
Activation of Granulocytes in Response to a High Protein Diet Leads to the Formation of Necrotic Lesions in the Liver


Abstract
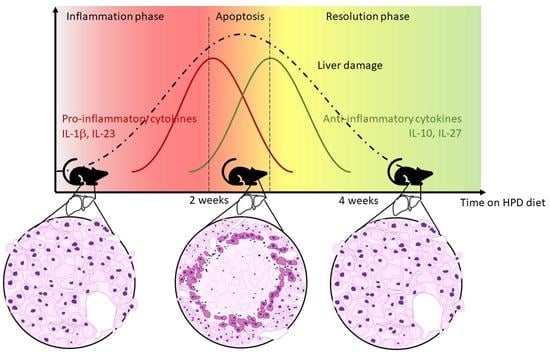
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Diets and Feeding
2.3. In Vivo Experiments
2.4. Cell Isolation and Flow Cytometry
2.5. Histology
2.6. Cytokine Quantification
3. Results
3.1. Short-Term Extreme Diets in Mice Do Not Cause Changes in Overall Morphological Parameters
3.2. Short-Term High-Protein Diet Feeding Causes Necrotic Liver Lesions Associated with Immune Cell Infiltrations
3.3. High Protein Diet Causes a Relative Increase of Pro-Inflammatory Cells of the Myeloid Lineage
3.4. Short-Term High-Protein Diet Is Associated with an Anti-Inflammatory Cytokine Profile
3.5. Neutrophils Cause Necrotic Liver Lesions in Response to Short-Term High-Protein Diet
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Diet, nutrition and the prevention of chronic diseases. World Health Organ Technol. Rep. Ser. 2003, 916, i–viii, 1–149. [Google Scholar]
- Stevens, G.A.; Singh, G.M.; Lu, Y.; Danaei, G.; Lin, J.K.; Finucane, M.M.; Bahalim, A.N.; McIntire, R.K.; Gutierrez, H.R.; Cowan, M.; et al. National, regional, and global trends in adult overweight and obesity prevalences. Popul. Health Metr. 2012, 10, 22. [Google Scholar] [CrossRef]
- Rakhra, V.; Galappaththy, S.L.; Bulchandani, S.; Cabandugama, P.K. Obesity and the Western Diet: How We Got Here. Mo. Med. 2020, 117, 536–538. [Google Scholar] [PubMed]
- O’Neill, B.; Raggi, P. The ketogenic diet: Pros and cons. Atherosclerosis 2020, 292, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Pesta, D.H.; Samuel, V.T. A high-protein diet for reducing body fat: Mechanisms and possible caveats. Nutr. Metab. 2014, 11, 53. [Google Scholar] [CrossRef]
- Freire, R. Scientific evidence of diets for weight loss: Different macronutrient composition, intermittent fasting, and popular diets. Nutrition 2020, 69, 110549. [Google Scholar] [CrossRef]
- Goldberg, E.L.; Shchukina, I.; Asher, J.L.; Sidorov, S.; Artyomov, M.N.; Dixit, V.D. Ketogenesis activates metabolically protective gammadelta T cells in visceral adipose tissue. Nat. Metab. 2020, 2, 50–61. [Google Scholar] [CrossRef]
- Mbikay, M.; Mayne, J.; Sirois, F.; Fedoryak, O.; Ray\mond, A.; Noad, J.; Chretien, M. Mice Fed a High-Cholesterol Diet Supplemented with Quercetin-3-Glucoside Show Attenuated Hyperlipidemia and Hyperinsulinemia Associated with Differential Regulation of PCSK9 and LDLR in their Liver and Pancreas. Mol. Nutr. Food Res. 2018, 62, e1700729. [Google Scholar] [CrossRef]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Wensveen, F.M.; Valentic, S.; Sestan, M.; Turk Wensveen, T.; Polic, B. The “Big Bang” in obese fat: Events initiating obesity-induced adipose tissue inflammation. Eur. J. Immunol. 2015, 45, 2446–2456. [Google Scholar] [CrossRef]
- Ellenbroek, J.H.; van Dijck, L.; Tons, H.A.; Rabelink, T.J.; Carlotti, F.; Ballieux, B.E.; de Koning, E.J. Long-term ketogenic diet causes glucose intolerance and reduced beta- and alpha-cell mass but no weight loss in mice. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E552–E558. [Google Scholar] [CrossRef]
