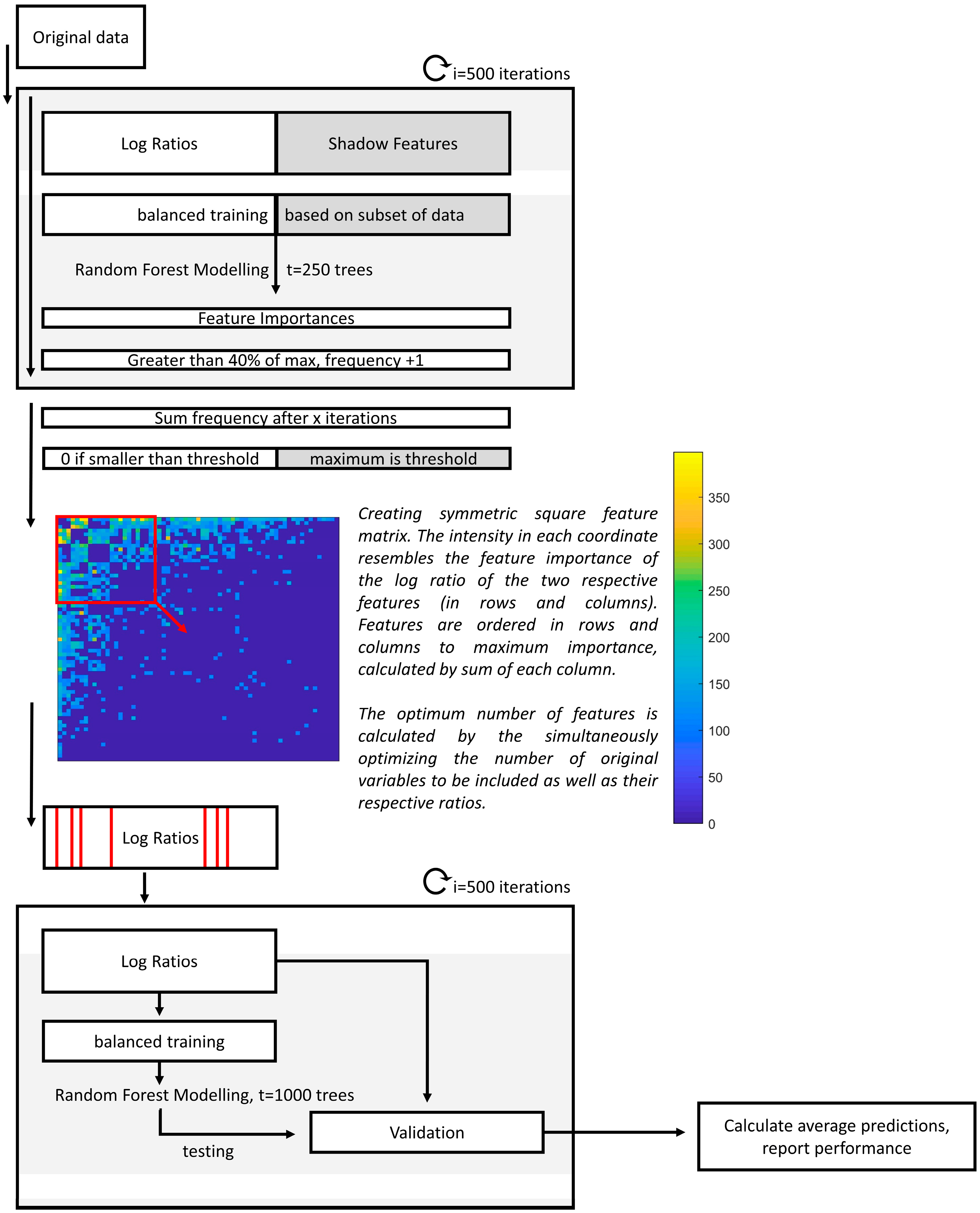
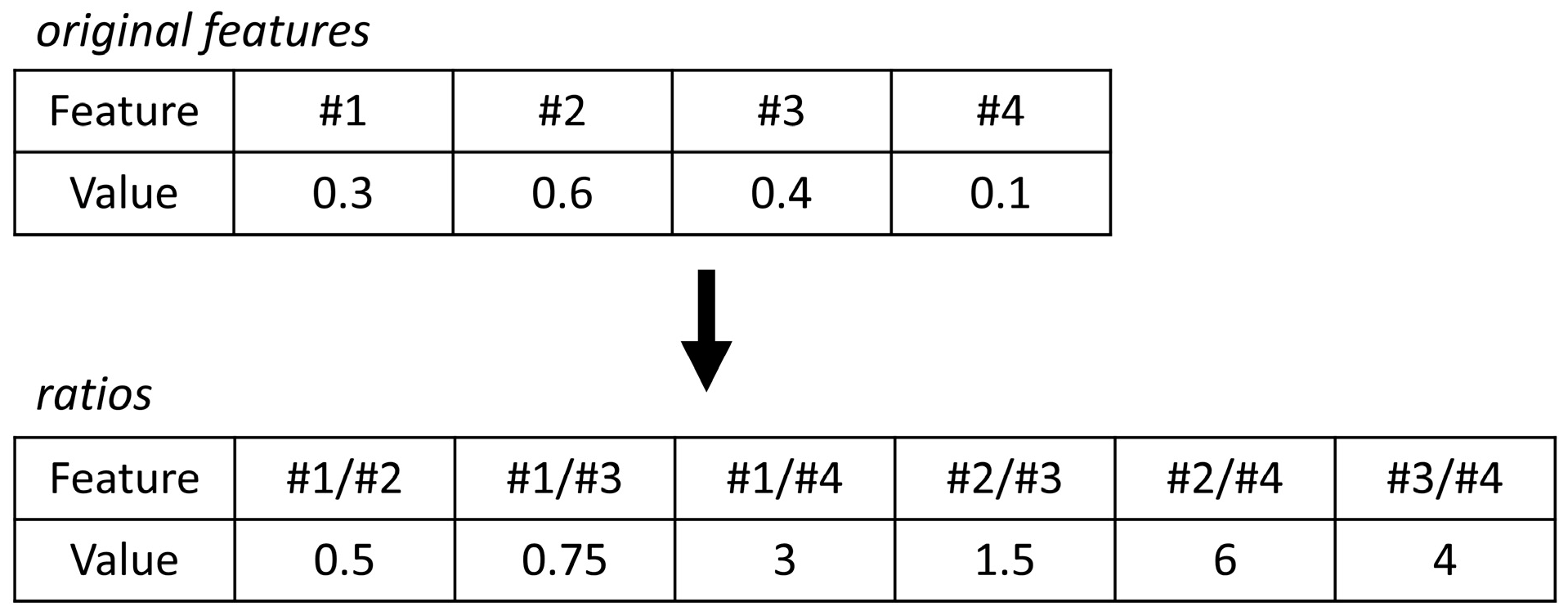
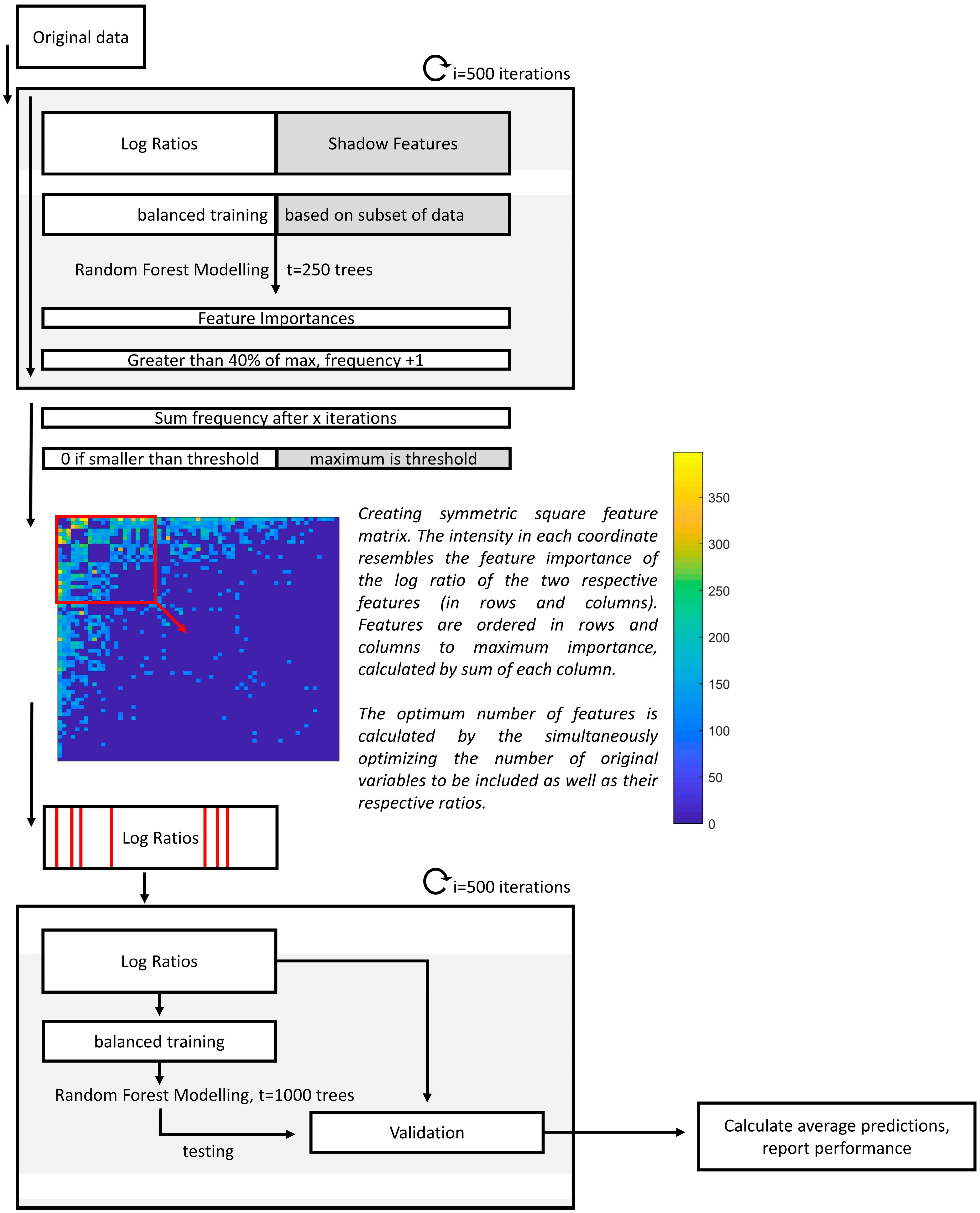
2.4.1. Interpreting Log Ratios with Random Forest
2.4.2. Data Fusion with Exhaled Breath
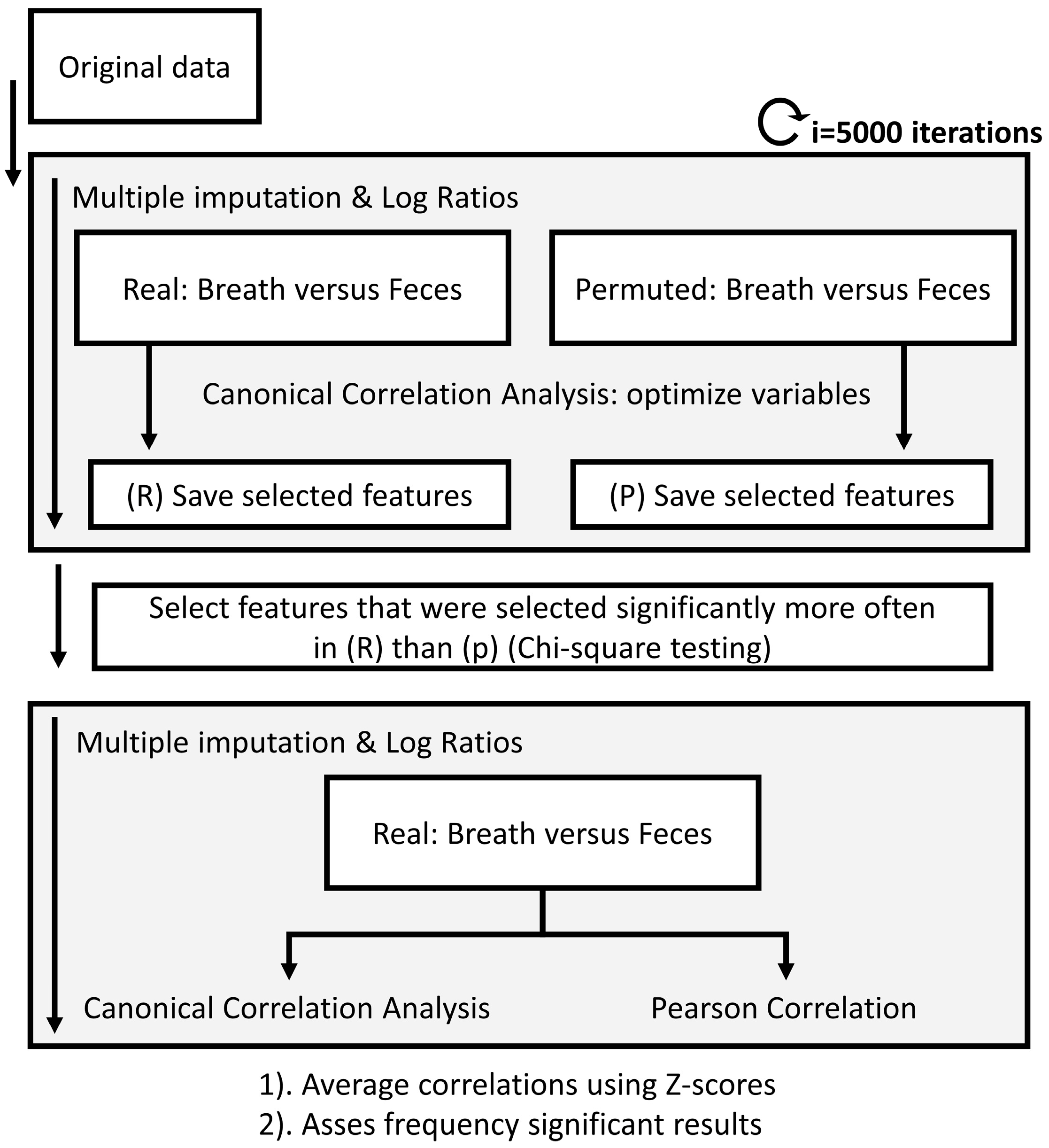
2.4.3. Correlating Feces, Exhaled Breath and Blood
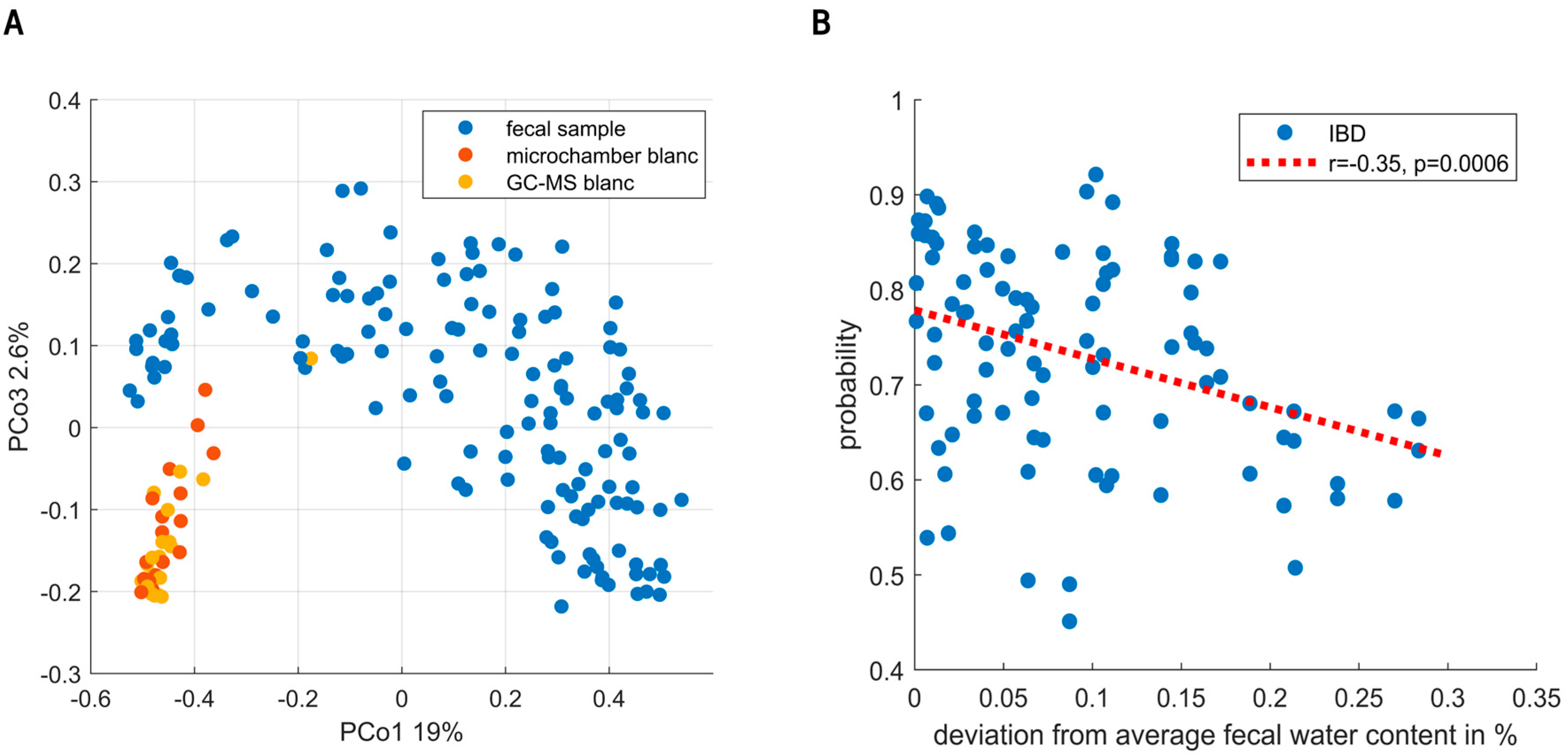
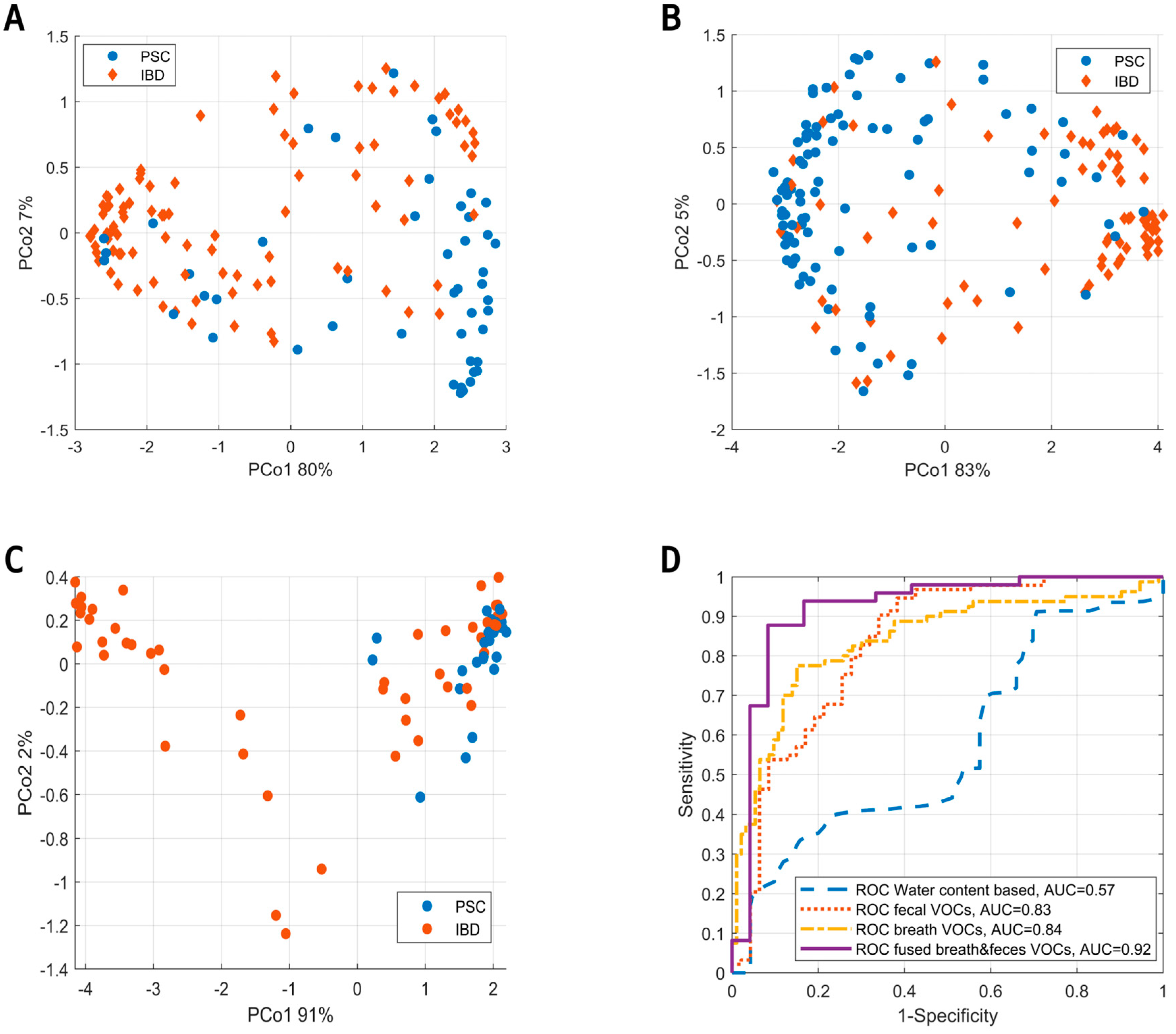
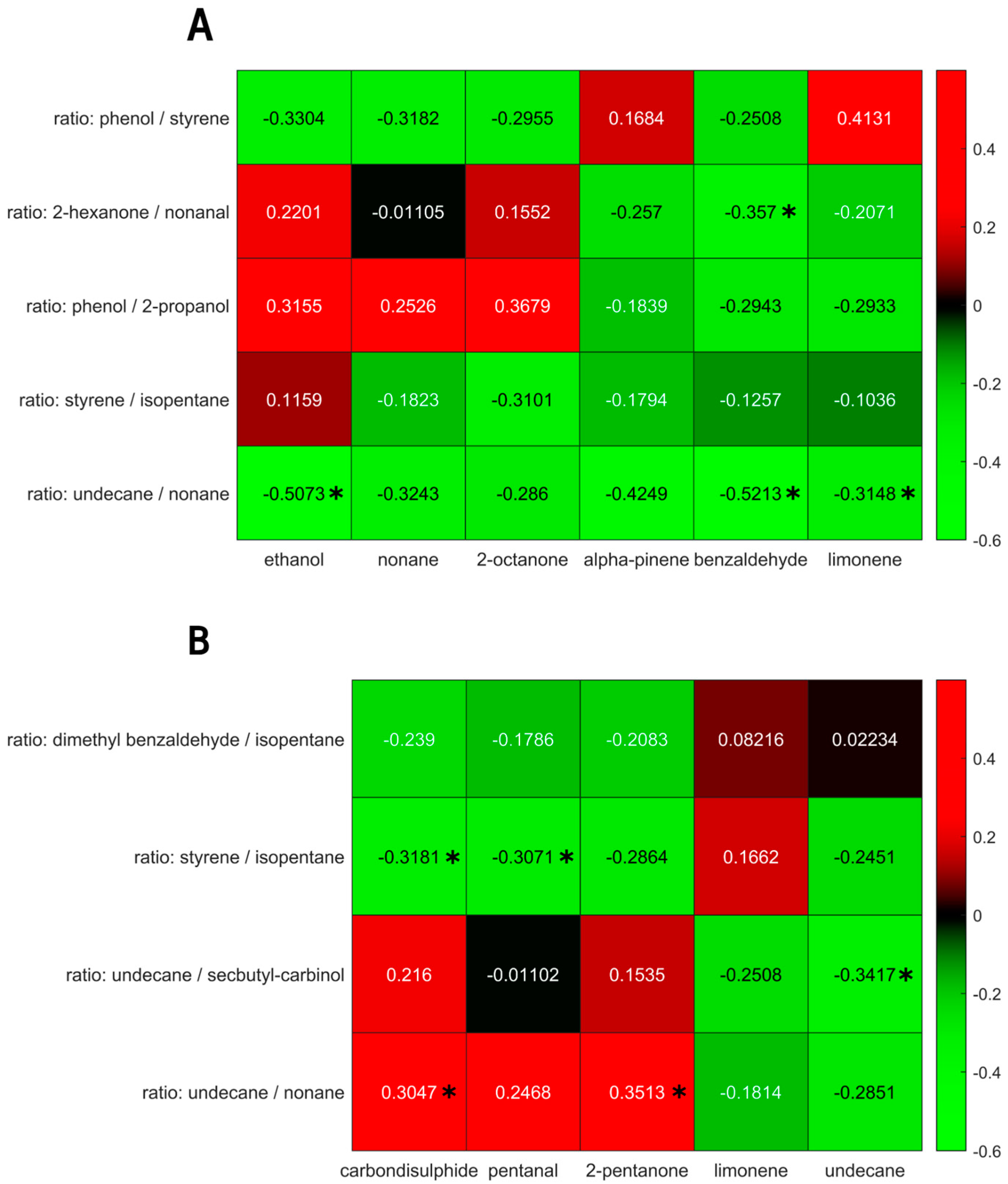
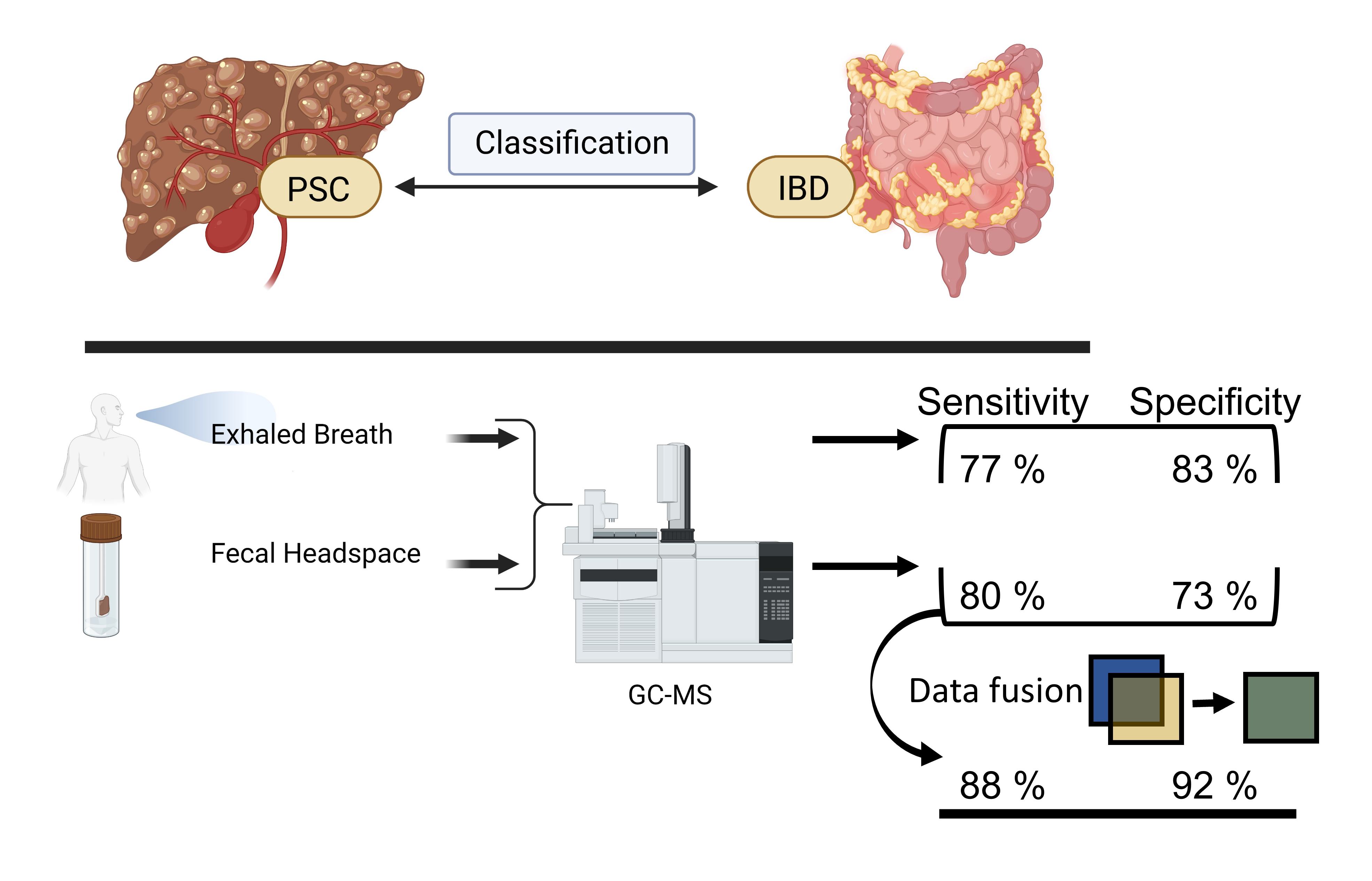
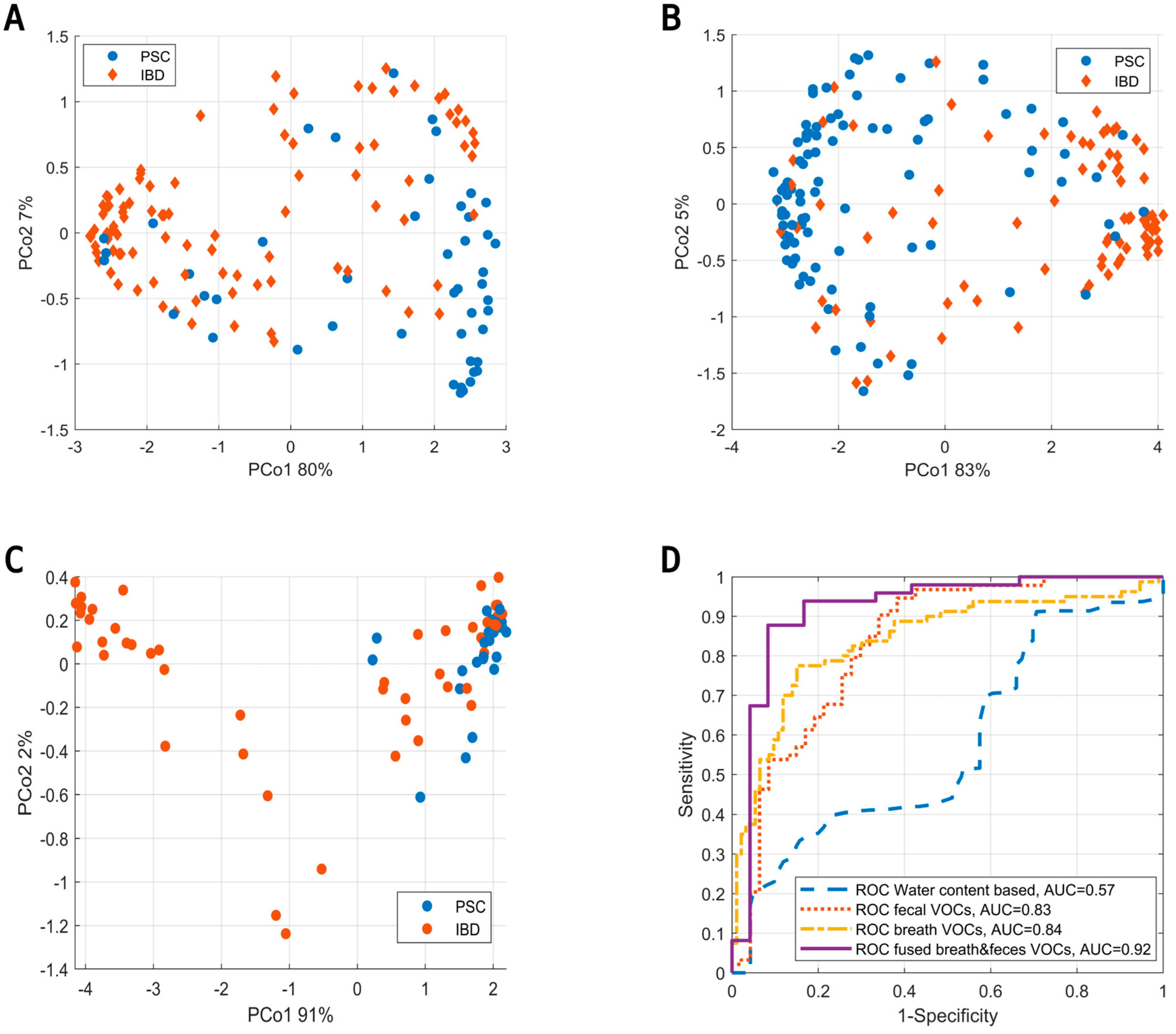
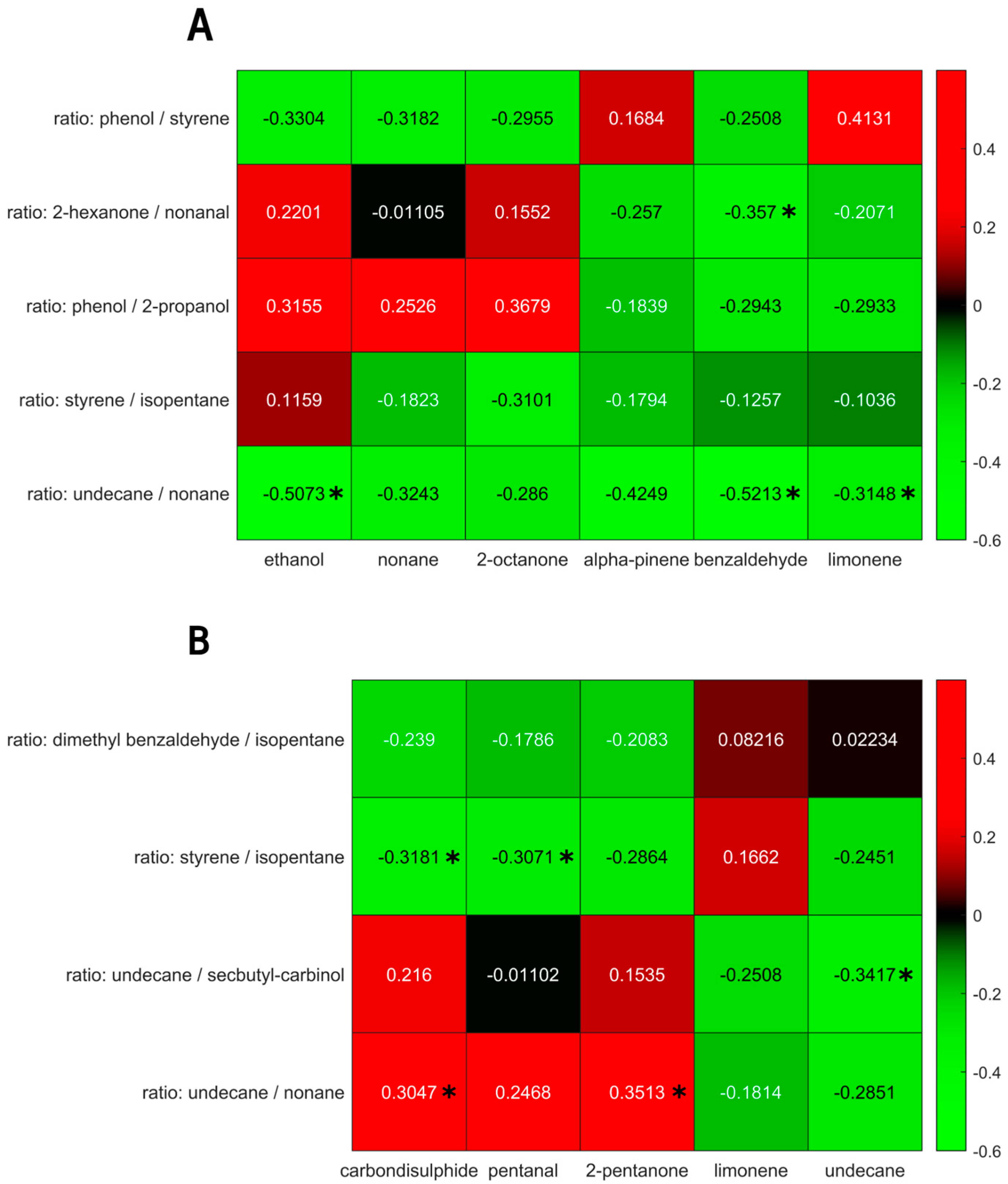
3. Results
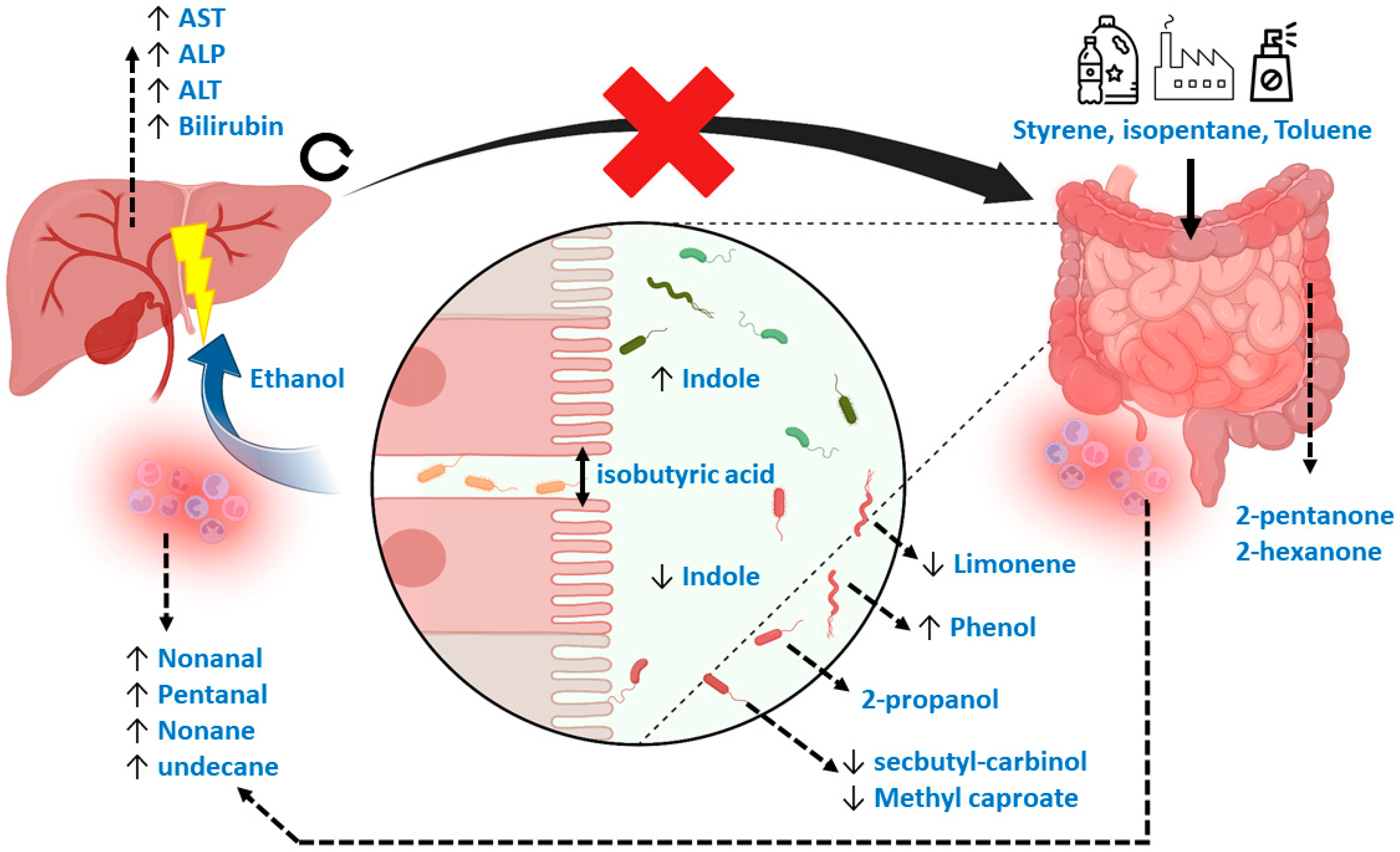
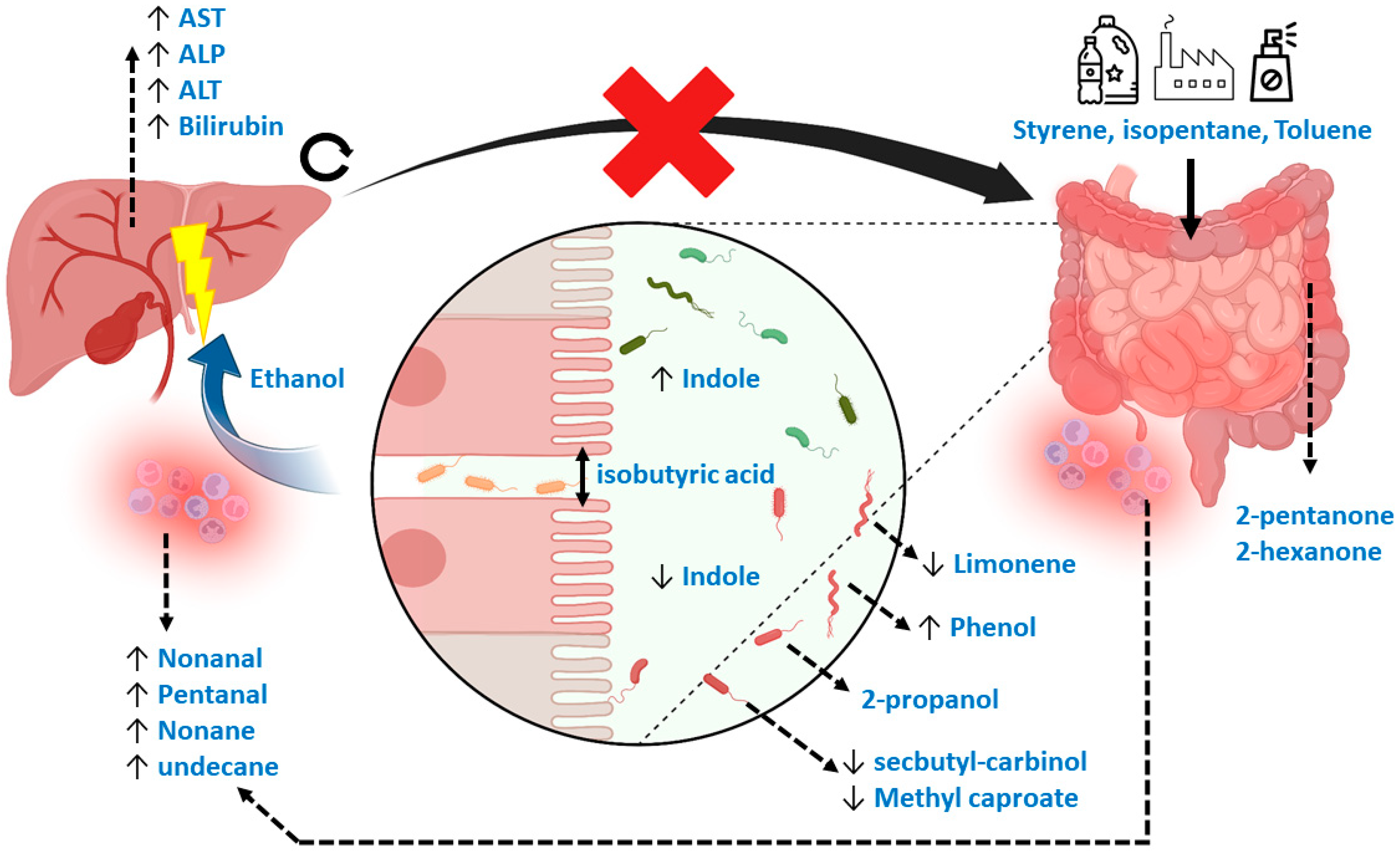
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karlsen, T.H.; Folseraas, T.; Thorburn, D.; Vesterhus, M. Primary sclerosing cholangitis—A comprehensive review. J. Hepatol. 2017, 67, 1298–1323. [Google Scholar] [CrossRef] [PubMed]
- Razumilava, N.; Gores, G.J. Surveillance for cholangiocarcinoma in patients with primary sclerosing cholangitis: Effective and justified? Clin. Liver Dis. 2016, 8, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhang, S.; Weng, J.-F.; Huang, D.; Gu, W.-L. Recent discoveries in microbiota dysbiosis, cholangiocytic factors, and models for studying the pathogenesis of primary sclerosing cholangitis. Open Med. 2022, 17, 915–929. [Google Scholar] [CrossRef] [PubMed]
- Dyson, J.K.; Beuers, U.; Jones, D.E.J.; Lohse, A.W.; Hudson, M. Primary sclerosing cholangitis. Lancet 2018, 391, 2547–2559. [Google Scholar] [CrossRef] [PubMed]
- Mjelle, A.B.; Fossdal, G.; Gilja, O.H.; Vesterhus, M. Liver Elastography in Primary Sclerosing Cholangitis Patients Using Three Different Scanner Systems. Ultrasound Med. Biol. 2020, 46, 1854–1864. [Google Scholar] [CrossRef] [PubMed]
- European Association for the Study of the Liver. EASL Clinical Practice Guidelines: Management of cholestatic liver diseases. J. Hepatol. 2009, 51, 237–267. [Google Scholar] [CrossRef] [PubMed]
- Burak, K.W.; Angulo, P.; Lindor, K.D. Is there a role for liver biopsy in primary sclerosing cholangitis? Am. J. Gastroenterol. 2003, 98, 1155–1158. [Google Scholar] [CrossRef]
