


LC-MS and NMR Based Plant Metabolomics: A Comprehensive Phytochemical Investigation of Symphytum anatolicum

 , ,
, ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemical and Reagents
2.2. Plant Material
2.3. General Procedures
2.4. LC-ESI/LTQOrbitrap/MS Analysis
2.5. Extraction and Isolation Procedures
2.6. NMR Analysis and Data Processing
2.7. Total Phenolic Content, DDPH, and TEAC Assays
2.8. α-Glucosidase Inhibition Assay
2.9. Tyrosinase Inhibition Assay
2.10. Statistical Analysis of Data
3. Results
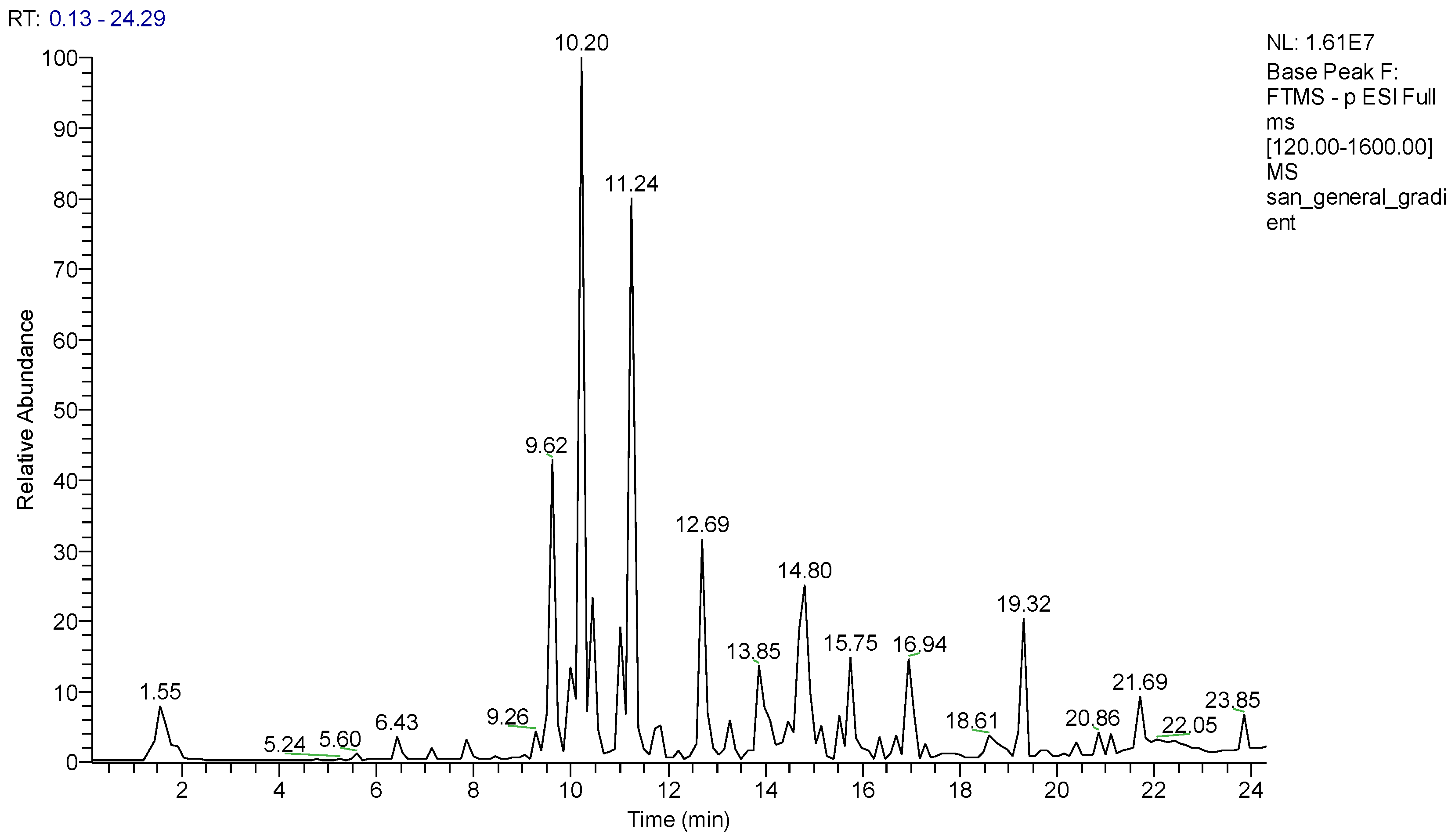
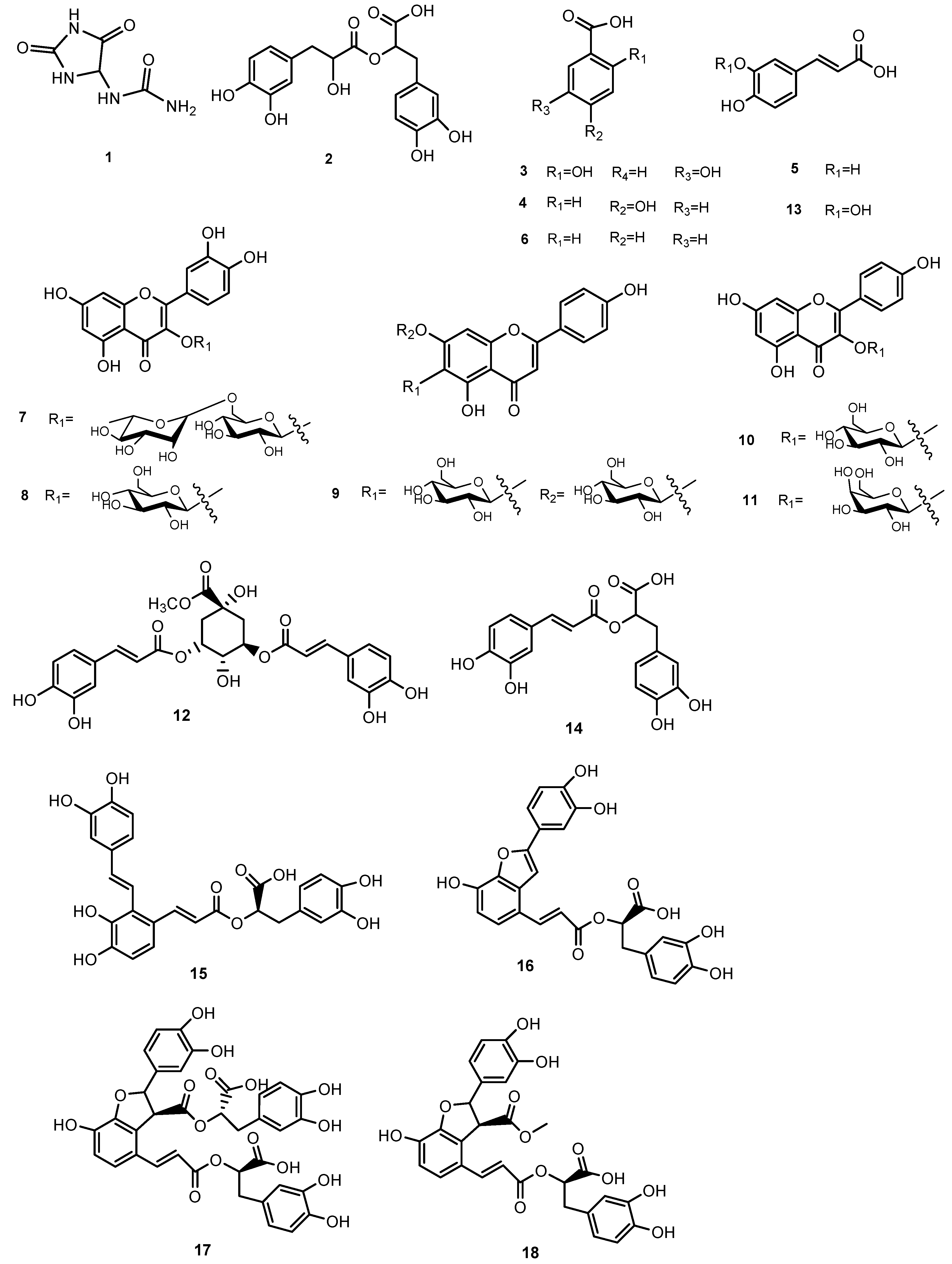
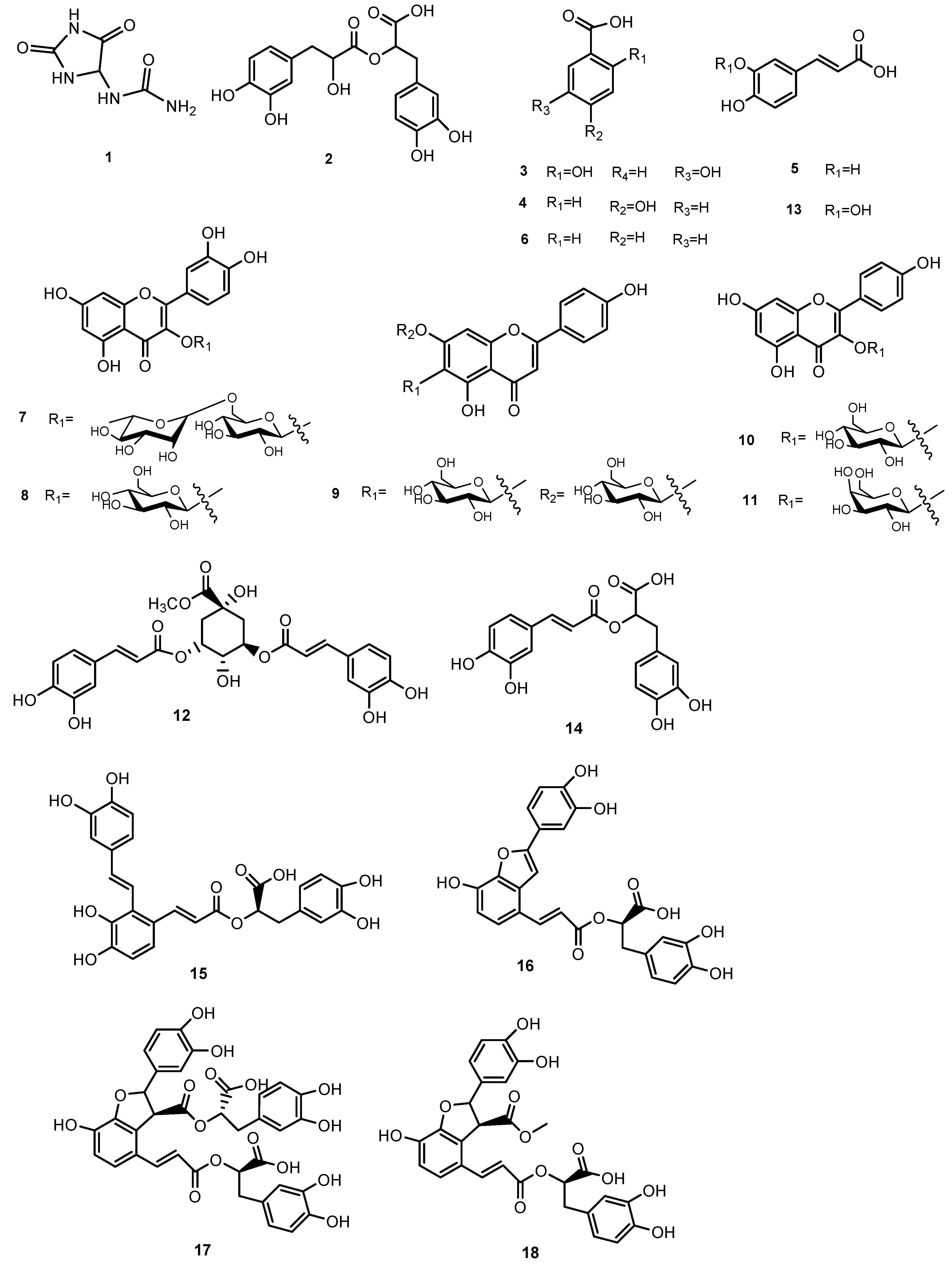
3.1. LCESI/Orbitrap/MS/MS Analysis of MeOH extract of S. anatolicum
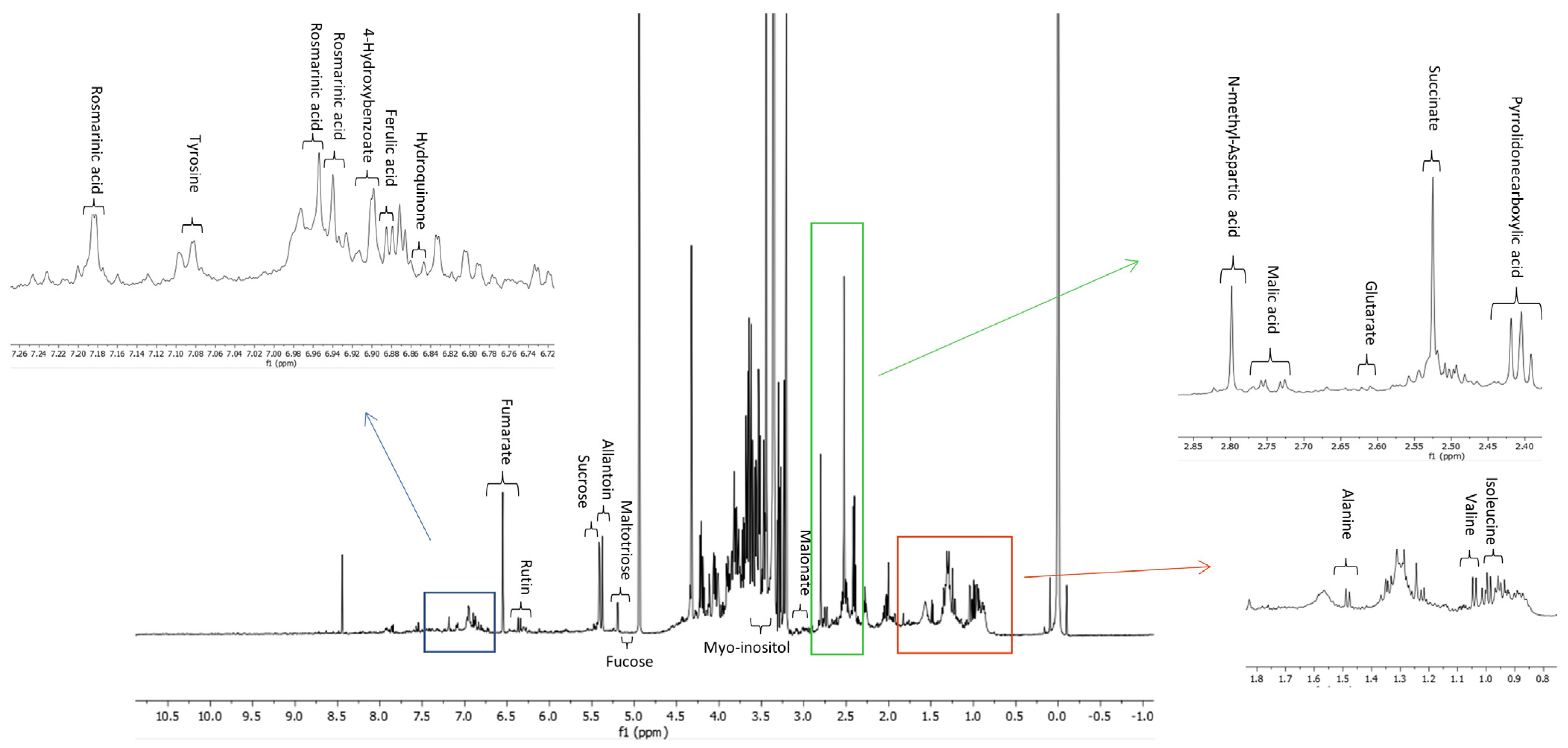
3.2. NMR Analysis of MeOH Extract of S. anatolicum
3.3. Evaluation of Phenolic Content and Antioxidant Activity
3.4. α-Glucosidase Inhibitory Activity
3.5. Tyrosinase Inhibitory Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Walker, T.W.N.; Alexander, J.M.; Allard, P.-M.; Baines, O.; Baldy, V.; Bardgett, R.D.; Capdevila, P.; Coley, P.D.; David, B.; Defossez, E.; et al. Functional Traits 2.0: The power of the metabolome for ecology. J. Ecol. 2022, 110, 4–20. [Google Scholar] [CrossRef]
- Ikhalaynen, Y.A.; Plyushchenko, I.V.; Rodin, I.A. Hopomics: Humulus lupulus Brewing Cultivars Classification Based on LC-MS Profiling and Nested Feature Selection. Metabolites 2022, 12, 945. [Google Scholar] [CrossRef]
- Ivanović, S.; Simić, K.; Lekić, S.; Jadranin, M.; Vujisić, L.; Gođevac, D. Plant Metabolomics as a Tool for Detecting Adulterants in Edible Plant: A Case Study of Allium ursinum. Metabolites 2022, 12, 849. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.P.; Wang, M.; Saroja, S.G.; Khan, I.A. NMR technique and methodology in botanical health product analysis and quality control. J. Pharm. Biomed. Anal. 2022, 207, 114376. