
Upregulation of Nrf2/HO-1 Signaling and Attenuation of Oxidative Stress, Inflammation, and Cell Death Mediate the Protective Effect of Apigenin against Cyclophosphamide Hepatotoxicity

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Results
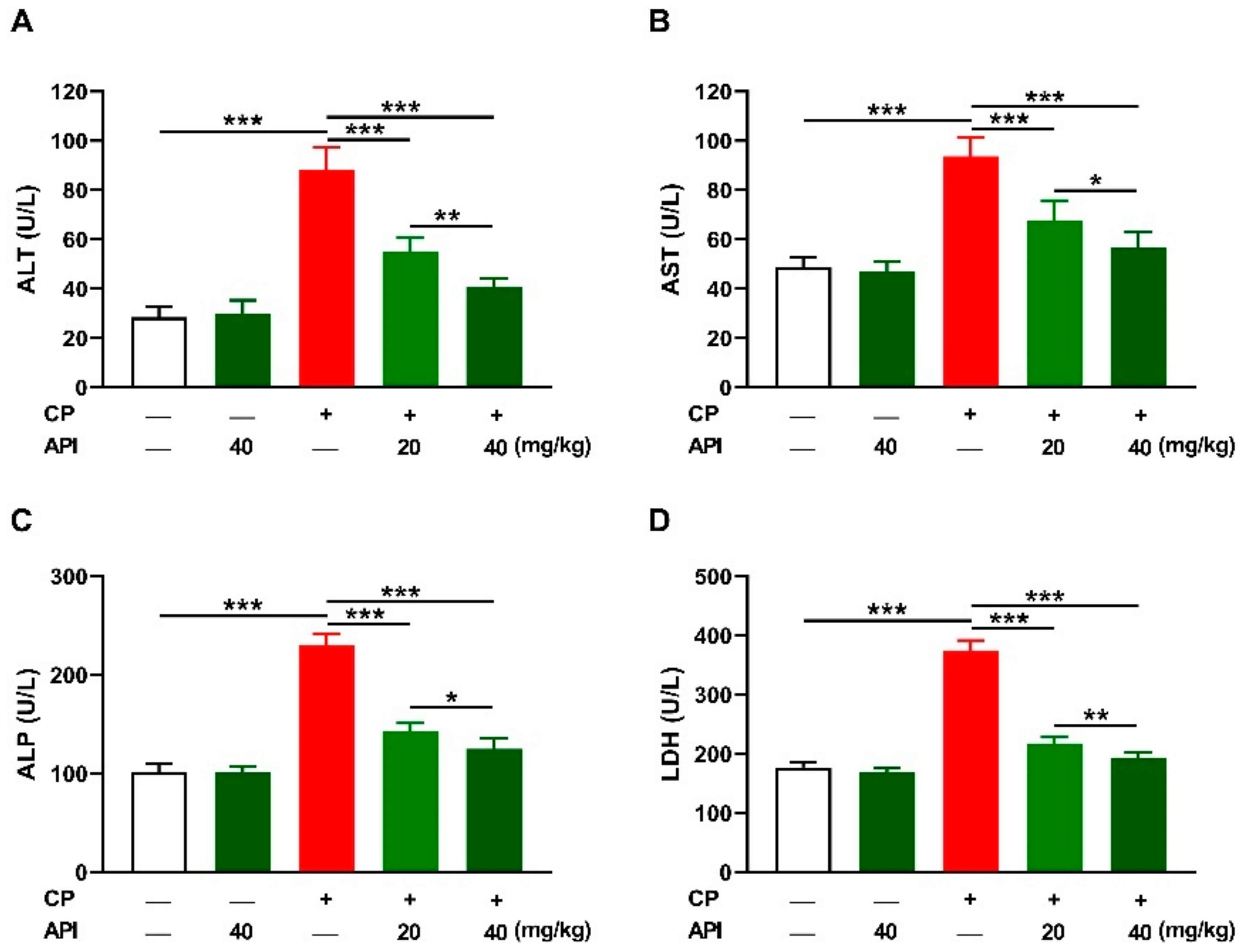
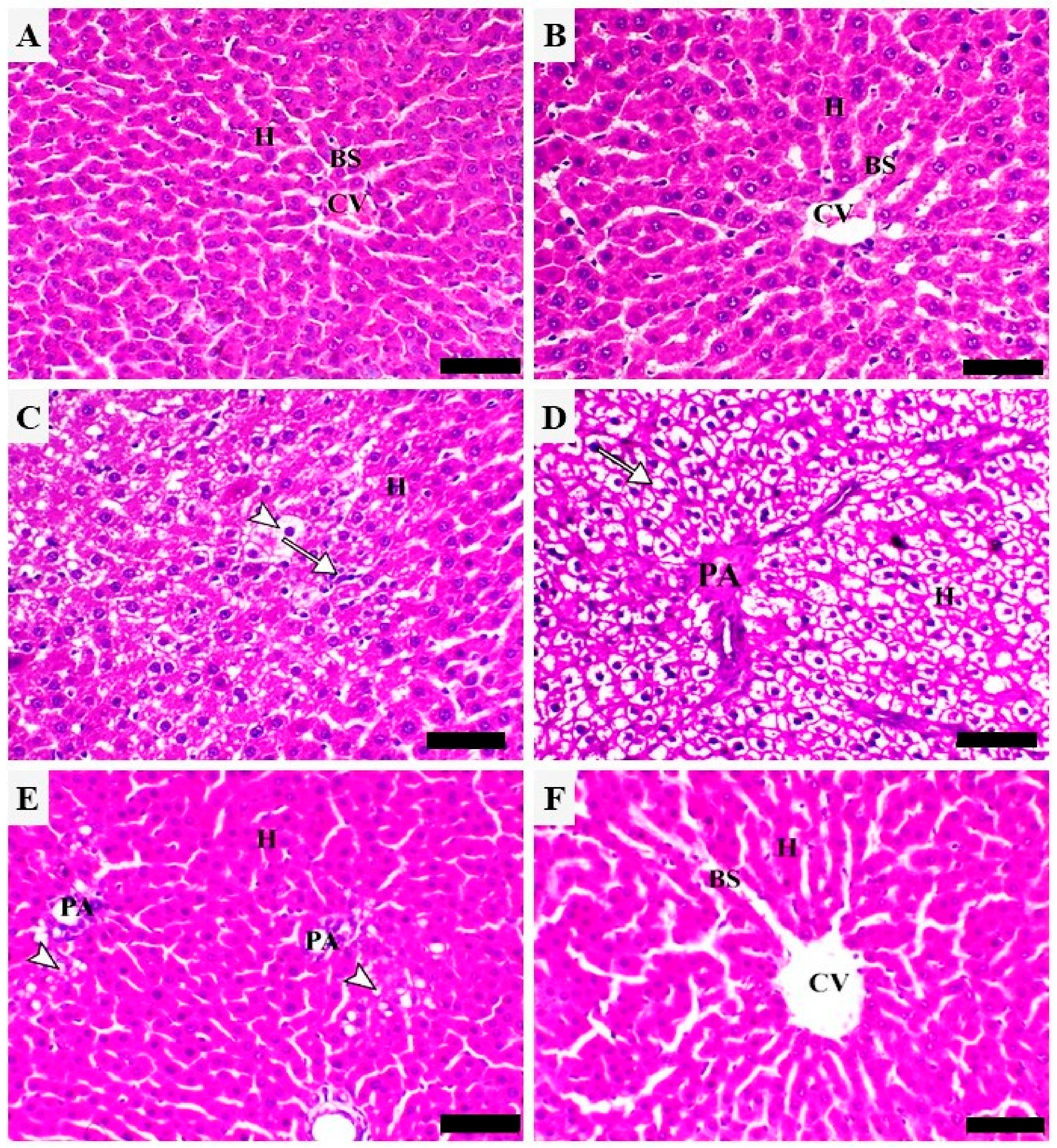
2.1. API Prevents CP-Induced Hepatocyte Injury in Rats
2.2. API Attenuates CP-Induced Oxidative Stress and Oxidative DNA Damage in Rats
2.3. API Activates Nrf2/HO-1 Signaling in CP-Intoxicated Rats
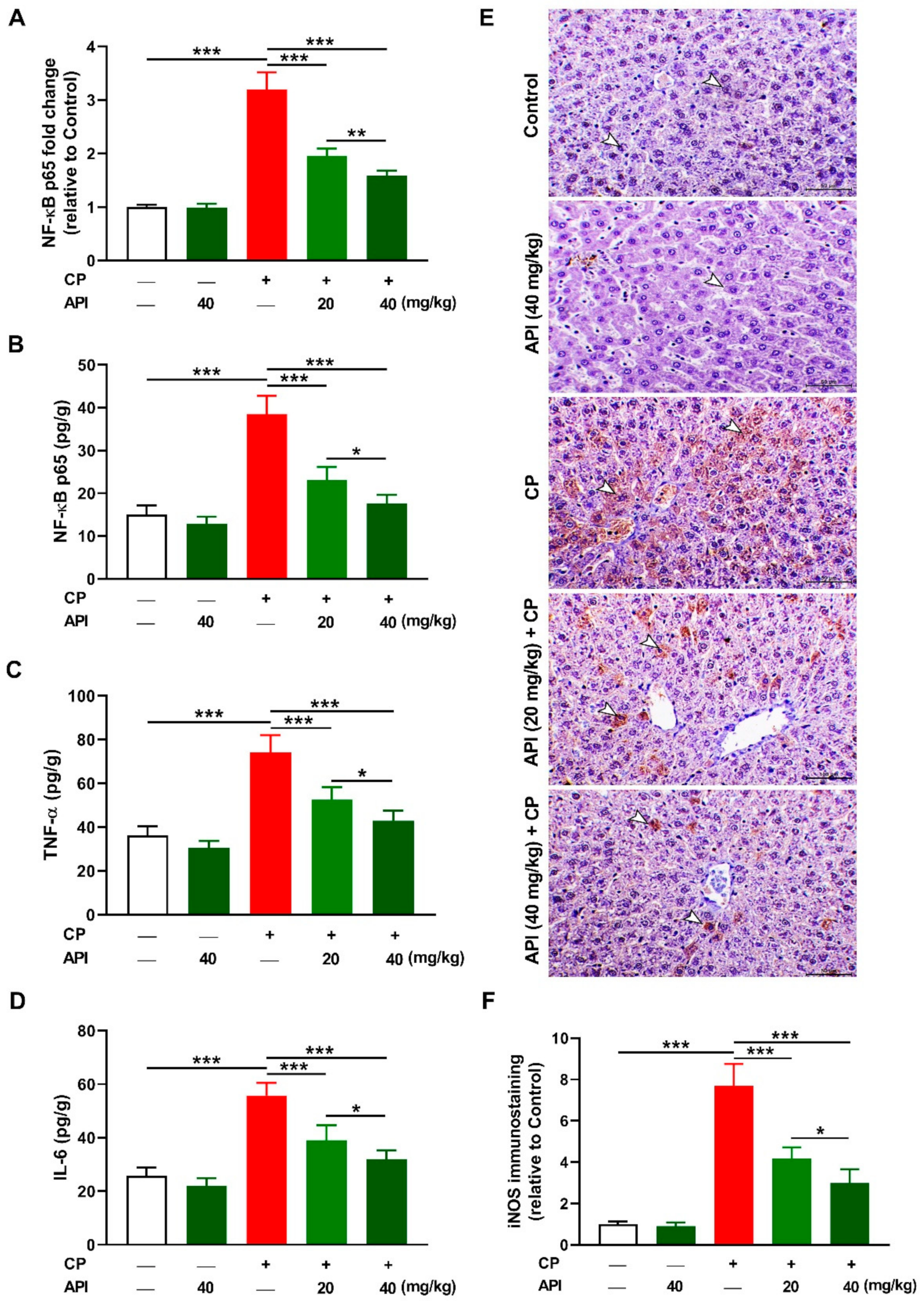
2.4. API Mitigates CP-Induced Hepatic Inflammation in Rats
2.5. API Prevents CP-Induced Hepatic Apoptosis in Rats
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Treatments
4.2. Determination of Liver Function Enzymes, Cytokines, and 8-Oxo-dG
4.3. Determination of ROS, LPO, NO, and Antioxidants
4.4. Histopathology and Immunohistochemistry (IHC) Examination
4.5. qRT-PCR
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Emadi, A.; Jones, R.J.; Brodsky, R.A. Cyclophosphamide and cancer: Golden anniversary. Nat. Rev. Clin. Oncol. 2009, 6, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Colvin, O.M. An overview of cyclophosphamide development and clinical applications. Curr. Pharm. Des. 1999, 5, 555–560. [Google Scholar] [PubMed]
- Moignet, A.; Hasanali, Z.; Zambello, R.; Pavan, L.; Bareau, B.; Tournilhac, O.; Roussel, M.; Fest, T.; Awwad, A.; Baab, K. Cyclophosphamide as a first-line therapy in lgl leukemia. Leukemia 2014, 28, 1134–1136. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.; Mortimer, J.; Livingston, R.B. High-dose cyclophosphamide in the treatment of refractory lymphomas and solid tumor malignancies. Cancer 1989, 63, 228–232. [Google Scholar] [CrossRef]
- Michot, J.M.; Annereau, M.; Danu, A.; Legoupil, C.; Bertin, L.; Chahine, C.; Achab, N.; Antosikova, A.; Cerutti, A.; Rossignol, J.; et al. High-dose cyclophosphamide for hard-to-treat patients with relapsed or refractory b-cell non-hodgkin’s lymphoma, a phase ii result. Eur. J. Haematol. 2020, 104, 281–290. [Google Scholar] [CrossRef]
- Abd El-Ghafar, O.A.M.; Hassanein, E.H.M.; Sayed, A.M.; Rashwan, E.K.; Shalkami, A.S.; Mahmoud, A.M. Acetovanillone prevents cyclophosphamide-induced acute lung injury by modulating pi3k/akt/mtor and nrf2 signaling in rats. Phytother. Res. PTR 2021, 35, 4499–4510. [Google Scholar] [CrossRef]