- Sharma, P. Value of Liver Function Tests in Cirrhosis. J. Clin. Exp. Hepatol. 2022, 12, 948–964. [Google Scholar] [CrossRef]
- W Walker, D.I.; Juran, B.D.; Cheung, A.C.; Schlicht, E.M.; Liang, Y.; Niedzwiecki, M.; LaRusso, N.F.; Gores, G.J.; Jones, D.P.; Miller, G.W.; et al. High-Resolution Exposomics and Metabolomics Reveals Specific Associations in Cholestatic Liver Diseases. Hepatol. Commun. 2022, 6, 965–979. [Google Scholar] [CrossRef]
- Tietz-Bogert, P.S.; Kim, M.; Cheung, A.; Tabibian, J.H.; Heimbach, J.K.; Rosen, C.B.; Nandakumar, M.; Lazaridis, K.N.; LaRusso, N.F.; Sung, J.; et al. Metabolomic Profiling of Portal Blood and Bile Reveals Metabolic Signatures of Primary Sclerosing Cholangitis. Int. J. Mol. Sci. 2018, 19, 3188. [Google Scholar] [CrossRef]
- Mindikoglu, A.L.; Coarfa, C.; Opekun, A.R.; Shah, V.H.; Arab, J.P.; Lazaridis, K.N.; Putluri, N.; Ambati, C.R.; Robertson, M.J.; Devaraj, S.; et al. Metabolomic biomarkers are associated with mortality in patients with cirrhosis caused by primary biliary cholangitis or primary sclerosing cholangitis. Futur. Sci. OA 2020, 6, FSO441. [Google Scholar] [CrossRef] [PubMed]
- Trottier, J.; Białek, A.; Caron, P.; Straka, R.J.; Heathcote, J.; Milkiewicz, P.; Barbier, O. Metabolomic profiling of 17 bile acids in serum from patients with primary biliary cirrhosis and primary sclerosing cholangitis: A pilot study. Dig. Liver Dis. 2012, 44, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Lammert, C.; Shin, A.S.; Xu, H.; Hemmerich, C.; O’Connell, T.M.; Chalasani, N. Short-chain fatty acid and fecal microbiota profiles are linked to fibrosis in primary biliary cholangitis. FEMS Microbiol. Lett. 2021, 368, fnab038. [Google Scholar] [CrossRef] [PubMed]
- Awoniyi, M.; Wang, J.; Ngo, B.; Meadows, V.; Tam, J.; Viswanathan, A.; Lai, Y.; Montgomery, S.; Farmer, M.; Kummen, M.; et al. Protective and aggressive bacterial subsets and metabolites modify hepatobiliary inflammation and fibrosis in a murine model of PSC. Gut 2022, 72, 671–685. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, B.; Li, Y.; Wei, Y.; Huang, B.; Liang, J.; You, Z.; Li, Y.; Qian, Q.; Wang, R.; et al. Altered faecal microbiome and metabolome in IgG4-related sclerosing cholangitis and primary sclerosing cholangitis. Gut 2022, 71, 899–909. [Google Scholar] [CrossRef]
