
Effects of Pregnancy on Plasma Sphingolipids Using a Metabolomic and Quantitative Analysis Approach
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Metabolomics Analysis
2.3. Targeted Quantitative Analysis
2.4. Statistical Analysis
2.5. Sphingolipid Naming Conventions
3. Results
3.1. Characteristics of Subjects
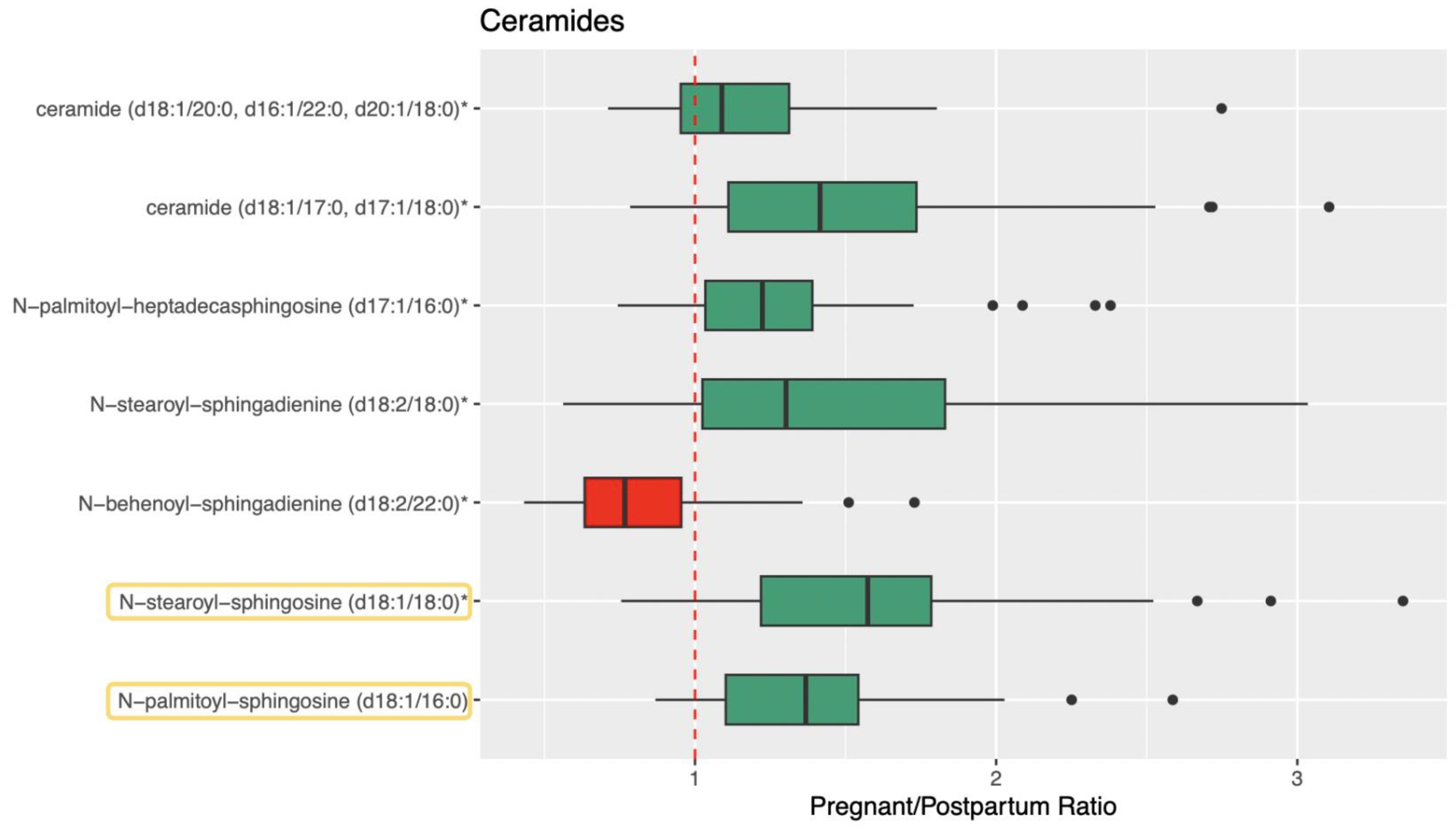
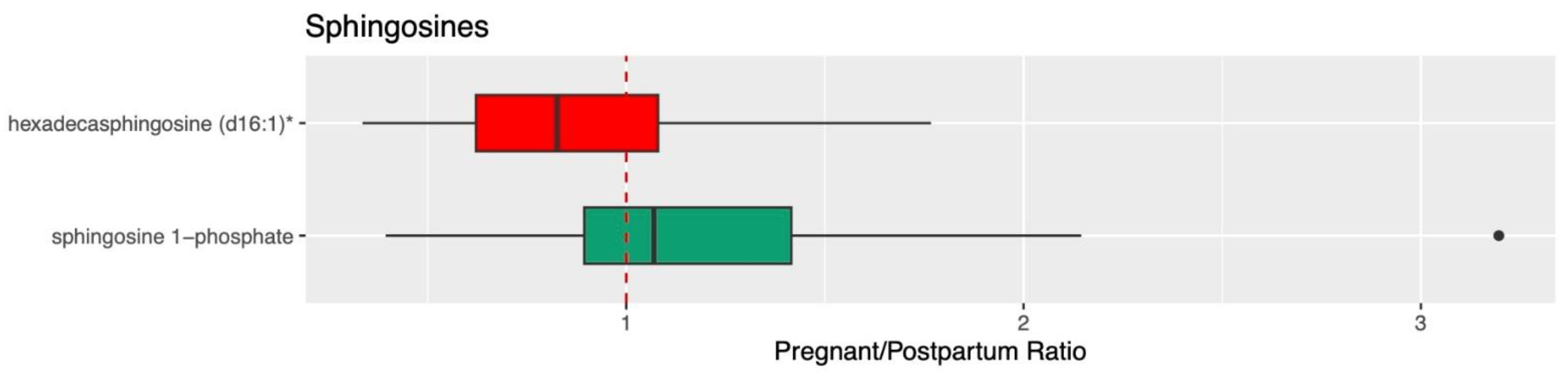
3.2. Ceramide, Sphingolipid, and Sphingosine
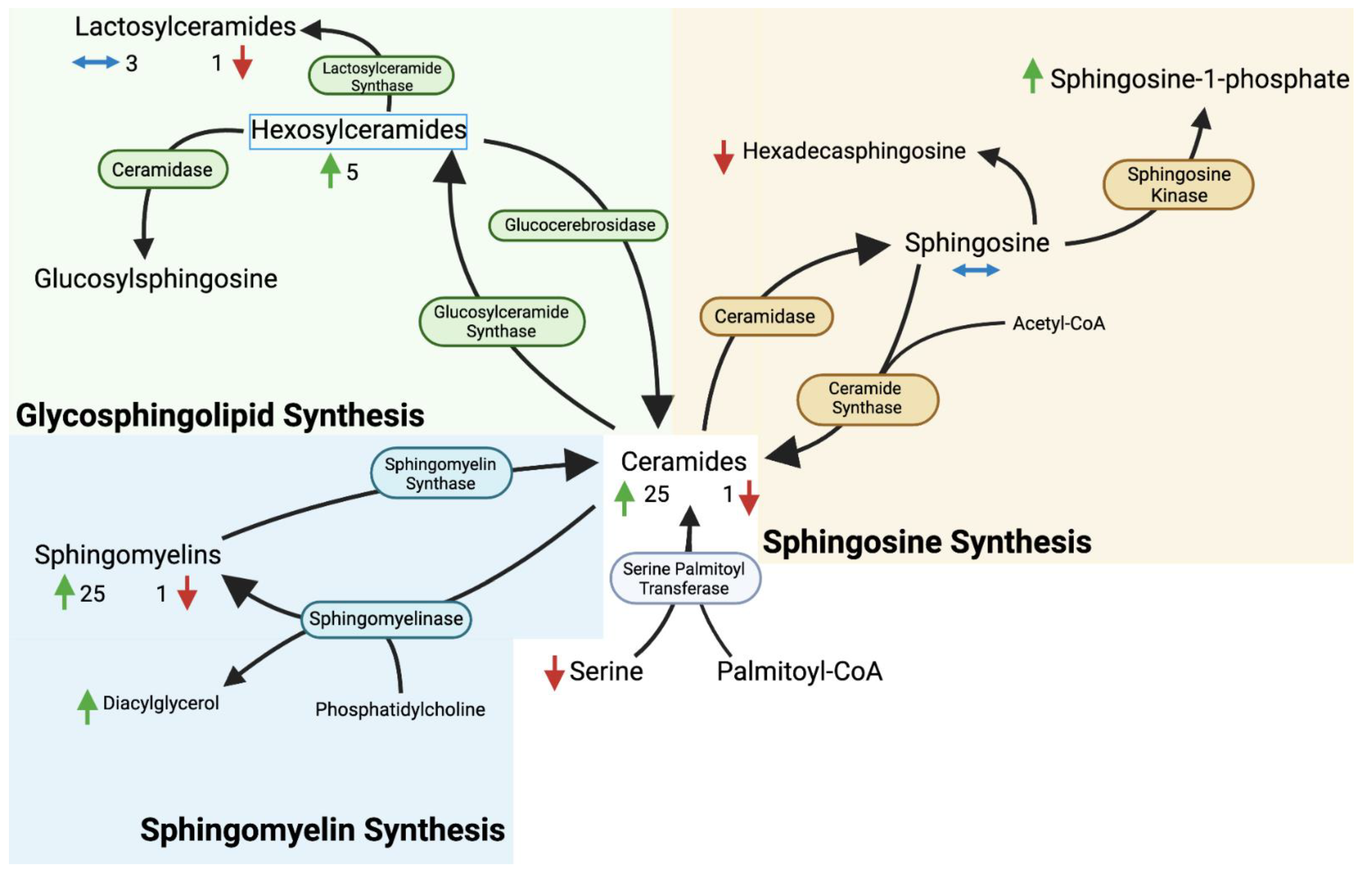
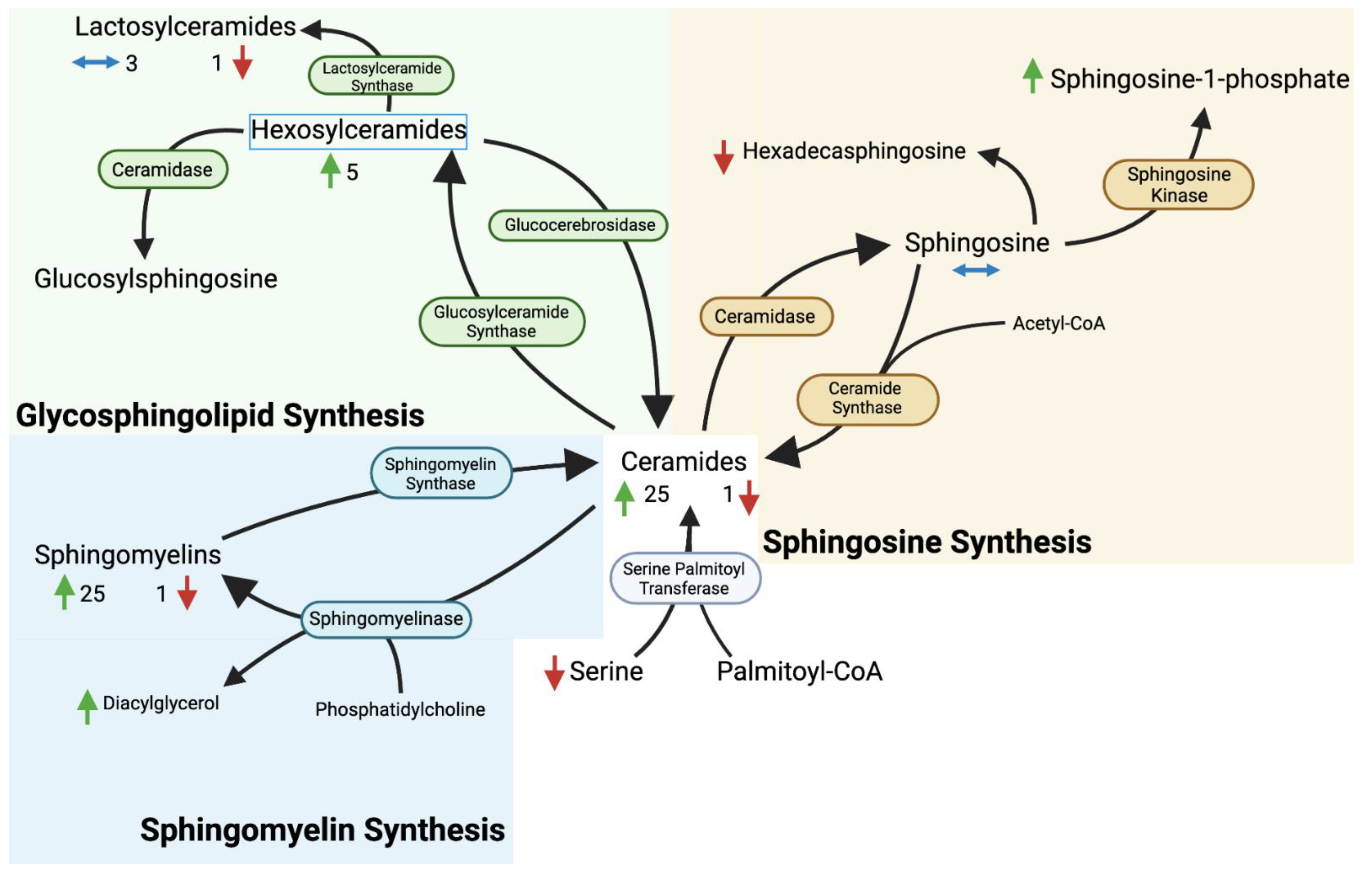
4. Discussion
4.1. Overview
4.2. Limitations
4.3. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soma-Pillay, P.; Nelson-Piercy, C.; Tolppanen, H.; Mebazaa, A. Physiological changes in pregnancy. Cardiovasc. J. Afr. 2016, 27, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Gault, C.R.; Obeid, L.M.; Hannun, Y.A. An overview of sphingolipid metabolism: From synthesis to breakdown. Adv. Exp. Med. Biol. 2010, 688, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Juchnicka, I.; Kuźmicki, M.; Zabielski, P.; Krętowski, A.; Błachnio-Zabielska, A.; Szamatowicz, J. Serum C18:1-Cer as a Potential Biomarker for Early Detection of Gestational Diabetes. J. Clin. Med. 2022, 11, 384. [Google Scholar] [CrossRef] [PubMed]
- Charkiewicz, K.; Goscik, J.; Blachnio-Zabielska, A.; Raba, G.; Sakowicz, A.; Kalinka, J.; Chabowski, A.; Laudanski, P. Sphingolipids as a new factor in the pathomechanism of preeclampsia—Mass spectrometry analysis. PLoS ONE 2017, 12, e0177601. [Google Scholar] [CrossRef]