- Hassanein, E.H.M.; Abd El-Ghafar, O.A.M.; Ahmed, M.A.; Sayed, A.M.; Gad-Elrab, W.M.; Ajarem, J.S.; Allam, A.A.; Mahmoud, A.M. Edaravone and acetovanillone upregulate nrf2 and pi3k/akt/mtor signaling and prevent cyclophosphamide cardiotoxicity in rats. Drug Des. Dev. Ther. 2020, 14, 5275–5288. [Google Scholar] [CrossRef]
- Martin, F.; Lauwerys, B.; Lefèbvre, C.; Devogelaer, J.P.; Houssiau, F.A. Side-effects of intravenous cyclophosphamide pulse therapy. Lupus 1997, 6, 254–257. [Google Scholar] [CrossRef]
- Stillwell, T.J.; Benson, R.C., Jr. Cyclophosphamide-induced hemorrhagic cystitis. A review of 100 patients. Cancer 1988, 61, 451–457. [Google Scholar] [CrossRef]
- Segura, A.; Yuste, A.; Cercos, A.; López-Tendero, P.; Gironés, R.; Pérez-Fidalgo, J.A.; Herranz, C. Pulmonary fibrosis induced by cyclophosphamide. Ann. Pharmacother. 2001, 35, 894–897. [Google Scholar] [CrossRef]
- Dhesi, S.; Chu, M.P.; Blevins, G.; Paterson, I.; Larratt, L.; Oudit, G.Y.; Kim, D.H. Cyclophosphamide-induced cardiomyopathy: A case report, review, and recommendations for management. J. Investig. Med. High Impact Case Rep. 2013, 1, 2324709613480346. [Google Scholar] [CrossRef] [PubMed]
- Akay, H.; Akay, T.; Secilmis, S.; Kocak, Z.; Donderici, O. Hepatotoxicity after low-dose cyclophosphamide therapy. South. Med. J. 2006, 99, 1399–1401. [Google Scholar] [CrossRef]
- De Jonge, M.; Huitema, A.; Beijnen, J.; Rodenhuis, S. High exposures to bioactivated cyclophosphamide are related to the occurrence of veno-occlusive disease of the liver following high-dose chemotherapy. Br. J. Cancer 2006, 94, 1226–1230. [Google Scholar] [CrossRef][Green Version]
- Moghe, A.; Ghare, S.; Lamoreau, B.; Mohammad, M.; Barve, S.; McClain, C.; Joshi-Barve, S. Molecular mechanisms of acrolein toxicity: Relevance to human disease. Toxicol. Sci. 2015, 143, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Hamzeh, M.; Hosseinimehr, S.J.; Khalatbary, A.R.; Mohammadi, H.R.; Dashti, A.; Amiri, F.T. Atorvastatin mitigates cyclophosphamide-induced hepatotoxicity via suppression of oxidative stress and apoptosis in rat model. Res. Pharm. Sci. 2018, 13, 440. [Google Scholar] [PubMed]