- Kummen, M.; Thingholm, L.B.; Rühlemann, M.C.; Holm, K.; Hansen, S.H.; Moitinho-Silva, L.; Liwinski, T.; Zenouzi, R.; Storm-Larsen, C.; Midttun, Ø.; et al. Altered gut microbial metabolism of essential nutrients in primary sclerosing cholangitis Europe PMC Funders Group. Gastroenterology 2021, 160, 1784–1798. [Google Scholar] [CrossRef]
- Little, R.; Wine, E.; Kamath, B.M.; Griffiths, A.M.; Ricciuto, A. Gut microbiome in primary sclerosing cholangitis: A review. World J. Gastroenterol. 2020, 26, 2768–2780. [Google Scholar] [CrossRef]
- Özdirik, B.; Müller, T.; Wree, A.; Tacke, F.; Sigal, M.; Turroni, S. Molecular Sciences The Role of Microbiota in Primary Sclerosing Cholangitis and Related Biliary Malignancies. J. Mol. Sci. 2021, 22, 6975. [Google Scholar] [CrossRef]
- Jardon, K.M.; Canfora, E.E.; Goossens, G.H.; Blaak, E.E. Dietary macronutrients and the gut microbiome: A precision nutrition approach to improve cardiometabolic health. Gut 2022, 71, 1214–1226. [Google Scholar] [CrossRef]
- Kulaksiz, H.; Rudolph, G.; Kloeters-Plachky, P.; Sauer, P.; Geiss, H.; Stiehl, A. Biliary candida infections in primary sclerosing cholangitis. J. Hepatol. 2006, 45, 711–716. [Google Scholar] [CrossRef]
- Lemoinne, S.; Kemgang, A.; Ben Belkacem, K.; Straube, M.; Jegou, S.; Corpechot, C.; Chazouillères, O.; Housset, C.; Sokol, H.; Network, S.-A.I. Fungi participate in the dysbiosis of gut microbiota in patients with primary sclerosing cholangitis. Gut 2020, 69, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shen, J.; Ran, Z. Crosstalk between the gut and the liver via susceptibility loci: Novel advances in inflammatory bowel disease and autoimmune liver disease. Clin. Immunol. 2017, 175, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Van Vorstenbosch, R.; Cheng, H.R.; Jonkers, D.; Penders, J.; Schoon, E.; Masclee, A.; van Schooten, F.-J.; Smolinska, A.; Mujagic, Z. Systematic Review: Contribution of the Gut Microbiome to the Volatile Metabolic Fingerprint of Colorectal Neoplasia. Metabolites 2023, 13, 55. [Google Scholar] [CrossRef] [PubMed]
