

Dam Body Condition Score Alters Offspring Circulating Cortisol and Energy Metabolites in Holstein Calves but Did Not Affect Neonatal Leptin Surge


Abstract
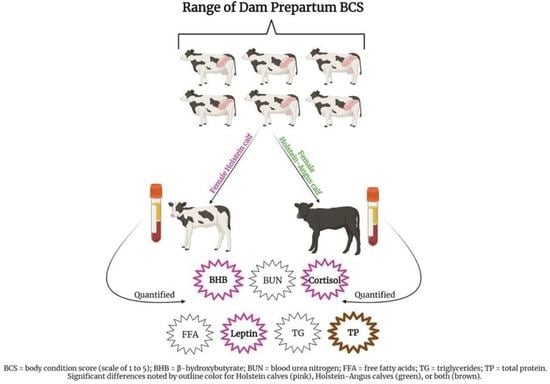
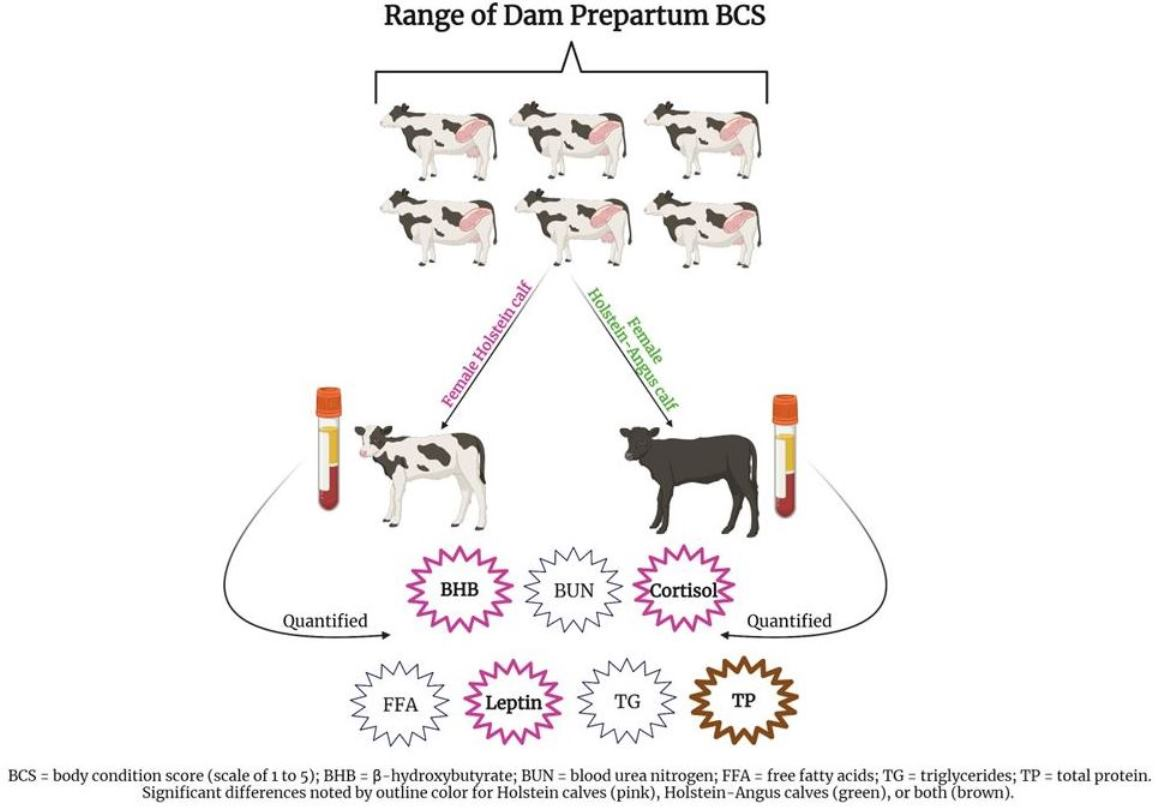
1. Introduction
2. Materials and Methods
2.1. Animal Use, Treatments, and Handling
2.2. Sample Collection and Analysis
2.3. Statistical Analysis
3. Results and Discussion
3.1. Leptin
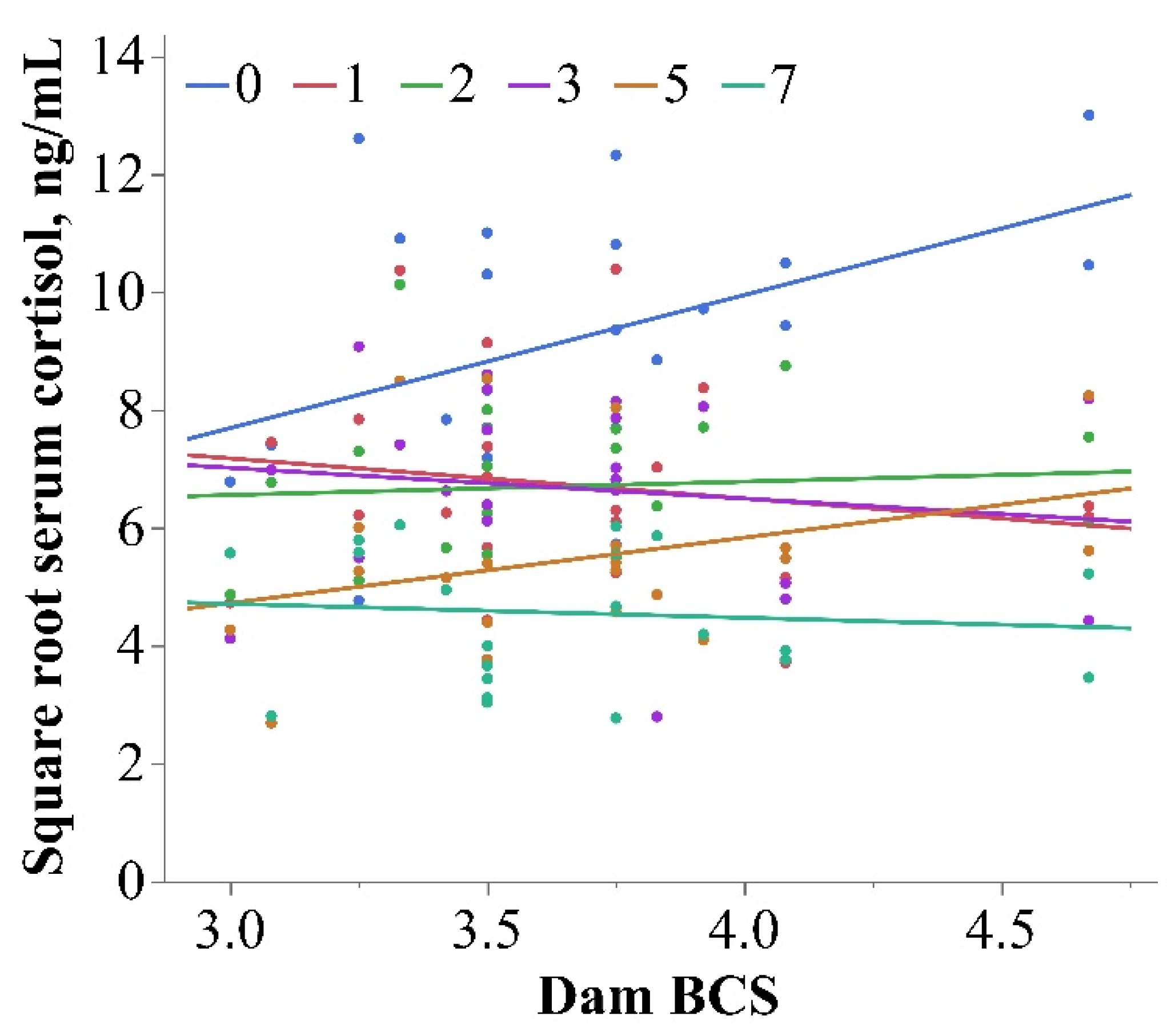
3.2. Cortisol
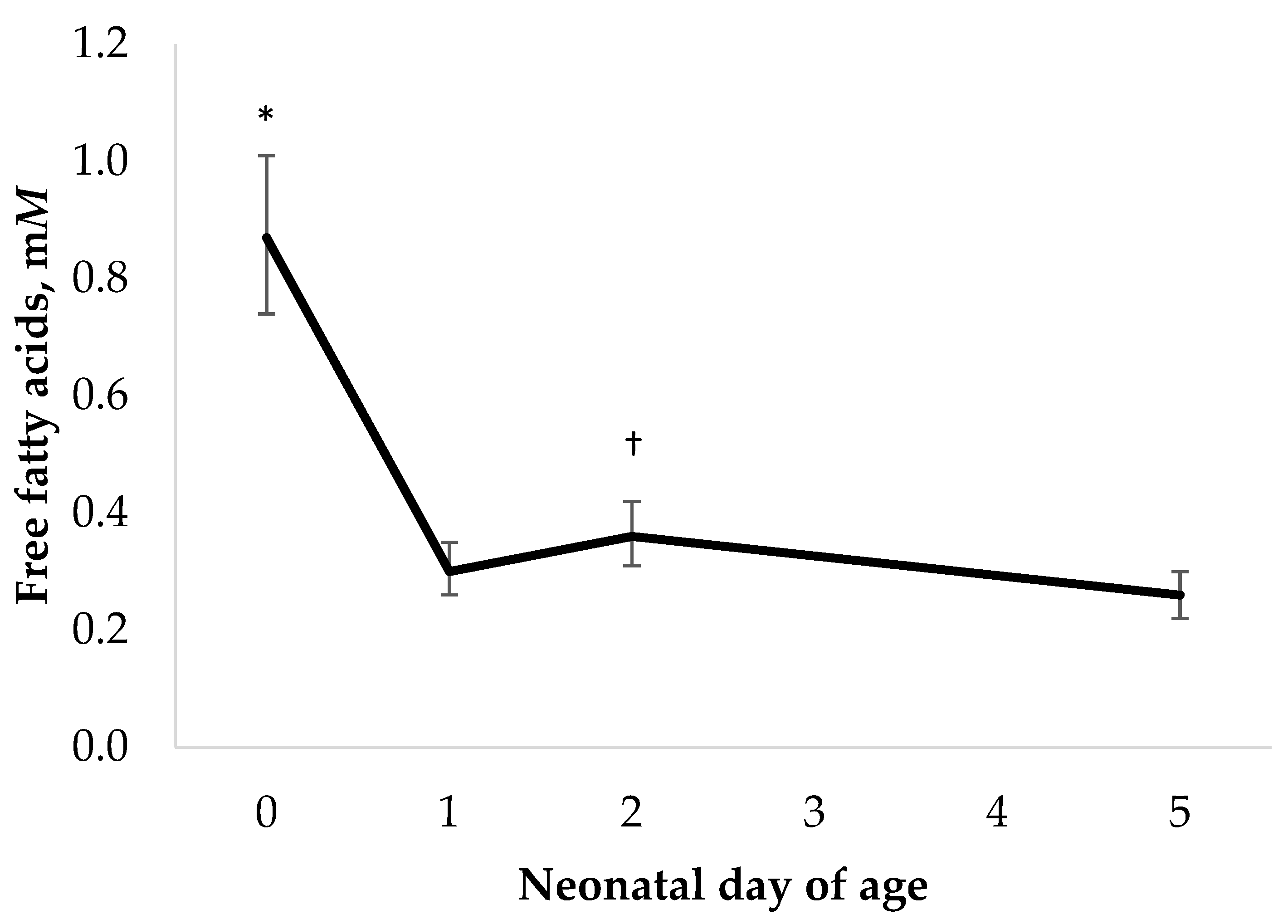
3.3. Other Serum Metabolites
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Funston, R.N.; Summers, A.F. Effect of Prenatal Programming on Heifer Development. Vet. Clin. Food Anim. 2013, 29, 517–536. [Google Scholar] [CrossRef]
- Du, M.; Ford, S.; Zhu, M. Optimizing Livestock Production Efficiency through Maternal Nutritional Management and Fetal Developmental Programming. Anim. Front. 2017, 7, 5–11. [Google Scholar] [CrossRef]
- Long, N.M.; George, L.A.; Uthlaut, A.B.; Smith, D.T.; Nijland, M.J.; Nathanielsz, P.W.; Ford, S.P. Maternal Obesity and Increased Nutrient Intake before and during Gestation in the Ewe Results in Altered Growth, Adiposity, and Glucose Tolerance in Adult Offspring. J. Anim. Sci. 2010, 88, 3546–3553. [Google Scholar] [CrossRef] [PubMed]
- Long, N.M.; Rule, D.C.; Tuersunjiang, N.; Nathanielsz, P.W.; Ford, S.P. Maternal Obesity in Sheep Increases Fatty Acid Synthesis, Upregulates Nutrient Transporters, and Increases Adiposity in Adult Male Offspring after a Feeding Challenge. PLoS ONE 2015, 10, e0122152. [Google Scholar] [CrossRef]
- Nivoit, P.; Morens, C.; Van Assche, F.A.; Jansen, E. Established Diet-Induced Obesity in Female Rats Leads to Offspring Hyperphagia, Adiposity and Insulin Resistance. Diabetologia 2009, 52, 1133–1142. [Google Scholar] [CrossRef]
- Kirk, S.L.; Samuelsson, A.M.; Argenton, M.; Dhonye, H.; Kalamatianos, T.; Poston, L.; Taylor, P.D.; Coen, C.W. Maternal Obesity Induced by Diet in Rats Permanently Influences Central Processes Regulating Food Intake in Offspring. PLoS ONE 2009, 4, e5870. [Google Scholar] [CrossRef] [PubMed]