- Zhu, H.; Long, M.-H.; Wu, J.; Wang, M.-M.; Li, X.-Y.; Shen, H.; Xu, J.-D.; Zhou, L.; Fang, Z.-J.; Luo, Y. Ginseng alleviates cyclophosphamide-induced hepatotoxicity via reversing disordered homeostasis of glutathione and bile acid. Sci. Rep. 2015, 5, 17536. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Germoush, M.O.; Alotaibi, M.F.; Hussein, O.E. Possible involvement of nrf2 and pparγ up-regulation in the protective effect of umbelliferone against cyclophosphamide-induced hepatotoxicity. Biomed. Pharmacother. 2017, 86, 297–306. [Google Scholar] [CrossRef]
- Mansour, D.F.; Saleh, D.O.; Mostafa, R.E. Genistein ameliorates cyclophosphamide-induced hepatotoxicity by modulation of oxidative stress and inflammatory mediators. Open Access Maced. J. Med. Sci. 2017, 5, 836. [Google Scholar] [CrossRef]
- Elgebaly, H.A.; Mosa, N.M.; Allach, M.; El-massry, K.F.; El-Ghorab, A.H.; Al Hroob, A.M.; Mahmoud, A.M. Olive oil and leaf extract prevent fluoxetine-induced hepatotoxicity by attenuating oxidative stress, inflammation and apoptosis. Biomed. Pharmacother. 2018, 98, 446–453. [Google Scholar] [CrossRef]
- Mohamed, H.M.; Mahmoud, A.M. Chronic exposure to the opioid tramadol induces oxidative damage, inflammation and apoptosis, and alters cerebral monoamine neurotransmitters in rats. Biomed. Pharmacother. 2019, 110, 239–247. [Google Scholar] [CrossRef]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.B.; Novellino, E. The therapeutic potential of apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.; Naz, F.; Jyoti, S.; Siddique, Y.H. Protective effect of apigenin against n-nitrosodiethylamine (ndea)-induced hepatotoxicity in albino rats. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2014, 767, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Chen, F.; Hu, X.; Yuan, Y. Protective effects of apigenin against furan-induced toxicity in mice. Food Funct. 2014, 5, 1804–1812. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, X.-Y.; Xue, J.; Gu, Z.-L.; Xie, M.-L. Protective effect of apigenin on mouse acute liver injury induced by acetaminophen is associated with increment of hepatic glutathione reductase activity. Food Funct. 2013, 4, 939–943. [Google Scholar] [CrossRef] [PubMed]
