
Effect of Selenium Nanoparticles and/or Bee Venom against STZ-Induced Diabetic Cardiomyopathy and Nephropathy

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Animal Housing, Treatment and Grouping
2.3. Animal Grouping
2.4. Induction and Assessment of Diabetes
2.5. Blood Sampling and Tissue Collection
2.6. Evaluation of Biochemical Parameters
2.7. Evaluation of Micro RNAs (miR-328a and miR21) and Other Gene Transcriptional Levels in Renal and Cardiac Tissues
2.8. Histopathological Studies
2.9. Statistical Analysis
3. Results
3.1. The Effects of SeNPs and/or BV on Biochemical Parameters in the Serum of STZ-Diabetic Rats
3.2. The Effect of SeNPs and/or BV on the Expression Levels of miR-21 Gene in Renal Tissues, and miR-21 and miR-328 Genes in Cardiac Tissues
3.3. The Effect of SeNPs and/or BV on the Expression Levels of TGF-β1, NF-κβ and SMAD-7 Genes in Renal Tissues
3.4. The Effect of SeNPs and/or Bee Venom on the Expression Levels of TGF-β1, TGF-βR, JAK-1, STAT-3 and SMAD-1 Genes in Cardiac Tissues
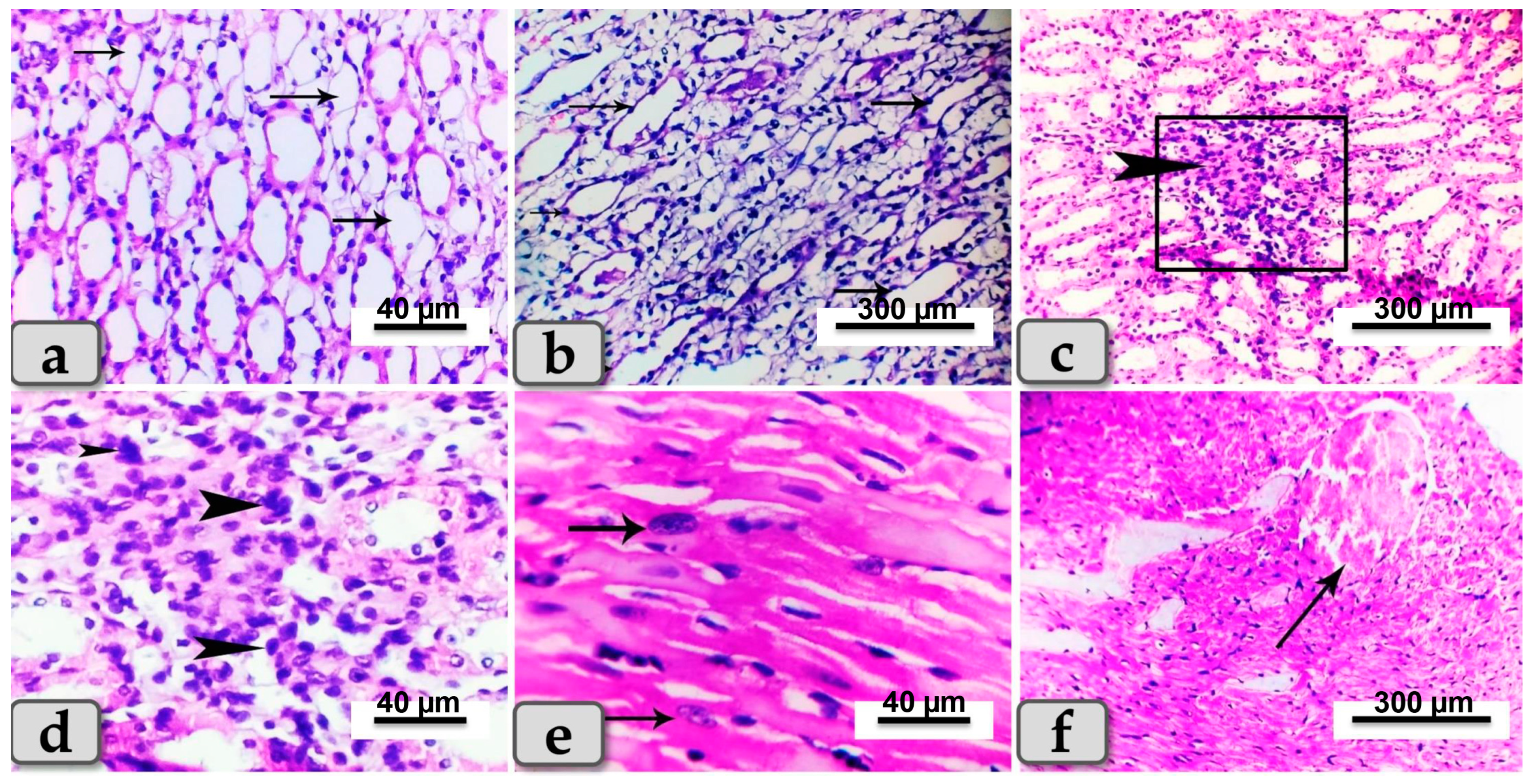
3.5. Histopathological Finding of Renal and Cardiac Tissues due to SeNP and/or BV Administration in STZ-Diabetic Rats
4. Discussion
4.1. Effect of SeNPs and/or Bee Venom on Blood Glucose and Insulin Concentrations
4.2. Biochemical and Histopathological Effects of SeNPs and/or BV on Renal and Cardiac Functions
4.3. Molecular Role of SeNPs and/or BV
4.3.1. Effect of SeNPS and/or BV on Expression Levels of miR-21 and miR-328 in Renal and Cardiac Tissues
4.3.2. Effect of SeNPs and /or BV on the Expression of Genes Involved in JAK/STAT, TGF-β/SMAD and TGF-β/NF-kβ Signaling Pathways in Renal and Cardiac Tissues
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2013, 36 (Suppl. S1), S67–S74. [Google Scholar] [CrossRef]
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef]
- Anfenan, M.L.K. Evaluation of nutritional and antidiabetic activity of different forms of Ginger in rats. Middle-East J. Sci. Res. 2014, 21, 56–62. [Google Scholar]
- El-Borady, O.M.; Othman, M.S.; Atallah, H.H.; Abdel Moneim, A.E. Hypoglycemic potential of selenium nanoparticles capped with polyvinyl-pyrrolidone in streptozotocin-induced experimental diabetes in rats. Heliyon 2020, 6, e04045. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.R.; Kassab, R.B.; Abdel Moneim, A.E.; Amin, H.K. Comparison among garlic, berberine, resveratrol, Hibiscus sabdariffa, genus zizyphus, hesperidin, red beetroot, catha edulis, portulaca oleracea, and mulberry leaves in the treatment of hypertension and type 2 DM: A comprehensive review. Nat. Prod. Commun. 2020, 15, 1934578X20921623. [Google Scholar]
- Jia, G.; Hill, M.A.; Sowers, J.M. Diabetic Cardiomyopathy. An Update of Mechanisms Contributing to This Clinical Entity. Circ. Res. 2018, 122, 624–638. [Google Scholar] [CrossRef]