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Sharopov, F.; Tumer, T.B.; Ozleyen, A.; Rodriguez-Perez, C.; Ezzat, S.M.; Azzini, E.; Hosseinabadi, T.; Butnariu, M.; Sarac, I.; et al. Symphytum Species: A Comprehensive Review on Chemical Composition, Food Applications and Phytopharmacology. Molecules 2019, 24, 2272. [Google Scholar] [CrossRef]
- Horinouchi, C.D.S.; Otuki, M.F. Botanical Briefs: Comfrey (Symphytum officinale). Cutis 2013, 91, 225–228. [Google Scholar]
- Trifan, A.; Zengin, G.; Sinan, K.I.; Esslinger, N.; Grubelnik, A.; Wolfram, E.; Skalicka-Woźniak, K.; Minceva, M.; Luca, S.V. Influence of the Post-Harvest Storage Time on the Multi-Biological Potential, Phenolic and Pyrrolizidine Alkaloid Content of Comfrey (Symphytum officinale L.) Roots Collected from Different European Regions. Plants 2021, 10, 1825. [Google Scholar] [CrossRef]
- Hacioglu, B.T.; Erik, S. Phylogeny of Symphytum L. (Boraginaceae) with special emphasis on Turkish species. Afr. J. Biotechnol. 2011, 10, 15483–15493. [Google Scholar] [CrossRef]
- Nastić, N.; Borrás-Linares, I.; Lozano-Sánchez, J.; Švarc-Gajić, J.; Segura-Carretero, A. Comparative Assessment of Phytochemical Profiles of Comfrey (Symphytum officinale L.) Root Extracts Obtained by Different Extraction Techniques. Molecules 2020, 25, 837. [Google Scholar] [CrossRef] [PubMed]
- Rode, D. Comfrey toxicity revisited. Trends Pharmacol. Sci. 2002, 23, 497–499. [Google Scholar] [CrossRef]
- Seigner, J.; Junker-Samek, M.; Plaza, A.; D’Urso, G.; Masullo, M.; Piacente, S.; Holper-Schichl, Y.M.; de Martin, R. A Symphytum officinale Root Extract Exerts Anti-inflammatory Properties by Affecting Two Distinct Steps of NF-kappa B Signaling. Front. Pharmacol. 2019, 10, 289. [Google Scholar] [CrossRef]
