
Probing Serum Albumins and Cyclodextrins as Binders of the Mycotoxin Metabolites Alternariol-3-Glucoside, Alternariol-9-Monomethylether-3-Glucoside, and Zearalenone-14-Glucuronide

 ,
,  , and
, and 
Abstract
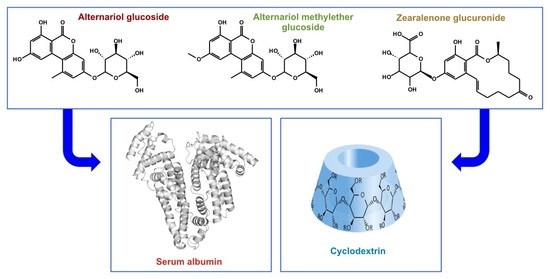
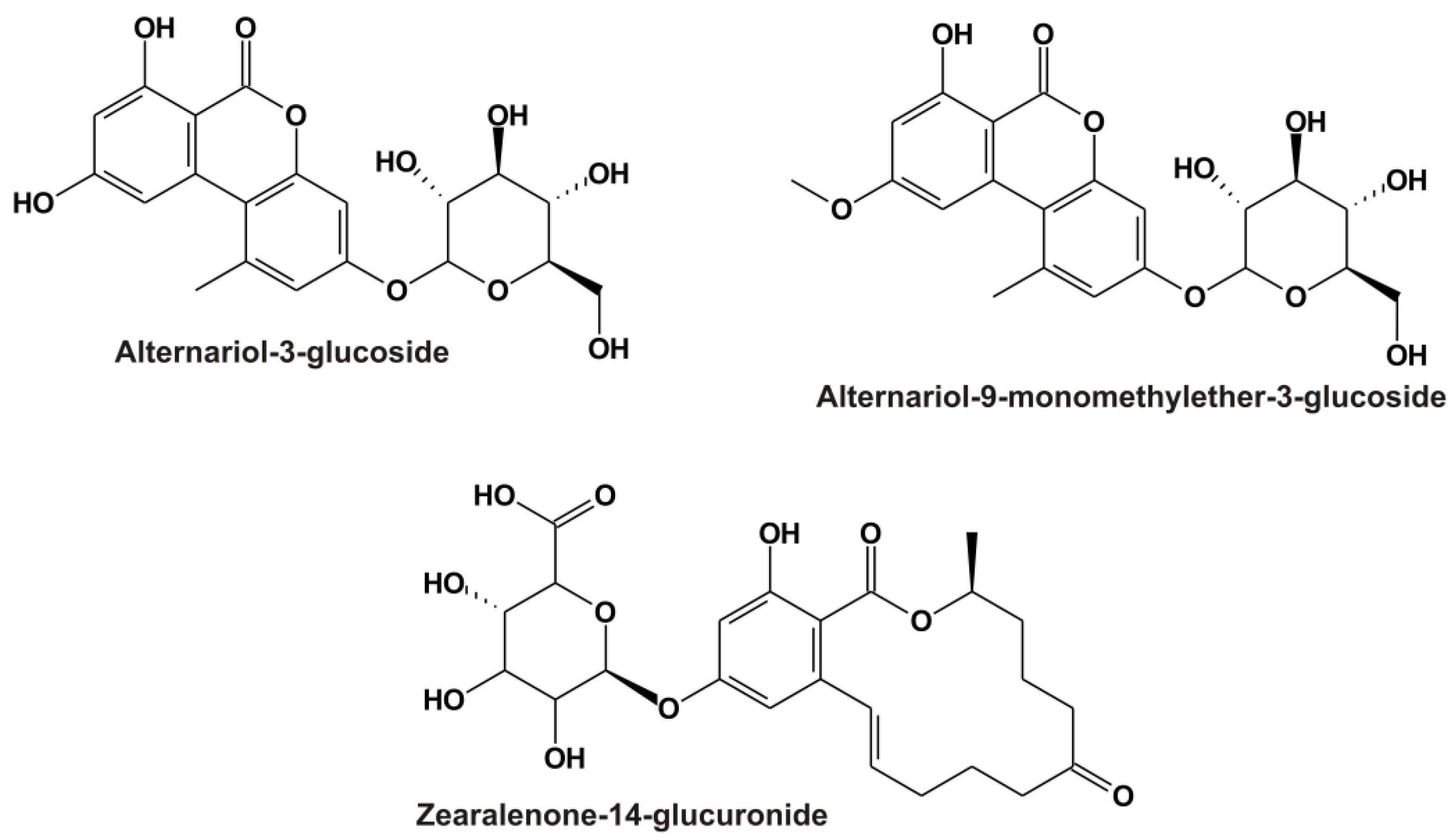
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Spectroscopic Studies
2.3. Ultracentrifugation Studies
2.4. Extraction of Mycotoxins with Cyclodextrin Bead Polymers
2.5. HPLC Analyses
2.6. Statistical Analyses
3. Results and Discussion
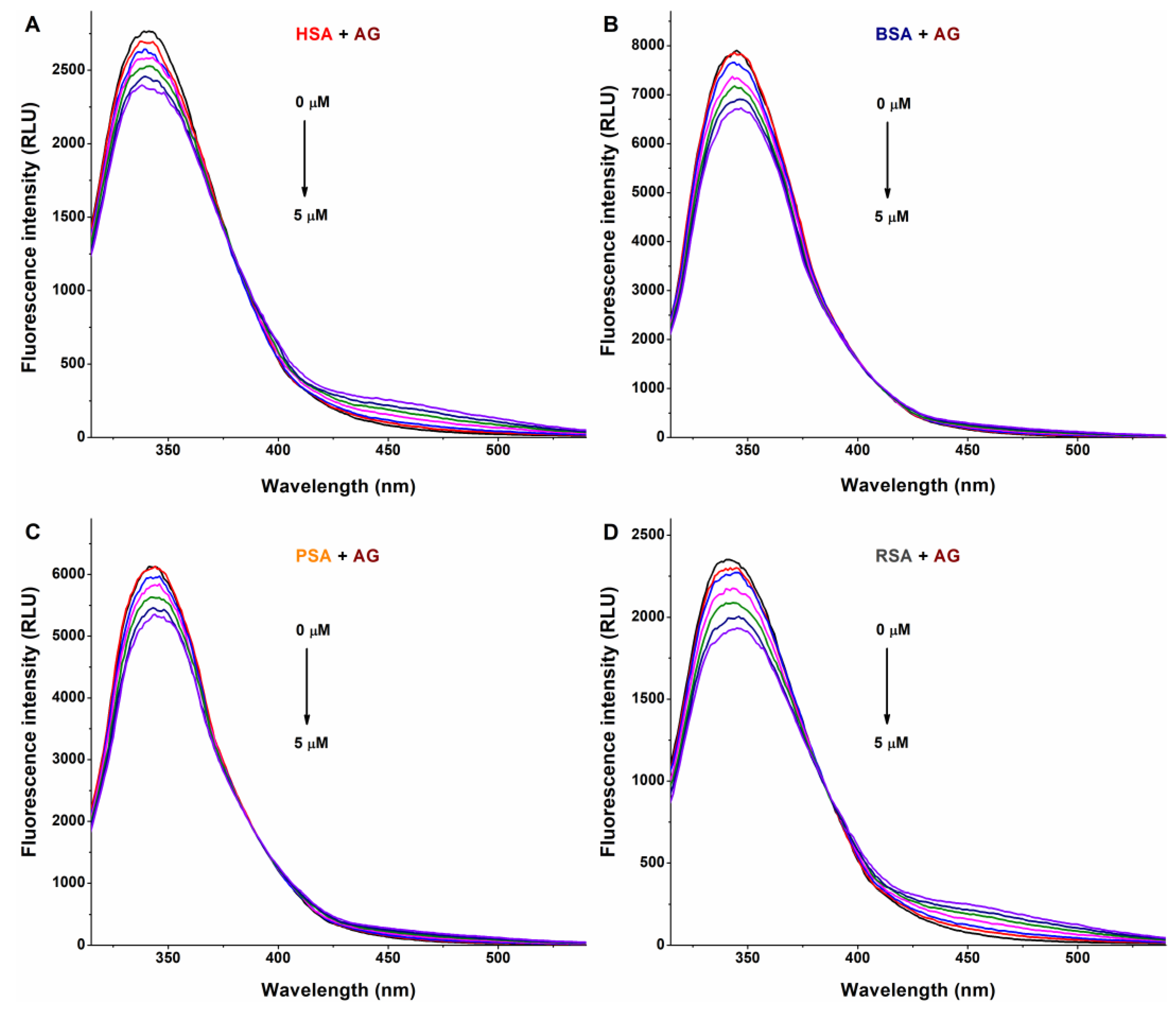
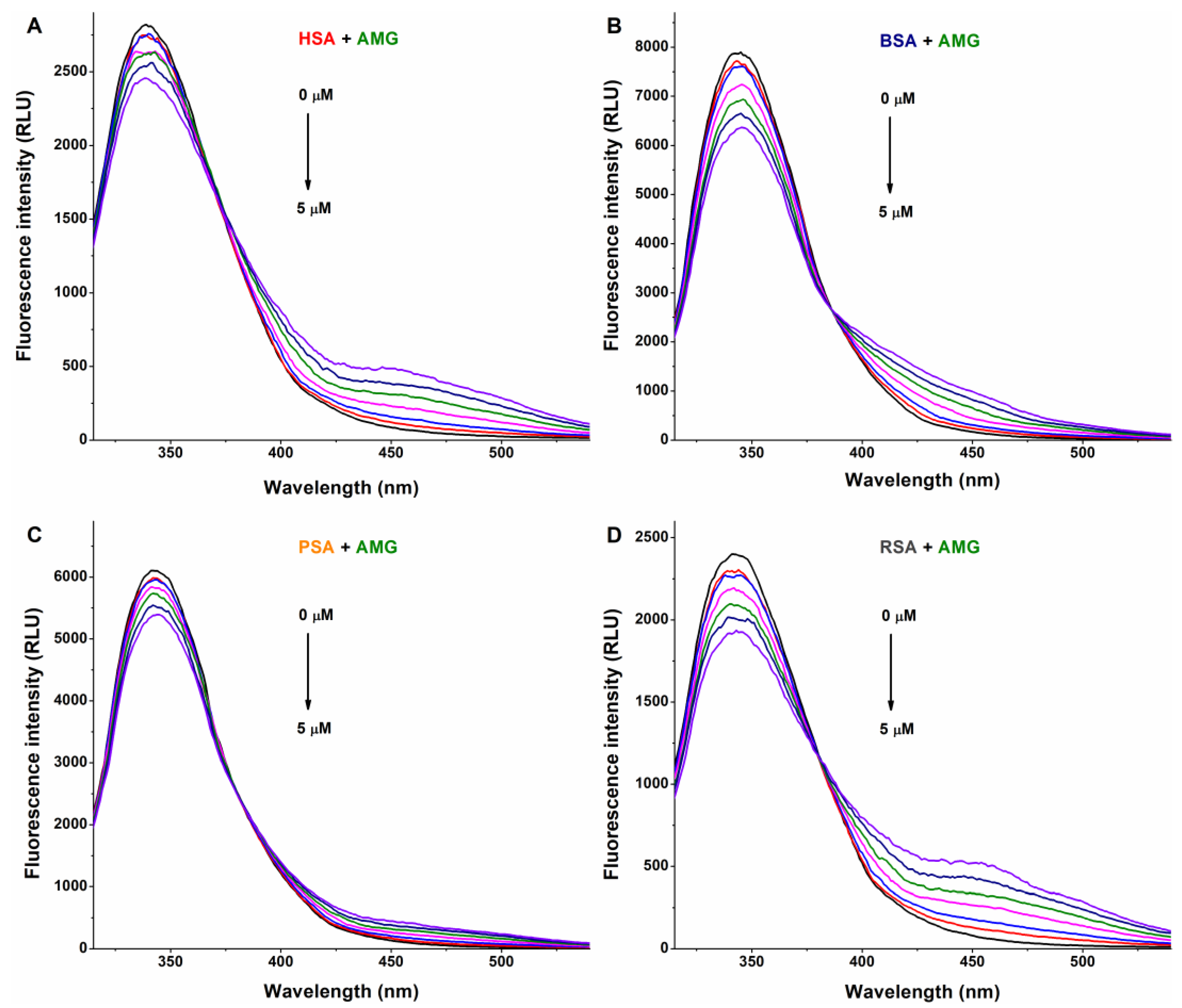
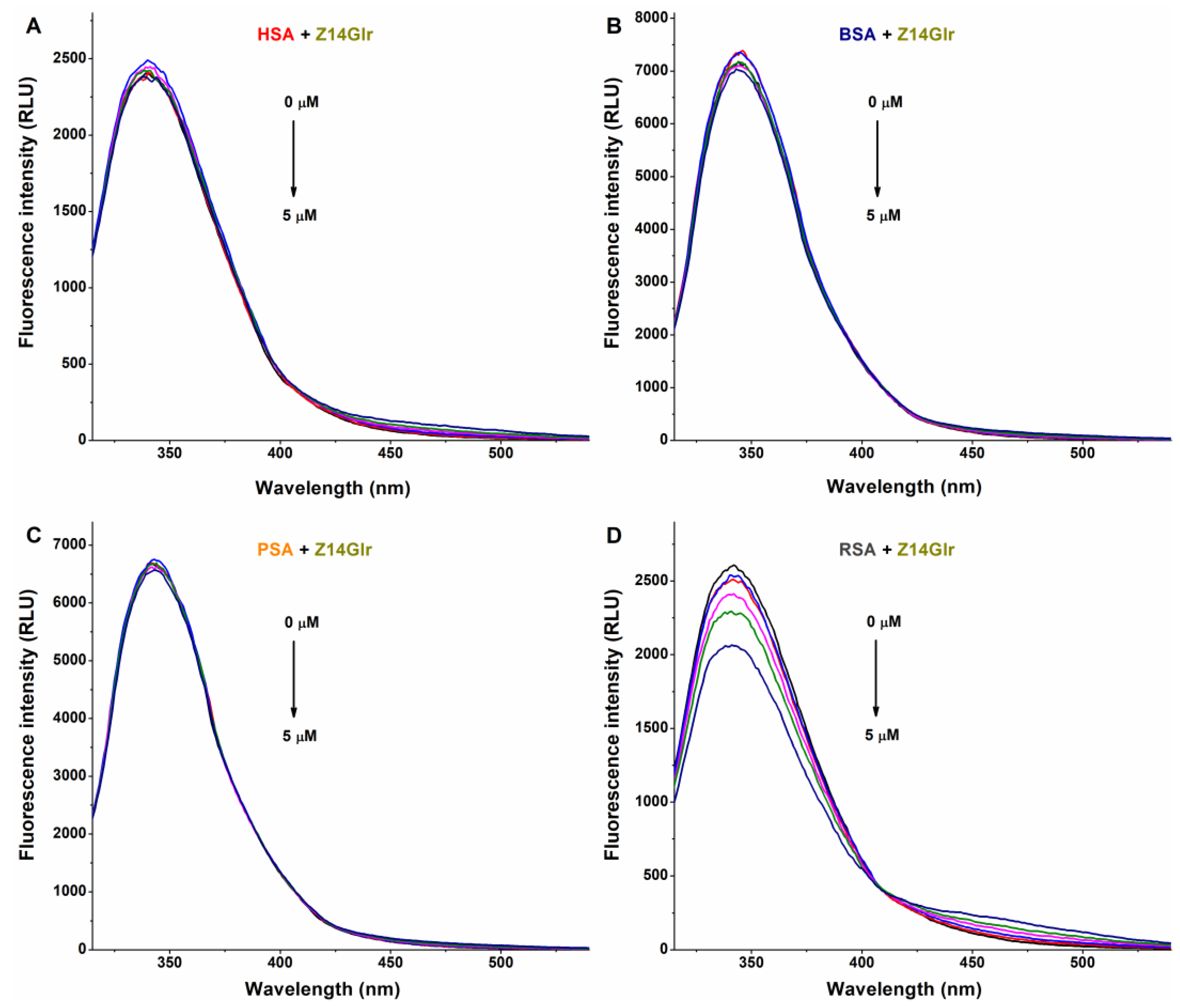
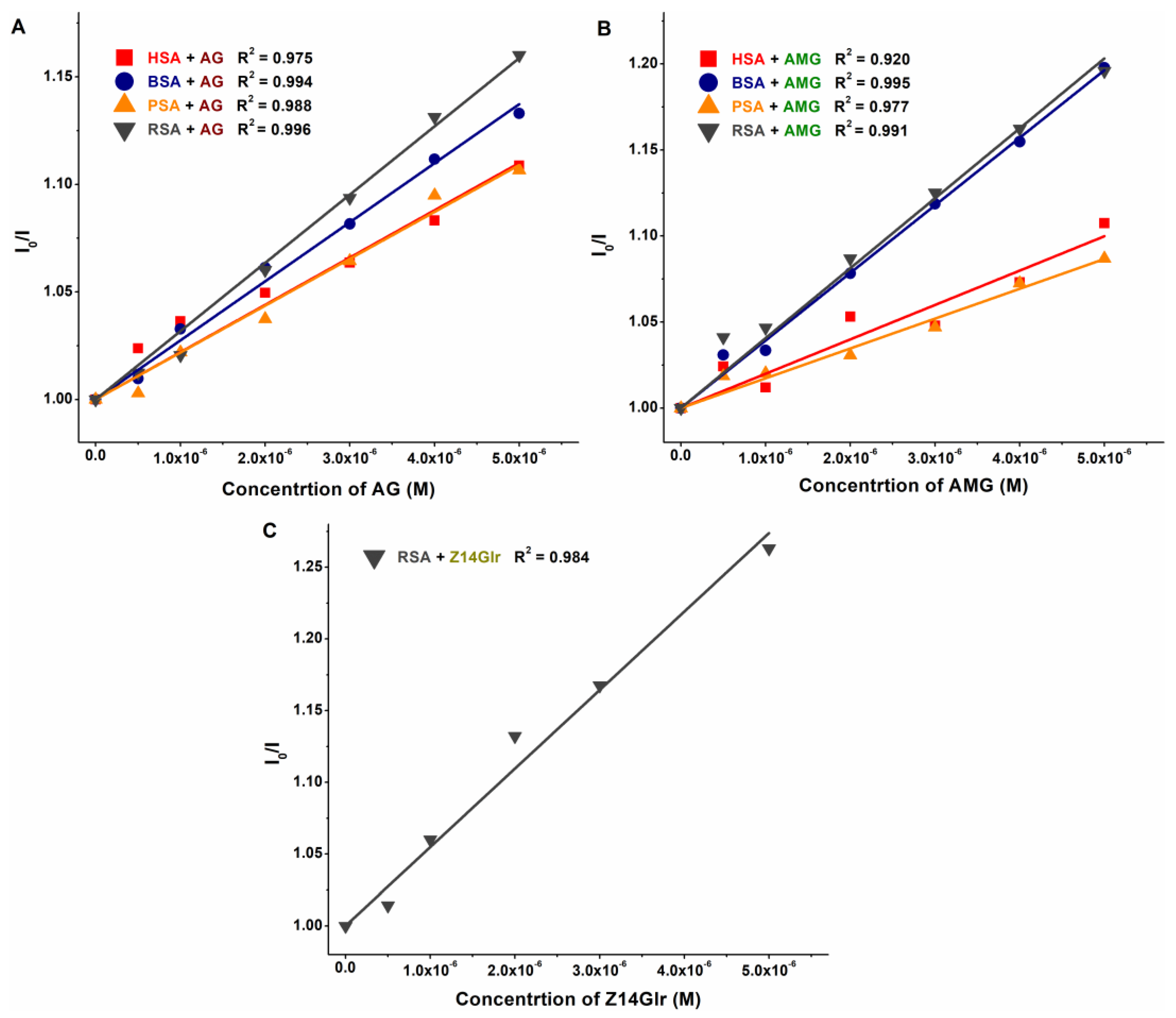
3.1. Interaction of AG, AMG, and Z14Glr with Serum Albumins Based on Fluorescence Quenching Studies
3.2. Interaction of AG, AMG, and Z14Glr with Serum Albumins Based on Ultracentrifugation Studies
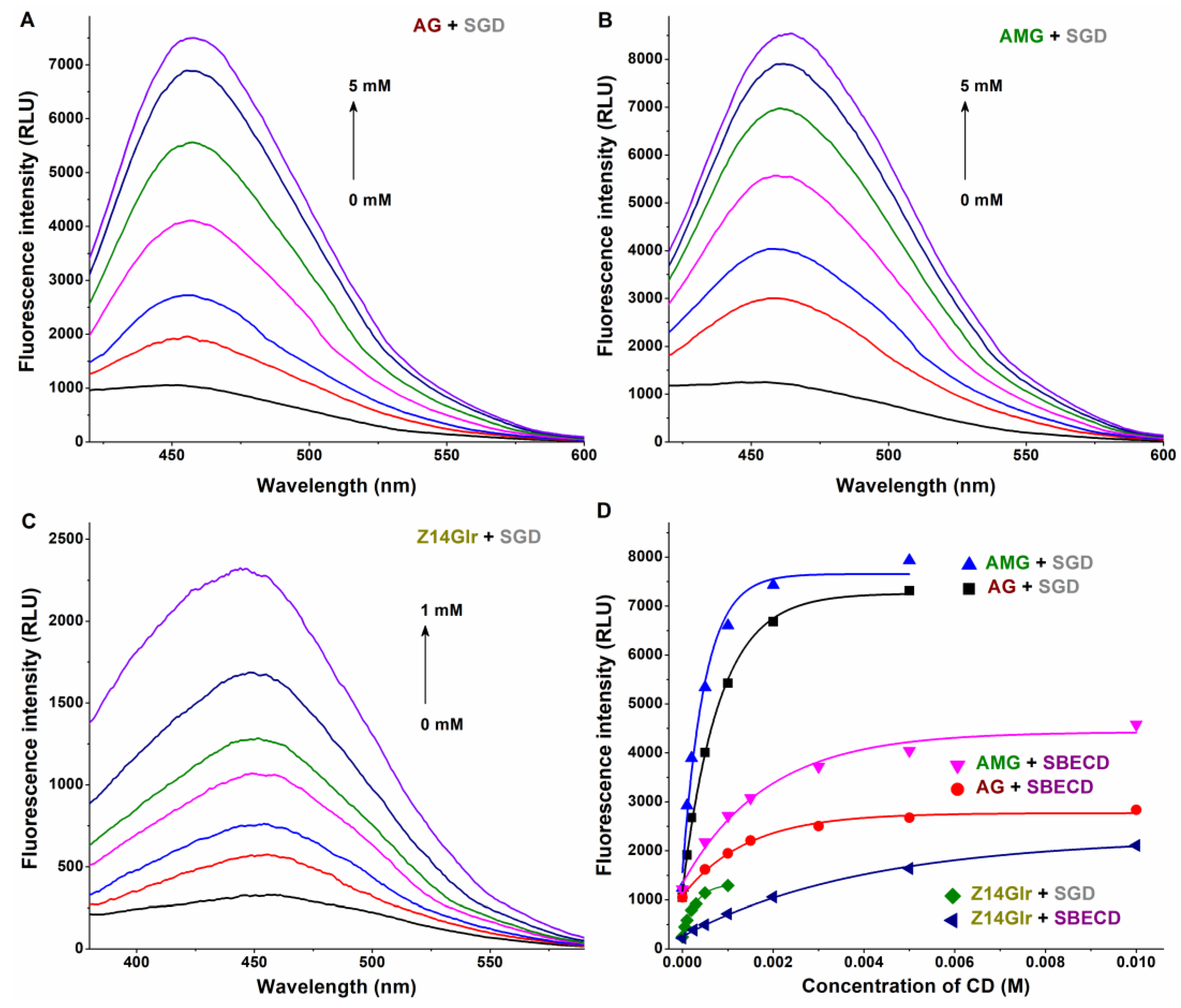
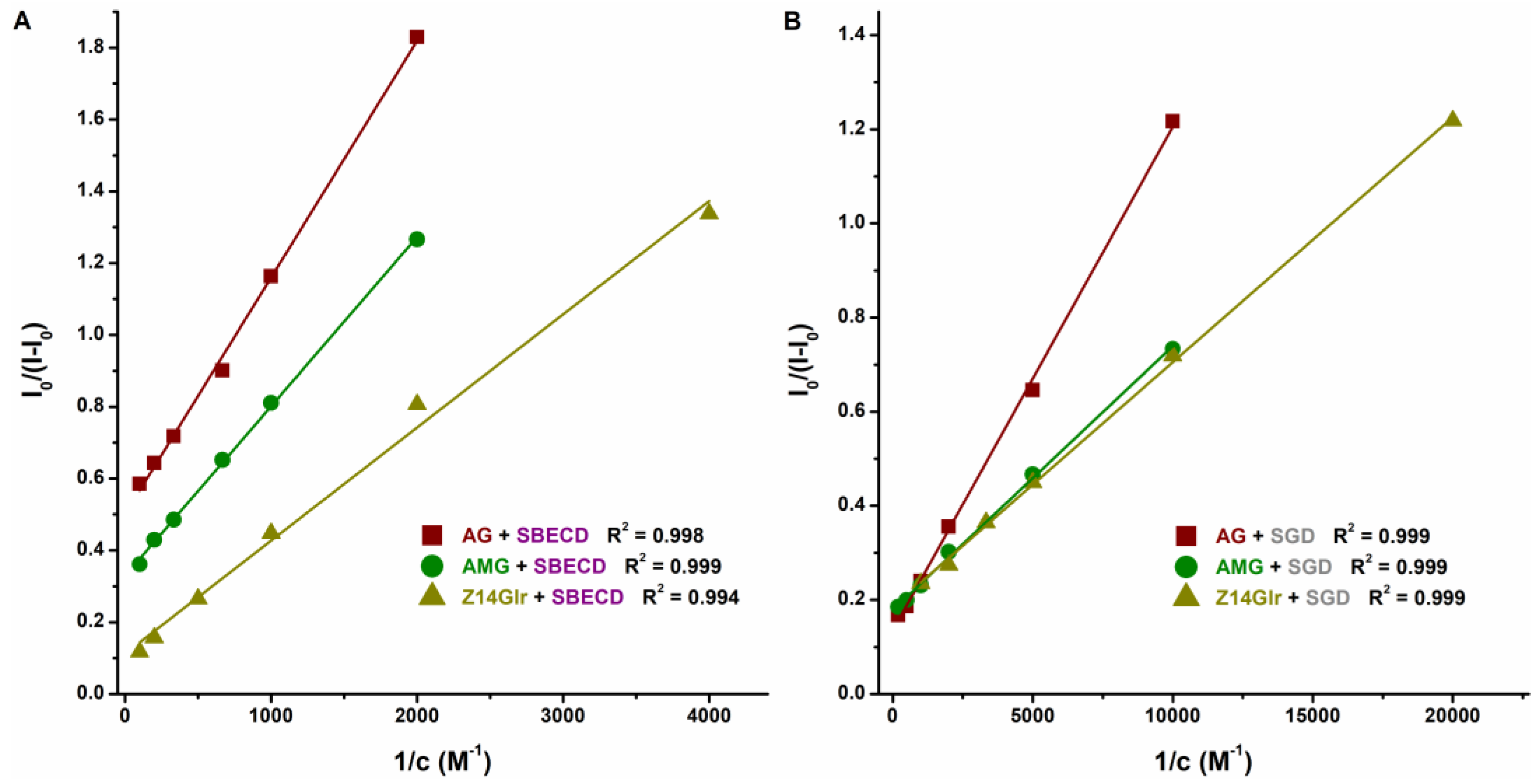
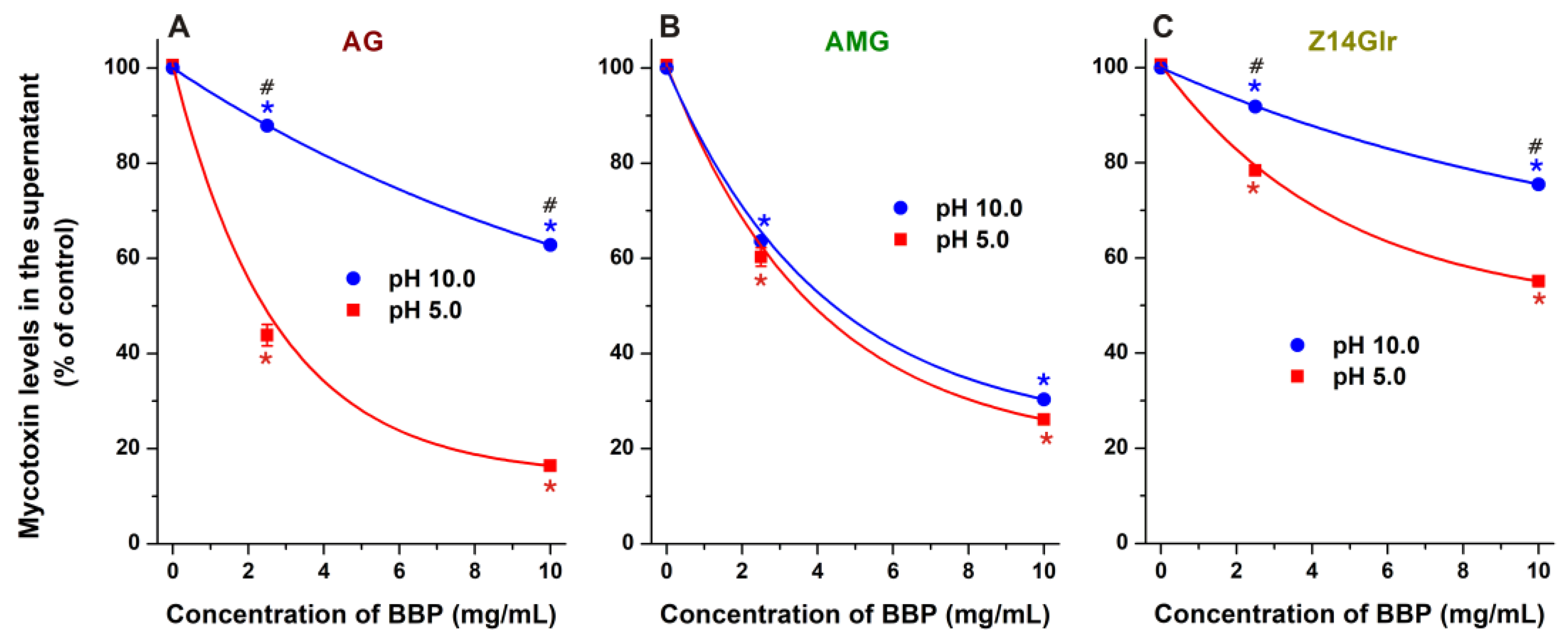
3.3. Interaction of AG, AMG, and Z14Glr with Sulfobutylether-β-Cyclodextrin and Sugammadex
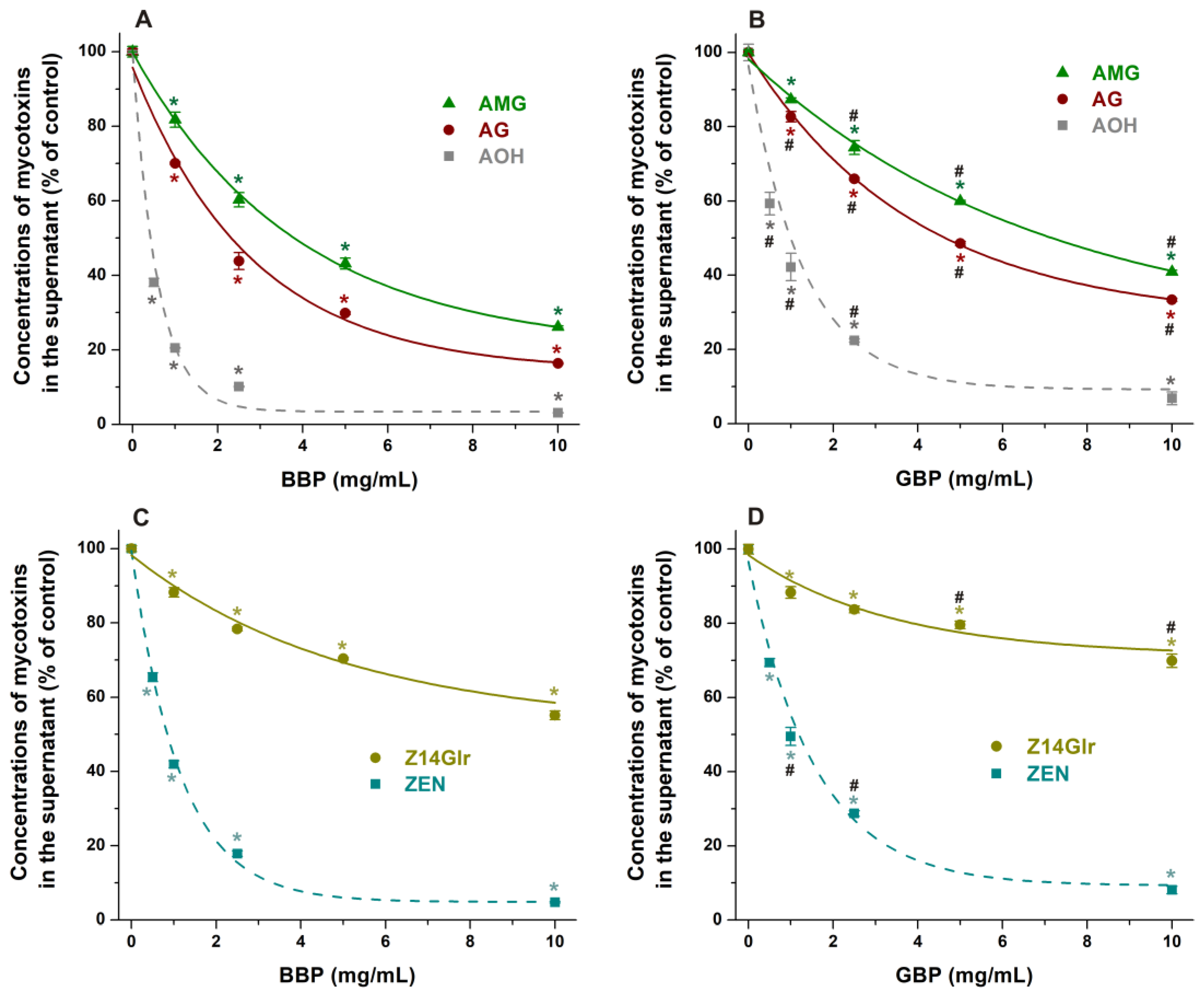
3.4. Extraction of Mycotoxins from Aqueous Solutions with Cyclodextrin Bead Polymers
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific Opinion on the Risks for Animal and Public Health Related to the Presence of Alternaria Toxins in Feed and Food. EFSA J. 2011, 9, 2407. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, e04851. [Google Scholar]
