Abstract
Taurine, the end product in the sulfur-containing amino acid pathway, is conjugated with bile acids (BAs) in the liver. The rate-limiting enzymes in both taurine synthesis and BA conjugation may be regulated by a nucleus receptor, FXR, that promotes BA homeostasis. However, it is controversial because BAs act as natural FXR agonists or antagonists in humans and mice, respectively, due to the species differences in BA synthesis. The present study evaluated the influences of different BA compositions on both pathways in the liver by comparing Cyp2a12−/−/Cyp2c70−/− mice with a human-like BA composition (DKO) and wild-type (WT) mice. The DKO liver contains abundant natural FXR agonistic BAs, and the taurine-conjugated BA proportion and the taurine concentration were significantly increased, while the total BA concentration was significantly decreased compared to those in the WT liver with natural FXR antagonistic BAs. The mRNA expression levels of the enzymes Bacs and Baat in BA aminations and Cdo and Fmo1 in the taurine synthesis, as well as Fxr and its target gene, Shp, were significantly higher in the DKO liver than in the WT liver. The present study, using a model with a human-like BA composition in the liver, confirmed, for the first time in mice, that both the taurine synthesis and BA amidation pathways are upregulated by FXR activation.
1. Introduction
Taurine (2-aminoethanesulfonic acid) is abundant in various cells and tissues in mammals. This is due to dietary intake and its biosynthesis from sulfur-containing amino acids, methionine and cysteine, which involves the rate-limiting enzymes, cysteine dioxygenase (CDO) [1] and cysteine sulfinate decarboxylase (CSD) (Figure 1C) [2,3,4,5]. Taurine has been reported to have many physiological and pharmacological roles with regard to maintaining various biological forms of homeostasis [6,7,8,9,10,11]. Among them, the most established and well-known role is the conjugation with bile acids (BAs) [12,13]. Bile acids are metabolized from cholesterol in the liver. Newly synthesized cholic acid (CA) and chenodeoxycholic acid (CDCA), the primary BAs in humans, are then conjugated at the C24 carboxyl group with taurine or glycine (Figure 1B) [14]. Through the conjugation with amino acids (amidation), the polarity of BAs is increased, and, consequently, the excretion into the bile, the formation of bile micelle, and the intestinal absorption of lipids and lipid-solved vitamins are promoted. In addition, the cytotoxicity of hydrophobic BAs is reduced [13,15]. The excreted primary BAs conjugated with taurine or glycine in the intestine are deconjugated by intestinal bacteria, and then converted to the secondary BAs, deoxycholic acid (DCA) and lithocholic acid (LCA) (from CA and CDCA, respectively), following dehydroxylation at the 7α-hydroxy group by intestinal bacteria (Figure 1A) [16]. The deconjugated BAs are conjugated again with the amino acids in the liver following intestinal absorption in the enterohepatic circulation. The BA amidation is carried out in the peroxisome and endoplasmic reticulum through sequential enzyme reactions involving two limiting enzymes: ATP-dependent microsomal BA coenzyme A (CoA) synthetase (BACS), which converts a BA to an acyl-CoA thioester; and BA-CoA:amino acid N-acetyltransferase (BAAT), which transfers the acyl-CoA thioester to taurine or glycine (Figure 1B) [17,18]. Almost all BAs are conjugated with taurine in rodents [19], while the ratio of taurine- and glycine-conjugation is 1:3~3.5 in humans [20,21].
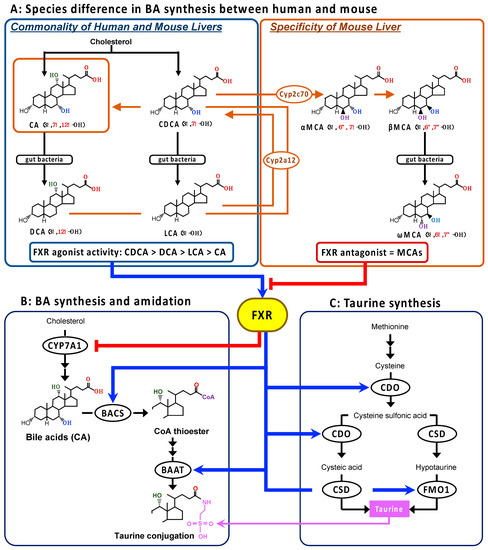
Figure 1.
The species differences in BA synthesis in the liver between humans and mice (A), and the FXR regulation of the BA synthesis, BA amidation, and taurine synthesis pathways (B,C). The primary BAs, CA and CDCA, are metabolized from cholesterol and then further converted to the secondary BAs, DCA and LCA, respectively, by the intestinal bacteria in humans. In the mouse liver, CDCA is further metabolized to MCAs by Cyp2c70, and the secondary BAs are converted back to the respective primary BA by Cyp2a12. Consequently, CA and MCAs are the main BAs in mice. The mouse-specific reactions are shown in orange. The human-type BAs act as endogenous FXR agonists, and the order of ligand activity is CDCA > DCA > LCA > CA. On the other hand, MCAs behave as endogenous FXR antagonists. The gene expression of the rate-limiting enzymes in BA synthesis, BA amidation, and taurine synthesis might be regulated by FXR. Blue arrow and red line with head show the acceleration and inhibition, respectively. The orange line and the arrow show the BAs in the mouse liver and the mouse-specific metabolic pathway. Abbreviations: BA, bile acid; BAAT, bile acid-coenzyme A:amino acid N-acetyltransferase; BACS, ATP-dependent microsomal bile acid coenzyme A synthetase; CA, cholic acid; CDCA, chenodeoxycholic acid; CDO, cysteine dioxygenase; CSD, cysteine sulfinate decarboxylase; CYP7, CYP, cytochrome P450; DCA, deoxycholic acid; FMO1, flavin containing monooxygenase 1; FXR, farnesoid X receptor; LCA, lithocholic acid; MCA, muricholic acid.
Bile acids play the role of the endogenous agonist of a nuclear receptor, farnesoid X receptor (FXR; NR1H4), to regulate the transcription of associated genes in the hepatic synthesis and enterohepatic circulation of BAs to avoid BA accumulation in the liver [22,23]. It is also involved in the downregulation of microsomal cholesterol 7α-hydroxylase (CYP7A1), the rate-limiting enzyme in the BA synthetic pathway (Figure 1B), through the upregulation of the atypical nuclear receptor, small heterodimer partner (SHP; NR0B2) [24], the upregulation of ABC transporter family including the bile salt export pump (BSEP; ABCB11), which actively excretes BAs into the bile canaliculi [24], and the downregulation of the basolateral sodium/taurocholate co-transporter peptide (NTCP; SLC10A1), which transports BAs into the liver from the portal vain [24].
In addition, the rate-limiting enzyme genes in BA amidation, BACS and BAAT, have been shown to be the direct target of FXR (Figure 1B) [25,26]. Pircher et al. showed that both BACS and BAAT genes are positively regulated via inverted repeat-1 elements cognate to the FXR in human and rat hepatocytes [26]. The increased transcriptional levels of both genes were also confirmed in the liver of rats treated with the synthetic FXR agonist GW4064 (3-[2-[2-chloro4-[[3-3(2,6-dichlorophenyl)-5-(1-methylethyl)-4-isoxazolyl]methoxy]phenyl]ethenyl]benzoic acid) [26]. Therefore, it makes sense that both the rate-limiting enzymes in BA amidation and taurine synthesis are positively regulated by FXR activation, based on the role of FXR in accelerating BA excretion from the liver tissue.