- Stavropoulos, G.; van Munster, K.; Ferrandino, G.; Sauca, M.; Ponsioen, C.; van Schooten, F.-J.; Smolinska, A. Liver impairment—The potential application of volatile organic compounds in hepatology. Metabolites 2021, 11, 618. [Google Scholar] [CrossRef]
- Stavropoulos, G.; van Vorstenbosch, R.; Jonkers, D.M.A.E.; Penders, J.; Hill, J.E.; van Schooten, F.-J.; Smolinska, A. Advanced data fusion: Random forest proximities and pseudo-sample principle towards increased prediction accuracy and variable interpretation. Anal. Chim. Acta 2021, 1183, 339001. [Google Scholar] [CrossRef]
- Stavropoulos, G.; van Munster, K.; van Vorstenbosch, R.; Pachen, D.; van Schooten, F.-J.; Ponsioen, C.; Smolinska, A. The Potential of Volatile Organic Compounds to Diagnose Primary Sclerosing Cholagitis; Maastricht University: Maastricht, The Netherlands, 2023; expected: Soon in press (currently Under Review); Available online: https://drive.google.com/file/d/1Xxe3oyixqIEFutUfZq6o8SMhYZFkn0rk/view?usp=sharing (accessed on 19 December 2023).
- Magro, F.; Gionchetti, P.; Eliakim, R.; Ardizzone, S.; Armuzzi, A.; Barreiro-de Acosta, M.; Burisch, J.; Gecse, K.B.; Hart, A.L.; Hindryckx, P.; et al. Third European evidence-based consensus on diagnosis and management of ulcerative colitis. Part 1: Definitions, diagnosis, extra-intestinal manifestations, pregnancy, cancer surveillance, surgery, and ileo-anal pouch disorders. J. Crohn’s Colitis 2017, 11, 649–670. [Google Scholar] [CrossRef] [PubMed]
- Ratiu, I.A.; Mametov, R.; Ligor, T.; Buszewski, B. Micro-Chamber/Thermal Extractor (µ-CTE) as a new sampling system for VOCs emitted by feces. Sci. Rep. 2021, 11, 18780. [Google Scholar] [CrossRef]
- Vork, L.; Wilms, E.; Penders, J.; Jonkers, D.M.A.E. Stool Consistency: Looking Beyond the Bristol Stool Form Scale. J. Neurogastroenterol. Motil. 2019, 25, 625. [Google Scholar] [CrossRef]
- Fijten, R.R.R.; Smolinska, A.; Drent, M.; Dallinga, J.W.; Mostard, R.; Pachen, D.M.; van Schooten, F.J.; Boots, A.W. The necessity of external validation in exhaled breath research: A case study of sarcoidosis. J. Breath Res. 2017, 12, 016004. [Google Scholar] [CrossRef]