- Aichinger, G.; Del Favero, G.; Warth, B.; Marko, D. Alternaria toxins-Still emerging? Compr. Rev. Food. Sci. Food Saf. 2021, 20, 4390–4406. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Das, M.; Tripathi, A. Occurrence and toxicity of a fusarium mycotoxin, zearalenone. Crit. Rev. Food Sci. Nutr. 2020, 60, 2710–2729. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority; Arcella, D.; Eskola, M.; Gómez Ruiz, J.A. Dietary Exposure Assessment to Alternaria Toxins in the European Population. EFSA J. 2016, 14, 4654. [Google Scholar]
- Puntscher, H.; Kütt, M.-L.; Skrinjar, P.; Mikula, H.; Podlech, J.; Fröhlich, J.; Marko, D.; Warth, B. Tracking emerging mycotoxins in food: Development of an LC-MS/MS method for free and modified Alternaria toxins. Anal. Bioanal. Chem. 2018, 410, 4481–4494. [Google Scholar] [CrossRef]
- Qin, Q.; Fan, Y.; Jia, Q.; Duan, S.; Liu, F.; Jia, B.; Wang, G.; Guo, W.; Wang, C. The Potential of Alternaria Toxins Production by A. alternata in Processing Tomatoes. Toxins 2022, 14, 827. [Google Scholar] [CrossRef] [PubMed]
- Appel, B.N.; Gottmann, J.; Schäfer, J.; Bunzel, M. Absorption and metabolism of modified mycotoxins of alternariol, alternariol monomethyl ether, and zearalenone in Caco-2 cells. Cereal Chem. 2021, 98, 109–122. [Google Scholar] [CrossRef]
- Fanali, G.; di Masi, A.; Trezza, V.; Marino, M.; Fasano, M.; Ascenzi, P. Human Serum Albumin: From Bench to Bedside. Mol. Aspects. Med. 2012, 33, 209–290. [Google Scholar] [CrossRef]