- Sharma, A.; Khanna, S.; Kaur, G.; Singh, I. Medicinal plants and their components for wound healing applications. Future J. Pharm. Sci. 2021, 7, 53. [Google Scholar] [CrossRef]
- Staiger, C. Comfrey: A Clinical Overview. Phytother. Res. 2012, 26, 1441–1448. [Google Scholar] [CrossRef] [PubMed]
- Sarikurkcu, C.; Ozer, M.S.; Tlili, N. LC–ESI–MS/MS characterization of phytochemical and enzyme inhibitory effects of different solvent extract of Symphytum anatolicum. Ind. Crop. Prod. 2019, 140, 111666. [Google Scholar] [CrossRef]
- Varvouni, E.-F.; Zengin, G.; Graikou, K.; Ganos, C.; Mroczek, T.; Chinou, I. Phytochemical analysis and biological evaluation of the aerial parts from Symphytum anatolicum Boiss. and Cynoglottis barrelieri (All.) Vural & Kit Tan (Boraginaceae). Biochem. Syst. Ecol. 2020, 92, 104128. [Google Scholar]
- Neagu, E.; Paun, G.; Albu, C.; Eremia, S.A.-M.V.; Radu, G.L. Artemisia abrotanum and Symphytum officinale Polyphenolic Compounds-Rich Extracts with Potential Application in Diabetes Management. Metabolites 2023, 13, 354. [Google Scholar] [CrossRef]
- Cicero, N.; Corsaro, C.; Salvo, A.; Vasi, S.; Giofre, S.V.; Ferrantelli, V.; Di Stefano, V.; Mallamace, D.; Dugo, G. The metabolic profile of lemon juice by proton HR-MAS NMR: The case of the PGI Interdonato Lemon of Messina. Nat. Prod. Res. 2015, 29, 1894–1902. [Google Scholar] [CrossRef] [PubMed]
- Cerulli, A.; Masullo, M.; Montoro, P.; Hosek, J.; Pizza, C.; Piacente, S. Metabolite profiling of “green” extracts of Corylus avellana leaves by H-1 NMR spectroscopy and multivariate statistical analysis. J. Pharm. Biomed. Anal. 2018, 160, 168–178. [Google Scholar] [CrossRef]
- Cerulli, A.; Masullo, M.; Piacente, S. Metabolite Profiling of Helichrysum italicum Derived Food Supplements by H-1-NMR-Based Metabolomics. Molecules 2021, 26, 6619. [Google Scholar] [CrossRef]