- Smolinska, A.; Hauschild, A.C.; Fijten, R.R.R.; Dallinga, J.W.; Baumbach, J.; Van Schooten, F.J. Current breathomics—A review on data pre-processing techniques and machine learning in metabolomics breath analysis. J. Breath Res. 2014, 8, 027105. [Google Scholar] [CrossRef]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic quotient normalization as robust method to account for dilution of complex biological mixtures. Application in1H NMR metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef] [PubMed]
- Malyjurek, Z.; de Beer, D.; Joubert, E.; Walczak, B. Working with log-ratios. Anal. Chim. Acta 2019, 1059, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Stavropoulos, G.; van Voorstenbosch, R.; van Schooten, F.-J.; Smolinska, A. Random Forest and Ensemble Methods. Compr. Chemom. 2020, 661–672. [Google Scholar] [CrossRef]
- Engel, J.; Blanchet, L.; Bloemen, B.; Heuvel, L.v.D.; Engelke, U.; Wevers, R.; Buydens, L. Regularized MANOVA (rMANOVA) in untargeted metabolomics. Anal. Chim. Acta 2015, 899, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kurada, S.; Alkhouri, N.; Fiocchi, C.; Dweik, R.; Rieder, F. Alimentary Pharmacology and Therapeutics Review article: Breath analysis in in flammatory bowel diseases. Aliment. Pharmacol. Ther. 2014, 41, 329–341. [Google Scholar] [CrossRef]
- Sadighara, P.; Akbari, N.; Mostashari, P.; Yazdanfar, N.; Shokri, S. The amount and detection method of styrene in foods: A systematic review and meta-analysis. Food Chem. X 2022, 13, 100238. [Google Scholar] [CrossRef]
- Guazzotti, V.; Hendrich, V.; Gruner, A.; Fiedler, D.; Störmer, A.; Welle, F. Migration of Styrene in Yogurt and Dairy Products Packaged in Polystyrene: Results from Market Samples. Foods 2022, 11, 2120. [Google Scholar] [CrossRef]
- Longo, V.; Forleo, A.; Ferramosca, A.; Notari, T.; Pappalardo, S.; Siciliano, P.; Capone, S.; Montano, L. Blood, urine and semen Volatile Organic Compound (VOC) pattern analysis for assessing health environmental impact in highly polluted areas in Italy. Environ. Pollut. 2021, 286, 117410. [Google Scholar] [CrossRef]