- Roche, J.R.; Friggens, N.C.; Kay, J.K.; Fisher, M.W.; Stafford, K.J.; Berry, D.P. Invited Review: Body Condition Score and Its Association with Dairy Cow Productivity, Health, and Welfare. J. Dairy Sci. 2009, 92, 5769–5801. [Google Scholar] [CrossRef]
- Alharthi, A.S.; Coleman, D.N.; Alhidary, I.A.; Abdelrahman, M.M.; Trevisi, E.; Loor, J.J. Maternal Body Condition during Late-Pregnancy Is Associated with in Utero Development and Neonatal Growth of Holstein Calves. J. Anim. Sci. Biotechnol. 2021, 12, 44. [Google Scholar] [CrossRef] [PubMed]
- Banos, G.; Brotherstone, S.; Coffey, M.P. Prenatal Maternal Effects on Body Condition Score, Female Fertility, and Milk Yield of Dairy Cows. J. Dairy Sci. 2007, 90, 3490–3499. [Google Scholar] [CrossRef] [PubMed]
- Bouret, S.G. Neurodevelopmental Actions of Leptin. Brain Res. 2010, 1350, 2–9. [Google Scholar] [CrossRef]
- Granado, M.; Fuente-Martín, E.; García-Cáceres, C.; Argente, J.; Chowen, J.A. Leptin in Early Life: A Key Factor for the Development of the Adult Metabolic Profile. Obes. Facts 2012, 5, 138–150. [Google Scholar] [CrossRef] [PubMed]
- McFadin, E.L.; Morrison, C.D.; Buff, P.R.; Whitley, N.C.; Keisler, D.H. Leptin Concentrations in Periparturient Ewes and Their Subsequent Offspring. J. Anim. Sci. 2000, 80, 738–743. [Google Scholar] [CrossRef]
- Long, N.M.; Ford, S.P.; Nathanielsz, P.W. Maternal Obesity Eliminates the Neonatal Lamb Plasma Leptin Peak. J. Physiol. 2011, 589, 1455–1462. [Google Scholar] [CrossRef]
- Kesser, J.; Korst, M.; Koch, C.; Romberg, F.; Rehage, J.; Müller, U.; Schmicke, M.; Eder, K. Different Milk Feeding Intensities during the First 4 Weeks of Rearing Dairy Calves: Part 2: Effects on the Metabolic and Endocrine Status during Calfhood and around the First Lactation. J. Dairy Sci. 2017, 100, 3109–3125. [Google Scholar] [CrossRef] [PubMed]
- Long, N.M.; Schafer, D.W. Sex Effects on Plasma Leptin Concentrations in Newborn and Postnatal Beef Calves. Prof. Anim. Sci. 2013, 29, 601–605. [Google Scholar] [CrossRef]
