
Lipid Metabolism Is Dysregulated in the Motor Cortex White Matter in Amyotrophic Lateral Sclerosis

 ,
,  ,
,  and
and
Abstract

1. Introduction
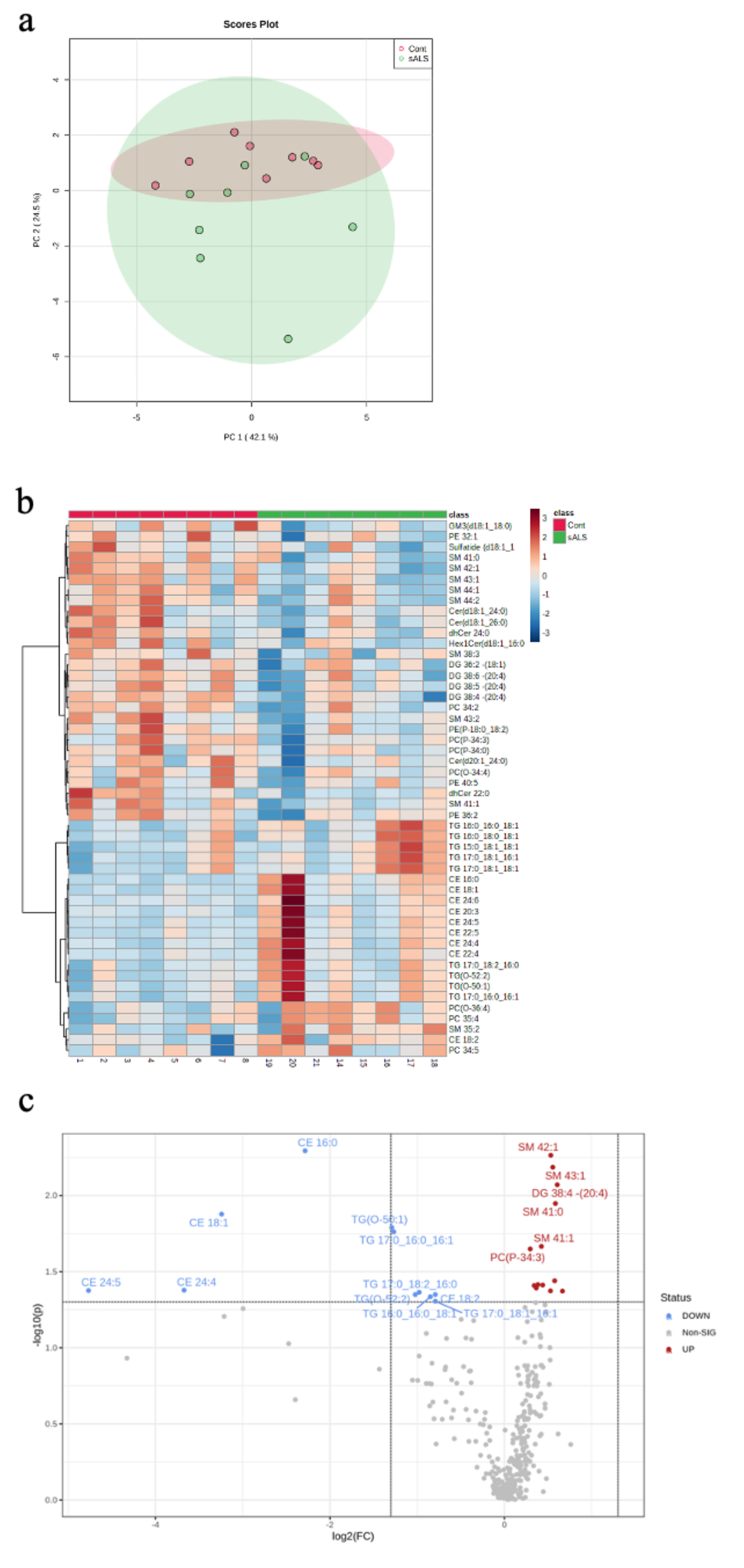
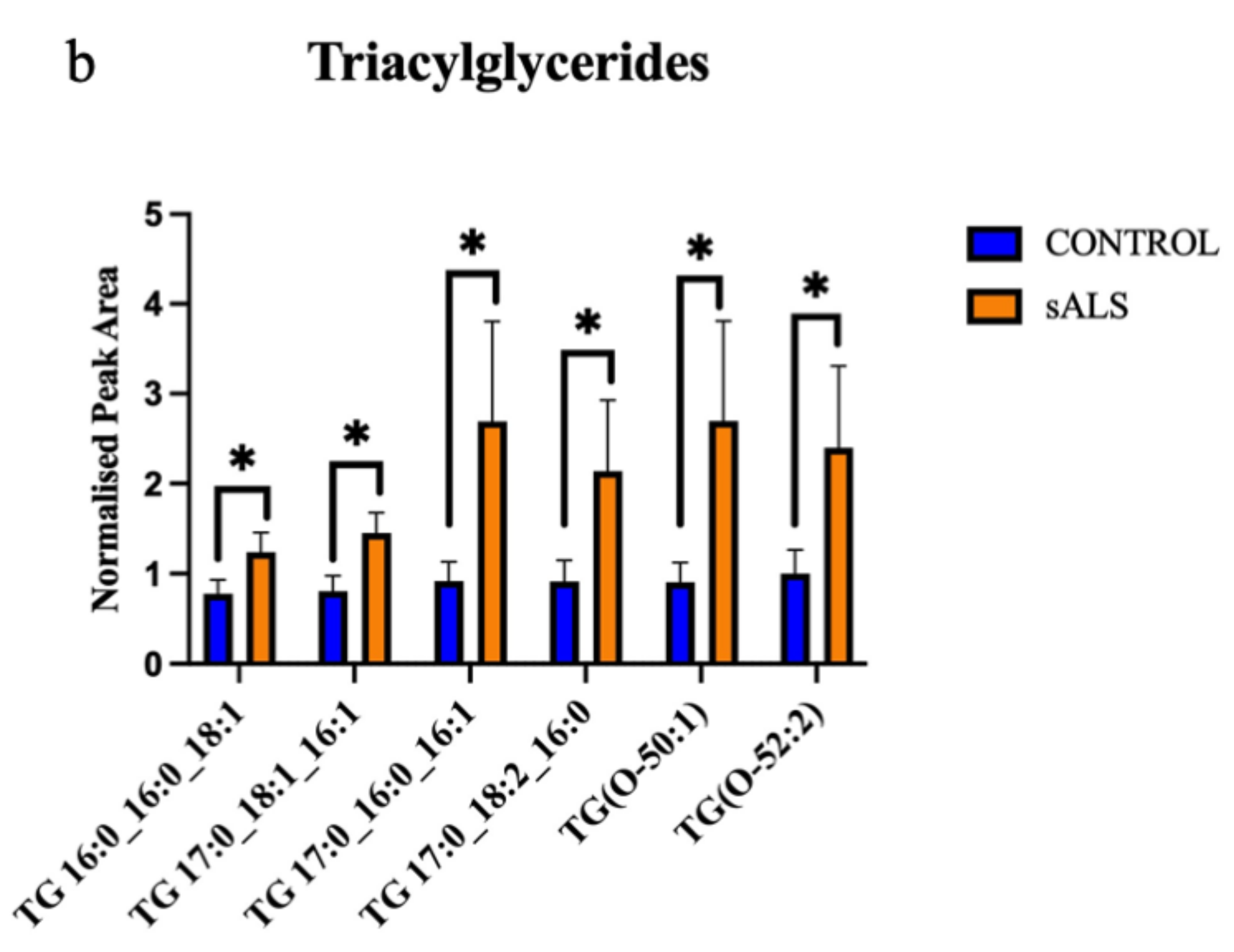