On the other hand, in vivo studies using mice showed that the Baat and Csd gene expression levels were significantly reduced by GW4064 treatment, suggesting that the taurine synthetic enzymes are negatively regulated by the FXR–SHP axis [27]. Thus, there is a discrepancy in mice with regard to the responses of BA amidation and taurine synthesis to the role of FXR on BA homeostasis. A possible reason for this is the species difference, particularly between humans and mice, in BA metabolism [28]. In the liver, CA and CDCA are the end-product, as the primary BAs in humans, while CDCA is further metabolized to muricholic acids (MCAs) by Cyp2c70 in mice (Figure 1A) [28]. Moreover, the secondary BAs are converted by Cyp2a12 back to their respective primary BAs in the mouse, but not the human, liver (Figure 1A) [28]. The potency order of the FXR agonist activity among the natural BAs is CDCA > DCA > LCA > CA [29,30], but MCAs act as FXR antagonists [31]. Thus, the hepatobiliary concentrations of BAs with more potent FXR agonists are almost deficient, whereas the concentrations of natural FXR antagonistic MCAs are abundant in mice.
In order to eliminate the species difference in the BA metabolism with regard to humans, we created a mouse model with human-like BA composition by double knockouts of Cyp2a12 and Cyp2c70 (DKO) [28,32,33,34,35]. The purpose of this study is to evaluate the influences of alteration in the composition of human-type BAs with higher FXR agonist activity on BA amidation and taurine biosynthesis by comparing the DKO mouse to the wild-type (WT) mouse, which has endogenous FXR antagonistic MCAs.
2. Materials and Methods
2.1. Materials
Cholic acid, glycocholic acid (GCA), taurocholic acid (TCA), CDCA, glycochenodeoxycholic acid (GCDCA), taurochenodeoxycholic acid (TCDCA), DCA, glycodeoxycholic acid (GDCA), taurodeoxycholic acid (TDCA), LCA, glycolithocholic acid (GLCA), taurolithocholic acid (TLCA), ursodeoxycholic acid (UDCA), glycoursodeoxycholic acid (GUDCA), αMCA, βMCA, ωMCA, tauro-αMCA (TαMCA), tauro-βMCA (TβMCA), and tauro-ωMCA (TωMCA) were obtained from Steraloids, Inc. (Newport, RI, USA). [2H4]cholic acid, [2H4]DCA, and [2H4]LCA were purchased from C/D/N Isotopes, Inc. (Pointe-Claire, QC, Canada). [2H4]taurocholic acid was obtained from Merck, KGaA (Darmstadt, Germany). [2H4]chenodeoxycholic acid was supplied from the Research Laboratory of Nippon Kayaku Co. (Toyo, Japan). Tauroursodeoxycholic acid (TUDCA), [2H4]UDCA, and [2H4]TUDCA were supplied by Tokyo Tanabe Company (Tokyo, Japan).
3-Aminopyridyl-N-hydroxysuccinimidyl carbamate (APDS), acetonitrile, amino acids mixture standard solutions (Type B, AN-2), ammonium acetate, APDSTAG Wako Amino Acids Internal Standard (IS) mixture solution, APDSTAG Wako Eluent, dicalcium phosphate, ethanol, formic acid, methanol, potassium phosphate, sodium borate buffer, sucrose, and taurine were purchased from FUJIFILM Wako Pure Chemical Corporation (Osaka, Japan).
The RNeasy Plus Mini Kit was purchased from Qiagen K.K. (Tokyo, Japan). The reverse transcription using the PrimeScript RT reagent kit and the mouse housekeeping gene primer set were purchased from Takara Bio, Inc. (Shiga, Japan). The FastStart DNA Master SYBR Green I and the LightCycler system were obtained from Roche Diagnostics (Mannheim, Germany).
2.2. Animals
Wild-type C57BL/6J mice (male, body weight 26.9 ± 0.6 g, n = 8) and DKO (Cyp2a12−/− and Cyp2c70−/−; male, 28.3 ± 0.5 g, n = 8) mice were used according to our previous studies [28,32,33]. The mice were bred and kept until 20 weeks of age at the Jackson Laboratory Japan, Inc. (Ishioka, Japan) under a regular 12 h light–dark cycle (6:00–18:00) with regular food (CRF-1, Oriental Yeast Co., Ltd., Tokyo, Japan) and water ad libitum in pathogen-free conditions. After fasting overnight with free access to water, mice were euthanized by exsanguination under combination anesthesia with medetomidine, midazolam, and butorphanol. Liver tissue was collected and frozen at –80 °C until analysis.
2.3. BA Analysis
Liver tissue was homogenized with ice-cold ten-times volume of phosphate-buffered saline (PBS), and the supernatant was collected after centrifugation at 3500× g for 10 min at 4 °C. In the supernatant, BAs were measured using a high-performance liquid chromatography–electrospray ionization tandem mass spectrometry (HPLC-ESI-MS/MS) system according to the previous reports [28,36]. In brief, 200 µL of the supernatant was mixed with 20 µL of IS mixture (41.6 ng of [2H4]CA, 57.5 ng of [2H4]CDCA, 32.8 ng of [2H4]DCA, 22.4 ng of [2H4]LCA, 34.4 ng of [2H4]UDCA, 34.4 ng of [2H4]TUDCA, 100 ng of [2H3]TCA in acetonitrile), and 2 mL of 0.5 M potassium phosphate buffer (pH 7.4). The sample was eluted using Bond Elut C18 cartridges (200 mg; Agilent Technologies, Santa Clara, CA, USA) and was evaporated at 100 °C under a nitrogen stream. Then, the dried residue was redissolved in 20 mM ammonium acetate–methanol buffer (pH 7.5) and centrifugated at 12,000× g for 1 min. An aliquot of the supernatant was injected into the HPLC-ESI-MS/MS system for analysis. For further information on the extraction of BAs, see the previous study [37].
2.4. Amino Acid Analysis
Amino acids in the liver, including taurine, methionine, and cysteine, were measured by a derivatization method with APDS using the HPLC-ESI-MS/MS system according to the previously reported method [38,39]. In brief, 50 µL of the supernatant of liver tissue homogenized with PBS was mixed with an equal volume of APDSTAG IS solution and two-times volume of acetonitrile. After centrifugation at 20,000× g for 10 min, 20 µL of the supernatant was mixed with an equal volume of APDS-acetonitrile solution (20 mg/mL) and three-times volume of 0.2 M sodium borate buffer (pH 8.8), and then incubated at 55 °C for 10 min. Next, the reaction mixture was added to 100 μL of formic acid solution (0.1% in water), and 5 μL was used for injection into the HPLC-ESI-MS/MS system. For further information on the general HPLC and MS/MS conditions, see the previous studies [38,40].