- Pelleymounter, M.A.; Cullen, M.J.; Baker, M.B.; Hecht, R.; Winters, D.; Boone, T.; Collins, F. Effects of the Obese Gene Product on Body Weight Regulation in Ob/Ob Mice. Science 1995, 269, 540–543. [Google Scholar] [CrossRef] [PubMed]
- Proulx, K.; Richard, D.; Walker, C.D. Leptin Regulates Appetite-Related Neuropeptides in the Hypothalamus of Developing Rats without Affecting Food Intake. Endocrinology 2002, 143, 4683–4692. [Google Scholar] [CrossRef]
- Attig, L.; Solomon, G.; Ferezou, J.; Abdennebi-Najar, L.; Taouis, M.; Gertler, A.; Djiane, J. Early Postnatal Leptin Blockage Leads to a Long-Term Leptin Resistance and Susceptibility to Diet-Induced Obesity in Rats. Int. J. Obes. 2008, 32, 1153–1160. [Google Scholar] [CrossRef]
- Shasa, D.R.; Odhiambo, J.F.; Long, N.M.; Tuersunjiang, N.; Nathanielsz, P.W.; Ford, S.P. Multigenerational Impact of Maternal Overnutrition/Obesity in the Sheep on the Neonatal Leptin Surge in Granddaughters. Int. J. Obes. 2015, 39, 695–701. [Google Scholar] [CrossRef]
- Holdorf, H.; Kendall, S.; Ruh, K.; Caputo, M.; Combs, G.; Henisz, S.; Brown, W.; Bresolin, T.; Ferrerira, R.; Dorea, J.; et al. Increasing the Prepartum Dose of Rumen-Protected Choline: Effects on Milk Production and Metabolism in High Producing Holstein Dairy Cows. J. Dairy Sci. 2023, accepted. [Google Scholar]
- Wildman, E.E.; Jones, G.M.; Wagner, P.E.; Boman, R.L.; Troutt, H.F.; Lesch, T.N. A Dairy Cow Body Condition Scoring System and Its Relationship to Selected Production Characteristics. J. Dairy Sci. 1982, 65, 495–501. [Google Scholar] [CrossRef]
- Holdorf, H.; Brown, W.; Combs, G.; Henisz, S.; Kendall, S.; Caputo, M.; Ruh, K.; White, H. Increasing the Prepartum Dose of Rumen-Protected Choline: Effects of Maternal Choline Supplementation on Growth, Feed Efficiency, and Metabolism in Holstein and Holstein x Angus Calves. J. Dairy Sci. 2023, accepted. [Google Scholar]
- Caputo Oliveira, R.; Sailer, K.J.; Holdorf, H.T.; Seely, C.R.; Pralle, R.S.; Hall, M.B.; Bello, N.M.; White, H.M. Postpartum Supplementation of Fermented Ammoniated Condensed Whey Improved Feed Efficiency and Plasma Metabolite Profile. J. Dairy Sci. 2019, 102, 2283–2297. [Google Scholar] [CrossRef]