- Fliszár-Nyúl, E.; Lemli, B.; Kunsági-Máté, S.; Dellafiora, L.; Dall’Asta, C.; Cruciani, G.; Pethő, G.; Poór, M. Interaction of Mycotoxin Alternariol with Serum Albumin. Int. J. Mol. Sci. 2019, 20, 2352. [Google Scholar] [CrossRef]
- Faisal, Z.; Lemli, B.; Szerencsés, D.; Kunsági-Máté, S.; Bálint, M.; Hetényi, C.; Kuzma, M.; Mayer, M.; Poór, M. Interactions of zearalenone and its reduced metabolites α-zearalenol and β-zearalenol with serum albumins: Species differences, binding sites, and thermodynamics. Mycotoxin Res. 2018, 34, 269–278. [Google Scholar] [CrossRef]
- Perry, J.L.; Christensen, T.; Goldsmith, M.R.; Toone, E.J.; Beratan, D.N.; Simon, J.D. Binding of Ochratoxin A to Human Serum Albumin Stabilized by a Protein-Ligand Ion Pair. J. Phys. Chem. B 2003, 107, 7884–7888. [Google Scholar] [CrossRef]
- Fliszár-Nyúl, E.; Szabó, Á.; Szente, L.; Poór, M. Extraction of mycotoxin alternariol from red wine and from tomato juice with beta-cyclodextrin bead polymer. J. Mol. Liq. 2020, 319, 114180. [Google Scholar] [CrossRef]
- Hong, C.-Y.; Chen, Y.-C. Selective enrichment of ochratoxin A using human serum albumin bound magnetic beads as the concentrating probes for capillary electrophoresis/electrospray ionization-mass spectrometric analysis. J. Chromatogr. A 2007, 1159, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Leal, T.; Abrunhosa, L.; Domingues, L.; Venâncio, A.; Oliveira, C. BSA-based sample clean-up columns for ochratoxin A determination in wine: Method development and validation. Food. Chem. 2019, 300, 125204. [Google Scholar] [CrossRef] [PubMed]