2.5. Total RNA Extraction and RT-PCR Analysis
Fifty mg of the liver tissue was homogenized with a 10-times volume of lysis buffer, and then total RNA was extracted using the RNeasy Plus Mini Kit. Five-handled ng of total RNA was reverse-transcribed to cDNA using the PrimeScript RT reagent kit. The mRNA expressions of Fxr, Shp, Cyp7a1, Cdo, Csd, Bacs, Baat, favin containing monooxygenase (Fmo) 1, Fmo3, Bsep, Ntcp, and hepatocyte nucleus factor 4α (Hnf4α) were quantified with real-time quantitative PCR using gene-specific primers (Table 1) in the FastStart DNA Master SYBR Green I and the LightCycler system (Roche Diagnostics). PCR amplification began with a 10 min preincubation step at 95 °C, followed by 40 cycles of denaturation at 95 °C for 10 s, annealing at 62 °C for 10 s, and elongation at 72 °C. The relative concentration of the PCR products derived from the target gene was calculated using the LightCycler System software. A standard curve for each run was constructed by plotting the crossover point against the log concentration. The concentration of target molecules in each sample was then calculated automatically with reference to this curve (r = −1.00), and the specificity of each PCR product was assessed by melting curve analysis. The mRNA expression of each gene was standardized to the expression of Ywhaz as a housekeeping gene that was selected from the mouse housekeeping gene primers including Atp5f1, B2m, Hprt1, Rplp1, Ppia, Rps18, Pgk1, Gusb, Tbp, Actb, Tfrc, Ywhaz, 18SrRNA, and Gapdh by geNorm and Bestkeeper algorisms (Supplementary Table S1) [41].

Table 1.
Gene sequences of the PCR primers.
2.6. Statistical Analysis
Statistical analysis was carried out using Jump, and the threshold for significant differences was set at a p-value of 0.05; the significance was assessed using unpaired Student’s t-tests. Data are expressed as the mean ± standard error (SE).
3. Results
3.1. Bile Acid Concentrations in the Liver
Unconjugated and taurine-conjugated forms of all the BA types are shown in Figure 2A. In the WT group, MCAs (αMCA, βMCA, and ωMCA), CA, and DCA of both forms were abundantly contained in the liver, but the CDCA, DCA, LCA, and UDCA levels were so low as to be undetectable (Figure 2A). On the other hand, MCAs of both forms were undetectable, while CDCA and LCA, in addition to CA and DCA, were found in the DKO liver (Figure 2A). Ursodeoxycholic acid level was still very low in the DKO group. In the DKO group, both unconjugated and taurine-conjugated CA concentrations were significantly lower than in the WT group, while the taurine-conjugated forms of other human-type BAs, TCDCA, TDCA, and TLCA, were significantly higher compared to those in the WT group (Figure 2A). Any types of BAs conjugated with glycine were undetectable in the liver of both groups.
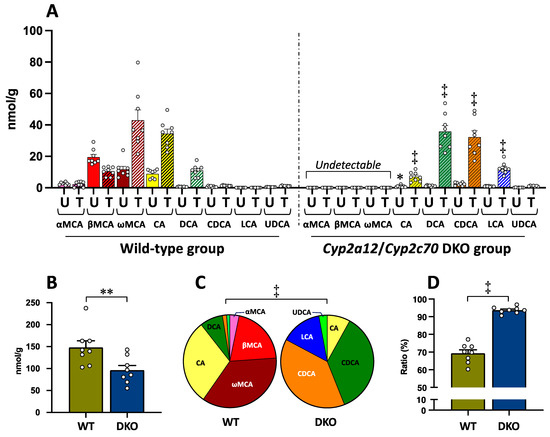
Figure 2.
Bile acid concentrations in the liver of the WT and DKO groups. (A) Concentration of all BA types conjugated with and without taurine. Data are the mean ± SE. The white circle in the column shows the individual value. * p < 0.05, ‡ p < 0.0001 compared to the respective BA in the WT group by two-way ANOVA post hoc Bonferroni’s multiple comparison test. Abbreviations: U, unconjugated form (plane column); T, taurine-conjugated form (striped column). (B) TBA concentration (the sum of all BA types of unconjugated and taurine-conjugated forms). Data are the mean ± SE. ** p < 0.01 according to unpaired Student’s t-test. (C) BA composition (the sum of unconjugated and taurine-conjugated forms). Pie graph was constructed using the mean of each BA. ‡ p < 0.0001 according to chi-square test. (D) The ratio of taurine-conjugated BAs. Data are the mean ± SE. ‡ p < 0.0001 according to unpaired Student’s t-test.
Total BA (TBA) concentration (the sum of all BA types with unconjugated and taurine-conjugated forms) in the liver was significantly decreased in the DKO group as compared to the WT group (Figure 2B).
Figure 2C shows the BA composition consisting of each BA type (the sum of unconjugated and taurine-conjugated forms) in the liver of both groups. There was a significant difference in the BA composition between the two groups (Figure 2C). In the WT group, the ratios of MCAs (the sum of αMCA, βMCA, and ωMCA) and human-type BAs (the sum of CA, CDCA, DCA, LCA, and UDCA) were 59.7 ± 1.6% and 40.3 ± 1.7%, respectively. On the other hand, all of the BA compositions were changed to the human-type BAs in the DKO group. Particularly, the ratios of DCA, CDCA, and LCA were 8.0 ± 0.8%, 1.3 ± 0.2%, and 0.05 ± 0.01%, respectively, in the WT group, while they were increased to 38.6 ± 0.9%, 35.7 ± 0.7%, and 14.2 ± 0.9%, respectively, instead of the disappearance of MCAs, in the DKO group (Figure 2C). The BA compositions in the DKO liver (Figure 2C) became similar to those of normal liver tissue in humans previously reported (CA 28.7%, CDCA 46.4%, DCA 15.7%, LCA 4.5%, and UDCA 4.6%) [42].
As shown in Figure 2D, the proportion of taurine-conjugated BAs (the sum of all BA types) in the liver was 69.3 ± 2.1% in the WT group; on the other hand, the proportion was significantly increased to 93.7 ± 0.7% in the DKO group.
3.2. Amino Acid Concentrations in the Liver
Figure 3 shows taurine and its precursor amino acids in the biosynthesis pathway, methionine and cysteine, in the livers of both groups. Taurine concentration was significantly higher in the DKO group than in the WT group (Figure 3). On the other hand, methionine concentration was significantly lower in the DKO group than in the WT group (Figure 3). Similarly, cysteine concentration tended to be lower, but not significantly, in the DKO group than in the WT group (Figure 3).
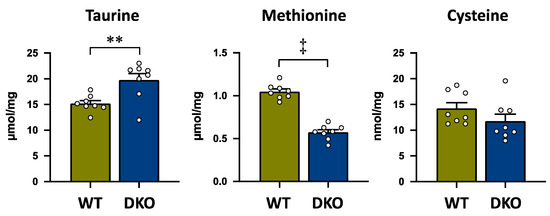
Figure 3.
Taurine and the sulfur-containing amino acids, methionine and cysteine, which are the precursors of taurine; concentrations in the liver of both groups. The data are presented as the mean ± SE. The white circle in the column shows the individual value. ** p < 0.01, and ‡ p < 0.0001 by unpaired Student’s t-test.