- Pralle, R.S.; Erb, S.J.; Holdorf, H.T.; White, H.M. Greater Liver PNPLA3 Protein Abundance in Vivo and in Vitro Supports Lower Triglyceride Accumulation in Dairy Cows. Sci. Rep. 2021, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Laporta, J.; Fabris, T.F.; Skibiel, A.L.; Powell, J.L.; Hayen, M.J.; Horvath, K.; Dahl, G.E. In Utero Exposure to Heat Stress during Late Gestation Has Prolonged Effects on the Activity Patterns and Growth of Dairy Calves. J. Dairy Sci. 2017, 100, 2976–2984. [Google Scholar] [CrossRef]
- Zenobi, M.; Gardinal, R.; Zuniga, J.; Dias, A.; Nelson, C.; Driver, J.; Barton, B.; Santos, J.; Staples, C. Effects of Supplementation with Ruminally Protected Choline on Performance of Multiparous Holstein Cows Did Not Depend upon Prepartum Caloric Intake. J. Dairy Sci. 2018, 101, 1088–1110. [Google Scholar] [CrossRef]
- Abuelo, A. Symposium Review: Late-Gestation Maternal Factors Affecting the Health and Development of Dairy Calves. J. Dairy Sci. 2020, 103, 3882–3893. [Google Scholar] [CrossRef]
- Schäff, C.T.; Rohrbeck, D.; Kanitz, E.; Sauerwein, H.; Bruckmaier, R.M. Effects of Colostrum versus Formula Feeding on Hepatic Glucocorticoid and A1- and Β2-Adrenergic Receptors in Neonatal Calves and Their Effect on Glucose and Lipid Metabolism. J. Dairy Sci. 2014, 97, 6344–6357. [Google Scholar] [CrossRef] [PubMed]
- Liermann, W.; Schäff, C.T.; Gruse, J.; Derno, M.; Weitzel, J.M.; Kanitz, E.; Otten, W.; Hoeflich, A.; Stefaniak, T.; Sauerwein, H.; et al. Effects of Colostrum Instead of Formula Feeding for the First 2 Days Postnatum on Whole-Body Energy Metabolism and Its Endocrine Control in Neonatal Calves. J. Dairy Sci. 2020, 103, 3577–3598. [Google Scholar] [CrossRef]
- Uken, K.L.; Vogel, L.; Gnott, M.; Görs, S.; Schäff, C.T.; Tuchscherer, A.; Hoeflich, A.; Weitzel, J.M.; Kanitz, E.; Tröscher, A.; et al. Effect of Maternal Supplementation with Essential Fatty Acids and Conjugated Linoleic Acid on Metabolic and Endocrine Development in Neonatal Calves. J. Dairy Sci. 2021, 104, 7295–7314. [Google Scholar] [CrossRef]