- Roberts, M.N.; Wallace, M.A.; Tomilov, A.A.; Zhou, Z.; Marcotte, G.R.; Tran, D.; Perez, G.; Gutierrez-Casado, E.; Koike, S.; Knotts, T.A.; et al. A Ketogenic Diet Extends Longevity and Healthspan in Adult Mice. Cell Metab. 2017, 26, 539–546.e535. [Google Scholar] [CrossRef] [PubMed]
- Sumiyoshi, M.; Sakanaka, M.; Kimura, Y. Chronic intake of high-fat and high-sucrose diets differentially affects glucose intolerance in mice. J. Nutr. 2006, 136, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, R.; Seino, Y.; Ogata, H.; Murase, M.; Iida, A.; Hosokawa, K.; Joo, E.; Harada, N.; Tsunekawa, S.; Hamada, Y.; et al. Chronic high-sucrose diet increases fibroblast growth factor 21 production and energy expenditure in mice. J. Nutr. Biochem. 2017, 49, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Riviere, S.; Soubeyre, V.; Jarriault, D.; Molinas, A.; Leger-Charnay, E.; Desmoulins, L.; Grebert, D.; Meunier, N.; Grosmaitre, X. High Fructose Diet inducing diabetes rapidly impacts olfactory epithelium and behavior in mice. Sci. Rep. 2016, 6, 34011. [Google Scholar] [CrossRef]
- Fan, A.; Oladiran, O.; Shi, X.Q.; Zhang, J. High-salt diet decreases mechanical thresholds in mice that is mediated by a CCR2-dependent mechanism. J. Neuroinflamm. 2020, 17, 179. [Google Scholar] [CrossRef]
- He, W.; Xu, J.; Mu, R.; Li, Q.; Lv, D.L.; Huang, Z.; Zhang, J.; Wang, C.; Dong, L. High-salt diet inhibits tumour growth in mice via regulating myeloid-derived suppressor cell differentiation. Nat. Commun. 2020, 11, 1732. [Google Scholar] [CrossRef]
- Jones, J.G. Hepatic glucose and lipid metabolism. Diabetologia 2016, 59, 1098–1103. [Google Scholar] [CrossRef] [PubMed]
- Paulusma, C.C.; Lamers, W.H.; Broer, S.; van de Graaf, S.F.J. Amino acid metabolism, transport and signalling in the liver revisited. Biochem. Pharmacol. 2022, 201, 115074. [Google Scholar] [CrossRef]
- Racanelli, V.; Rehermann, B. The liver as an immunological organ. Hepatology 2006, 43, S54–S62. [Google Scholar] [CrossRef]
- Bellentani, S. The epidemiology of non-alcoholic fatty liver disease. Liver Int. 2017, 37 (Suppl. S1), 81–84. [Google Scholar] [CrossRef]
- Jimenez, J.V.; Carrillo-Perez, D.L.; Rosado-Canto, R.; Garcia-Juarez, I.; Torre, A.; Kershenobich, D.; Carrillo-Maravilla, E. Electrolyte and Acid-Base Disturbances in End-Stage Liver Disease: A Physiopathological Approach. Dig. Dis. Sci. 2017, 62, 1855–1871. [Google Scholar] [CrossRef] [PubMed]
- Charlton, M.; Krishnan, A.; Viker, K.; Sanderson, S.; Cazanave, S.; McConico, A.; Masuoko, H.; Gores, G. Fast food diet mouse: Novel small animal model of NASH with ballooning, progressive fibrosis, and high physiological fidelity to the human condition. Am. J. Physiol. Gastrointest Liver Physiol. 2011, 301, G825–G834. [Google Scholar] [CrossRef] [PubMed]
- Basaranoglu, M.; Neuschwander-Tetri, B.A. Nonalcoholic Fatty Liver Disease: Clinical Features and Pathogenesis. Gastroenterol. Hepatol. 2006, 2, 282–291. [Google Scholar]
- Yang, P.; Wang, Y.; Tang, W.; Sun, W.; Ma, Y.; Lin, S.; Jing, J.; Jiang, L.; Shi, H.; Song, Z.; et al. Western diet induces severe nonalcoholic steatohepatitis, ductular reaction, and hepatic fibrosis in liver CGI-58 knockout mice. Sci. Rep. 2020, 10, 4701. [Google Scholar] [CrossRef] [PubMed]