- Fan, W. Epidemiology in diabetes mellitus and cardiovascular disease. Cardiovasc. Endocrinol. 2017, 6, 8–16. [Google Scholar] [CrossRef]
- Du, W.; Piek, A.; Schouten, E.M.; van de Kolk, C.W.A.; Mueller, C.; Mebazaa, A.; Voors, A.A.; de Boer, R.A.; Silljé, H.H.W. Plasma levels of heart failure biomarkers are primarily a reflection of extracardiac production. Theranostics 2018, 8, 4155–4169. [Google Scholar] [CrossRef] [PubMed]
- Gheith, O.; Farouk, N.; Nampoory, N.; Halim, M.A.; Al-Otaibi, T. Diabetic kidney disease: Worldwide difference of prevalence and risk factors. J. Nephropharmacol. 2015, 5, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Rabkin, R. Diabetic nephropathy. Clin. Cornerstone 2003, 5, 1–11. [Google Scholar] [CrossRef]
- American Diabetes Association. Standards of medical care in diabetes—2014. Diabetes Care 2014, 37 (Suppl. S1), S14–S80. [Google Scholar] [CrossRef]
- Lim, A.K.H. Diabetic nephropathy—Complications and treatment. Int. J. Nephrol. Renovasc. Dis. 2014, 7, 361–381. [Google Scholar] [CrossRef]
- Forbes, J.M.; Coughlan, M.T.; Cooper, M.E. Oxidative stress as a major culprit in kidney disease in diabetes. Diabetes 2008, 57, 1446–1454. [Google Scholar] [CrossRef] [PubMed]
- Molitch, M.E.; DeFronzo, R.A.; Franz, M.J.; Keane, W.F.; Mogensen, C.E.; Parving, H.H.; Steffes, M.W. American Diabetes Association Nephropathy in diabetes. Diabetes Care 2004, 27 (Suppl. S1), S79–S83. [Google Scholar]
- Kamianowska, M.; Szczepański, M.; Wasilewska, A. Tubular and Glomerular Biomarkers of Acute Kidney Injury in Newborns. Curr. Drug Metab. 2019, 20, 332–349. [Google Scholar] [CrossRef] [PubMed]
- Moghaddasi Kouchaksaraei, Y.; Ghazalian, F.; Abediankenari, S.; Ebrahim, K.; Abednatanzi, H. Determination of CRP blood level in type 1 diabetic patients and the effect of aerobic and resistance training on the level of this biomarker. Casp. J. Intern. Med. 2022, 13, 38–43. [Google Scholar] [CrossRef]
- Colhoun, H.M.; Schalkwijk, C.; Rubens, M.B.; Stehouwer, C.D. C-reactive protein in type 1 diabetes and its relationship to coronary artery calcification. Diabetes Care 2002, 25, 1813–1817. [Google Scholar] [CrossRef]
- Sproston, N.R.; Ashworth, J.J. Role of C—Reactive protein at Sites of Inflammation and Infection. Front. Immunol. 2018, 9, 754. [Google Scholar] [CrossRef]
- Van Genderen, M.E.; Lima, A.; de Geus, H.; Klijn, E.; Wijnhoven, B.; Gommers, D.; van Bommel, J. Serum c-reactive protein as a predictor of morbidity and mortality in intensive care unit patients after esophagectomy. Ann. Thorac. Surg. 2011, 14, 1775–1779. [Google Scholar] [CrossRef]