- Kilinc, H.; Masullo, M.; Lauro, G.; D’Urso, G.; Alankus, O.; Bifulco, G.; Piacente, S. Scabiosa atropurpurea: A rich source of iridoids with-glucosidase inhibitory activity evaluated by in vitro and in silico studies. Phytochemistry 2023, 205, 113471. [Google Scholar] [CrossRef]
- Oh, K.E.; Shin, H.; Lee, M.K.; Park, B.; Lee, K.Y. Characterization and Optimization of the Tyrosinase Inhibitory Activity of Vitis amurensis Root Using LC-Q-TOF-MS Coupled with a Bioassay and Response Surface Methodology. Molecules 2021, 26, 446. [Google Scholar] [CrossRef] [PubMed]
- Trifan, A.; Opitz, S.E.W.; Josuran, R.; Grubelnik, A.; Esslinger, N.; Peter, S.; Bram, S.; Meier, N.; Wolfram, E. Is comfrey root more than toxic pyrrolizidine alkaloids? Salvianolic acids among antioxidant polyphenols in comfrey (Symphytum officinale L.) roots. Food Chem. Toxicol. 2018, 112, 178–187. [Google Scholar] [CrossRef]
- Wang, J.; Xu, J.; Gong, X.; Yang, M.; Zhang, C.; Li, M. Biosynthesis, Chemistry, and Pharmacology of Polyphenols from Chinese Salvia Species: A Review. Molecules 2019, 24, 155. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, A.; Cerulli, A.; Pizza, C.; Piacente, S. Multi-class polar lipid profiling in fresh and roasted hazelnut (Corylus avellana cultivar “Tonda di Giffoni”) by LC-ESI/LTQOrbitrap/MS/MS(n). Food Chem. 2018, 269, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Cerulli, A.; Masullo, M.; Pizza, C.; Piacente, S. Metabolite Profiling of “Green” Extracts of Cynara cardunculus subsp. scolymus, Cultivar “Carciofo di Paestum” PGI by H-1 NMR and HRMS-Based Metabolomics. Molecules 2022, 27, 3328. [Google Scholar] [CrossRef]
- Borim de Souza, A.J.; Ocampos, F.M.M.; Catoia Pulgrossi, R.; Dokkedal, A.L.; Colnago, L.A.; Cechin, I.; Saldanha, L.L. NMR-Based Metabolomics Reveals Effects of Water Stress in the Primary and Specialized Metabolisms of Bauhinia ungulata L. (Fabaceae). Metabolites 2023, 13, 381. [Google Scholar] [CrossRef]