- Goudarzi, M.; Kalantar, M.; Sadeghi, E.; Karamallah, M.H.; Kalantar, H. Protective effects of apigenin on altered lipid peroxidation, inflammation, and antioxidant factors in methotrexate-induced hepatotoxicity. Naunyn Schmiedeberg’s Arch. Pharmacol. 2021, 394, 523–531. [Google Scholar] [CrossRef]
- Satta, S.; Mahmoud, A.M.; Wilkinson, F.L.; Yvonne Alexander, M.; White, S.J. The role of nrf2 in cardiovascular function and disease. Oxidative Med. Cell. Longev. 2017, 2017, 9237263. [Google Scholar] [CrossRef] [PubMed]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting molecular cross-talk between nrf2 and nf-κb response pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Wang, H.; Wang, X.; Zhu, L.; Mao, L. The absence of nrf2 enhances nf-kappab-dependent inflammation following scratch injury in mouse primary cultured astrocytes. Mediat. Inflamm. 2012, 2012, 217580. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Al Dera, H.S. 18β-glycyrrhetinic acid exerts protective effects against cyclophosphamide-induced hepatotoxicity: Potential role of pparγ and nrf2 upregulation. Genes Nutr. 2015, 10, 41. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Hozayen, W.G.; Ramadan, S.M. Berberine ameliorates methotrexate-induced liver injury by activating nrf2/ho-1 pathway and pparγ, and suppressing oxidative stress and apoptosis in rats. Biomed. Pharmacother. 2017, 94, 280–291. [Google Scholar] [CrossRef]
- Aladaileh, S.H.; Abukhalil, M.H.; Saghir, S.A.; Hanieh, H.; Alfwuaires, M.A.; Almaiman, A.A.; Bin-Jumah, M.; Mahmoud, A.M. Galangin activates nrf2 signaling and attenuates oxidative damage, inflammation, and apoptosis in a rat model of cyclophosphamide-induced hepatotoxicity. Biomolecules 2019, 9, 346. [Google Scholar] [CrossRef] [PubMed]
- Kamel, E.M.; Mahmoud, A.M.; Ahmed, S.A.; Lamsabhi, A.M. A phytochemical and computational study on flavonoids isolated from trifolium resupinatum l. And their novel hepatoprotective activity. Food Funct. 2016, 7, 2094–2106. [Google Scholar] [CrossRef] [PubMed]
- Vincenzi, B.; Armento, G.; Spalato Ceruso, M.; Catania, G.; Leakos, M.; Santini, D.; Minotti, G.; Tonini, G. Drug-induced hepatotoxicity in cancer patients-implication for treatment. Expert Opin. Drug Saf. 2016, 15, 1219–1238. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M. Hesperidin protects against cyclophosphamide-induced hepatotoxicity by upregulation of pparγ and abrogation of oxidative stress and inflammation. Can. J. Physiol. Pharmacol. 2014, 92, 717–724. [Google Scholar] [CrossRef]
