
Prenatal SSRI Exposure Increases the Risk of Autism in Rodents via Aggravated Oxidative Stress and Neurochemical Changes in the Brain



Abstract
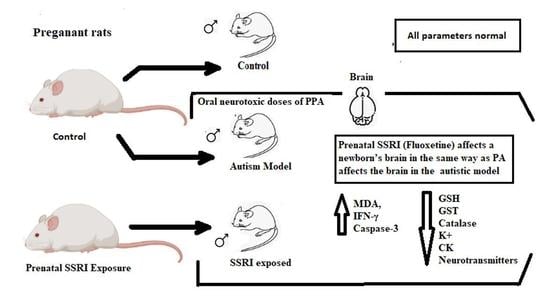
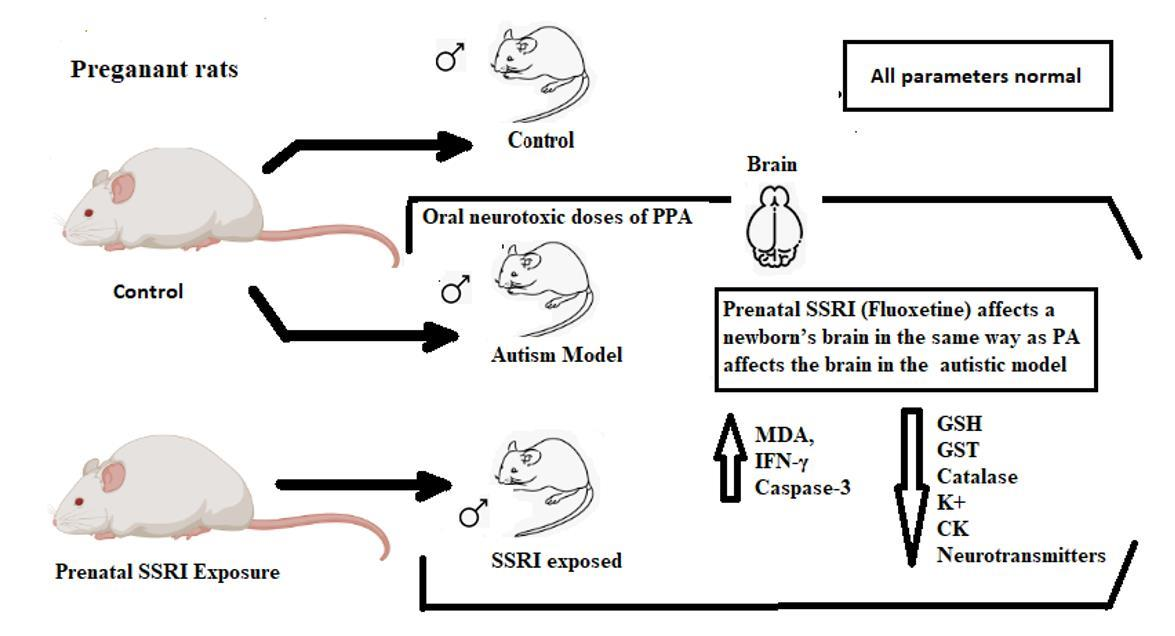
1. Introduction
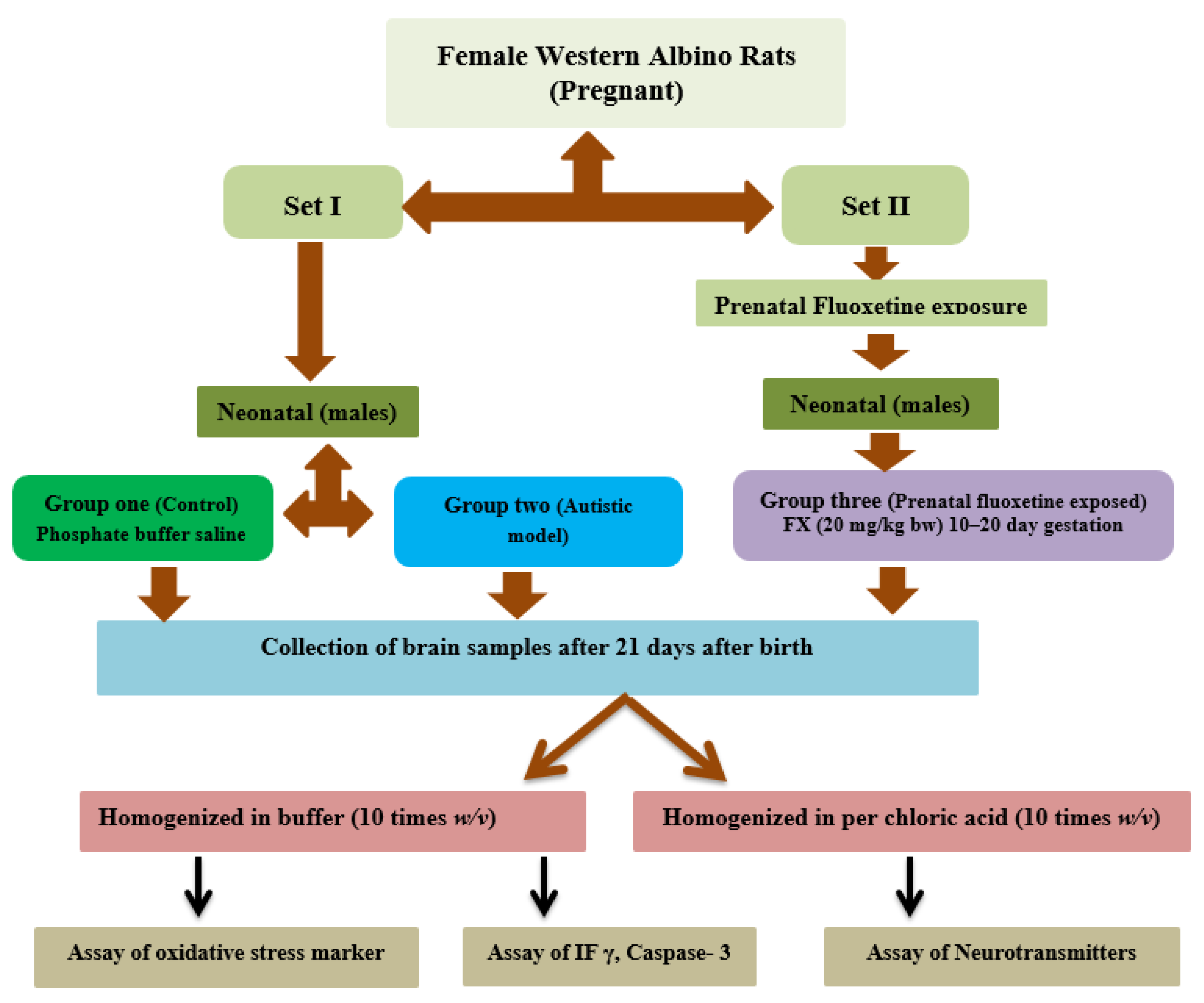
2. Materials and Methods
2.1. Animals
2.2. Brain Tissue Collection
2.3. Biochemical Analyses
2.3.1. Lipid Peroxidation
2.3.2. Glutathione
2.3.3. Glutathione-S-Transferase
2.3.4. Catalase
2.3.5. Potassium
2.3.6. Neurotransmitter
2.3.7. Creatine Kinase
2.3.8. Caspase-3
2.3.9. Interferon Gamma
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Israelyan, N.; Margolis, K.G. Serotonin as a link between the gut-brain-microbiome axis in autism spectrum disorders. Pharmacol. Res. 2018, 132, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.; Courtiol, E.; Castellanos, F.X.; Teixeira, C.M. Abnormal Serotonin Levels During Perinatal Development Lead to Behavioral Deficits in Adulthood. Front. Behav. Neurosci. 2018, 12, 114. [Google Scholar] [CrossRef] [PubMed]
- Lesch, K.-P.; Waider, J. Serotonin in the Modulation of Neural Plasticity and Networks: Implications for Neurodevelopmental Disorders. Neuron 2012, 76, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Azmitia, E.C. Modern views on an ancient chemical: Serotonin effects on cell proliferation, maturation, and apoptosis. Brain Res. Bull. 2001, 56, 413–424. [Google Scholar] [CrossRef]
- Saldanha, D.; Kumar, N.; Ryali, V.; Srivastava, K.; Pawar, A. Serum Serotonin Abnormality in Depression. Med. J. Armed Forces India 2009, 65, 108–112. [Google Scholar] [CrossRef]
- Abdel-Salam, O.M.; Morsy, S.M.Y.; Sleem, A.A. The effect of different antidepressant drugs on oxidative stress after lipopolysaccharide administration in mice. EXCLI J. 2011, 10, 290–302. [Google Scholar] [PubMed] [PubMed Central]
- Ornoy, A.; Koren, G. SSRIs and SNRIs (SRI) in Pregnancy: Effects on the Course of Pregnancy and the Offspring: How Far Are We from Having All the Answers? Int. J. Mol. Sci. 2019, 20, 2370. [Google Scholar] [CrossRef]
- Rotem-Kohavi, N.; Williams, L.J.; Virji-Babul, N.; Bjornson, B.; Brain, U.; Werker, J.F.; Grunau, R.E.; Miller, S.; Oberlander, T.F. Alterations in Resting-State Networks Following In Utero Selective Serotonin Reuptake Inhibitor Exposure in the Neonatal Brain. Biol. Psychiatry: Cogn. Neurosci. Neuroimaging 2018, 4, 39–49. [Google Scholar] [CrossRef]