- Abdul Hamid, N.A.; Mediani, A.; Maulidiani, M.; Abas, F.; Park, Y.S.; Leontowicz, H.; Leontowicz, M.; Namiesnik, J.; Gorinstein, S. Characterization of metabolites in different kiwifruit varieties by NMR and fluorescence spectroscopy. J. Pharm. Biomed. Anal. 2017, 138, 80–91. [Google Scholar] [CrossRef]
- Corol, D.I.; Harflett, C.; Beale, M.H.; Ward, J.L. An Efficient High Throughput Metabotyping Platform for Screening of Biomass Willows. Metabolites 2014, 4, 946–976. [Google Scholar] [CrossRef]
- Ohyama, T.; Isaka, M.; Saito, A.; Higuchi, K. Effects of Nodulation on Metabolite Concentrations in Xylem Sap and in the Organs of Soybean Plants Supplied with Different N Forms. Metabolites 2023, 13, 319. [Google Scholar] [CrossRef]
- Selamoglu, Z.; Dusgun, C.; Akgul, H.; Gulhan, M.F. In-Vitro Antioxidant Activities of the Ethanolic Extracts of Some Contained-Allantoin Plants. Iran. J. Pharm. Res. 2017, 16, 92–98. [Google Scholar]
- Dawood, M.F.A.; Tahjib-Ul-Arif, M.; Sohag, A.A.; Latef, A.A.H.A.; Ragaey, M.M. Mechanistic Insight of Allantoin in Protecting Tomato Plants against Ultraviolet C Stress. Plants 2021, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Bao, T.Q.; Li, Y.; Qu, C.; Zheng, Z.G.; Yang, H.; Li, P. Antidiabetic Effects and Mechanisms of Rosemary (Rosmarinus officinalis L.) and its Phenolic Components. Am. J. Chin. Med. 2020, 48, 1353–1368. [Google Scholar] [CrossRef]
- Chen, S.L.; Shang, H.M.; Yang, J.Y.; Li, R.; Wu, H.X. Effects of different extraction techniques on physicochemical properties and activities of polysaccharides from comfrey (Symphytum officinale L.) root. Ind. Crop. Prod. 2018, 121, 18–25. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef]