- Guo, R.; Nair, S. Role of microRNA in diabetic cardiomyopathy: From mechanism to intervention. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2070–2077. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, S. Role of microRNAs in the pathogenesis of diabetic cardiomyopathy. Biomed. Rep. 2017, 6, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Kiu, H.; Nicholson, S.E. Biology and significance of the JAK/STAT signalling pathways. Growth Factors 2012, 30, 88–106. [Google Scholar] [CrossRef]
- Kishore, R.; Verma, S.K. Roles of STATs signaling in cardiovascular diseases. Jak-Stat 2012, 1, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Xuan, Y.T.; Guo, Y.; Han, H.; Zhu, Y.; Bolli, R. An essential role of the JAK-STAT pathway in ischemic preconditioning. Proc. Natl. Acad. Sci. USA 2001, 98, 9050–9055. [Google Scholar] [CrossRef]
- Dobaczewski, M.; Chen, W.; Frangogiannis, N.G. Transforming growth factor (TGF)-β signaling in cardiac remodeling. J. Mol. Cell Cardiol. 2011, 51, 600–606. [Google Scholar] [CrossRef]
- Ling, H.; Fabbri, M.; Calin, G.A. MicroRNAs and other non-coding RNAs as targets for anticancer drug development. Nat. Rev. Drug Discov. 2013, 12, 847–865. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.Y.; Chang, D.C.; Lin, S.L. The microRNA (miRNA): Overview of the RNA genes that modulate gene function. Mol. Biotechnol. 2008, 38, 257–268. [Google Scholar] [CrossRef]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef]
- Moura, J.; Børsheim, E.; Carvalho, E. The Role of MicroRNAs in Diabetic Complications-Special Emphasis on Wound Healing. Genes 2014, 29, 926–956. [Google Scholar] [CrossRef]
- Margaritis, K.; Margioula-Siarkou, G.; Giza, S.; Kotanidou, E.P.; Tsinopoulou, V.R.; Christoforidis, A.; Galli-Tsinopoulou, A. Micro-RNA Implications in Type-1 Diabetes Mellitus: A Review of Literature. Int. J. Mol. Sci. 2021, 22, 12165. [Google Scholar] [CrossRef]
- Dey, N.; Das, F.; Mariappan, M.M.; Mandal, C.C.; Ghosh-Choudhury, N.; Kasinath, B.S.; Choudhury, G.G. MicroRNA-21 Orchestrates High Glucose-Induced Signals to TOR Complex 1, Resulting in Renal Cell Pathology in Diabetes. J. Biol. Chem. 2011, 286, 25586–25603. [Google Scholar] [CrossRef]
- Liu, S.; Wu, W.; Liao, J.; Tang, F.; Gao, G.; Peng, J.; Fu, X.; Zhan, Y.; Chen, Z.; Xu, W.; et al. MicroRNA-21: A Critical Pathogenic Factor of Diabetic Nephropathy. Front. Endocrinol. 2022, 13, 895010. [Google Scholar] [CrossRef]
- Krzywińska, O.; Bracha, M.; Jeanniere, C.; Recchia, E.; Kędziora Kornatowska, K.; Kozakiewicz, M. Meta-Analysis of the Potential Role of miRNA-21 in Cardiovascular System Function Monitoring. Biomed. Res. Int. 2020, 2020, 4525410. [Google Scholar] [CrossRef] [PubMed]
- Surina, F.R.A.; Scisciola, L.; Marfella, R.; Paolisso, G.; Barbieri, M. miR-21 in Human Cardiomyopathies. Front. Cardiovasc. Med. 2021, 8, 767064. [Google Scholar] [CrossRef] [PubMed]