- Lemli, B.; Vilmányi, P.; Fliszár-Nyúl, E.; Zsidó, B.Z.; Hetényi, C.; Szente, L.; Poór, M. Testing Serum Albumins and Cyclodextrins as Potential Binders of the Mycotoxin Metabolites Alternariol-3-Sulfate, Alternariol-9-Monomethylether and Alternariol-9-Monomethylether-3-Sulfate. Int. J. Mol. Sci. 2022, 23, 14353. [Google Scholar] [CrossRef]
- Crini, G. Review: A history of cyclodextrins. Chem. Rev. 2014, 114, 10940–10975. [Google Scholar] [CrossRef]
- Fliszár-Nyúl, E.; Bock, I.; Csepregi, R.; Szente, L.; Szabó, I.; Csenki, Z.; Poór, M. Testing the protective effects of cyclodextrins vs. alternariol-induced acute toxicity in HeLa cells and in zebrafish embryos. Environ. Toxicol. Pharmacol. 2022, 95, 103965. [Google Scholar] [CrossRef]
- Fliszár-Nyúl, E.; Lemli, B.; Kunsági-Máté, S.; Szente, L.; Poór, M. Interactions of Mycotoxin Alternariol with Cyclodextrins and Its Removal from Aqueous Solution by Beta-Cyclodextrin Bead Polymer. Biomolecules 2019, 9, 428. [Google Scholar] [CrossRef] [PubMed]
- Poór, M.; Faisal, Z.; Zand, A.; Bencsik, T.; Lemli, B.; Kunsági-Máté, S.; Szente, L. Removal of Zearalenone and Zearalenols from Aqueous Solutions Using Insoluble Beta-Cyclodextrin Bead Polymer. Toxins 2018, 10, 216. [Google Scholar] [CrossRef]
- Faisal, Z.; Fliszár-Nyúl, E.; Dellafiora, L.; Galaverna, G.; Dall’Asta, C.; Lemli, B.; Kunsági-Máté, S.; Szente, L.; Poór, M. Interaction of zearalenone-14-sulfate with cyclodextrins and the removal of the modified mycotoxin from aqueous solution by beta-cyclodextrin bead polymer. J. Mol. Liq. 2020, 310, 113236. [Google Scholar] [CrossRef]