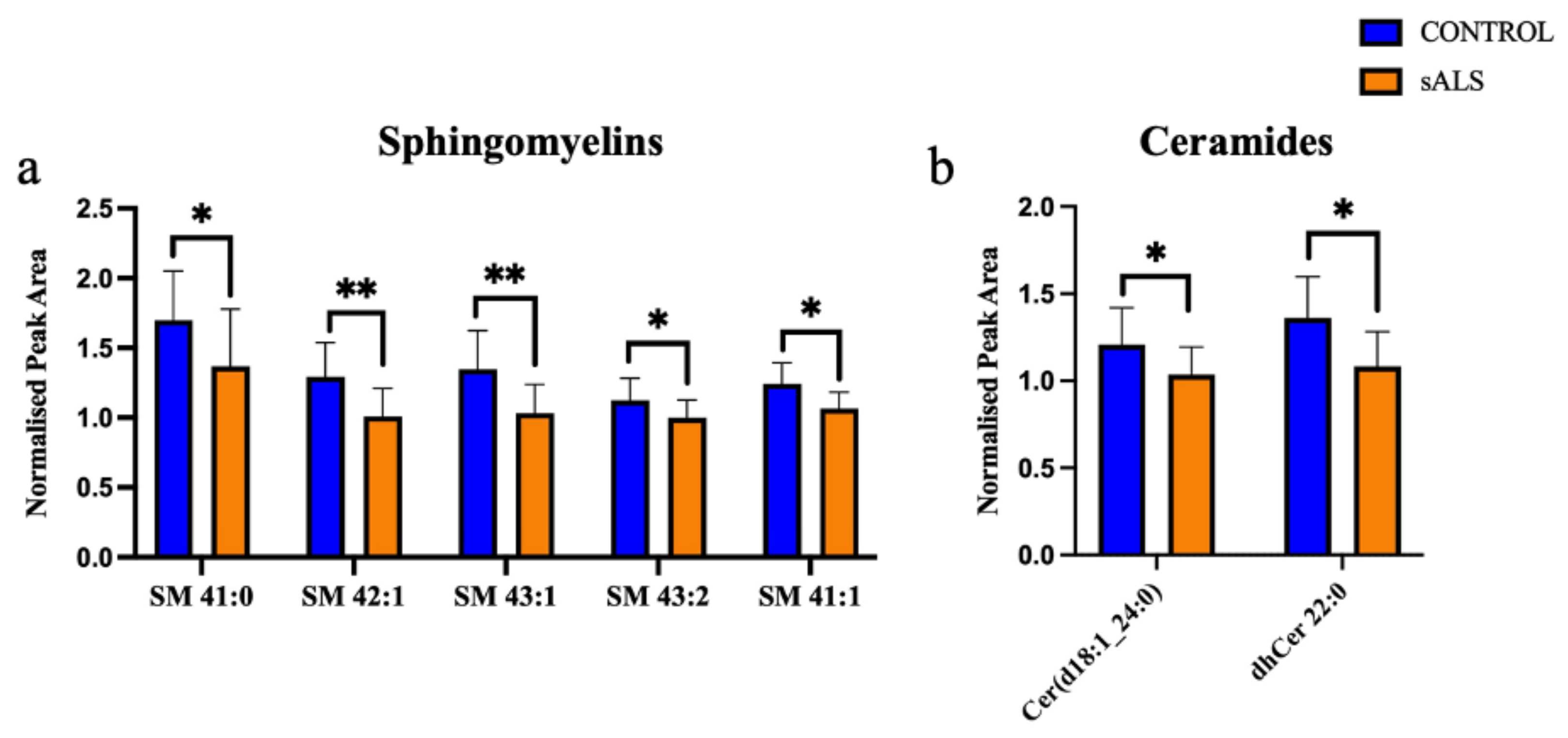
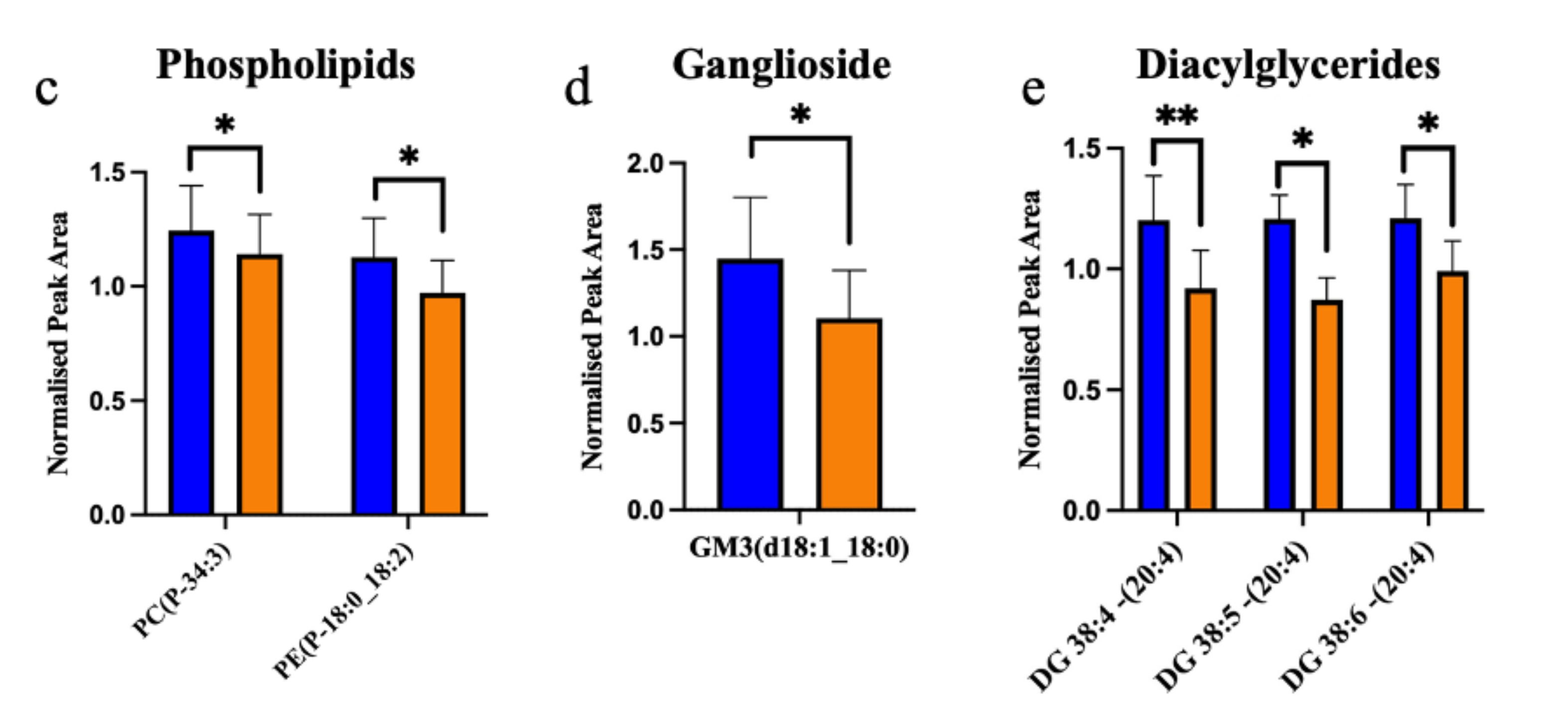
2. Results