- Yonkers, K.A.; Blackwell, K.A.; Glover, J.; Forray, A. Antidepressant Use in Pregnant and Postpartum Women. Annu. Rev. Clin. Psychol. 2014, 10, 369–392. [Google Scholar] [CrossRef]
- Pei, S.; Liu, L.; Zhong, Z.; Wang, H.; Lin, S.; Shang, J. Risk of prenatal depression and stress treatment: Alteration on serotonin system of offspring through exposure to Fluoxetine. Sci. Rep. 2016, 6, 33822. [Google Scholar] [CrossRef]
- Kristensen, J.H.; Ilett, K.F.; Hackett, L.P.; Yapp, P.; Paech, M.; Begg, E.J. Distribution and excretion of fluoxetine and norfluoxetine in human milk. Br. J. Clin. Pharmacol. 1999, 48, 521–527. [Google Scholar] [CrossRef]
- Zengeler, K.E.; Shapiro, D.A.; Bruch, K.R.; Lammert, C.R.; Ennerfelt, H.; Lukens, J.R. SSRI treatment modifies the effects of maternal inflammation on in utero physiology and offspring neurobiology. Brain, Behav. Immun. 2023, 108, 80–97. [Google Scholar] [CrossRef]
- Estes, M.L.; McAllister, A.K. Maternal immune activation: Implications for neuropsychiatric disorders. Science 2016, 353, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Han, V.X.; Patel, S.; Jones, H.F.; Dale, R.C. Maternal immune activation and neuroinflammation in human neurodevelopmental disorders. Nat. Rev. Neurol. 2021, 17, 564–579. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.S.; Derkits, E.J. Prenatal Infection and Schizophrenia: A Review of Epidemiologic and Translational Studies. Am. J. Psychiatry 2010, 167, 261–280. [Google Scholar] [CrossRef]
- Casper, R.C.; Fleisher, B.E.; Lee-Ancajas, J.C.; Gilles, A.; Gaylor, E.; DeBattista, A.; Hoyme, H. Follow-up of children of depressed mothers exposed or not exposed to antidepressant drugs during pregnancy. J. Pediatr. 2003, 142, 402–408. [Google Scholar] [CrossRef]
- Pedersen, L.H.; Henriksen, T.B.; Olsen, J. Fetal exposure to antidepressantand normal milestone development at 6 and 19 months of age. Pediatrics 2010, 125, e600–e608. [Google Scholar] [CrossRef] [PubMed]
- Oberlander, T.F.; Grunau, R.E.; Fitzgerald, C.; Papsdorf, M.; Rurak, D.; Riggs, W. Pain reactivity in 2-month-old infants after prenatal and postnatal selectiveserotonin reuptake inhibitor medication exposure. Pediatrics 2005, 115, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Croen, L.A.; Grether, J.K.; Yoshida, C.K.; Odouli, R.; Hendrick, V. Antidepressant use during pregnancy and childhood autism spectrum disorders. Arch Gen Psychiatry 2011, 68, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Muller, C.; Anacker, A.; Veenstra-VanderWeele, J. The serotonin system in autism spectrum disorder: From biomarker to animal models. Neuroscience 2016, 321, 24–41. [Google Scholar] [CrossRef]