- Mohos, V.; Faisal, Z.; Fliszár-Nyúl, E.; Szente, L.; Poór, M. Testing the extraction of 12 mycotoxins from aqueous solutions by insoluble beta-cyclodextrin bead polymer. Environ. Sci. Pollut. Res. 2022, 29, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Faisal, Z.; Garai, E.; Csepregi, R.; Bakos, K.; Fliszár-Nyúl, E.; Szente, L.; Balázs, A.; Cserháti, M.; Kőszegi, T.; Urbányi, B.; et al. Protective effects of beta-cyclodextrins vs. zearalenone-induced toxicity in HeLa cells and Tg(vtg1:mCherry) zebrafish embryos. Chemosphere 2020, 240, 124948. [Google Scholar] [CrossRef]
- Tan, H.; Zhou, H.; Guo, T.; Zhou, Y.; Zhang, Q.; Zhang, Y.; Ma, L. Recent advances on formation, transformation, occurrence, and analytical strategy of modified mycotoxins in cereals and their products. Food Chem. 2023, 405, 134752. [Google Scholar] [CrossRef]
- Morin-Crini, N.; Crini, G. Environmental applications of water-insoluble β-cyclodextrin–epichlorohydrin polymers. Prog. Polym. Sci. 2013, 38, 344–368. [Google Scholar]
- Moulahcene, L.; Skiba, M.; Senhadji, O.; Milon, N.; Benamor, M.; Lahiani-Skiba, M. Inclusion and removal of pharmaceutical residues from aqueous solution using water-insoluble cyclodextrin polymers. Chem. Eng. Res. Des. 2015, 97, 145–158. [Google Scholar] [CrossRef]