2.1. ALS Patients Have an Altered Metabolic Signature in Their Motor Cortex White Matter
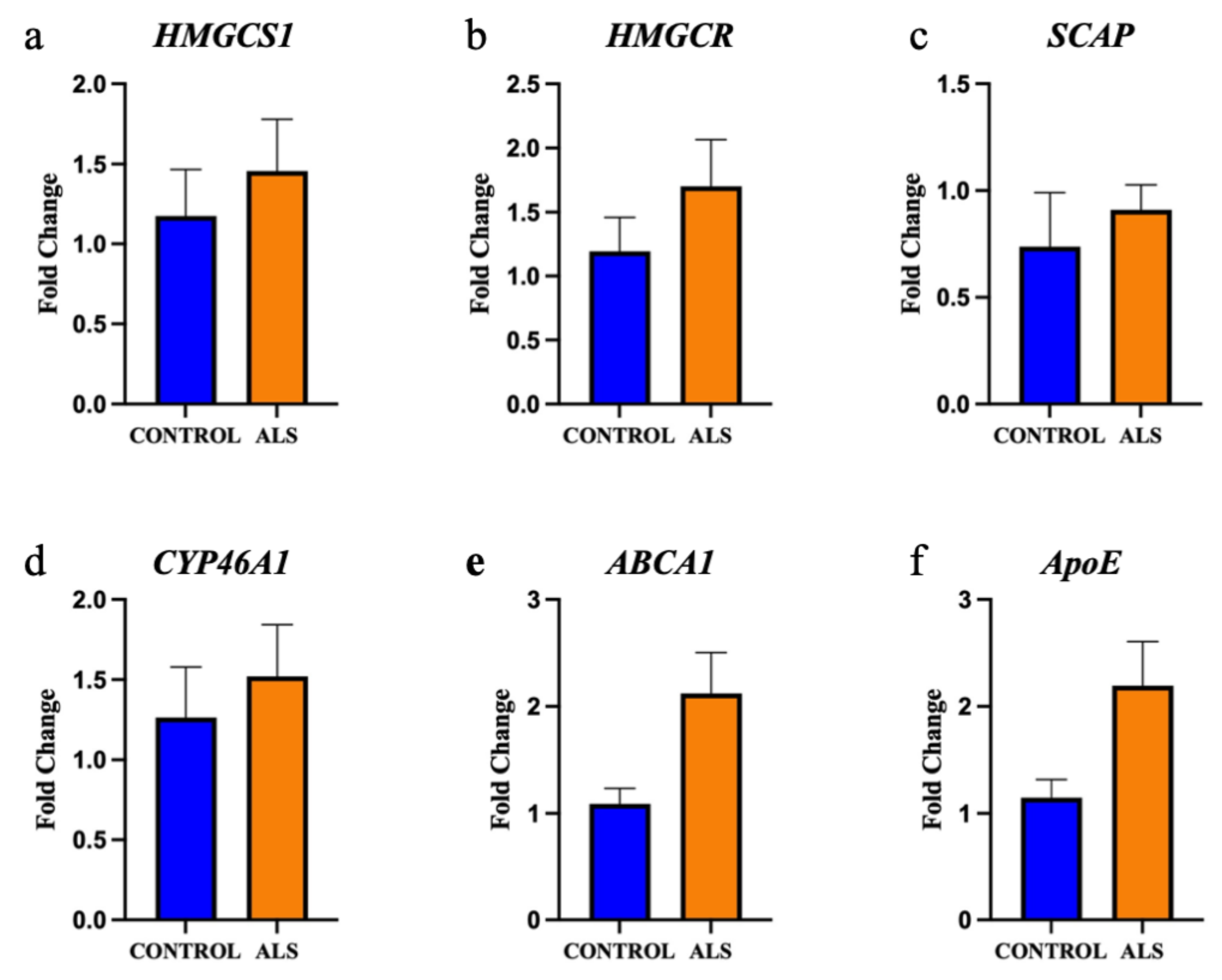
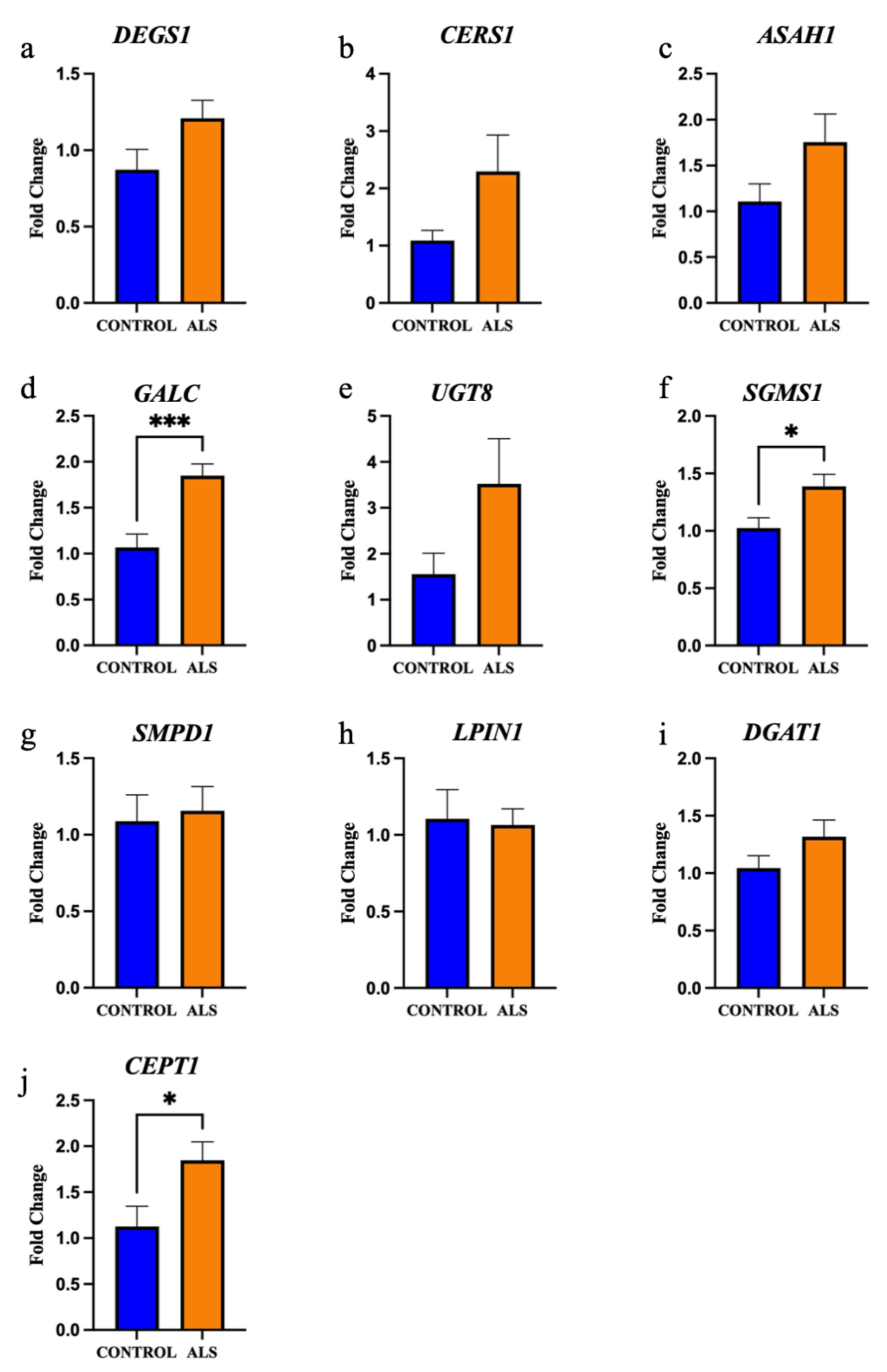
2.2. The Enzymes Responsible for Lipid