3.3. The mRNA Expression Levels in the Liver
The mRNA expressions of Fxr and its direct target gene, Shp, were significantly increased in the DKO group as compared to the WT group, but there was no difference in Cyp7a1 mRNA between both groups (Figure 4). The mRNA expressions of Bacs and Baat, the rate-limiting enzymes in BA amidation, were also significantly higher in the DKO group compared to those in the WT group. In the rate-limiting enzymes of the taurine synthesis pathway, the mRNA expression level of Cdo was significantly higher in the DKO group than in the WT group, while there was no difference in Csd mRNA expression between both groups. The mRNA expression levels of Fmo1, which is also a key enzyme in taurine biosynthesis [42], and Fmo3, which is the direct FXR target gene [43], were significantly higher in the DKO group compared to those in the WT group. In addition, the mRNA expression levels of Hnf4α (NR2A1), which is a nucleus receptor, and Bsep, which is an efflux BA transporter, were significantly higher in the DKO group than in the WT group, while there was no significant difference in Ntcp, which is an influx BA transporter, between both groups.
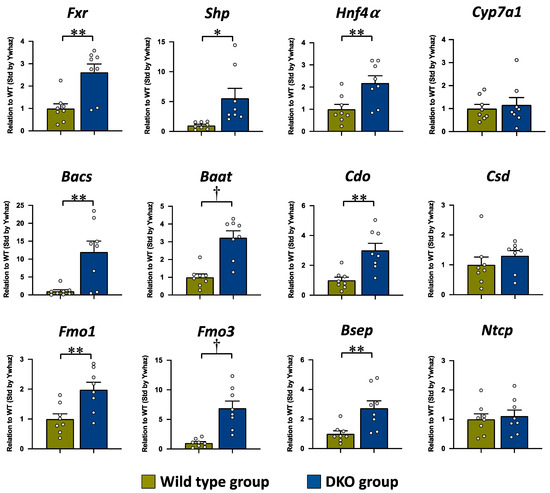
Figure 4.
The mRNA expressions of Fxr and its target genes involved in BA synthesis, BA amidation, taurine synthesis, and BA transport in the liver tissue. The mRNA expression level is expressed in relation to that of the WT group, after standardization by Ywhaz. The data are presented as the mean ± the SE. The white circle in the column shows the individual value. * p < 0.05, ** p < 0.01, and † p < 0.001 according to unpaired Student’s t-test.
4. Discussion
The present study evaluated the alterations of BA amidation and taurine biosynthesis properties in the livers of the DKO mice, which have abundant FXR agonist human-type BAs, in comparison to the WT mice, which have FXR antagonistic BAs. As shown in our previous studies [28,33], the BA composition in the liver was completely changed to human-type BAs by the DKO of Cyp2C70 and Cyp2a12, which are the enzymes that convert CDCA to αMCA, and the secondary BAs to the primary BAs. In the DKO liver, the ratio of CDCA and DCA, which have the most and second highest FXR agonist activity, was increased up to approximately 40%, while the WT liver was occupied by MCAs, which are the endogenous FXR antagonist, and CA, which has little activity as an FXR agonist. Upon the change to a human-like BA composition, the proportion of taurine-conjugated BA in the DKO liver reached over 90%, a 20% point increase compared to the WT liver. Furthermore, the taurine concentration was significantly increased by 25% in the DKO liver compared to the WT liver.
In the evaluation of the mRNA expression levels, the significant increase in Shp in the DKO liver implies that the FXR was activated by the increase in endogenous FXR agonists CDCA and DCA. This FXR activation should induce the enhancements of BA conjugation with taurine and taurine biosynthesis reactions through the upregulations of FXR target genes involving these reactions. In BA amidation, the mRNA expressions of Bacs and Baat were significantly increased in the DKO liver; particularly, the increase in the Bacs gene was markedly higher than the Baat expression. Both Bacs and Baat have been shown to be the direct target of FXR in human and rat hepatocytes, and the upregulations of them were induced in the synthetic FXR agonist GW4064-treated rat liver [26]. The present results agree with the previous observations of Pircher et al. [26]. Furthermore, the mRNA expression of Bsep, which is an efflux BA transporter, was significantly increased in the DKO liver. According to the increased BA-taurine conjugation ratio, along with the upregulation of mRNA expressions of the BA amidation enzymes as well as the efflux BA transporter, the significantly decreased TBA concentration in the DKO liver was highly suggestive of the enhancement of efficient biliary BA excretion from the liver. In the taurine biosynthesis pathway, the present study showed significantly increased mRNA expression levels of Cdo, but not Csd, in the DKO liver, and this can be considered to be related to the significant increase in taurine concentration in the DKO liver. From this result, the gene expression of Cdo is suggested to be upregulated by a direct or indirect FXR–SHP axis pathway. In the Cyp2c70 single KO mice with higher CDCA concentration, but not in the Cyp2a12 single KO mice with MCAs, reported in our previous study [28], the increases of BA-taurine conjugation ratio and mRNA levels of these enzymes in BA amidation and taurine synthesis were also observed in the liver, supporting the positive regulations on the reactions by FXR activation (Supplementary Figure S1).
However, the previous study reported opposite responses with regard to the gene expressions in BA–taurine conjugation and taurine biosynthesis in the WT mice treated with GW4064, i.e., the significantly decreased mRNA expression levels of Baat and Csd in the synthetic FXR-agonist-treated liver [27]. A possible reason for the opposite response to the FXR agonist in the WT mouse liver as compared to the present and previous findings [26] might be related to the balance of the endogenous FXR agonists and antagonists that were permanently present. The opposite results for these gene expressions under GW4064 treatment between mice [27] and rats [26] were likely due to the different ratios of MCA and CDCA contents in the liver, because rats do not metabolize all CDCA to MCA, and the amount of CDCA is still maintained to the MCA level in rats [44]. However, further study is needed to clarify how the different balances of endogenous FXR agonists and antagonists influence the response of gene expressions to the synthetic FXR agonist.
Unexpectedly, the Fxr mRNA level was significantly higher in the DKO liver than in the WT liver. Although the regulation of Fxr transcription has been clarified, chronic treatment of the semisynthetic FXR agonist 6α-ethyl CDCA (INT-747, obeticholic acid) has been reported to increase Fxr expression in the penile tissue of rabbits and rats [45]. Therefore, there is a possibility that Fxr transcription might be ligand-dependent and positively regulated. However, the increased Fxr mRNA expression was not found in the Cyp2c70 single KO liver [28], and it is unclear if the Fxr transcriptional system in the mouse liver has a similar system in the Fxr transcription to the penile tissue in rabbits and rats. Thus, this point also requires more investigation.