- Dobierzewska, A.; Soman, S.; Illanes, S.E.; Morris, A.J. Plasma cross-gestational sphingolipidomic analyses reveal potential first trimester biomarkers of preeclampsia. PLoS ONE 2017, 12, e0175118. [Google Scholar] [CrossRef]
- Spijkers, L.J.; van den Akker, R.F.; Janssen, B.J.; Debets, J.J.; De Mey, J.G.; Stroes, E.S.; van den Born, B.-J.H.; Wijesinghe, D.S.; Chalfant, C.E.; MacAleese, L.; et al. Hypertension is associated with marked alterations in sphingolipid biology: A potential role for ceramide. PLoS ONE 2011, 6, e21817. [Google Scholar] [CrossRef]
- Yang, J.; Yu, Y.; Sun, S.; Duerksen-Hughes, P.J. Ceramide and other sphingolipids in cellular responses. Cell Biochem. Biophys. 2004, 40, 323–350. [Google Scholar] [CrossRef]
- Iqbal, J.; Walsh, M.T.; Hammad, S.M.; Hussain, M.M. Sphingolipids and Lipoproteins in Health and Metabolic Disorders. Trends Endocrinol. Metab. 2017, 28, 506–518. [Google Scholar] [CrossRef]
- Enthoven, L.F.; Shi, Y.; Fay, E.E.; Moreni, S.; Mao, J.; Honeyman, E.M.; Smith, C.K.; Whittington, D.; Brockerhoff, S.E.; Isoherranen, N.; et al. The Effects of Pregnancy on Amino Acid Levels and Nitrogen Disposition. Metabolites 2023, 13, 242. [Google Scholar] [CrossRef]
- Shum, S.; Yadav, A.; Fay, E.; Moreni, S.; Mao, J.; Czuba, L.; Wang, C.; Isoherranen, N.; Hebert, M.F. Infant Dextromethorphan and Dextrorphan Exposure via Breast Milk From Mothers Who Are CYP2D6 Extensive Metabolizers. J. Clin. Pharmacol. 2021, 62, 747–755. [Google Scholar] [CrossRef]