- Zaidi, K.U.; Ali, S.A.; Ali, A.; Naaz, I. Natural Tyrosinase Inhibitors: Role of Herbals in the Treatment of Hyperpigmentary Disorders. Mini Rev. Med. Chem. 2019, 19, 796–808. [Google Scholar] [CrossRef]
- Shenoy, A.; Madan, R. Post-Inflammatory Hyperpigmentation: A Review of Treatment Strategies. J. Drugs Dermatol. 2020, 19, 763–768. [Google Scholar] [CrossRef]
- Zhang, X.W.; Bian, G.L.; Kang, P.Y.; Cheng, X.J.; Yan, K.; Liu, Y.L.; Gao, Y.X.; Li, D.Q. Recent advance in the discovery of tyrosinase inhibitors from natural sources via separation methods. J. Enzyme Inhib. Med. Chem. 2021, 36, 2104–2117. [Google Scholar]
- Zhu, L.; Ma, S.; Li, K.; Xiong, P.; Qin, S.; Cai, W. Systematic Screening of Chemical Constituents in the Traditional Chinese Medicine Arnebiae Radix by UHPLC-Q-Exactive Orbitrap Mass Spectrometry. Molecules 2022, 27, 2631. [Google Scholar] [CrossRef]
- Thuong, P.T.; Kang, K.W.; Kim, J.K.; Seo, D.B.; Lee, S.J.; Kim, S.H.; Oh, W.K. Lithospermic acid derivatives from Lithospermum erythrorhizon increased expression of serine palmitoyltransferase in human HaCaT cells. Bioorg. Med. Chem. Lett. 2009, 19, 1815–1817. [Google Scholar] [CrossRef]
- Ryu, M.J.; Kim, M.; Ji, M.; Lee, C.; Yang, I.; Hong, S.B.; Chin, J.; Seo, E.K.; Paik, M.J.; Lim, K.M.; et al. Discrimination of Lycium chinense and L. barbarum Based on Metabolite Analysis and Hepatoprotective Activity. Molecules 2020, 25, 5835. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | Rt | [M-H]− | Molecular Formula | ppm | MS/MS | Identity |
|---|---|---|---|---|---|---|
| 1 | 1.54 | 157.0362 | C4H6O3N4 | −0.63 | 140.01/114.03/97.00 | Allantoin * |
| 2 | 1.86 | 377.0857 | C18H18O9 | −2.6 | 197.0471 | dan shen suan C * |
| 3 | 4.40 | 153.0184 | C7H6O4 | 1.9 | - | 2,5-dihydroxybenzoic acid |
| 4 | 5.60 | 137.0240 | C7H6O3 | 1.4 | - | 4-hydroxybenzoic acid |
| 5 | 7.14 | 179.0346 | C9H8O4 | 1.1 | - | caffeic acid |