- Blum, J.W.; Zbinden, Y.; Hammon, H.M.; Chilliard, Y. Plasma Leptin Status in Young Calves: Effects of Pre-Term Birth, Age, Glucocorticoid Status, Suckling, and Feeding with an Automatic Feeder or by Bucket. Domest. Anim. Endocrinol. 2005, 28, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Lewis, L.K.; Ricks, R.E.; Long, N.M. Short Communication: Manipulation of Neonatal Leptin Profile via Exogenous Glucocorticoids in Beef Calves. Animal 2019, 13, 1982–1985. [Google Scholar] [CrossRef]
- Lemaster, C.T.; Taylor, R.K.; Ricks, R.E.; Long, N.M. The Effects of Late Gestation Maternal Nutrient Restriction with or without Protein Supplementation on Endocrine Regulation of Newborn and Postnatal Beef Calves. Theriogenology 2017, 87, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Osorio, J.S.; Trevisi, E.; Ballou, M.A.; Bertoni, G.; Drackley, J.K.; Loor, J.J. Effect of the Level of Maternal Energy Intake Prepartum on Immunometabolic Markers, Polymorphonuclear Leukocyte Function, and Neutrophil Gene Network Expression in Neonatal Holstein Heifer Calves. J. Dairy Sci. 2013, 96, 3573–3587. [Google Scholar] [CrossRef] [PubMed]
- Hammon, H.M.; Steinhoff-Wagner, J.; Flor, J.; Schönhusen, U.; Metges, C.C. LACTATION BIOLOGY SYMPOSIUM: Role of Colostrum and Colostrum Components on Glucose Metabolism in Neonatal Calves. J. Anim. Sci. 2013, 91, 685–695. [Google Scholar] [CrossRef]
- Hammon, H.M.; Blum, J.W. Metabolic and Endocrine Traits of Neonatal Calves Are Influenced by Feeding Colostrum for Different Durations or Only Milk Replacer. J. Nutr. 1998, 128, 624–632. [Google Scholar] [CrossRef]
- Van Soest, B.; Cullens, F.; VandeHaar, M.J.; Nielsen, M.W. Short Communication: Effects of Transition Milk and Milk Replacer Supplemented with Colostrum Replacer on Growth and Health of Dairy Calves. J. Dairy Sci. 2020, 103, 12104–12108. [Google Scholar] [CrossRef]
- Soliman, M.; Kimura, K.; Ahmed, M.; Yamaji, D.; Matsushita, Y.; Okamatsu-Ogura, Y.; Makondo, K.; Saito, M. Inverse Regulation of Leptin MRNA Expression by Short- and Long-Chain Fatty Acids in Cultured Bovine Adipocytes. Domest. Anim. Endocrinol. 2007, 33, 400–409. [Google Scholar] [CrossRef]