- Collet, T.H.; Sonoyama, T.; Henning, E.; Keogh, J.M.; Ingram, B.; Kelway, S.; Farooqi, I.S. A Metabolomic Signature of Acute Caloric Restriction. J. Clin. Endocrinol. Metab. 2017, 102, 4486–4495. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, R.N.; Yu, C.; Hoofnagle, A.; Hari, N.; Jensen, P.N.; Fretts, A.M.; Umans, J.G.; Howard, B.V.; Sitlani, C.M.; Siscovick, D.S.; et al. Circulating Sphingolipids, Insulin, HOMA-IR, and HOMA-B: The Strong Heart Family Study. Diabetes 2018, 67, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- MacLean, B.; Tomazela, D.M.; Shulman, N.; Chambers, M.; Finney, G.L.; Frewen, B.; Kern, R.; Tabb, D.L.; Liebler, D.C.; MacCoss, M.J. Skyline: An open source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 2010, 26, 966–968. [Google Scholar] [CrossRef] [PubMed]
- Storey, J.D.; Bass, A.J.; Dabney, A.; Robinson, D. Qvalue: Q-Value Estimation for False Discovery Rate Control, Version 2.28.0.; R Package: 2022. Available online: http://github.com/jdstorey/qvalue (accessed on 15 January 2023).
- Mejia, J.F.; Hirschi, K.M.; Tsai, K.Y.F.; Long, M.G.; Tullis, B.C.; Bitter, E.E.K.; Bikman, B.T.; Reynolds, P.R.; Arroyo, J.A. Differential placental ceramide levels during gestational diabetes mellitus (GDM). Reprod. Biol. Endocrinol. 2019, 17, 81. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.L.; Feng, Y.A.; Fiehn, O.; Albert, P.S.; Tsai, M.Y.; Zhu, Y.; Wang, X.; Tekola-Ayele, F.; Liang, L.; Zhang, C. Plasma lipidomics profile in pregnancy and gestational diabetes risk: A prospective study in a multiracial/ethnic cohort. BMJ Open Diabetes Res. Care 2021, 9, e001551. [Google Scholar] [CrossRef]