- Morin-Crini, N.; Winterton, P.; Fourmentin, S.; Wilson, L.D.; Fenyvesi, É.; Crini, G. Water-insoluble β-cyclodextrin–epichlorohydrin polymers for removal of pollutants from aqueous solutions by sorption processes using batch studies: A review of inclusion mechanisms. Prog. Polym. Sci. 2018, 78, 1–23. [Google Scholar] [CrossRef]
- Wang, T.; Zeng, L.-H.; Li, D.-L. A Review on the Methods for Correcting the Fluorescence Inner-Filter Effect of Fluorescence Spectrum. Appl. Spectrosc. Rev. 2017, 52, 883–908. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd ed.; Springer: New York, NY, USA, 2006; ISBN 978-0-387-31278-1. [Google Scholar]
- Gans, P.; Sabatini, A.; Vacca, A. Investigation of Equilibria in Solution. Determination of Equilibrium Constants with the HYPERQUAD Suite of Programs. Talanta 1996, 43, 1739–1753. [Google Scholar] [CrossRef]
- Verrone, R.; Catucci, L.; Cosma, P.; Fini, P.; Agostiano, A.; Lippolis, V.; Pascale, M. Effect of b-cyclodextrin on spectroscopic properties of ochratoxin A in aqueous solution. J. Incl. Phenom. Macrocycl. Chem. 2007, 57, 475–479. [Google Scholar] [CrossRef]
- Fliszár-Nyúl, E.; Faisal, Z.; Skaper, R.; Lemli, B.; Bayartsetseg, B.; Hetényi, C.; Gömbös, P.; Szabó, A.; Poór, M. Interaction of the Emerging Mycotoxins Beauvericin, Cyclopiazonic Acid, and Sterigmatocystin with Human Serum Albumin. Biomolecules 2022, 12, 1106. [Google Scholar] [PubMed]
- van de Weert, M.; Stella, L. Fluorescence Quenching and Ligand Binding: A Critical Discussion of a Popular Methodology. J. Mol. Struct. 2011, 998, 144–150. [Google Scholar] [CrossRef]
- Dobretsov, G.E.; Syrejschikova, T.I.; Smolina, N.V. On Mechanisms of Fluorescence Quenching by Water. Biophysics 2014, 59, 231–237. [Google Scholar]