- Cammisotto, P.G.; Gélinas, Y.; Deshaies, Y.; Bukowiecki, L.J. Regulation of Leptin Secretion from White Adipocytes by Free Fatty Acids. Am. J. Physiol.—Endocrinol. Metab. 2003, 285, E521–E526. [Google Scholar] [CrossRef]
- Leal-Cerro, A.; Soto, A.; Martínez, M.A.; Dieguez, C.; Casanueva, F.F. Influence of Cortisol Status on Leptin Secretion. Pituitary 2001, 4, 111–116. [Google Scholar] [CrossRef]
- Houseknecht, K.L.; Portocarrero, C.P.; Ji, S.; Lemenager, R.; Spurlock, M.E. Growth Hormone Regulates Leptin Gene Expression in Bovine Adipose Tissue: Correlation with Adipose IGF-1 Expression. J. Endocrinol. 2000, 164, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Forhead, A.J.; Thomas, L.; Crabtree, J.; Hoggard, N.; Gardner, D.S.; Giussani, D.A.; Fowden, A.L. Plasma Leptin Concentration in Fetal Sheep during Late Gestation: Ontogeny and Effect of Glucocorticoids. Endocrinology 2002, 143, 1166–1173. [Google Scholar] [CrossRef] [PubMed]
- Long, N.; Smith, D.; Ford, S.; Nathanielsz, P. Elevated Glucocorticoids during Ovine Pregnancy Increase Appetite and Produce Glucose Dysregulation and Adiposity in Their Granddaughters in Response to Ad Libitum Feeding at One Year of Age. Am. J. Obs. Gynecol. 2014, 209, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Diesch, T.J.; Mellor, D.J.; Stafford, K.J.; Ward, R.N. Blood Metabolites and Body Temperature of Single Calves between 1 and 4 Days of Age in a Dairy Herd in New Zealand. N. Z. Vet. J. 2004, 52, 256–260. [Google Scholar] [CrossRef]
- Deelen, S.M.; Leslie, K.E.; Steele, M.A.; Eckert, E.; Brown, H.E.; DeVries, T.J. Validation of a Calf-Side β-Hydroxybutyrate Test and Its Utility for Estimation of Starter Intake in Dairy Calves around Weaning. J. Dairy Sci. 2016, 99, 7624–7633. [Google Scholar] [CrossRef]
- Eckert, E.; Brown, H.E.; Leslie, K.E.; DeVries, T.J.; Steele, M.A. Weaning Age Affects Growth, Feed Intake, Gastrointestinal Development, and Behavior in Holstein Calves Fed an Elevated Plane of Nutrition during the Preweaning Stage. J. Dairy Sci. 2015, 98, 6315–6326. [Google Scholar] [CrossRef] [PubMed]