- De Grandis, E.; Serrano, M.; Pérez-Dueñas, B.; Ormazábal, A.; Montero, R.; Veneselli, E.; Pineda, M.; González, V.; Sanmartí, F.; Fons, C.; et al. Cerebrospinal fluid alterations of the serotonin product, 5-hydroxyindolacetic acid, in neurological disorders. J. Inherit. Metab. Dis. 2010, 33, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Mulder, E.J.; Anderson, G.M.; Kema, I.P.; De Bildt, A.; Van Lang, N.D.; Den Boer, J.A.; Minderaa, R.B. Platelet serotonin levels in pervasive developmental disorders and mental retardation: Diagnostic group differences, within-group distribution, and behavioral correlates. J. Am. Acad. Child Adolesc. Psychiatry 2004, 43, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Hodges, H.; Fealko, C.; Soares, N. Autism spectrum disorder: Definition, epidemiology, causes, and clinical evaluation. Transl. Pediatr. 2020, 9, S55. [Google Scholar] [CrossRef] [PubMed]
- Malwane, M.I.; Nguyen, E.B.; Trejo, S.; Kim, E.Y.; Cucalón-Calderón, J.R. A Delayed Diagnosis of Autism Spectrum Disorder in the Setting of Complex Attention Deficit Hyperactivity Disorder. Cureus 2022, 14, e25825. [Google Scholar] [CrossRef]
- Maenner, M.J.; Shaw, K.A.; Bakian, A.V.; Bilder, D.A.; Durkin, M.S.; Esler, A.; Furnier, S.M.; Hallas, L.; Hall-Lande, J.; Hudson, A.; et al. Prevalence and characteristics of autism spectrum disorder among children aged 8 years—Autism and Developmental Disabilities Monitoring Network, 11 sites, United States, 2018. MMWR Surveill Summ. 2021, 70, 1–16. [Google Scholar] [CrossRef]
- Yang, T.; Chen, L.; Dai, Y.; Jia, F.; Hao, Y.; Li, L.; Zhang, J.; Wu, L.; Ke, X.; Yi, M.; et al. Vitamin A Status Is More CommonlyAssociated with Symptoms and Neurodevelopment in Boys with Autism Spectrum Disorders-A Multicenter Study in China. Front. Nutr. 2022, 9, 851980. [Google Scholar] [CrossRef]
- Wang, L.; Wang, B.; Wu, C.; Wang, J.; Sun, M. Autism Spectrum Disorder: Neurodevelopmental Risk Factors, Biological Mechanism, and Precision Therapy. Int. J. Mol. Sci. 2023, 24, 1819. [Google Scholar] [CrossRef]
- Kapra, O.; Rotem, R.; Gross, R. The Association Between Prenatal Exposure to Antidepressants and Autism: Some Research and Public Health Aspects. Front. Psychiatry 2020, 11, 55740. [Google Scholar] [CrossRef]