- Mikula, H.; Hametner, C.; Berthiller, F.; Warth, B.; Krska, R.; Adam, G.; Fröhlich, J. Fast Reproducible Chemical Synthesis of Zearalenone-14-β,D-Glucuronide. World Mycotoxin J. 2012, 5, 289–296. [Google Scholar] [CrossRef]
- Nasseri, S.A.; Betschart, L.; Opaleva, D.; Rahfeld, P.; Withers, S.G. A Mechanism-Based Approach to Screening Metagenomic Libraries for Discovery of Unconventional Glycosidases. Angew. Chem. Int. Ed. 2018, 57, 11359–11364. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HSA logKSV ± SEM | BSA logKSV ± SEM | PSA logKSV ± SEM | RSA logKSV ± SEM | |
|---|---|---|---|---|
| AG | 4.48 ± 0.08 | 4.44 ± 0.04 | 4.38 ± 0.03 | 4.49 ± 0.03 |
| AMG | 4.38 ± 0.05 | 4.57 ± 0.02 | 4.27 ± 0.03 | 4.62 ± 0.02 |
| Z14Glr | – | – | – | 4.65 ± 0.05 |
| HSA logK ± SEM | BSA logK ± SEM | PSA logK ± SEM | RSA logK ± SEM | |
|---|---|---|---|---|
| AG | 4.52 ± 0.10 | 4.46 ± 0.04 | 4.43 ± 0.03 | 4.53 ± 0.02 |
| AMG | 4.40 ± 0.05 | 4.59 ± 0.02 | 4.29 ± 0.02 | 4.65 ± 0.02 |
| Z14Glr | – | – | – | 4.71 ± 0.03 |
| HSA pH 7.4 logK ± SEM | HSA pH 5.0 logK ± SEM | HSA pH 8.5 logK ± SEM | |
|---|---|---|---|
| AG | 4.24 ± 0.02 | 4.48 ± 0.03 | 4.40 ± 0.06 |
| AMG | 4.11 ± 0.03 | 3.82 ± 0.02 | 5.05 ± 0.07 |
| Z14Glr | 4.22 ± 0.03 | 4.59 ± 0.14 | 4.23 ± 0.16 |
| SBECD logK ± SEM | SGD logK ± SEM | |
|---|---|---|
| AG | 2.92 ± 0.05 | 3.19 ± 0.08 |
| AMG | 2.79 ± 0.08 | 3.45 ± 0.04 |