- McGill, M.R. The past and present of serum aminotransferases and the future of liver injury biomarkers. EXCLI J. 2016, 15, 817–828. [Google Scholar]
- Germoush, M.O.; Mahmoud, A.M. Berberine mitigates cyclophosphamide-induced hepatotoxicity by modulating antioxidant status and inflammatory cytokines. J. Cancer Res. Clin. Oncol. 2014, 140, 1103–1109. [Google Scholar] [CrossRef]
- Smathers, R.L.; Galligan, J.J.; Stewart, B.J.; Petersen, D.R. Overview of lipid peroxidation products and hepatic protein modification in alcoholic liver disease. Chem. Biol. Interact. 2011, 192, 107–112. [Google Scholar] [CrossRef]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef]
- Messaadia, L.; Bekkar, Y.; Benamira, M.; Lahmar, H. Predicting the antioxidant activity of some flavonoids of arbutus plant: A theoretical approach. Chem. Phys. Impact 2020, 1, 100007. [Google Scholar] [CrossRef]
- Zheng, Y.-Z.; Chen, D.-F.; Deng, G.; Guo, R. The substituent effect on the radical scavenging activity of apigenin. Molecules 2018, 23, 1989. [Google Scholar] [CrossRef]
- Khadrawy, S.M.; Mohamed, H.M.; Mahmoud, A.M. Mesenchymal stem cells ameliorate oxidative stress, inflammation, and hepatic fibrosis via nrf2/ho-1 signaling pathway in rats. Environ. Sci. Pollut. Res. 2021, 28, 2019–2030. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Desouky, E.M.; Hozayen, W.G.; Bin-Jumah, M.; El-Nahass, E.S.; Soliman, H.A.; Farghali, A.A. Mesoporous silica nanoparticles trigger liver and kidney injury and fibrosis via altering tlr4/nf-κb, jak2/stat3 and nrf2/ho-1 signaling in rats. Biomolecules 2019, 9, 528. [Google Scholar] [CrossRef]
- Al-Amarat, W.; Abukhalil, M.H.; Althunibat, O.Y.; Alfwuaires, M.A.; Alnamshan, M.M.; Alqosaibi, A.I.; Ahmeda, A.F.; Kamel, E.M.; Arab, H.H.; Mahmoud, A.M. Galangin attenuates liver injury, oxidative stress and inflammation, and upregulates nrf2/ho-1 signaling in streptozotocin-induced diabetic rats. Processes 2021, 9, 1562. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Zaki, A.R.; Hassan, M.E.; Mostafa-Hedeab, G. Commiphora molmol resin attenuates diethylnitrosamine/phenobarbital-induced hepatocarcinogenesis by modulating oxidative stress, inflammation, angiogenesis and nrf2/are/ho-1 signaling. Chem. Biol. Interact. 2017, 270, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Gonzalez, X.; Fuentes, F.; Jeffery, S.; Saw, C.L.; Shu, L.; Su, Z.Y.; Kong, A.N. Induction of nrf2-mediated gene expression by dietary phytochemical flavones apigenin and luteolin. Biopharm. Drug Dispos. 2015, 36, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.-S.; Yen, J.-H.; Kou, M.-C.; Wu, M.-J. Luteolin and apigenin attenuate 4-hydroxy-2-nonenal-mediated cell death through modulation of upr, nrf2-are and mapk pathways in pc12 cells. PLoS ONE 2015, 10, e0130599. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, J.; Zhao, G.; Lin, M.; Lang, Y.; Zhang, D.; Feng, D.; Tu, C. Apigenin protects human melanocytes against oxidative damage by activation of the nrf2 pathway. Cell Stress Chaperones 2020, 25, 277–285. [Google Scholar] [CrossRef]
- Feng, X.; Yu, W.; Li, X.; Zhou, F.; Zhang, W.; Shen, Q.; Li, J.; Zhang, C.; Shen, P. Apigenin, a modulator of pparγ, attenuates hfd-induced nafld by regulating hepatocyte lipid metabolism and oxidative stress via nrf2 activation. Biochem. Pharmacol. 2017, 136, 136–149. [Google Scholar] [CrossRef]
- Luedde, T.; Schwabe, R.F. Nf-κb in the liver--linking injury, fibrosis and hepatocellular carcinoma. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 108–118. [Google Scholar] [CrossRef]
- Kolios, G.; Valatas, V.; Kouroumalis, E. Role of kupffer cells in the pathogenesis of liver disease. World J. Gastroenterol. 2006, 12, 7413–7420. [Google Scholar] [CrossRef]
- Alqahtani, S.; Mahmoud, A.M. Gamma-glutamylcysteine ethyl ester protects against cyclophosphamide-induced liver injury and hematologic alterations via upregulation of pparγ and attenuation of oxidative stress, inflammation, and apoptosis. Oxid. Med. Cell Longev. 2016, 2016, 4016209. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Chen, J.; Weng, C.; Chen, R.; Zheng, Y.; Chen, Q.; Tang, H. Identification of the protein-protein contact site and interaction mode of human vdac1 with bcl-2 family proteins. Biochem. Biophys. Res. Commun. 2003, 305, 989–996. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Et Biophys. Acta BBA Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef] [PubMed]