- Kennel, P.J.; Schulze, P.C. A Review on the Evolving Roles of MiRNA-Based Technologies in Diagnosing and Treating Heart Failure. Cells 2021, 10, 3191. [Google Scholar] [CrossRef]
- Magenta, A.; Greco, S.; Gaetano, C.; Martelli, F. Oxidative stress and microRNAs in vascular diseases. Int. J. Mol. Sci. 2013, 14, 17319–17346. [Google Scholar] [CrossRef]
- La Sala, L.; Mrakic-Sposta, S.; Micheloni, S.; Prattichizzo, F.; Ceriello, A. Glucose-sensing microRNA-21 disrupts ROS homeostasis and impairs antioxidant responses in cellular glucose variability. Cardiovasc. Diabetol. 2018, 17, 105. [Google Scholar] [CrossRef]
- Assar, M.E.; Angulo, J.; Rodríguez-Mañas, L. Diabetes and ageing-induced vascular inflammation. J. Physiol. 2016, 594, 2125–2146. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Ponnusamy, M.; Liu, C.; Gao, J.; Wang, K.; Li, P. MicroRNA as a Therapeutic Target in Cardiac Remodeling. Biomed. Res. Int. 2017, 2017, 1278436. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Chen, H.; Liang, X.; Chen, X.; Chen, X.; Chen, C. Upregulated miR-328-3p and its high risk in atrial fibrillation: A systematic review and meta-analysis with meta-regression. Medicine 2022, 101, e28980. [Google Scholar] [CrossRef] [PubMed]
- Masè, M.; Grasso, M.; Avogaro, L.; Nicolussi Giacomaz, M.; D’Amato, E.; Tessarolo, F.; Graffigna, A.; Denti, M.A.; Ravelli, F. Upregulation of miR-133b and miR-328 in Patients with Atrial Dilatation: Implications for Stretch-Induced Atrial Fibrillation. Front. Physiol. 2019, 10, 1133. [Google Scholar] [CrossRef] [PubMed]
- Saadat, S.; Noureddini, M.; Mahjoubin-Tehran, M.; Nazemi, S.; Shojaie, L.; Aschner, M.; Maleki, B.; Abbasi-Kolli, M.; Rajabi Moghadam, H.; Alani, B.; et al. Pivotal Role of TGF-β/Smad Signaling in Cardiac Fibrosis: Non-coding RNAs as Effectual Players. Front. Cardiovasc. Med. 2021, 7, 588347. [Google Scholar] [CrossRef] [PubMed]
- Sureshbabu, A.; Muhsin, S.A.; Choi, M.E. TGF-β signaling in the kidney: Profibrotic and protective effects. Am. J. Physiol. Renal. Physiol. 2016, 310, F596–F606. [Google Scholar] [CrossRef]
- Hwang, D.; Seo, S.; Kim, Y.; Kim, C.; Shim, S.; Jee, S.; Lee, S.; Jang, M.; Kim, M.; Yim, S.; et al. Selenium acts as an insulin-like molecule for the down-regulation of diabetic symptoms via endoplasmic reticulum stress and insulin signalling proteins in diabetes-induced non-obese diabetic mice. J. Biosci. 2007, 32, 723–735. [Google Scholar] [CrossRef]
- Zolfagharian, H.; Mohajeri, M.; Babaie, M. Honey Bee Venom (Apis mellifera) Contains Anticoagulation Factors and Increases the Blood-clotting Time. J. Pharmacopunct. 2015, 18, 7–11. [Google Scholar] [CrossRef]