| 6 | 7.26 | 121.0301 | C7H6O2 | 3.0 | - | benzoic acid |
| 7 | 9.26 | 609.1460 | C27H30O16 | 0.8 | 301.0356 | rutin |
| 8 | 9.62 | 463.0879 | C21H20O12 | 0.7 | 301.0341 | quercetin-3-O-glucoside |
| 9 | 9.98 | 593.1509 | C27H30O15 | 0.5 | 285.0405 | saponarin |
| 10 | 10.20 | 447.0930 | C21H20O11 | 0.67 | 285.0407 | kaempferol 3-O-glucoside |
| 11 | 10.43 | 447.1348 | C15H28O15 | −0.44 | 285.0407 | kaempferol 3-O-galactoside |
| 12 | 10.62 | 515.1190 | C25H24O12 | 0.19 | 353.0872 | 3,5-O-DCQA |
| 13 | 11.12 | 193.0505 | C10H10O4 | 2.5 | - | ferulic acid |
| 14 | 11.24 | 359.0770 | C18H16O8 | 1.11 | 161.0249/179.0359/197.0472 | rosmarinic acid |
| 15 | 11.85 | 493.1136 | C26H22O10 | 0.40 | 295.0613 | salvianolic acid A |
| 16 | 12.69 | 491.0980 | C26H20O10 | 0.40 | 311.0564 | salvianolic acid C |
| 17 | 12.80 | 717.1460 | C36H30O16 | 0.69 | 519.0928/339.0498/321.0403 | salvianolic acid B |
| 18 | 13.23 | 551.1200 | C28H24O12 | 2.8 | 519.0922/371.0768/353.0666/339.0506 | monomethyl lithospermate |
| 19 | 13.85 | 327.2180 | C18H32O5 | 2.7 | 309.2076/291.1965/211.1346/171.1030 | 9,12,13-TriHODE (10,15) * |
| 20 | 14.76 | 329.2333 | C18H34O5 | 1.51 | 171.1030/229.1445/293.2128/311.2228 | 9,12,13 triHOME (10) * |
| 21 | 15.15 | 327.2179 | C18H32O5 | 2.5 | 229.1445/211.1346/171.1030/291.1965 | Oxo-9,10DiHOME (11) * |
| mM | mg/g MeOH Extract | |
|---|---|---|
| Amino acids | ||
| Alanine | 0.0148 ± 0.0052 | 0.1293 ± 0.0454 |
| Isoleucine | 0.0254 ± 0.0085 | 0.3266 ± 0.1093 |
| Tyrosine | 0.1716 ± 0.0018 | 3.0483 ± 0.0320 |
| N-Methyl-aspartic acid | 0.2474 ± 0.0349 | 3.5686 ± 0.5034 |
| Valine | 0.0424 ± 0.0037 | 0.4870 ± 0.0425 |
| Sugars | ||
| Sucrose | 0.3021 ± 0.0028 | 10.1381 ± 0.0940 |
| Fucose | 0.5127 ± 0.0374 | 8.2515 ± 0.6019 |
| Maltotriose | 0.3855 ± 0.0293 | 19.0634 ± 1.4489 |
| Organic acids | ||
| Pyrrolidonecarboxylic acid | 0.3492 ± 0.0975 | 4.4201 ± 1.2341 |