In the DKO liver, the taurine precursor amino acids, methionine and cysteine, were lower, while taurine was higher, compared to those in the WT liver. The decreased level of these amino acids might be due to the enhanced synthesis of taurine. Recently, Veeravalli et al. reported that the subtype 1 of FMO (FMO1) is the new candidate to be the key enzyme in the taurine synthesis pathway to oxidize hypotaurine to taurine (Figure 1) [42]. In the DKO liver, Fmo1 and another subtype, Fmo3, expressions were significantly higher compared to those in the WT liver. Flavin-containing monooxygenase type 3 is the direct FXR target gene and oxidizes trimethylamine (TMA), which is metabolized from choline and carnitine in the gut by intestinal bacteria, to trimethylamine oxide in the liver [46]. It is notable that methionine is the metabolic source of the methyl group donor in many metabolic pathways of trimethyl products, including choline and carnitine, and is not only used in taurine biosynthesis. Therefore, the significant decrease in methionine is likely due to the utilization of other metabolites such as choline and carnitine. Similar to Fmo3, Fmo1 is suggested to be also regulated by FXR and might contribute to the significant increase of taurine concentration in the DKO liver. However, further studies are needed to clarify the details of the regulation of Fmo1 by FXR.
The DKO mouse with the human-like BA composition is considered to be a model that extrapolates FXR regulation on BA amidation and taurine synthesis in humans. However, there is a species difference in the taurine synthesis property; rodents can highly synthesize taurine, while taurine synthesis is very low in humans [47]. If taurine biosynthesis is promoted by FXR activation, a question arises as to why taurine synthesis is high in mice with MCAs. The reason is still unclear, but there is a high possibility that the taurine synthesis pathway is mainly regulated by another uncertain mechanism, in addition FXR, under the presence of MCAs. However, it is also certain that the taurine synthesis pathway might be additionally promoted by FXR activation in liver under human-like BA compositions.
In addition, the proportion of BA amidation is different among animal species; all BAs in mice are conjugated with taurine, while the conjugation ratio with taurine and glycine is 1:3~3.5 in humans [20,21]. In humans, the reason for the low taurine conjugation ratio is considered to be due to the lower taurine pool in the liver compared to mice, and the taurine pool in humans is capable of only one quarter of the conjugation demand of BAs in the liver [21]. On the other hand, there is no species difference in the affinity of BAAT to taurine, and the affinity of BAAT in the human liver is higher with taurine than with glycine [48]. Furthermore, the ratio of conjugation with taurine is increased by taurine supplementation even in humans, while glycine supplementation does not increase the ratio conjugated with glycine [13,21]. Therefore, because the determining factor of the lower ratio of BA–taurine conjugation in humans is the lower taurine pool, the proportion of BA conjugation with taurine would be enhanced by the promotion of taurine synthesis through FXR activation. Because the conjugation with taurine increases the hydrophilicity of the BA molecule, as opposed to the conjugation with glycine [49], and because taurine itself has many beneficial effects on various tissues and cells [6,7,8,9,10,11,25], the increase in the taurine pool would lead to important physiological and pathological effects on bile excretion, cholesterol reduction, and hepatic protection. Recently, the relationship between taurine level in the body and healthy life span has been reported in the study of various animals including humans [50]; taurine abundance decreased during aging and the prevention of this decline by taurine supplementation increased health span and lifespan in the monkeys and rodents. Because the decrease of taurine synthetic ability is a high possible reason for the age-related decline of taurine pool, clarification of the regulation of taurine synthesis contributes healthy life and longevity. In the clinic, therapeutic effects for BA analogs and synthetic FXR agonists on various liver injury and metabolic disorders, including primary biliary cholangitis, nonalcoholic fatty liver disease, nonalcohol hepatitis, and lipodystrophy, have been reported [51]. Considering the present results, the enhancement of BA amidation and taurine synthesis might contribute to the therapeutic effects of these FXR agonists.
In the present and our previous studies [28], the mRNA expressions of Cyp7a1 and Ntcp, which are negatively regulated by FXR, were not reduced, but the FXR–SHP pathway was activated, in the DKO liver. Competitively to the downregulations by FXR activation, the transcriptions of Cyp7a1 [52,53,54] and Ntcp [52,55] are positively regulated by HNF4α, and the transactivation of Hnf4α is inhibited by BAs [56]. The marked increases of Cyp7a1 and Ntcp expressions have been observed in the liver with decreased BAs of the patients with cerebrotendinous xanthomatosis which is the inherited deficiency of key-enzyme (sterol 27-hydroxylase) in BA synthesis, although the FXR–SHP pathway was normally activated [57]. The significantly increased Hnf4a expression in the DKO liver was induced concomitantly with the significant decreases of TBA in the liver and the body pool in the present and previous studies [28]. Furthermore, the significantly increased taurine concentration in the DKO liver is likely to be a factor to increase Cyp7a1 expression, because taurine itself has the effect to increase Cyp7a1 mRNA expression as well as its activity in rodents [58,59,60]. In addition, taurine has been reported to act as an agonist of the nucleus receptor, liver X receptor α (LXRα; NR1H3) [61], which positively regulates Cyp7a1 expression [62,63]. In the DKO liver, we showed that the expressions of target genes for Lxrα, including sterol regulatory element-binding protein 1 and ATP-binding cassette transporter A1, and the biliary proportion of cholesterol were significantly increased, suggesting the activation of Lxrα [28]. Thus, the change to human BA composition also causes a concomitant decrease in BA content and an increase in taurine content in the DKO liver. Therefore, the absence of Cyp7a1 and Ntcp expression suppression despite Fxr activity may be related to competitive regulation by activation of Hnf4α and Lxrα in the DKO liver.
5. Conclusions
In the present study, using a model with a human-like BA composition in the liver, we confirmed, for the first time in mice, that a high content of BAs with FXR agonist activity upregulates taurine biosynthesis in addition to BA conjugation with taurine. Because the hydrophilicity of taurine-conjugated BAs is higher than that of glycine-conjugated BAs, and because taurine itself has various beneficial actions, the promotion of taurine conjugation and biosynthesis through FXR activation is considered meaningful for human health.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/metabo13070824/s1, Figure S1: The ratio of taurine-conjugated bile acids and the mRNA expressions of Fxr target genes in BA amidation and taurine synthesis in the liver of Cyp2a12 KO and Cyp2c70 KO mice.; Table S1: Evaluation stability and ranking of 14 candidate reference genes evaluated by geNorm and Bestkeeper algorisms.
Author Contributions
Conceptualization, T.M. and A.H.; methodology, T.M., H.U. and A.H.; validation, T.I.; formal analysis, T.M.; data curation, T.M.; writing—original draft preparation, T.M.; writing—review and editing, A.H.; visualization, T.M.; supervision, T.I.; project administration, T.M. and A.H.; funding acquisition, T.M., A.H. and T.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by a KAKENHI grant (15K15300, 17H04167, 18K08017, 18K07920, 21K11603, and 22H02853) from the Japanese Society for the Promotion of Science.
Institutional Review Board Statement
This experimental protocol was approved by the Animal Experiment Committees of The Jackson Laboratory Japan and Tokyo Medical University (Permission #H30-0069, H31- 0064, R2-0013, R3-0010, and R4-0048).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article.