- Nemati, M.; Amanlou, H.; Khorvash, M.; Moshiri, B.; Mirzaei, M.; Khan, M.A.; Ghaffari, M.H. Rumen Fermentation, Blood Metabolites, and Growth Performance of Calves during Transition from Liquid to Solid Feed: Effects of Dietary Level and Particle Size of Alfalfa Hay. J. Dairy Sci. 2015, 98, 7131–7141. [Google Scholar] [CrossRef] [PubMed]
- Omidi-Mirzaei, H.; Khorvash, M.; Ghorbani, G.R.; Moshiri, B.; Mirzaei, M.; Pezeshki, A.; Ghaffari, M.H. Effects of the Step-up/Step-down and Step-down Milk Feeding Procedures on the Performance, Structural Growth, and Blood Metabolites of Holstein Dairy Calves. J. Dairy Sci. 2015, 98, 7975–7981. [Google Scholar] [CrossRef]
- Hammon, H.M.; Schiessler, G.; Nussbaum, A.; Blum, J.W. Feed Intake Patterns, Growth Performance, and Metabolic and Endocrine Traits in Calves Fed Unlimited Amounts of Colostrum and Milk by Automate, Starting in the Neonatal Period. J. Dairy Sci. 2002, 85, 3352–3362. [Google Scholar] [CrossRef]
- Mccoy, G.C.; Reneau, J.K.; Hunter, A.G.; Williams, J.B. Effects of Diet and Time on Blood Serum Proteins in the Newborn Calf. J. Dairy Sci. 1970, 53, 358–362. [Google Scholar] [CrossRef]
- Rauprich, A.B.E.; Hammon, H.M.; Blum, J.W. Influence of Feeding Different Amounts of First Colostrum on Metabolic, Endocrine, and Health Status and on Growth Performance in Neonatal Calves. J. Anim. Sci. 2000, 78, 896–908. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dam BCS 2 | p-Value | |||||
|---|---|---|---|---|---|---|
| Item | BCS Estimate | 95% CI | BCS | Time | BCS × Time | Diet |
| Holstein, n = 22 | ||||||
| Leptin, ng/mL 3 | −0.003 | [−0.007, 0.001] | 0.12 | 0.09 | 0.79 | - |
| Cortisol, ng/mL 4 | −0.24 | [−1.87, 1.40] | 0.43 | 0.08 | 0.02 | - |
| BHB, mM 5 | 0.87 | [0.31, 1.43] | <0.01 | 0.19 | 0.28 | 0.06 |
| BUN, mg/dL 4 | −0.30 | [−0.83, 0.23] | 0.66 | 0.46 | 0.32 | - |
| FFA, mM 4 | 0.08 | [−0.28, 0.44] | 0.45 | 0.01 | 0.26 | - |
| TG, mg/dL 6 | 0.005 | [−0.004, 0.013] | 0.24 | 0.26 | 0.39 | - |