| Z14Glr | 2.32 ± 0.12 | 3.46 ± 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poór, M.; Lemli, B.; Vilmányi, P.; Dombi, Á.; Nagymihály, Z.; Both, E.B.; Lambert, N.; Czömpöly, T.; Szente, L. Probing Serum Albumins and Cyclodextrins as Binders of the Mycotoxin Metabolites Alternariol-3-Glucoside, Alternariol-9-Monomethylether-3-Glucoside, and Zearalenone-14-Glucuronide. Metabolites 2023, 13, 446. https://doi.org/10.3390/metabo13030446
Poór M, Lemli B, Vilmányi P, Dombi Á, Nagymihály Z, Both EB, Lambert N, Czömpöly T, Szente L. Probing Serum Albumins and Cyclodextrins as Binders of the Mycotoxin Metabolites Alternariol-3-Glucoside, Alternariol-9-Monomethylether-3-Glucoside, and Zearalenone-14-Glucuronide. Metabolites. 2023; 13(3):446. https://doi.org/10.3390/metabo13030446
Chicago/Turabian StylePoór, Miklós, Beáta Lemli, Péter Vilmányi, Ágnes Dombi, Zoltán Nagymihály, Eszter Borbála Both, Nándor Lambert, Tamás Czömpöly, and Lajos Szente. 2023. "Probing Serum Albumins and Cyclodextrins as Binders of the Mycotoxin Metabolites Alternariol-3-Glucoside, Alternariol-9-Monomethylether-3-Glucoside, and Zearalenone-14-Glucuronide" Metabolites 13, no. 3: 446. https://doi.org/10.3390/metabo13030446
APA StylePoór, M., Lemli, B., Vilmányi, P., Dombi, Á., Nagymihály, Z., Both, E. B., Lambert, N., Czömpöly, T., & Szente, L. (2023). Probing Serum Albumins and Cyclodextrins as Binders of the Mycotoxin Metabolites Alternariol-3-Glucoside, Alternariol-9-Monomethylether-3-Glucoside, and Zearalenone-14-Glucuronide. Metabolites, 13(3), 446. https://doi.org/10.3390/metabo13030446