| Succinic acid | 0.1020 ± 0.0154 | 1.1607 ± 0.1752 |
| 4-Hydroxybenzoic acid | 0.0129 ± 0.0052 | 0.1734 ± 0.0699 |
| Malonic acid | 0.3249 ± 0.0287 | 3.2506 ± 0.2871 |
| Glutaric acid | 0.0509 ± 0.0200 | 0.6543 ± 0.2571 |
| Fumaric acid | 0.0663 ± 0.0023 | 0.7414 ± 0.0257 |
| Malic acid | 0.0986 ± 0.0251 | 1.2962 ± 0.3300 |
| Phenolic compounds | ||
| Rutin | 0.1435 ± 0.0152 | 8.5889 ± 0.9098 |
| Rosmarinic acid | 0.0900 ± 0.0062 | 3.1791 ± 0.2190 |
| 3,4-dihydroxyhydrocinnamic acid | 0.0934 ± 0.0063 | 1.6482 ± 0.1112 |
| Others | ||
| Myo-inositol | 0.2131 ± 0.0999 | 3.7639 ± 1.7645 |
| Hydroquinone | 0.0160 ± 0.0021 | 0.1727 ± 0.0227 |
| Allantoin | 0.1595 ± 0.0237 | 2.4726 ± 0.3674 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kılınc, H.; D’Urso, G.; Paolillo, A.; Alankus, O.; Piacente, S.; Masullo, M. LC-MS and NMR Based Plant Metabolomics: A Comprehensive Phytochemical Investigation of Symphytum anatolicum. Metabolites 2023, 13, 1051. https://doi.org/10.3390/metabo13101051
Kılınc H, D’Urso G, Paolillo A, Alankus O, Piacente S, Masullo M. LC-MS and NMR Based Plant Metabolomics: A Comprehensive Phytochemical Investigation of Symphytum anatolicum. Metabolites. 2023; 13(10):1051. https://doi.org/10.3390/metabo13101051
Chicago/Turabian StyleKılınc, Hilal, Gilda D’Urso, Annunziata Paolillo, Ozgen Alankus, Sonia Piacente, and Milena Masullo. 2023. "LC-MS and NMR Based Plant Metabolomics: A Comprehensive Phytochemical Investigation of Symphytum anatolicum" Metabolites 13, no. 10: 1051. https://doi.org/10.3390/metabo13101051
APA StyleKılınc, H., D’Urso, G., Paolillo, A., Alankus, O., Piacente, S., & Masullo, M. (2023). LC-MS and NMR Based Plant Metabolomics: A Comprehensive Phytochemical Investigation of Symphytum anatolicum. Metabolites, 13(10), 1051. https://doi.org/10.3390/metabo13101051