- Mathew, S.; Bichenapally, S.; Khachatryan, V.; Muazzam, A.; Hamal, C.; Velugoti, L.S.D.R.; Tabowei, G.; Gaddipati, G.N.; Mukhtar, M.; Alzubaidee, M.J.; et al. Role of Serotoninergic Antidepressants in the Development of Autism Spectrum Disorders: A Systematic Review. Cureus 2022, 14, e28505. [Google Scholar] [CrossRef]
- Sujan, A.C.; Öberg, A.S.; Quinn, P.D.; D’Onofrio, B.M. Annual Research Review: Maternal antidepressant use during pregnancy and offspring neurodevelopmental problems—A critical review and recommendations for future research. J. Child Psychol. Psychiatry 2018, 60, 356–376. [Google Scholar] [CrossRef]
- El-Ansary, A.K.; Ben Bacha, A.; Kotb, M. Etiology of autistic features: The persisting neurotoxic effects of propionic acid. J. Neuroinflammation 2012, 9, 74. [Google Scholar] [CrossRef] [PubMed]
- Jiji, K.N.; Muralidharan, P. Evaluation of the protective effect of Clitoria ternatea L. against propionic acid induced autistic spectrum disorders in rat model. Bull. Natl. Res. Cent. 2022, 46, 71. [Google Scholar] [CrossRef]
- Abuaish, S.; Al-Otaibi, N.M.; Abujamel, T.S.; Alzahrani, S.A.; Alotaibi, S.M.; AlShawakir, Y.A.; Aabed, K.; El-Ansary, A. Fecal Transplant and Bifidobacterium Treatments Modulate Gut Clostridium Bacteria and Rescue Social Impairment and Hippocampal BDNF Expression in a Rodent Model of Autism. Brain Sci. 2021, 11, 1038. [Google Scholar] [CrossRef]
- Pawluski, J.L.; Van Donkelaar, E.; Abrams, Z.; Houbart, V.; Fillet, M.; Steinbusch, H.W.M.; Charlier, T.D. Fluoxetine Dose and Administration Method Differentially Affect Hippocampal Plasticity in Adult Female Rats. Neural Plast. 2014, 2014, 123026. [Google Scholar] [CrossRef]
- Bhat, R.S.; GezeerY, A.E.; Bachan, A.B.; AlonazI, M.A.; Alsuhaibani, L.S.; El-Ansary, A. Prenatal exposure to the fluoride containing psychiatric drug fluoxetine and anti-oxidative alterations in the neonatal rat brain. Biocell 2019, 43, 65–71. [Google Scholar]
- Ruiz-Larrea, M.B.; Leal, A.M.; Liza, M.; Lacort, M.; de Groot, H. Antioxidant effects of estradiol and 2-hydroxyestradiol on iron-induced lipid peroxidation of rat liver microsomes. Steroids 1994, 59, 383–388. [Google Scholar] [CrossRef]
- Beutle, E.; Duran, O.; Kelly, B.M. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Chance, B. Catalases and peroxidess, part II. Special methods. Methods Biochem. Anal. 1954, 1, 408. [Google Scholar]
- Terri, A.E.; Sesin, P.G. Determination of potassium in blood serum. Am. Soc. Clin. Pathol. 1958, 29, 86–89. [Google Scholar] [CrossRef]