- Patanapirunhakit, P.; Karlsson, H.; Mulder, M.; Ljunggren, S.; Graham, D.; Freeman, D. Sphingolipids in HDL—Potential markers for adaptation to pregnancy? Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 158955. [Google Scholar] [CrossRef]
- Handelman, S.K.; Romero, R.; Tarca, A.L.; Pacora, P.; Ingram, B.; Maymon, E.; Chaiworapongsa, T.; Hassan, S.S.; Erez, O. The plasma metabolome of women in early pregnancy differs from that of non-pregnant women. PLoS ONE 2019, 14, e0224682. [Google Scholar] [CrossRef]
- Hytten, F. Blood volume changes in normal pregnancy. Clin. Haematol. 1985, 14, 601–612. [Google Scholar] [CrossRef]
- Fakhr, Y.; Brindley, D.N.; Hemmings, D.G. Physiological and pathological functions of sphingolipids in pregnancy. Cell Signal. 2021, 85, 110041. [Google Scholar] [CrossRef]
- Lantzanaki, M.; Veneti, S.; Mintziori, G.; Begou, O.; Pappas, P.D.; Gika, H.; Goulis, D.G.; Bili, H.; Taousani, E.; Vavilis, D. Plasma Ceramide Concentrations in Full-Term Pregnancies Complicated with Gestational Diabetes Mellitus: A Case-Control Study. Metabolites 2022, 12, 1123. [Google Scholar] [CrossRef]
- Jacquemyn, Y.; Osmanovic, F.; Martens, G. Preeclampsia and birthweight by gestational age in singleton pregnancies in Flanders, Belgium: A prospective study. Clin Exp Obs. Gynecol. 2006, 33, 96–98. [Google Scholar]
- Nofer, J.R.; van der Giet, M.; Tölle, M.; Wolinska, I.; Lipinski, K.v.W.; Baba, H.A.; Tietge, U.J.; Gödecke, A.; Ishii, I.; Kleuser, B.; et al. HDL induces NO-dependent vasorelaxation via the lysophospholipid receptor S1P3. J. Clin. Investig. 2004, 113, 569–581. [Google Scholar] [CrossRef]