- Garner, C.E.; Smith, S.; Lacy Costello, B.; White, P.; Spencer, R.; Probert, C.S.J.; Ratcliffem, N.M. Volatile organic compounds from feces and their potential for diagnosis of gastrointestinal disease. FASEB J. 2007, 21, 1675–1688. [Google Scholar] [CrossRef]
- De Preter, V.; Machiels, K.; Joossens, M.; Arijs, I.; Matthys, C.; Vermeire, S.; Rutgeerts, P.; Verbeke, K. Faecal metabolite profiling identifies medium-chain fatty acids as discriminating compounds in IBD. Gut 2015, 64, 447–458. [Google Scholar] [CrossRef]
- Ahmed, I.; Greenwood, R.; Costello, B.; Ratcliffe, N.; Probert, C.S. Investigation of faecal volatile organic metabolites as novel diagnostic biomarkers in inflammatory bowel disease. Aliment. Pharmacol. Ther. 2016, 43, 596–611. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; MacDonald, G.A.; Morrison, M.; Holtmann, G. Targeting the Gut Microbiome as a Treatment for Primary Sclerosing Cholangitis: A Conceptional Framework. Am. J. Gastroenterol. 2020, 115, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Couch, R.D.; Dailey, A.; Zaidi, F.; Navarro, K.; Forsyth, C.B.; Mutlu, E.; Engen, P.A.; Keshavarzian, A. Alcohol induced alterations to the human fecal VOC metabolome. PLoS ONE 2015, 10, e0119362. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sperandino, V. Indole Signaling at the host-microbiota-pathogen interface. MBio 2019, 10, 01031-19. [Google Scholar] [CrossRef] [PubMed]
- Smolinska, A.; Tedjo, D.I.; Blanchet, L.; Bodelier, A.; Pierik, M.J.; Masclee, A.A.; Dallinga, J.; Savelkoul, P.H.; Jonkers, D.M.; Penders, J.; et al. Volatile metabolites in breath strongly correlate with gut microbiome in CD patients. Anal. Chim. Acta 2018, 1025, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fabietti, F.; Delise, M.; Piccioli Bocca, A. Investigation into the benzene and toluene content of soft drinks. Food Control 2001, 12, 505–509. [Google Scholar] [CrossRef]
- Madhogaria, B.; Bhowmik, P.; Kundu, A. Correlation between human gut microbiome and diseases. Infect. Med. 2022, 1, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, Z.; Li, H.; Zhao, J.; Wei, X.; Lin, W.; Zhao, X.; Jiang, A.; Yuan, J. Endogenous ethanol produced by intestinal bacteria induces mitochondrial dysfunction in non-alcoholic fatty liver disease. J. Gastroenterol. Hepatol. 2020, 35, 2009–2019. [Google Scholar] [CrossRef]