- Zagrodzka, J.; Romaniuk, A.; Wieczorek, M.; Boguszewski, P. Bicuculline administration into ventromedial hypothalamus: Effects on fear and regional brainmonoamines and GABA concentrations in rats. Acta Neurobiol. Exp. 2000, 60, 333–343. [Google Scholar]
- Boukhris, T.; Sheehy, O.; Mottron, L.; Bérard, A. Antidepressant use during pregnancy and the risk of autism spectrum disorder in children. JAMA Pediatr. 2016, 170, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.K.; Hussain-Shamsy, N.; Lunsky, Y.; Dennis, C.L.E.; Vigod, S.N. The Association between antenatal exposure to selective serotonin reuptake inhibitors and autism: A systematic review and meta-analysis. J. Clin. Psychiatry 2017, 78, e48. [Google Scholar] [CrossRef] [PubMed]
- Gidaya, N.B.; Lee, B.K.; Burstyn, I.; Yudell, M.; Mortensen, E.L.; Newschaffer, C.J. In utero exposure to selective serotonin reuptake inhibitors and risk for autism spectrum disorder. J. Autism. Dev. Disord. 2014, 44, 2558–2567. [Google Scholar] [CrossRef] [PubMed]
- Rai, D.; Lee, B.K.; Dalman, C.; Golding, J.; Lewis, G.; Magnusson, C. Parental depression, maternalantidepressant use during pregnancy, and risk of autism spectrum disorders:population based case-control study. BMJ 2013, 346, f2059. [Google Scholar] [CrossRef]
- Shen, H.H. News Feature: Better models for brain disease. Proc. Natl. Acad. Sci. USA 2016, 113, 5461–5464. [Google Scholar] [CrossRef]
- Bölte, S.; Girdler, S.; Marschik, P.B. The contribution of environmental exposure to the etiology of autism spectrum disorder. Cell. Mol. Life Sci. 2019, 76, 1275–1297. [Google Scholar] [CrossRef]
- Pugsley, K.; Scherer, S.W.; Bellgrove, M.A.; Hawi, Z. Environmental exposures associated with elevated risk for autism spectrum disorder may augment the burden of deleterious de novo mutations among probands. Mol. Psychiatry 2021, 27, 710–730. [Google Scholar] [CrossRef]
- Parker, W.; Hornik, C.D.; Bilbo, S.; Holzknecht, Z.E.; Gentry, L.; Rao, R.; Lin, S.S.; Herbert, M.R.; Nevison, C.D. The role of oxidative stress, inflammation and acetaminophen exposure from birth to early childhood in the induction of autism. J. Int. Med. Res. 2017, 45, 407–438. [Google Scholar] [CrossRef]
- Rossignol, D.A.; Genuis, S.J.; Frye, R.E. Environmental toxicants and autism spectrum disorders: A systematic review. Transl. Psychiatry 2014, 4, e360. [Google Scholar] [CrossRef]
- Quaak, I.; Brouns, M.R.; Van de Bor, M. The dynamics of autism spectrum disorders: How neurotoxic compounds and neurotransmitters interact. Int. J. Environ. Res. Public Health 2013, 10, 3384–3408. [Google Scholar] [CrossRef]