- Tsaroucha, A.K.; Tsiaousidou, A.; Ouzounidis, N.; Tsalkidou, E.; Lambropoulou, M.; Giakoustidis, D.; Chatzaki, E.; Simopoulos, C. Intraperitoneal administration of apigenin in liver ischemia/reperfusion injury protective effects. Saudi J. Gastroenterol. 2016, 22, 415–422. [Google Scholar] [PubMed]
- Du, H.; Hao, J.; Liu, F.; Lu, J.; Yang, X. Apigenin attenuates acute myocardial infarction of rats via the inhibitions of matrix metalloprotease-9 and inflammatory reactions. Int. J. Clin. Exp. Med. 2015, 8, 8854. [Google Scholar]
- Hozayen, W.G.; Mahmoud, A.M.; Desouky, E.M.; El-Nahass, E.S.; Soliman, H.A.; Farghali, A.A. Cardiac and pulmonary toxicity of mesoporous silica nanoparticles is associated with excessive ros production and redox imbalance in wistar rats. Biomed. Pharmacother. Biomed. Pharmacother. 2019, 109, 2527–2538. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Grisham, M.B.; Johnson, G.G.; Lancaster, J.R., Jr. Quantitation of nitrate and nitrite in extracellular fluids. Methods Enzymol. 1996, 268, 237–246. [Google Scholar]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Cohen, G.; Dembiec, D.; Marcus, J. Measurement of catalase activity in tissue extracts. Anal. Biochem. 1970, 34, 30–38. [Google Scholar] [CrossRef]
- Abraham, N.G.; Lutton, J.D.; Levere, R.D. Heme metabolism and erythropoiesis in abnormal iron states: Role of delta-aminolevulinic acid synthase and heme oxygenase. Exp. Hematol. 1985, 13, 838–843. [Google Scholar]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Jennes, L. Chapter 7-immunohistochemical detection of neuropeptides/transmitters in mammalian brain. In Methods in Cell Biology; Conn, P.M., Ed.; Academic Press: Cambridge, MA, USA, 2013; Volume 113, pp. 123–147. [Google Scholar]
- Mahmoud, A.M.; Hussein, O.E.; Hozayen, W.G.; Abd El-Twab, S.M. Methotrexate hepatotoxicity is associated with oxidative stress, and down-regulation of ppargamma and nrf2: Protective effect of 18beta-glycyrrhetinic acid. Chem. Biol. Interact. 2017, 270, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2011, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| Nrf2 | TTGTAGATGACCATGAGTCGC | TGTCCTGCTGTATGCTGCTT |