- Ratcliffe, N.; Wieczorek, T.; Drabińska, N.; Gould, O.; Osborne, A.; Costello, B.P.J.d.L. A mechanistic study and review of volatile products from peroxidation of unsaturated fatty acids: An aid to understanding the origins of volatile organic compounds from the human body. J. Breath Res. 2020, 14, 034001. [Google Scholar] [CrossRef]
- Holz, O.; van Vorstenbosch, R.; Guenther, F.; Schuchardt, S.; Trinkmann, F.; van Schooten, F.-J.; Smolinska, A.; Hohlfeld, J.M. Changes of breath volatile organic compounds in healthy volunteers following segmental and inhalation endotoxin challenge. J. Breath Res. 2022, 16, 037102. [Google Scholar] [CrossRef]
- Visconti, A.; Le Roy, C.I.; Rosa, F.; Rossi, N.; Martin, T.C.; Mohney, R.P.; Li, W.; de Rinaldis, E.; Bell, J.T.; Venter, J.C.; et al. Interplay between the human gut microbiome and host metabolism. Nat. Commun. 2019, 10, 4505. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| sulfur dioxide | methyl formate | formic acid methyl ester | ethanol | trimethylamine |
| acrolein | acetone | 2-propanol | isoprene | pentane |
| dimethyl sulfide | metylacetate | carbon disulfide | 1-propanol | acetic acid |
| butanal | 2-butanone | hexane | 2-butanol | Dimethyl carbonate |
| isobutyl alcohol | methyl propionate | 1-butanol | benzene | 2-pentanone |
| propanoic acid | cyclopentanol | pentanal | acetic acid propyl ester | butanoic acid methyl ester |
| isobutyric acid | dimethyl disulfide | pyrrole | toluene | butanoic acid |
| 2-hexanone | butyric acid ethyl ester | hexanal | octane | propionic acid propyl ester |
| acetic acid butyl ester | isovaleric acid | 2-methyl butyric acid | N,N-dimethyl acetamide | pentanoic acid |
| nonane | styrene | dimethylsulfone | hexanoic acid methyl ester | hexanoic acid |
| benzaldehyde | phenol | dimethyltrisulfide | octanal | 3-carene |
| limonene | p-cresol | nonanal | decanal | methane amine |
| indole | 6-methyl indole |
| PSC (n = 8, d = 16) | PSC/IBD (n = 16, d = 31) | IBD (n = 49, d = 93) | p-Value | |
|---|---|---|---|---|