| Total Protein, g/dL 3 | 0.00 | [−0.004, 0.004] | 0.32 | 0.64 | 0.03 | - |
| Holstein × Angus, n = 11 | ||||||
| Leptin, ng/mL 3 | −0.006 | [−0.018, 0.007] | 0.68 | 0.51 | 0.58 | - |
| Cortisol, ng/mL 4 | −1.73 | [−4.96, 1.49] | 0.13 | 0.88 | 0.91 | - |
| BHB, mM | 0.02 | [−0.09, 0.12] | 0.31 | 0.53 | 0.58 | 0.06 |
| BUN, mg/dL | 0.65 | [−0.85, 2.16] | 0.98 | 0.39 | 0.41 | - |
| FFA, mM 4 | −0.01 | [−0.34, 0.32] | 0.43 | 0.27 | 0.49 | - |
| TG, mg/dL 3 | −0.001 | [−0.003, 0.001] | 0.15 | 0.86 | 0.91 | - |
| Total Protein, g/dL | 2.35 | [0.47, 4.23] | 0.01 | 0.45 | 0.20 | 0.02 |
| Day | Cortisol, mg/dL | Total Protein, g/dL | ||
|---|---|---|---|---|
| 0 | 84.7 | [72.5, 97.9] | 5.5 | [5.3, 5.6] |
| 1 | 45.0 | [36.3, 54.8] | 8.3 | [7.8, 8.8] |
| 2 | 44.9 | [36.1, 54.6] | 7.7 | [7.4, 8.2] |
| 3 | 44.3 | [35.6, 54.0] | - | |
| 5 | 29.8 | [22.8, 37.9] | 7.7 | [7.4, 8.2] |
| 7 | 20.6 | [14.8, 27.4] | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brown, W.E.; Holdorf, H.T.; Kendall, S.J.; White, H.M. Dam Body Condition Score Alters Offspring Circulating Cortisol and Energy Metabolites in Holstein Calves but Did Not Affect Neonatal Leptin Surge. Metabolites 2023, 13, 631. https://doi.org/10.3390/metabo13050631
Brown WE, Holdorf HT, Kendall SJ, White HM. Dam Body Condition Score Alters Offspring Circulating Cortisol and Energy Metabolites in Holstein Calves but Did Not Affect Neonatal Leptin Surge. Metabolites. 2023; 13(5):631. https://doi.org/10.3390/metabo13050631
Chicago/Turabian StyleBrown, William E., Henry T. Holdorf, Sophia J. Kendall, and Heather M. White. 2023. "Dam Body Condition Score Alters Offspring Circulating Cortisol and Energy Metabolites in Holstein Calves but Did Not Affect Neonatal Leptin Surge" Metabolites 13, no. 5: 631. https://doi.org/10.3390/metabo13050631
APA StyleBrown, W. E., Holdorf, H. T., Kendall, S. J., & White, H. M. (2023). Dam Body Condition Score Alters Offspring Circulating Cortisol and Energy Metabolites in Holstein Calves but Did Not Affect Neonatal Leptin Surge. Metabolites, 13(5), 631. https://doi.org/10.3390/metabo13050631