- McGill, M.R. The past and present of serum aminotransferases and the future of liver injury biomarkers. EXCLI J. 2016, 15, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Sitia, G.; Isogawa, M.; Kakimi, K.; Wieland, S.F.; Chisari, F.V.; Guidotti, L.G. Depletion of neutrophils blocks the recruitment of antigen-nonspecific cells into the liver without affecting the antiviral activity of hepatitis B virus-specific cytotoxic T lymphocytes. Proc. Natl. Acad. Sci. USA 2002, 99, 13717–13722. [Google Scholar] [CrossRef]
- Scapini, P.; Lapinet-Vera, J.A.; Gasperini, S.; Calzetti, F.; Bazzoni, F.; Cassatella, M.A. The neutrophil as a cellular source of chemokines. Immunol. Rev. 2000, 177, 195–203. [Google Scholar] [CrossRef]
- Karlmark, K.R.; Weiskirchen, R.; Zimmermann, H.W.; Gassler, N.; Ginhoux, F.; Weber, C.; Merad, M.; Luedde, T.; Trautwein, C.; Tacke, F. Hepatic recruitment of the inflammatory Gr1+ monocyte subset upon liver injury promotes hepatic fibrosis. Hepatology 2009, 50, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Batten, M.; Kljavin, N.M.; Li, J.; Walter, M.J.; de Sauvage, F.J.; Ghilardi, N. Cutting edge: IL-27 is a potent inducer of IL-10 but not FoxP3 in murine T cells. J. Immunol. 2008, 180, 2752–2756. [Google Scholar] [CrossRef]
- Kamari, Y.; Shaish, A.; Vax, E.; Shemesh, S.; Kandel-Kfir, M.; Arbel, Y.; Olteanu, S.; Barshack, I.; Dotan, S.; Voronov, E.; et al. Lack of interleukin-1alpha or interleukin-1beta inhibits transformation of steatosis to steatohepatitis and liver fibrosis in hypercholesterolemic mice. J. Hepatol. 2011, 55, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.C.; Harper, A.E. Adaptation of rats to diets containing different levels of protein: Effects on food intake, plasma and brain amino acid concentrations and brain neurotransmitter metabolism. J. Nutr. 1985, 115, 382–398. [Google Scholar] [CrossRef] [PubMed]
- Pichon, L.; Huneau, J.F.; Fromentin, G.; Tome, D. A high-protein, high-fat, carbohydrate-free diet reduces energy intake, hepatic lipogenesis, and adiposity in rats. J. Nutr. 2006, 136, 1256–1260. [Google Scholar] [CrossRef] [PubMed]
- Garcia Caraballo, S.C.; Comhair, T.M.; Dejong, C.H.; Lamers, W.H.; Kohler, S.E. A high-protein diet is anti-steatotic and has no pro-inflammatory side effects in dyslipidaemic APOE2 knock-in mice. Br. J. Nutr. 2014, 112, 1251–1265. [Google Scholar] [CrossRef] [PubMed]
- Kiilerich, P.; Myrmel, L.S.; Fjaere, E.; Hao, Q.; Hugenholtz, F.; Sonne, S.B.; Derrien, M.; Pedersen, L.M.; Petersen, R.K.; Mortensen, A.; et al. Effect of a long-term high-protein diet on survival, obesity development, and gut microbiota in mice. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E886–E899. [Google Scholar] [CrossRef]
- Vu, J.P.; Luong, L.; Parsons, W.F.; Oh, S.; Sanford, D.; Gabalski, A.; Lighton, J.R.; Pisegna, J.R.; Germano, P.M. Long-Term Intake of a High-Protein Diet Affects Body Phenotype, Metabolism, and Plasma Hormones in Mice. J. Nutr. 2017, 147, 2243–2251. [Google Scholar] [CrossRef]
- Cui, X.; Kim, E. Dual Effects of High Protein Diet on Mouse Skin and Colonic Inflammation. Clin. Nutr. Res. 2018, 7, 56–68. [Google Scholar] [CrossRef]
- Snelson, M.; Clarke, R.E.; Nguyen, T.V.; Penfold, S.A.; Forbes, J.M.; Tan, S.M.; Coughlan, M.T. Long Term High Protein Diet Feeding Alters the Microbiome and Increases Intestinal Permeability, Systemic Inflammation and Kidney Injury in Mice. Mol. Nutr. Food Res. 2021, 65, e2000851. [Google Scholar] [CrossRef]