| Age (years, mean) | 56.4 | 47.6 | 47.5 | 0.20 |
| Sex | 2/6 | 10/6 | 26/23 | 0.25 |
| BMI | 23.5 | 24.6 | 25.7 | 0.27 |
| Smoking | 5/3/0 | 11/4/1 | 25/20/4 | 0.79 |
| Diet | 2/6 | 3/13 | 8/41 | 0.8 |
| Medication | 8/0 | 16/0 | 44/5 | 0.47 |
| Ursodeoxycholic acid | 7/1 | 13/3 | 0/43 | <0.001 |
| Corticosteroids | 0/8 | 0/16 | 3/41 | 0.7 |
| Thiopurines | 0/8 | 2/14 | 4/40 | 0.68 |
| Mesalazine | 0/8 | 8/8 | 25/19 | <0.01 |
| Biologicals (infliximab) | 0/8 | 2/14 | 15/29 | 0.06 |
| Supplements | 4/4 (no/yes) | 3/13 (no/yes) | 18/31 (no/yes) | 0.26 |
| Alkane phosphatase (ALP, mean) | 191 | 199 | 78 | <0.001 |
| Aspartate aminotransferase (AST, mean) | 40 | 52 | 27 | 0.027 |
| Alanine aminotransferase (ALT, mean) | 52 | 59 | 28 | 0.027 |
| Fecal water content (mean, %) | 74.02 | 80.33 | 73.79 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Vorstenbosch, R.; van Munster, K.; Pachen, D.; Mommers, A.; Stavropoulos, G.; van Schooten, F.-J.; Ponsioen, C.; Smolinska, A. The Detection of Primary Sclerosing Cholangitis Using Volatile Metabolites in Fecal Headspace and Exhaled Breath. Metabolites 2024, 14, 23. https://doi.org/10.3390/metabo14010023
van Vorstenbosch R, van Munster K, Pachen D, Mommers A, Stavropoulos G, van Schooten F-J, Ponsioen C, Smolinska A. The Detection of Primary Sclerosing Cholangitis Using Volatile Metabolites in Fecal Headspace and Exhaled Breath. Metabolites. 2024; 14(1):23. https://doi.org/10.3390/metabo14010023
Chicago/Turabian Stylevan Vorstenbosch, Robert, Kim van Munster, Danielle Pachen, Alex Mommers, Georgios Stavropoulos, Frederik-Jan van Schooten, Cyriel Ponsioen, and Agnieszka Smolinska. 2024. "The Detection of Primary Sclerosing Cholangitis Using Volatile Metabolites in Fecal Headspace and Exhaled Breath" Metabolites 14, no. 1: 23. https://doi.org/10.3390/metabo14010023
APA Stylevan Vorstenbosch, R., van Munster, K., Pachen, D., Mommers, A., Stavropoulos, G., van Schooten, F.-J., Ponsioen, C., & Smolinska, A. (2024). The Detection of Primary Sclerosing Cholangitis Using Volatile Metabolites in Fecal Headspace and Exhaled Breath. Metabolites, 14(1), 23. https://doi.org/10.3390/metabo14010023