- Carpena, M.; Nuñez-Estevez, B.; Soria-Lopez, A.; Simal-Gandara, J. Bee Venom: An Updating Review of Its Bioactive Molecules and Its Health Applications. Nutrients 2020, 12, 3360. [Google Scholar] [CrossRef]
- Chen, J.; Guan, S.M.; Sun, W.; Fu, H. Melittin, the Major Pain-Producing Substance of Bee Venom. Neurosci. Bull. 2016, 32, 265–272. [Google Scholar] [CrossRef]
- Ceremuga, M.; Stela, M.; Janik, E.; Gorniak, L.; Synowiec, E.; Sliwinski, T.; Sitarek, P.; Saluk-Bijak, J.; Bijak, M. Melittin-A Natural Peptide from Bee Venom Which Induces Apoptosis in Human Leukaemia Cells. Biomolecules 2020, 10, 247. [Google Scholar] [CrossRef]
- Al-Quraishy, S.; Adkhil, M.; Moneim, A.E.A. Anti-hyperglycemic activity of selenium nanoparticles in streptozotocin-induced diabetic rats. Int. J. Nanomed. 2015, 10, 6741–6756. [Google Scholar] [CrossRef]
- El Adham, E.K.; Hassan, A.I.; Dawoud, M.M.A. Evaluating the role of propolis and bee venom on the oxidative stress induced by gamma rays in rats. Sci. Rep. 2022, 12, 2656. [Google Scholar] [CrossRef]
- Bhatt, L.K.; Addepalli, V. Attenuation of diabetic retinopathy by enhanced inhibition of MMP-2 and MMP-9 using aspirin and minocycline in streptozotocin-diabetic rats. Am. J. Transl. Res. 2010, 2, 181–189. [Google Scholar] [PubMed]
- Tietz, N.W. Clinical Guide to Laboratory Tests, 3rd ed.; WB Saunders: Philadelphia, PA, USA, 1995; pp. 268–273. [Google Scholar]
- Chevenne, D.; letailleur, A.; Trivin, F.; Porquet, D. Effect of hemolysis on the concentration of insulin in serum determined by RIA and IRMA. Clin. Chem. 1998, 44, 345–346. [Google Scholar] [CrossRef]
- Fawcett, J.K.; Scott, J.E. A rapid and precise method for the determination of urea. J. Clin. Pathol. 1960, 13, 156–159. [Google Scholar] [CrossRef]
- Bartels, H.; Böhmer, M.; Heierli, C. Serum Kreatinibestimmung ohne Enteiweissen [Serum creatinine determination without protein precipitation]. Clin. Chim Acta. 1972, 37, 193–197. [Google Scholar] [CrossRef]
- Doumas, B.T.; Watson, W.A.; Biggs, H.G. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta. 1971, 31, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Eckersall, P.D. Recent advances and future prospects for the use of acute phase proteins as markers of disease in animals. Rev. Med. Vet. 2000, 151, 577–584. [Google Scholar]
- Reitman, S.; Frankel, S.A. Colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C (T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Suvarna, S.K.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques, 8th ed.; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Alejandro, E.U.; Gregg, B.; Blandino-Rosano, M.; Cras-Méneur, C.; Bernal-Mizrachi, E. Natural history of β-cell adaptation and failure in type 2 diabetes. Mol. Asp. Med. 2015, 42, 19–41. [Google Scholar] [CrossRef] [PubMed]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Oguntibeju, O.O. Type 2 diabetes mellitus, oxidative stress and inflammation: Examining the links. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 45–63. [Google Scholar]