- Zeigler, M.B.; Fay, E.E.; Moreni, S.L.; Mao, J.; Totah, R.A.; Hebert, M.F. Plasma hydrogen sulfide, nitric oxide, and thiocyanate levels are lower during pregnancy compared to postpartum in a cohort of women from the Pacific northwest of the United States. Life Sci. 2023, 322, 121625. [Google Scholar] [CrossRef]
- Khalil, A.A.; Cooper, D.J.; Harrington, K.F. Pulse wave analysis: A preliminary study of a novel technique for the prediction of pre-eclampsia. BJOG Int. J. Obstet. Gynaecol. 2009, 116, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Qu, T.; Wang, W.; Li, C.; Yang, X.; He, X.; Wang, Y.; Xing, G.; Xu, X.; Yang, L.; et al. Untargeted lipidomics analysis in women with intrahepatic cholestasis of pregnancy: A cross-sectional study. BJOG Int. J. Obstet. Gynaecol. 2022, 129, 880–888. [Google Scholar] [CrossRef] [PubMed]
- Calzada, C.; Vors, C.; Penhoat, A.; Cheillan, D.; Michalski, M.C. Role of circulating sphingolipids in lipid metabolism: Why dietary lipids matter. Front. Nutr. 2023, 9, 1108098. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Pregnant (n = 47) | Postpartum (n = 47) | p-Value |
|---|---|---|---|
| Gestational Age (weeks) or Time Postpartum (weeks) | 27.0 ± 1.3 | 15.0 ± 2.1 | NA |
| Height (cm) | 163.6 ± 16.8 | NA | |
| Weight (kg) | 71.6 ± 10.6 | 67.4 ± 9.3 | 1 × 10−5 |
| IBW (kg) | 58.2 ± 6.4 | NA | |
| BMI (kg/m2) | 25.7 ± 3.2 | NA | |
| Albumin (g/dL) | 3.6 ± 0.2 | 4.6 ± 0.2 | 9 × 10−28 |
| Bilirubin (mg/dL) | 0.5 ± 0.2 | 0.7 ± 0.4 | 7 × 10−7 |
| Serum Creatinine (mg/dL) | 0.5 ± 0.1 | 0.7 ± 0.1 | 9 × 10−21 |
| BUN (mg/dL) | 7.8 ± 1.9 | 14.0 ± 4.0 | 1 × 10−14 |
| CrCl (mL/min) | 194.3 ± 81.4 | 130.9 ± 25.6 | 3 × 10−6 |