- Lang, S.; Martin, A.; Farowski, F.; Wisplinghoff, H.; Vehreschild, M.; Liu, J.; Krawczyk, M.; Nowag, A.; Kretzschmar, A.; Herweg, J.; et al. High Protein Intake Is Associated With Histological Disease Activity in Patients With NAFLD. Hepatol. Commun. 2020, 4, 681–695. [Google Scholar] [CrossRef]
- Diaz-Rua, R.; Keijer, J.; Palou, A.; van Schothorst, E.M.; Oliver, P. Long-term intake of a high-protein diet increases liver triacylglycerol deposition pathways and hepatic signs of injury in rats. J. Nutr. Biochem. 2017, 46, 39–48. [Google Scholar] [CrossRef]
- Hou, Y.; Hu, S.; Li, X.; He, W.; Wu, G. Amino Acid Metabolism in the Liver: Nutritional and Physiological Significance. Adv. Exp. Med. Biol. 2020, 1265, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Moundras, C.; Remesy, C.; Demigne, C. Dietary protein paradox: Decrease of amino acid availability induced by high-protein diets. Am. J. Physiol. 1993, 264, G1057–G1065. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.W.D.; Bradshaw, P.C. Effects of a high protein diet and liver disease in an in silico model of human ammonia metabolism. Theor. Biol. Med. Model. 2019, 16, 11. [Google Scholar] [CrossRef]
- Bigot, A.; Tchan, M.C.; Thoreau, B.; Blasco, H.; Maillot, F. Liver involvement in urea cycle disorders: A review of the literature. J. Inherit. Metab. Dis. 2017, 40, 757–769. [Google Scholar] [CrossRef]
- De Chiara, F.; Heeboll, S.; Marrone, G.; Montoliu, C.; Hamilton-Dutoit, S.; Ferrandez, A.; Andreola, F.; Rombouts, K.; Gronbaek, H.; Felipo, V.; et al. Urea cycle dysregulation in non-alcoholic fatty liver disease. J. Hepatol. 2018, 69, 905–915. [Google Scholar] [CrossRef]
- Jalan, R.; De Chiara, F.; Balasubramaniyan, V.; Andreola, F.; Khetan, V.; Malago, M.; Pinzani, M.; Mookerjee, R.P.; Rombouts, K. Ammonia produces pathological changes in human hepatic stellate cells and is a target for therapy of portal hypertension. J. Hepatol. 2016, 64, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Nagamani, S.C.S.; Ali, S.; Izem, R.; Schady, D.; Masand, P.; Shneider, B.L.; Leung, D.H.; Burrage, L.C. Biomarkers for liver disease in urea cycle disorders. Mol. Genet. Metab. 2021, 133, 148–156. [Google Scholar] [CrossRef]
- Klune, J.R.; Bartels, C.; Luo, J.; Yokota, S.; Du, Q.; Geller, D.A. IL-23 mediates murine liver transplantation ischemia-reperfusion injury via IFN-gamma/IRF-1 pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G991–G1002. [Google Scholar] [CrossRef]
- Zang, M.; Li, Y.; He, H.; Ding, H.; Chen, K.; Du, J.; Chen, T.; Wu, Z.; Liu, H.; Wang, D.; et al. IL-23 production of liver inflammatory macrophages to damaged hepatocytes promotes hepatocellular carcinoma development after chronic hepatitis B virus infection. Biochim. Biophys Acta Mol. Basis Dis. 2018, 1864, 3759–3770. [Google Scholar] [CrossRef]
- Schagen, F.H.; Wensveen, F.M.; Carette, J.E.; Dermody, T.S.; Gerritsen, W.R.; van Beusechem, V.W. Genetic targeting of adenovirus vectors using a reovirus sigma1-based attachment protein. Mol. Ther. 2006, 13, 997–1005. [Google Scholar] [CrossRef]