- Brown, R.J.; Rother, K.I. Effects of beta-cell rest on beta-cell function: A review of clinical and preclinical data. Pediatr. Diabetes. 2008, 3 Pt 2, 14–22. [Google Scholar] [CrossRef]
- Zahran, F.; Mohamad, A.; Zein, N. Bee venom ameliorates cardiac dysfunction in diabetic hyperlipidemic rats. Exp. Biol. Med. 2021, 246, 2630–2644. [Google Scholar] [CrossRef]
- Afkarian, M. Diabetic kidney disease in children and adolescents. Pediatr. Nephrol. 2015, 30, 65–74. [Google Scholar] [CrossRef]
- Lang, J.; Katz, R.; Ix, J.H.; Gutierrez, O.M.; Peralta, C.A.; Parikh, C.R.; Satterfield, S.; Petrovic, S.; Devarajan, P.; Bennett, M.; et al. Association of serum albumin levels with kidney function decline and incident chronic kidney disease in elders. Nephrol. Dial. Transplant. 2018, 33, 986–992. [Google Scholar] [CrossRef]
- Stanimirovic, J.; Radovanovic, J.; Banjac, K.; Obradovic, M.; Essack, M.; Zafirovic, S.; Gluvic, Z.; Gojobori, T.; Isenovic, E.R. Role of C—Reactive protein in Diabetic Inflammation. Mediat. Inflamm. 2022, 2022, 3706508. [Google Scholar] [CrossRef] [PubMed]
- García-García, P.M.; Getino-Melián, M.A.; Domínguez-Pimentel, V.; Navarro-González, J.F. Inflammation in diabetic kidney disease. World J. Diabetes 2014, 5, 431–443. [Google Scholar] [CrossRef]
- Matheus, A.S.; Tannus, L.R.; Cobas, R.A.; Palma, C.C.; Negrato, C.A.; Gomes, M.B. Impact of diabetes on cardiovascular disease: An update. Int. J. Hypertens 2013, 2013, 653789. [Google Scholar] [CrossRef] [PubMed]
- Jacob, R.; Khan, M. Cardiac Biomarkers: What Is and What Can Be. Indian J. Cardiovasc. Dis. Women WINCARS 2018, 3, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Doenst, T.; Nguyen, T.D.; Abel, E.D. Cardiac metabolism in heart failure: Implications beyond ATP production. Circ. Res. 2013, 113, 709–724. [Google Scholar] [CrossRef]
- Sivitz, W.I.; Yorek, M.A. Mitochondrial dysfunction in diabetes: From molecular mechanisms to functional significance and therapeutic opportunities. Antioxid. Redox Signal. 2010, 12, 537–577. [Google Scholar] [CrossRef]
- Babuin, L.; Jaffe, A.S. Troponin: The biomarker of choice for the detection of cardiac injury. Can. Med. Assoc. J. 2005, 173, 1191–1202. [Google Scholar] [CrossRef]
- Bang, C.; Batkai, S.; Dangwal, S.; Gupta, S.K.; Foinquinos, A.; Holzmann, A.; Just, A.; Remke, J.; Zimmer, K.; Zeug, A.; et al. Cardiac fibroblast-derived microRNA passenger strand-enriched exosomes mediate cardiomyocyte hypertrophy. J. Clin. Investig. 2014, 124, 2136–2146. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Liang, H.; Gao, X.; Li, X.; Zhang, Y.; Pan, Z.; Li, C.; Wang, Y.; Liu, Y.; Yuan, W.; et al. MicroRNA-328, a Potential Anti-Fibrotic Target in Cardiac Interstitial Fibrosis. Cell Physiol. Biochem. 2016, 39, 827–836. [Google Scholar] [CrossRef]