| NQO-1 | GGCCATCATTTGGGCAAGTC | TCCTTGTGGAACAAAGGCGA |
| HO-1 | GTAAATGCAGTGTTGGCCCC | ATGTGCCAGGCATCTCCTTC |
| NF-κB p65 | CCTCATCTTTCCCTCAGAGCC | GGTCCCGTGTAGCCATTGAT |
| Caspase-3 | GGAGCTTGGAACGCGAAGAA | ACACAAGCCCATTTCAGGGT |
| Bax | AGGACGCATCCACCAAGAAG | CAGTTGAAGTTGCCGTCTGC |
| Bcl-2 | ACTCTTCAGGGATGGGGTGA | TGACATCTCCCTGTTGACGC |
| β-actin | AGGAGTACGATGAGTCCGGC | CGCAGCTCAGTAACAGTCCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Amarat, W.; Abukhalil, M.H.; Alruhaimi, R.S.; Alqhtani, H.A.; Aldawood, N.; Alfwuaires, M.A.; Althunibat, O.Y.; Aladaileh, S.H.; Algefare, A.I.; Alanezi, A.A.; et al. Upregulation of Nrf2/HO-1 Signaling and Attenuation of Oxidative Stress, Inflammation, and Cell Death Mediate the Protective Effect of Apigenin against Cyclophosphamide Hepatotoxicity. Metabolites 2022, 12, 648. https://doi.org/10.3390/metabo12070648
Al-Amarat W, Abukhalil MH, Alruhaimi RS, Alqhtani HA, Aldawood N, Alfwuaires MA, Althunibat OY, Aladaileh SH, Algefare AI, Alanezi AA, et al. Upregulation of Nrf2/HO-1 Signaling and Attenuation of Oxidative Stress, Inflammation, and Cell Death Mediate the Protective Effect of Apigenin against Cyclophosphamide Hepatotoxicity. Metabolites. 2022; 12(7):648. https://doi.org/10.3390/metabo12070648
Chicago/Turabian StyleAl-Amarat, Wesam, Mohammad H. Abukhalil, Reem S. Alruhaimi, Haifa A. Alqhtani, Nouf Aldawood, Manal A. Alfwuaires, Osama Y. Althunibat, Saleem H. Aladaileh, Abdulmohsen I. Algefare, Abdulkareem A. Alanezi, and et al. 2022. "Upregulation of Nrf2/HO-1 Signaling and Attenuation of Oxidative Stress, Inflammation, and Cell Death Mediate the Protective Effect of Apigenin against Cyclophosphamide Hepatotoxicity" Metabolites 12, no. 7: 648. https://doi.org/10.3390/metabo12070648
APA StyleAl-Amarat, W., Abukhalil, M. H., Alruhaimi, R. S., Alqhtani, H. A., Aldawood, N., Alfwuaires, M. A., Althunibat, O. Y., Aladaileh, S. H., Algefare, A. I., Alanezi, A. A., AbouEl-ezz, A. M., Ahmeda, A. F., & Mahmoud, A. M. (2022). Upregulation of Nrf2/HO-1 Signaling and Attenuation of Oxidative Stress, Inflammation, and Cell Death Mediate the Protective Effect of Apigenin against Cyclophosphamide Hepatotoxicity. Metabolites, 12(7), 648. https://doi.org/10.3390/metabo12070648