| Sphingolipid | Pregnant (nM) | Postpartum (nM) | Pregnancy/ Postpartum | Corrected p-Value |
|---|---|---|---|---|
| HexCer (d18:1/16:0) | 120 ± 20 | 80 ± 20 | 1.56 ± 0.28 | 8.08 × 10−17 |
| Lactosylceramide (d18:1/16:0) | 540 ± 120 | 350 ± 70 | 1.54 ± 0.28 | 6.85 × 10−16 |
| HexCer (d18:1/20:0) | 17.7 ± 5 | 11.6 ± 3 | 1.57 ± 0.34 | 8.19 × 10−14 |
| HexCer (d18:1/18:0) | 16.9 ± 5 | 10.4 ± 3 | 1.68 ± 0.48 | 6.45 × 10−12 |
| HexCer (d18:1/22:0) | 140 ± 30 | 110 ± 20 | 1.40 ± 0.31 | 8.88 × 10−10 |
| Ceramide (d18:1/14:0) | 7.04 ± 2 | 5.06 ± 1 | 1.45 ± 0.41 | 2.44 × 10−9 |
| Palmitoyl sphingomyelin (d18:1/16:0) | 128,000 ± 15,800 | 111,000 ± 16,400 | 1.16 ± 0.14 | 2.00 × 10−8 |
| HexCer (d18:1/24:0) | 140 ± 30 | 110 ± 30 | 1.27 ± 0.24 | 9.92 × 10−8 |
| Stearoyl sphingomyelin (d18:1/18:0) | 4290 ± 780 | 3510 ± 670 | 1.24 ± 0.22 | 1.66 × 10−7 |
| Ceramide (d18:1/24:0) | 2300 ± 570 | 3030 ± 710 | 0.79 ± 0.21 | 3.84 × 10−7 |
| Sphingomyelin (d18:1/20:0) | 2270 ± 380 | 1880 ± 350 | 1.23 ± 0.21 | 4.01 × 10−7 |
| N-palmitoyl-sphingosine (d18:1/16:0) | 230 ± 40 | 190 ± 40 | 1.26 ± 0.26 | 1.19 × 10−6 |
| Ceramide (d18:1/20:0) | 60 ± 10 | 46.2 ± 10 | 1.37 ± 0.34 | 1.72 × 10−6 |
| N-stearoyl-sphingosine (d18:1/18:0) | 64 ± 20 | 45.7 ± 20 | 1.53 ± 0.52 | 1.76 × 10−6 |
| Sphingomyelin (d18:1/14:0) | 5300 ± 1230 | 4540 ± 1020 | 1.19 ± 0.22 | 4.99 × 10−5 |
| Ceramide (d18:1/22:0) | 400 ± 90 | 470 ± 140 | 0.91 ± 0.25 | 2.47 × 10−2 |
| Lactosylceramide (d18:1/24:0) | 66.3 ± 10 | 74.8 ± 20 | 0.91 ± 0.19 | 4.57 × 10−2 |
| Ceramide (d18:1/24:1) | 250 ± 50 | 220 ± 60 | 1.16 ± 0.28 | 7.51 × 10−2 |