Acknowledgments
The authors thank Raku Kato, Hideto Takahashi, Yutaro Wada, Shiro Uehara, Takashi Arakawa, and Takahiro Machiura (The Jackson Laboratory Japan, Inc.) for the breeding and care of the mice.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| APDS | 3-Aminopyridyl-N-hydroxysuccinimidyl carbamate |
| BA | bile acid |
| BAAT | bile acid-coenzyme A:amino acid N-acetyltransferase |
| BACS | ATP-dependent microsomal bile acid coenzyme A synthetase |
| BSEP | bile salt export pump |
| CA | cholic acid |
| CDCA | chenodeoxycholic acid |
| CDO | cysteine dioxygenase |
| CoA | coenzyme A |
| CSD | cysteine sulfinate decarboxylase |
| CYP | cytochrome P450 |
| DCA | deoxycholic acid |
| DKO | double knockout |
| FXR | farnesoid X receptor |
| FMO1/3 | flavin containing monooxygenase 1/3 |
| GCA | glycocholic acid |
| GCDCA | glycochenodeoxycholic acid |
| GDCA | glycodeoxycholic acid |
| GLCA | glycolithocholic acid |
| GUDCA | glycoursodeoxycholic acid |
| GW4064 | 3-[2-[2-chloro4-[[3-3(2,6-dichlorophenyl-5-(1-methylethyl-4-isoxazolyl]methoxy]phenyl]ethenyl]benzoic acid |
| HNF4α | hepatocyte nuclear factor 4α |
| HPLC-ESI-MS/MS | high-performance liquid chromatography–electrospray ionization tandem mass spectrometry |
| IS | internal standard |
| LCA | lithocholic acid |
| LXRα | liver X receptor α |
| MCA | muricholic acid |
| NTCP | sodium/taurocholate co-transporter peptide |
| PBS | phosphate-buffered saline |
| SE | standard error |
| SHP | small heterodimer partner |
| TBA | total bile acid |
| TCA | taurocholic acid |
| TCDCA | taurochenodeoxycholic acid |
| TDCA | taurodeoxycholic acid |
| TLCA | taurolithocholic acid |
| TUDCA | Tauroursodeoxycholic acid |
| TαMCA | tauro-α-muricholic acid |
| TβMCA | tauro-β-muricholic acid |
| TωMCA | tauro-ω-muricholic acid |
| UDCA | ursodeoxycholic acid |
| WT | wild type |
References
- Hosokawa, Y.; Matsumoto, A.; Oka, J.; Itakura, H.; Yamaguchi, K. Isolation and characterization of a cDNA for rat liver cysteine dioxygenase. Biochem. Biophys. Res. Commun. 1990, 168, 473–478. [Google Scholar] [CrossRef]
- Do, K.Q.; Tappaz, M.L. Specificity of cysteine sulfinate decarboxylase (CSD) for sulfur-containing amino-acids. Neurochem. Int. 1996, 28, 363–371. [Google Scholar] [CrossRef]
- Kaisaki, P.J.; Jerkins, A.A.; Goodspeed, D.C.; Steele, R.D.; Kaisakia, P.J. Cloning and characterization of rat cysteine sulfinic acid decarboxylase. Biochim. Biophys. Acta 1995, 1262, 79–82. [Google Scholar] [CrossRef]
- Reymond, I.; Sergeant, A.; Tappaz, M. Molecular cloning and sequence analysis of the cDNA encoding rat liver cysteine sulfinate decarboxylase (CSD). Biochim. Biophys. Acta 1996, 1307, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Tappaz, M.; Bitoun, M.; Reymond, I.; Sergeant, A. Characterization of the cDNA coding for rat brain cysteine sulfinate decarboxylase: Brain and liver enzymes are identical proteins encoded by two distinct mRNAs. J. Neurochem. 1999, 73, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Matsuzaki, Y. Taurine and liver diseases: A focus on the heterogeneous protective properties of taurine. Amino Acids 2014, 46, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Nakamura-Shinya, Y.; Ebina, K.; Komine, S.; Ra, S.G.; Ishikura, K.; Ohmori, H.; Honda, A. N-acetyltaurine and acetylcarnitine production for the mitochondrial acetyl-CoA regulation in skeletal muscles during endurance exercises. Metabolites 2021, 11, 522. [Google Scholar] [CrossRef]
- Huxtable, R.J. Does taurine have a function? Introduction. Fed. Proc. 1980, 39, 2678–2679. [Google Scholar]
- Huxtable, R.J. Physiological actions of taurine. Physiol. Rev. 1992, 72, 101–163. [Google Scholar] [CrossRef]
- Jacobsen, J.G.; Smith, L.H. Biochemistry and physiology of taurine and taurine derivatives. Physiol. Rev. 1968, 48, 424–491. [Google Scholar] [CrossRef]
- Miyazaki, T.; Ito, T.; Baseggio Conrado, A.; Murakami, S. Editorial for special issue on Regulation and effect of taurine on metabolism. Metabolites 2022, 12, 795. [Google Scholar] [CrossRef] [PubMed]
- Danielsson, H. Present status of research on catabolism and excretion of cholesterol. Adv. Lipid Res. 1963, 1, 335–385. [Google Scholar] [CrossRef] [PubMed]
- Sjovall, J. Dietary glycine and taurine on bile acid conjugation in man; bile acids and steroids 75. Proc. Soc. Exp. Biol. Med. 1959, 100, 676–678. [Google Scholar] [CrossRef] [PubMed]
- Falany, C.N.; Johnson, M.R.; Barnes, S.; Diasio, R.B. Glycine and taurine conjugation of bile acids by a single enzyme. Molecular cloning and expression of human liver bile acid CoA:Amino acid n-acyltransferase. J. Biol. Chem. 1994, 269, 19375–19379. [Google Scholar] [CrossRef]
- Danielsson, H.; Sjovall, J. Bile acid metabolism. Annu. Rev. Biochem. 1975, 44, 233–253. [Google Scholar] [CrossRef]
- Katafuchi, T.; Makishima, M. Molecular basis of bile acid-FXR-FGF15/19 signaling axis. Int. J. Mol. Sci. 2022, 23, 6046. [Google Scholar] [CrossRef]
- Solaas, K.; Ulvestad, A.; Soreide, O.; Kase, B.F. Subcellular organization of bile acid amidation in human liver: A key issue in regulating the biosynthesis of bile salts. J. Lipid Res. 2000, 41, 1154–1162. [Google Scholar] [CrossRef]
- Ferdinandusse, S.; Houten, S.M. Peroxisomes and bile acid biosynthesis. Biochim. Biophys. Acta 2006, 1763, 1427–1440. [Google Scholar] [CrossRef]
- Bremer, J. Species differences in the conjugation of free bile acids with taurine and glycine. Biochem. J. 1956, 63, 507–513. [Google Scholar] [CrossRef]
- Bruusgaard, A.; Thaysen, E.H. Increased ratio of glycine-taurine conjugated bile acids in the early diagnosis of terminal ileopathy. Preliminary report. Acta Med. Scand. 1970, 188, 547–548. [Google Scholar] [CrossRef]