- Dai, X.; Fang, M.; Li, S.; Yan, Y.; Zhong, Y.; Du, B. miR-21 is involved in transforming growth factor β1-induced chemoresistance and invasion by targeting PTEN in breast cancer. Oncol. Lett. 2017, 14, 6929–6936. [Google Scholar] [CrossRef]
- Zhong, X.; Chung, A.C.; Chen, H.Y.; Meng, X.M.; Lan, H.Y. Smad3-mediated upregulation of miR-21 promotes renal fibrosis. J. Am. Soc. Nephrol. 2011, 22, 1668–1681. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.S.; Hathaway, C.K.; Smithies, O.; Kakoki, M. Transforming growth factor-β1 and diabetic nephropathy. Am. J. Physiol. Ren. Physiol. 2016, 310, F689–F696. [Google Scholar] [CrossRef]
- Frangogiannis, N. Transforming growth factor-β in tissue fibrosis. J. Exp. Med. 2020, 217, e20190103. [Google Scholar] [CrossRef]
- Liu, M.; Li, Y.; Liang, B.; Li, Z.; Jiang, Z.; Chu, C.; Yang, J. Hydrogen sulfide attenuates myocardial fibrosis in diabetic rats through the JAK/STAT signaling pathway. Int. J. Mol. Med. 2018, 41, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Nowotny, K.; Jung, T.; Höhn, A.; Weber, D.; Grune, T. Advanced glycation end products and oxidative stress in type 2 diabetes mellitus. Biomolecules 2015, 5, 194–222. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Chung, A.C.K.; Chen, H.Y.; Dong, Y.; Meng, X.M.; Li, R.; Yang, W.; Hou, F.F.; Lan, H.Y. miR-21 is a Key Therapeutic Target for Renal Injury in a Mouse Model of Type 2 Diabetes. Diabetologia 2013, 56, 663–674. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Organ | Accession No. | Expected Size |
|---|---|---|---|---|
| TGFβ1 | 5′-AGGGCTACCATGCCAACTTC-3′ 5′-CCACGTAGTAGACGATGGGC-3′ | Heart and kidney | NM_021578.2 | 186 |
| TGFβR | 5′-CAGGAGGTGAAAGTCCCCG-3′ 5′-CACTGTTCAACTTGCCTCGC-3′ | Heart | NM_017256.1 | 172 |
| JAK-1 | 5′-AGGAATGTACTGGGCGTCTT-3′ 5′-GGTTGTCCAGTGTCCCTGAAA-3′ | Heart | NM_053466.1 | 107 |
| STAT-3 | 5′-GGTACAATCCCGCTCGGTG-3′ 5′-AGCTGGTTCCACTGAGCCAT-3′ | Heart | NM_012747.2 | 169 |
| SMAD-1 | 5′-TCAATAGAGGAGATGTTCAAGCAGT-3′ 5′-GAAACCATCCACCAACACGC-3′ | Heart | NM_013130.3 | 134 |
| β-actin | 5′-CCCGCGAGTACAACCTTCTT-3′ 5′-CGCAGCGATATCGTCATCCA-3′ | Heart | NM_031144.3 | 83 |
| NF-кB | 5′-CAGGACCAGGAACAGTTCGAA-3′ 5′-CCAGGTTCTGGAAGCTATGGAT-3′ | Kidney | NM_199267.2 | 150 |
| SMAD7 | 5′-GAGTCTCGGAGGAAGAGGCT-3′ 5′-CTGCTCGCATAAGCTGCTGG-3′ | Kidney | NM_030858.2 | 84 |
| GAPDH | 5′-GCATCTTCTTGTGCAGTGCC-3′ 5′-GGTAACCAGGCGTCCGATAC-3′ | Kidney | NM_017008.4 | 91 |
| rno-miR-21-5p | RT primer 5′-GTCGTATCCAGTGCAGGGT- CCGAGGTATTCGCACTGGATACGACTCAACA-3′ | F5′-AGCGACTAGCTTATCAGACT-3′ R 5′-GTCGTATCCAGTCAGGGT-3′ | ||