- Janecka, M.; Kodesh, A.; Levine, S.Z.; Lusskin, S.I.; Viktorin, A.; Rahman, R.; Buxbaum, J.D.; Schlessinger, A.; Sandin, S.; Reichenberg, A. Association of Autism Spectrum Disorder With Prenatal Exposure to Medication Affecting Neurotransmitter Systems. JAMA Psychiatry 2018, 75, 1217–1224. [Google Scholar] [CrossRef]
- Goines, P.E.; Ashwood, P. Cytokine dysregulation in autism spectrum disorders (ASD): Possible role of the environment. Neurotoxicology Teratol. 2012, 36, 67–81. [Google Scholar] [CrossRef]
- Goines, P.E.; Croen, L.A.; Braunschweig, D.; Yoshida, C.K.; Grether, J.; Hansen, R.; Kharrazi, M.; Ashwood, P.; Van de Water, J. Increased midgestational IFN-γ, IL-4 and IL-5 in women bearing a child with autism: A case-control study. Mol. Autism 2011, 2, 13. [Google Scholar] [CrossRef]
- Robinson-Agramonte, M.D.L.A.; García, E.N.; Guerra, J.F.; Hurtado, Y.V.; Antonucci, N.; Semprún-Hernández, N.; Schultz, S.; Siniscalco, D. Immune Dysregulation in Autism Spectrum Disorder: What Do We Know about It? Int. J. Mol. Sci. 2022, 23, 3033. [Google Scholar] [CrossRef]
- Frye, R.E. Mitochondrial Dysfunction in Autism Spectrum Disorder: Unique Abnormalities and Targeted Treatments. Semin. Pediatr. Neurol. 2020, 35, 100829. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Parameters | Groups | N | Min. | Max. | Mean ± S.D. | Percent Change | p Value a | p Value b |
|---|---|---|---|---|---|---|---|---|
| MDA (µmoles/mL) | Control | 10 | 0.31 | 0.35 | 0.330 ± 0.014 | 100.00 | 0.001 | |
| PPA | 10 | 0.45 | 0.47 | 0.460 ± 0.009 | 139.39 | 0.001 | ||
| FLX | 10 | 0.35 | 0.37 | 0.360 ± 0.009 | 109.09 | 0.001 | ||
| GSH (ug/mL) | Control | 10 | 33.20 | 37.55 | 36.19 ± 1.66 | 100.00 | 0.001 | |
| PPA | 10 | 28.99 | 31.20 | 30.04 ± 0.83 | 83.00 | 0.001 | ||
| FLX | 10 | 15.60 | 17.60 | 16.45 ± 0.86 | 45.45 | 0.001 | ||
| Catalase (U/dl) | Control | 10 | 5.25 | 7.50 | 6.23 ± 0.87 | 100.00 | 0.001 | |
| PPA | 10 | 4.20 | 5.30 | 4.67 ± 0.45 | 75.01 | 0.002 | ||
| FLX | 10 | 3.89 | 5.20 | 4.59 ± 0.56 | 73.73 | 0.001 | ||
| GST (U/mL) | Control | 10 | 13.22 | 18.22 | 15.47 ± 1.71 | 100.00 | 0.001 | |
| PPA | 10 | 11.22 | 13.80 | 12.55 ± 0.97 | 81.11 | 0.002 | ||
| FLX | 10 | 7.10 | 9.60 | 8.56 ± 0.84 | 55.33 | 0.001 | ||
| K+ (mmol/L) | Control | 10 | 5.10 | 6.33 | 5.60 ± 0.53 | 100.00 | 0.001 | |
| PPA | 10 | 4.31 | 5.22 | 4.82 ± 0.34 | 86.01 | 0.010 | ||
| FLX | 10 | 2.50 | 3.44 | 2.94 ± 0.35 | 52.43 | 0.001 | ||
| CK (IU/L) | Control | 10 | 250.33 | 311.80 | 284.91 ± 22.10 | 100.00 | 0.001 | |