| Sphingomyelin (d18:1/22:0) | 3150 ± 550 | 2880 ± 600 | 1.12 ± 0.21 | 0.207 |
| Lactosylceramide (d18:1/18:0) | 9.74 ± 3 | 9.03 ± 2 | 1.11 ± 0.27 | 0.769 |
| Sphingomyelin (d18:1/24:0) | 17,100 ± 3100 | 16,000 ± 3340 | 1.09 ± 0.21 | 1 |
| Lactosylceramide (d18:1/20:0) | 2.80 ± 1 | 25.6 ± 10 | 1.16 ± 0.41 | 1 |
| Lactosylceramide (d18:1/22:0) | 10.2 ± 3 | 10.7 ± 3 | 1.00 ± 0.34 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enthoven, L.F.; Shi, Y.; Fay, E.; Kim, A.; Moreni, S.; Mao, J.; Isoherranen, N.; Totah, R.A.; Hebert, M.F. Effects of Pregnancy on Plasma Sphingolipids Using a Metabolomic and Quantitative Analysis Approach. Metabolites 2023, 13, 1026. https://doi.org/10.3390/metabo13091026
Enthoven LF, Shi Y, Fay E, Kim A, Moreni S, Mao J, Isoherranen N, Totah RA, Hebert MF. Effects of Pregnancy on Plasma Sphingolipids Using a Metabolomic and Quantitative Analysis Approach. Metabolites. 2023; 13(9):1026. https://doi.org/10.3390/metabo13091026
Chicago/Turabian StyleEnthoven, Luke F., Yuanyuan Shi, Emily Fay, Agnes Kim, Sue Moreni, Jennie Mao, Nina Isoherranen, Rheem A. Totah, and Mary F. Hebert. 2023. "Effects of Pregnancy on Plasma Sphingolipids Using a Metabolomic and Quantitative Analysis Approach" Metabolites 13, no. 9: 1026. https://doi.org/10.3390/metabo13091026
APA StyleEnthoven, L. F., Shi, Y., Fay, E., Kim, A., Moreni, S., Mao, J., Isoherranen, N., Totah, R. A., & Hebert, M. F. (2023). Effects of Pregnancy on Plasma Sphingolipids Using a Metabolomic and Quantitative Analysis Approach. Metabolites, 13(9), 1026. https://doi.org/10.3390/metabo13091026