- Hardison, W.G. Hepatic taurine concentration and dietary taurine as regulators of bile acid conjugation with taurine. Gastroenterology 1978, 75, 71–75. [Google Scholar] [CrossRef]
- Modica, S.; Gadaleta, R.M.; Moschetta, A. Deciphering the nuclear bile acid receptor FXR paradigm. Nucl. Recept. Signal. 2010, 8, e005. [Google Scholar] [CrossRef]
- Parks, D.J.; Blanchard, S.G.; Bledsoe, R.K.; Chandra, G.; Consler, T.G.; Kliewer, S.A.; Stimmel, J.B.; Willson, T.M.; Zavacki, A.M.; Moore, D.D.; et al. Bile acids: Natural ligands for an orphan nuclear receptor. Science 1999, 284, 1365–1368. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.Y. Regulation of bile acid synthesis: Pathways, nuclear receptors, and mechanisms. J. Hepatol. 2004, 40, 539–551. [Google Scholar] [CrossRef]
- Miyazaki, T.; Honda, A.; Matsuzaki, Y. Regulation of taurine conjugation and biosynthesis by bile acids through farnesoid x receptor activation. Hepatol. Res. 2014, 44, E1–E2. [Google Scholar] [CrossRef] [PubMed]
- Pircher, P.C.; Kitto, J.L.; Petrowski, M.L.; Tangirala, R.K.; Bischoff, E.D.; Schulman, I.G.; Westin, S.K. Farnesoid x receptor regulates bile acid-amino acid conjugation. J. Biol. Chem. 2003, 278, 27703–27711. [Google Scholar] [CrossRef] [PubMed]
- Kerr, T.A.; Matsumoto, Y.; Matsumoto, H.; Xie, Y.; Hirschberger, L.L.; Stipanuk, M.H.; Anakk, S.; Moore, D.D.; Watanabe, M.; Kennedy, S.; et al. Cysteine sulfinic acid decarboxylase regulation: A role for farnesoid x receptor and small heterodimer partner in murine hepatic taurine metabolism. Hepatol. Res. 2014, 44, E218–E228. [Google Scholar] [CrossRef]
- Honda, A.; Miyazaki, T.; Iwamoto, J.; Hirayama, T.; Morishita, Y.; Monma, T.; Ueda, H.; Mizuno, S.; Sugiyama, F.; Takahashi, S.; et al. Regulation of bile acid metabolism in mouse models with hydrophobic bile acid composition. J. Lipid Res. 2020, 61, 54–69. [Google Scholar] [CrossRef]
- Makishima, M.; Okamoto, A.Y.; Repa, J.J.; Tu, H.; Learned, R.M.; Luk, A.; Hull, M.V.; Lustig, K.D.; Mangelsdorf, D.J.; Shan, B. Identification of a nuclear receptor for bile acids. Science 1999, 284, 1362–1365. [Google Scholar] [CrossRef]
- Carotti, A.; Marinozzi, M.; Custodi, C.; Cerra, B.; Pellicciari, R.; Gioiello, A.; Macchiarulo, A. Beyond bile acids: Targeting farnesoid x receptor (FXR) with natural and synthetic ligands. Curr. Top. Med. Chem. 2014, 14, 2129–2142. [Google Scholar] [CrossRef]
- Sayin, S.I.; Wahlstrom, A.; Felin, J.; Jantti, S.; Marschall, H.U.; Bamberg, K.; Angelin, B.; Hyotylainen, T.; Oresic, M.; Backhed, F. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist. Cell. Metab. 2013, 17, 225–235. [Google Scholar] [CrossRef]
- Ueda, H.; Honda, A.; Miyazaki, T.; Morishita, Y.; Hirayama, T.; Iwamoto, J.; Nakamoto, N.; Ikegami, T. Sex-, age-, and organ-dependent improvement of bile acid hydrophobicity by ursodeoxycholic acid treatment: A study using a mouse model with human-like bile acid composition. PLoS ONE 2022, 17, e0271308. [Google Scholar] [CrossRef]
- Iwamoto, J.; Honda, A.; Miyazaki, T.; Monma, T.; Ueda, H.; Morishita, Y.; Yara, S.I.; Hirayama, T.; Ikegami, T. Western diet changes gut microbiota and ameliorates liver injury in a mouse model with human-like bile acid composition. Hepatol. Commun. 2021, 5, 2052–2067. [Google Scholar] [CrossRef]
- Yamashita, M.; Honda, A.; Shimoyama, S.; Umemura, M.; Ohta, K.; Chida, T.; Noritake, H.; Kurono, N.; Ichimura-Shimizu, M.; Tsuneyama, K.; et al. Breach of tolerance versus burden of bile acids: Resolving the conundrum in the immunopathogenesis and natural history of primary biliary cholangitis. J. Autoimmun. 2023, 136, 103027. [Google Scholar] [CrossRef]
- Sato, Y.; Atarashi, K.; Plichta, D.R.; Arai, Y.; Sasajima, S.; Kearney, S.M.; Suda, W.; Takeshita, K.; Sasaki, T.; Okamoto, S.; et al. Novel bile acid biosynthetic pathways are enriched in the microbiome of centenarians. Nature 2021, 599, 458–464. [Google Scholar] [CrossRef]
- Murakami, M.; Iwamoto, J.; Honda, A.; Tsuji, T.; Tamamushi, M.; Ueda, H.; Monma, T.; Konishi, N.; Yara, S.; Hirayama, T.; et al. Detection of gut dysbiosis due to reduced clostridium subcluster XIVa using the fecal or serum bile acid profile. Inflamm. Bowel Dis. 2018, 24, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Shoda, J.; Mahara, R.; Osuga, T.; Tohma, M.; Ohnishi, S.; Miyazaki, H.; Tanaka, N.; Matsuzaki, Y. Similarity of unusual bile acids in human umbilical cord blood and amniotic fluid from newborns and in sera and urine from adult patients with cholestatic liver diseases. J. Lipid Res. 1988, 29, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Sasaki, S.I.; Toyoda, A.; Wei, F.Y.; Shirai, M.; Morishita, Y.; Ikegami, T.; Tomizawa, K.; Honda, A. Impaired bile acid metabolism with defectives of mitochondrial-tRNA taurine modification and bile acid taurine conjugation in the taurine depleted cats. Sci. Rep. 2020, 10, 4915. [Google Scholar] [CrossRef]
- Namikawa-Kanai, H.; Miyazaki, T.; Matsubara, T.; Shigefuku, S.; Ono, S.; Nakajima, E.; Morishita, Y.; Honda, A.; Furukawa, K.; Ikeda, N. Comparison of the amino acid profile between the nontumor and tumor regions in patients with lung cancer. Am. J. Cancer Res. 2020, 10, 2145–2159. [Google Scholar] [PubMed]
- Shimbo, K.; Oonuki, T.; Yahashi, A.; Hirayama, K.; Miyano, H. Precolumn derivatization reagents for high-speed analysis of amines and amino acids in biological fluid using liquid chromatography/electrospray ionization tandem mass spectrometry. Rapid Commun. Mass. Spectrom. 2009, 23, 1483–1492. [Google Scholar] [CrossRef]
- Bustin, S.A.; Beaulieu, J.F.; Huggett, J.; Jaggi, R.; Kibenge, F.S.; Olsvik, P.A.; Penning, L.C.; Toegel, S. MIQE precis: Practical implementation of minimum standard guidelines for fluorescence-based quantitative real-time pcr experiments. BMC Mol. Biol. 2010, 11, 74. [Google Scholar] [CrossRef]