- Steen, E.H.; Wang, X.; Balaji, S.; Butte, M.J.; Bollyky, P.L.; Keswani, S.G. The Role of the Anti-Inflammatory Cytokine Interleukin-10 in Tissue Fibrosis. Adv. Wound Care 2020, 9, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Chen, M.H.; Guo, Q.L.; Chen, Z.X.; Chen, Q.D.; Wang, X.Z. Interleukin-10 induces senescence of activated hepatic stellate cells via STAT3-p53 pathway to attenuate liver fibrosis. Cell Signal 2020, 66, 109445. [Google Scholar] [CrossRef] [PubMed]
- Matheson, R.; Deng, K.; Huai, G.; Lee, K.M.; Feeney, N.; Coe, T.M.; Cloonan, D.; Serifis, N.; Fu, Q.; Robson, S.C.; et al. Interleukin-27 in liver xenotransplantation: A rational target to mitigate ischemia reperfusion injury and increase xenograft survival. Trans. Rev. 2022, 36, 100674. [Google Scholar] [CrossRef] [PubMed]
- Guillot, A.; Gasmi, I.; Brouillet, A.; Ait-Ahmed, Y.; Calderaro, J.; Ruiz, I.; Gao, B.; Lotersztajn, S.; Pawlotsky, J.M.; Lafdil, F. Interleukins-17 and 27 promote liver regeneration by sequentially inducing progenitor cell expansion and differentiation. Hepatol. Commun. 2018, 2, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Hamberg, O.; Nielsen, K.; Vilstrup, H. Effects of an increase in protein intake on hepatic efficacy for urea synthesis in healthy subjects and in patients with cirrhosis. J. Hepatol. 1992, 14, 237–243. [Google Scholar] [CrossRef]
- Gallego-Duran, R.; Ampuero, J.; Pastor-Ramirez, H.; Alvarez-Amor, L.; Del Campo, J.A.; Maya-Miles, D.; Montero-Vallejo, R.; Cardenas-Garcia, A.; Pareja, M.J.; Gato-Zambrano, S.; et al. Liver injury in non-alcoholic fatty liver disease is associated with urea cycle enzyme dysregulation. Sci. Rep. 2022, 12, 3418. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Name | Diet Composition | (kcal/g) |
|---|---|---|
| Normal chow diet (NCD) | Mucedola 4RF21 diet 18.5% w/w crude protein 3% w/w crude fats and oils 6% w/w crude fibers 7% w/w crude ash 53% w/w NFE (carbs): −42.6% starch −3.7% sucrose | 3.13 |
| High fat diet (HFD) | 60% w/w NCD pellets 40% w/w fat (lard) | 11 |
| Steatotic diet (SSD) | 40% w/w NCD pellets 40% w/w fat (lard) 20% w/w fructose 2% w/w cholesterol | 11.3 |
| High cholesterol diet (HChD) | NCD pellets 2% w/w cholesterol | 3.13 |
| High fructose diet (HFrD) | 40% NCD pellets 60% fructose | 3.6 |
| High sodium diet (HSD) | NCD pellets 4% NaCl 1% NaCl in drinking water | 3.13 |
| High protein diet (HPD) | 50% w/w NCD pellets 50% w/w casein powder | 5.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benić, A.; Mikašinović, S.; Wensveen, F.M.; Polić, B. Activation of Granulocytes in Response to a High Protein Diet Leads to the Formation of Necrotic Lesions in the Liver. Metabolites 2023, 13, 153. https://doi.org/10.3390/metabo13020153
Benić A, Mikašinović S, Wensveen FM, Polić B. Activation of Granulocytes in Response to a High Protein Diet Leads to the Formation of Necrotic Lesions in the Liver. Metabolites. 2023; 13(2):153. https://doi.org/10.3390/metabo13020153
Chicago/Turabian StyleBenić, Ante, Sanja Mikašinović, Felix M. Wensveen, and Bojan Polić. 2023. "Activation of Granulocytes in Response to a High Protein Diet Leads to the Formation of Necrotic Lesions in the Liver" Metabolites 13, no. 2: 153. https://doi.org/10.3390/metabo13020153
APA StyleBenić, A., Mikašinović, S., Wensveen, F. M., & Polić, B. (2023). Activation of Granulocytes in Response to a High Protein Diet Leads to the Formation of Necrotic Lesions in the Liver. Metabolites, 13(2), 153. https://doi.org/10.3390/metabo13020153