| PPA | 10 | 275.55 | 300.22 | 287.07 ± 10.38 | 100.76 | 0.954 | ||
| FLX | 10 | 320.77 | 340.89 | 330.94 ± 7.17 | 116.16 | 0.001 | ||
| IF γ (pg/100 mg) | Control | 10 | 87.50 | 90.44 | 89.18 ± 1.15 | 100.00 | 0.001 | |
| PPA | 10 | 111.44 | 115.33 | 113.50 ± 1.42 | 127.27 | 0.001 | ||
| FLX | 10 | 90.40 | 99.55 | 93.10 ± 3.28 | 104.39 | 0.013 | ||
| Norepinephrine (ng/100 mg) | Control | 10 | 4.55 | 5.50 | 5.05 ± 0.32 | 100.00 | 0.001 | |
| PPA | 10 | 3.09 | 3.88 | 3.46 ± 0.27 | 68.55 | 0.001 | ||
| FLX | 10 | 3.99 | 5.11 | 4.82 ± 0.42 | 95.45 | 0.416 | ||
| Serotonin (ng/100 mg) | Control | 10 | 5.95 | 7.10 | 6.63 ± 0.39 | 100.00 | 0.001 | |
| PPA | 10 | 3.99 | 4.42 | 4.23 ± 0.14 | 63.87 | 0.001 | ||
| FLX | 10 | 4.50 | 5.02 | 4.77 ± 0.18 | 72.03 | 0.001 | ||
| Dopamine (ng/100 mg) | Control | 10 | 20.97 | 22.50 | 21.88 ± 0.50 | 100.00 | 0.001 | |
| PPA | 10 | 14.59 | 16.00 | 15.08 ± 0.51 | 68.92 | 0.001 | ||
| FLX | 10 | 15.17 | 17.09 | 16.08 ± 0.76 | 73.48 | 0.001 | ||
| Caspase-3 (pg/100 mg) | Control | 10 | 108.40 | 113.18 | 110.83 ± 1.76 | 100.00 | 0.001 | |
| PPA | 10 | 138.90 | 145.30 | 142.56 ± 2.40 | 128.62 | 0.001 | ||
| FLX | 10 | 132.99 | 135.15 | 134.03 ± 0.93 | 120.93 | 0.001 |
| Dependent Variable | Predictor Variable | Coefficient | p Value | Adjusted R2 | 95% CI | |
|---|---|---|---|---|---|---|
| Lower | Upper | |||||
| MDA | IF γ | 0.005 | 0.000 | 0.950 | 0.004 | 0.006 |
| K+ | GSH | 0.134 | 0.000 | 0.887 | 0.110 | 0.159 |
| CK | −0.017 | 0.010 | −0.029 | −0.005 | ||
| CK | K+ | −16.297 | 0.000 | 0.922 | −22.599 | −9.996 |
| Catalase | 10.859 | 0.000 | 6.435 | 15.283 | ||
| GST | −3.047 | 0.021 | −5.552 | −0.542 | ||
| IF γ | MDA | 187.386 | 0.000 | 0.950 | 165.202 | 209.569 |
| Serotonin | Caspase-3 | −0.076 | 0.000 | 0.950 | −0.085 | −0.067 |
| Dopamine | Caspase-3 | −0.192 | 0.000 | 0.958 | −0.222 | −0.163 |
| GST | 0.202 | 0.005 | 0.072 | 0.331 | ||
| Caspase-3 | Serotonin | −5.793 | 0.002 | 0.984 | −9.090 | −2.495 |
| Dopamine | −0.800 | 0.209 | −2.107 | 0.507 | ||
| MDA | 82.120 | 0.000 | 44.826 | 119.413 | ||
| K+ | −2.005 | 0.011 | −3.478 | −0.532 | ||
| Parameters | Groups | AUC | Cut-off Value | Sensitivity % | Specificity % | p Value | 95% CI |
|---|---|---|---|---|---|---|---|
| MDA (µmoles/mL) | PPA | 1.000 | 0.400 | 100.0% | 100.0% | 0.004 | 1.000–1.000 |
| FLX | 0.972 | 0.345 | 100.0% | 83.3% | 0.006 | 0.889–1.056 | |
| GSH (ug/mL) | PPA | 1.000 | 32.200 | 100.0% | 100.0% | 0.004 | 1.000–1.000 |
| FLX | 1.000 | 25.400 | 100.0% | 100.0% | 0.004 | 1.000–1.000 | |