- Veeravalli, S.; Phillips, I.R.; Freire, R.T.; Varshavi, D.; Everett, J.R.; Shephard, E.A. Flavin-containing monooxygenase 1 catalyzes the production of taurine from hypotaurine. Drug. Metab. Dispos. 2020, 48, 378–385. [Google Scholar] [CrossRef]
- Bennett, B.J.; de Aguiar Vallim, T.Q.; Wang, Z.; Shih, D.M.; Meng, Y.; Gregory, J.; Allayee, H.; Lee, R.; Graham, M.; Crooke, R.; et al. Trimethylamine-N-oxide, a metabolite associated with atherosclerosis, exhibits complex genetic and dietary regulation. Cell. Metab. 2013, 17, 49–60. [Google Scholar] [CrossRef]
- Uchida, K.; Nomura, Y.; Kadowaki, M.; Takase, H.; Takano, K.; Takeuchi, N. Age-related changes in cholesterol and bile acid metabolism in rats. J. Lipid Res. 1978, 19, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Morelli, A.; Vignozzi, L.; Maggi, M.; Adorini, L. Farnesoid x receptor activation improves erectile dysfunction in models of metabolic syndrome and diabetes. Biochim. Biophys. Acta 2011, 1812, 859–866. [Google Scholar] [CrossRef]
- Yancey, P.H.; Blake, W.R.; Conley, J. Unusual organic osmolytes in deep-sea animals: Adaptations to hydrostatic pressure and other perturbants. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 133, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Sturman, J.; Hayes, K. The biology of taurine in nutrition and development. In Advances in Nutritional Research; H. H. Draper. 9; Springer: Boston, MA, USA, 1980; Chapter 9; pp. 231–299. [Google Scholar]
- Styles, N.A.; Shonsey, E.M.; Falany, J.L.; Guidry, A.L.; Barnes, S.; Falany, C.N. Carboxy-terminal mutations of bile acid CoA:N-acyltransferase alter activity and substrate specificity. J. Lipid Res. 2016, 57, 1133–1143. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, M.J.; Carey, M.C. The hydrophobic-hydrophilic balance of bile salts. Inverse correlation between reverse-phase high performance liquid chromatographic mobilities and micellar cholesterol-solubilizing capacities. J. Lipid Res. 1982, 23, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Gollapalli, K.; Mangiola, S.; Schranner, D.; Yusuf, M.A.; Chamoli, M.; Shi, S.L.; Lopes Bastos, B.; Nair, T.; Riermeier, A.; et al. Taurine deficiency as a driver of aging. Science 2023, 380, eabn9257. [Google Scholar] [CrossRef]
- Li, C.; Yang, J.; Wang, Y.; Qi, Y.; Yang, W.; Li, Y. Farnesoid x receptor agonists as therapeutic target for cardiometabolic diseases. Front. Pharmacol. 2020, 11, 1247. [Google Scholar] [CrossRef]
- Inoue, Y.; Yu, A.M.; Yim, S.H.; Ma, X.; Krausz, K.W.; Inoue, J.; Xiang, C.C.; Brownstein, M.J.; Eggertsen, G.; Bjorkhem, I.; et al. Regulation of bile acid biosynthesis by hepatocyte nuclear factor 4alpha. J. Lipid Res. 2006, 47, 215–227. [Google Scholar] [CrossRef]
- Crestani, M.; Sadeghpour, A.; Stroup, D.; Galli, G.; Chiang, J.Y. Transcriptional activation of the cholesterol 7alpha-hydroxylase gene (CYP7A) by nuclear hormone receptors. J. Lipid Res. 1998, 39, 2192–2200. [Google Scholar] [CrossRef]
- Stroup, D.; Chiang, J.Y. Hnf4 and COUP-TFII interact to modulate transcription of the cholesterol 7alpha-hydroxylase gene (CYP7A1). J. Lipid Res. 2000, 41, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hayhurst, G.P.; Lee, Y.H.; Lambert, G.; Ward, J.M.; Gonzalez, F.J. Hepatocyte nuclear factor 4alpha (nuclear receptor 2A1) is essential for maintenance of hepatic gene expression and lipid homeostasis. Mol. Cell. Biol. 2001, 21, 1393–1403. [Google Scholar] [CrossRef]
- Zhang, M.; Chiang, J.Y. Transcriptional regulation of the human sterol 12alpha-hydroxylase gene (CYP8B1): Roles of heaptocyte nuclear factor 4alpha in mediating bile acid repression. J. Biol. Chem. 2001, 276, 41690–41699. [Google Scholar] [CrossRef]
- Honda, A.; Salen, G.; Matsuzaki, Y.; Batta, A.K.; Xu, G.; Hirayama, T.; Tint, G.S.; Doy, M.; Shefer, S. Disrupted coordinate regulation of farnesoid X receptor target genes in a patient with cerebrotendinous xanthomatosis. J. Lipid Res. 2005, 46, 287–296. [Google Scholar] [CrossRef]
- Yokogoshi, H.; Mochizuki, H.; Nanami, K.; Hida, Y.; Miyachi, F.; Oda, H. Dietary taurine enhances cholesterol degradation and reduces serum and liver cholesterol concentrations in rats fed a high-cholesterol diet. J. Nutr. 1999, 129, 1705–1712. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, N.; Umeda, C.; Oda, H.; Yokogoshi, H. The effect of taurine on the cholesterol metabolism in rats fed diets supplemented with cholestyramine or high amounts of bile acid. J. Nutr. Sci. Vitaminol. (Tokyo) 2003, 49, 21–26. [Google Scholar] [CrossRef]
- Murakami, S.; Fujita, M.; Nakamura, M.; Sakono, M.; Nishizono, S.; Sato, M.; Imaizumi, K.; Mori, M.; Fukuda, N. Taurine ameliorates cholesterol metabolism by stimulating bile acid production in high-cholesterol-fed rats. Clin. Exp. Pharmacol. Physiol. 2016, 43, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Hoang, M.H.; Jia, Y.; Jun, H.J.; Lee, J.H.; Hwang, K.Y.; Choi, D.W.; Um, S.J.; Lee, B.Y.; You, S.G.; Lee, S.J. Taurine is a liver X receptor-alpha ligand and activates transcription of key genes in the reverse cholesterol transport without inducing hepatic lipogenesis. Mol. Nutr. Food Res. 2012, 56, 900–911. [Google Scholar] [CrossRef]
- Peet, D.J.; Turley, S.D.; Ma, W.; Janowski, B.A.; Lobaccaro, J.M.; Hammer, R.E.; Mangelsdorf, D.J. Cholesterol and bile acid metabolism are impaired in mice lacking the nuclear oxysterol receptor LXR alpha. Cell 1998, 93, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, B.; Watson, M.A.; Kim, H.; Miao, J.; Kemper, J.K.; Kliewer, S.A. Differential regulation of rat and human CYP7A1 by the nuclear oxysterol receptor liver X receptor-alpha. Mol. Endocrinol. 2003, 17, 386–394. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).