| rno-miR-328a-5p | RT primer 5′-GTTGGCTCTGGTGCAGGGT- CCGAGGTATTCGCACCAGAGCCAACTGAGCC-3′ | F 5′-GTTTTTGGGGGGCAGGAG-3′ R 5′-GTGCAGGGTCCGAGGT-3′ | ||
| snRNA U6 | RT primer 5′-AACGCTTCACGAATTTGCGT-3′ | F 5′-CTCGCTTCGGCAGCACA-3′ R 5′-AACGCTTCACGAATTTCG-T-3′ | ||
| Control | Diabetic | Diabetic + SeNPs | Diabetic + Bee Venom | Diabetic + SeNPs + Bee Venom | |
|---|---|---|---|---|---|
| Glucose conc. (mg/dL) | 167.51 ± 9.37 c | 334.05 ± 13.68 a | 261.18 ± 11.20 b | 233.27 ± 12.28 c | 206.04 ± 11.39 d |
| Serum insulin conc. (µlU/mL). | 1.63 ± 0.24 a | 0.45 ± 0.04 e | 1.06 ± 0.20 c | 0.76 ± 0.12 d | 1.41 ± 0.09 b |
| Serum BUN (mg/dL) | 14.25 ± 1.67 d | 33.65 ± 3.89 a | 23.13 ± 3.2 b | 21.88± 2.94 b | 17.41 ± 1.64 c |
| serum creatinine conc. (mg/dL) | 1.1 ± 0.02 d | 1.89 ± 0.23 a | 1.43 ± 0.18 b | 1.38 ± 0.08 b | 1.21 ± 0.04 c |
| Serum albumin (gm%) | 6.1 ± 0.23 d | 2.65 ± 0.12 a | 3.74 ± 0.27 b | 3.82 ± 0.21 b | 4.68 ± 0.22 c |
| serum CRP (ng/mL) | 4.85 ± 0.39 d | 11.52 ± 1.42 a | 8.42 ± 1.06 b | 7.95 ± 1.1 b | 5.42 ± 0.68 c |
| serum CK-MB (IU/L) | 184.5 ± 9.45 d | 862.8 ± 19.6 a | 683.2 ± 18.7 b | 582.5 ± 9.1 b | 312.8 ± 10.8 c |
| serum AST conc. (IU/L) | 151.44 ± 11.38 e | 521.33 ± 19.83 a | 411.2 ± 18.6 b | 378.54 ± 16.1 c | 225.28 ± 13.53 d |
| serum LDH conc. (IU/L) | 320.5 ± 8.5 e | 809.8 ± 15.5 a | 511.9 ± 26.5 b | 491.6 ± 16.5 c | 403.4 ± 12.7 d |
| cTnI (ng/mL) | 0.65 ± 0.15 e | 1.83 ± 0.18 a | 1.29 ± 0.08 b | 1.26 ± 0.09 c | 0.94 ± 0.07 d |
| cTnT (pg/mL) | 42.81 ±5.42 e | 276.8 ± 21.8 a | 182.9 ± 9.7 b | 167.6 ± 11.6 c | 88.4 ± 7.52 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lotfy, M.M.; Dowidar, M.F.; Ali, H.A.; Ghonimi, W.A.M.; AL-Farga, A.; Ahmed, A.I. Effect of Selenium Nanoparticles and/or Bee Venom against STZ-Induced Diabetic Cardiomyopathy and Nephropathy. Metabolites 2023, 13, 400. https://doi.org/10.3390/metabo13030400
Lotfy MM, Dowidar MF, Ali HA, Ghonimi WAM, AL-Farga A, Ahmed AI. Effect of Selenium Nanoparticles and/or Bee Venom against STZ-Induced Diabetic Cardiomyopathy and Nephropathy. Metabolites. 2023; 13(3):400. https://doi.org/10.3390/metabo13030400
Chicago/Turabian StyleLotfy, Mona M., Mohamed F. Dowidar, Haytham A. Ali, Wael A. M. Ghonimi, Ammar AL-Farga, and Amany I. Ahmed. 2023. "Effect of Selenium Nanoparticles and/or Bee Venom against STZ-Induced Diabetic Cardiomyopathy and Nephropathy" Metabolites 13, no. 3: 400. https://doi.org/10.3390/metabo13030400
APA StyleLotfy, M. M., Dowidar, M. F., Ali, H. A., Ghonimi, W. A. M., AL-Farga, A., & Ahmed, A. I. (2023). Effect of Selenium Nanoparticles and/or Bee Venom against STZ-Induced Diabetic Cardiomyopathy and Nephropathy. Metabolites, 13(3), 400. https://doi.org/10.3390/metabo13030400