| Catalase (U/dl) | PPA | 0.972 | 5.370 | 100.0% | 83.3% | 0.006 | 0.889–1.056 |
| FLX | 1.000 | 5.225 | 100.0% | 100.0% | 0.004 | 1.000–1.000 | |
| GST (U/mL) | PPA | 0.958 | 14.100 | 100.0% | 83.3% | 0.008 | 0.851–1.065 |
| FLX | 1.000 | 11.410 | 100.0% | 100.0% | 0.004 | 1.000–1.000 | |
| K+ (mmol/L) | PPA | 0.917 | 5.050 | 83.3% | 100.0% | 0.016 | 0.742–1.091 |
| FLX | 1.000 | 4.270 | 100.0% | 100.0% | 0.004 | 1.000–1.000 | |
| CK (IU/L) | PPA | 0.528 | 288.980 | 66.7% | 50.0% | 0.873 | 0.168–0.888 |
| FLX | 1.000 | 316.285 | 100.0% | 100.0% | 0.004 | 1.000–1.000 | |
| IF g (pg/100 mg) | PPA | 1.000 | 100.940 | 100.0% | 100.0% | 0.004 | 1.000–1.000 |
| FLX | 0.972 | 90.300 | 100.0% | 83.3% | 0.006 | 0.889–1.056 | |
| Norepinephrine (ng/100 mg) | PPA | 1.000 | 4.215 | 100.0% | 100.0% | 0.004 | 1.000–1.000 |
| FLX | 0.694 | 5.030 | 83.3% | 66.7% | 0.262 | 0.377–1.012 | |
| Serotonin (ng/100 mg) | PPA | 1.000 | 5.185 | 100.0% | 100.0% | 0.004 | 1.000–1.000 |
| FLX | 1.000 | 5.485 | 100.0% | 100.0% | 0.004 | 1.000–1.000 | |
| Dopamine (ng/100 mg) | PPA | 1.000 | 18.485 | 100.0% | 100.0% | 0.004 | 1.000–1.000 |
| FLX | 1.000 | 19.030 | 100.0% | 100.0% | 0.004 | 1.000–1.000 | |
| Caspase-3 (pg/100 mg) | PPA | 1.000 | 126.040 | 100.0% | 100.0% | 0.004 | 1.000–1.000 |
| FLX | 1.000 | 123.085 | 100.0% | 100.0% | 0.004 | 1.000–1.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhat, R.S.; Alonazi, M.; Al-Daihan, S.; El-Ansary, A. Prenatal SSRI Exposure Increases the Risk of Autism in Rodents via Aggravated Oxidative Stress and Neurochemical Changes in the Brain. Metabolites 2023, 13, 310. https://doi.org/10.3390/metabo13020310
Bhat RS, Alonazi M, Al-Daihan S, El-Ansary A. Prenatal SSRI Exposure Increases the Risk of Autism in Rodents via Aggravated Oxidative Stress and Neurochemical Changes in the Brain. Metabolites. 2023; 13(2):310. https://doi.org/10.3390/metabo13020310
Chicago/Turabian StyleBhat, Ramesa Shafi, Mona Alonazi, Sooad Al-Daihan, and Afaf El-Ansary. 2023. "Prenatal SSRI Exposure Increases the Risk of Autism in Rodents via Aggravated Oxidative Stress and Neurochemical Changes in the Brain" Metabolites 13, no. 2: 310. https://doi.org/10.3390/metabo13020310
APA StyleBhat, R. S., Alonazi, M., Al-Daihan, S., & El-Ansary, A. (2023). Prenatal SSRI Exposure Increases the Risk of Autism in Rodents via Aggravated Oxidative Stress and Neurochemical Changes in the Brain. Metabolites, 13(2), 310. https://doi.org/10.3390/metabo13020310