Metabolism Are Also Altered in the Motor Cortex White Matter of ALS Patients
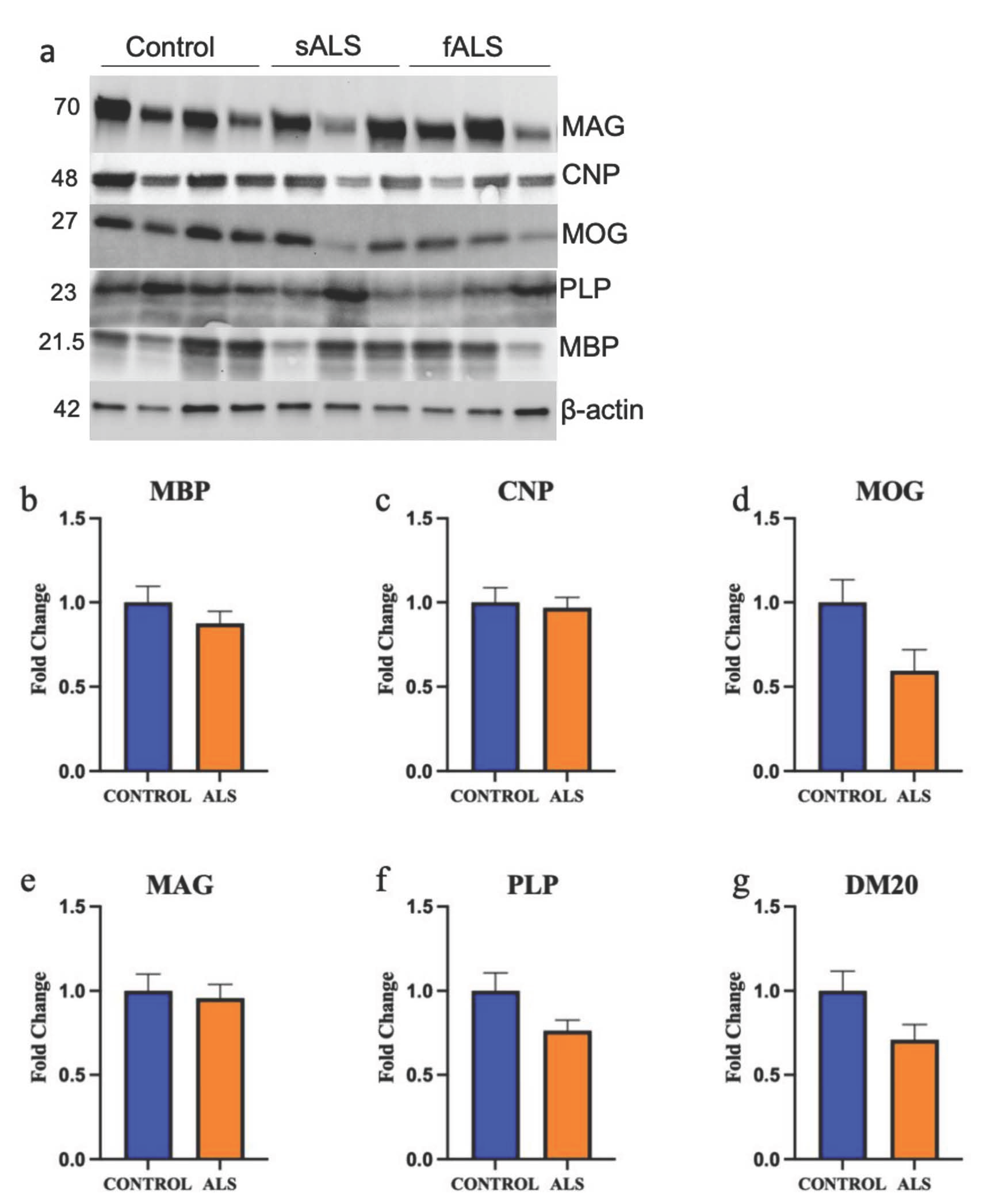
2.3. The Expression of Myelin Proteins Was Not Altered in the White Matter of the Motor Cortex in ALS
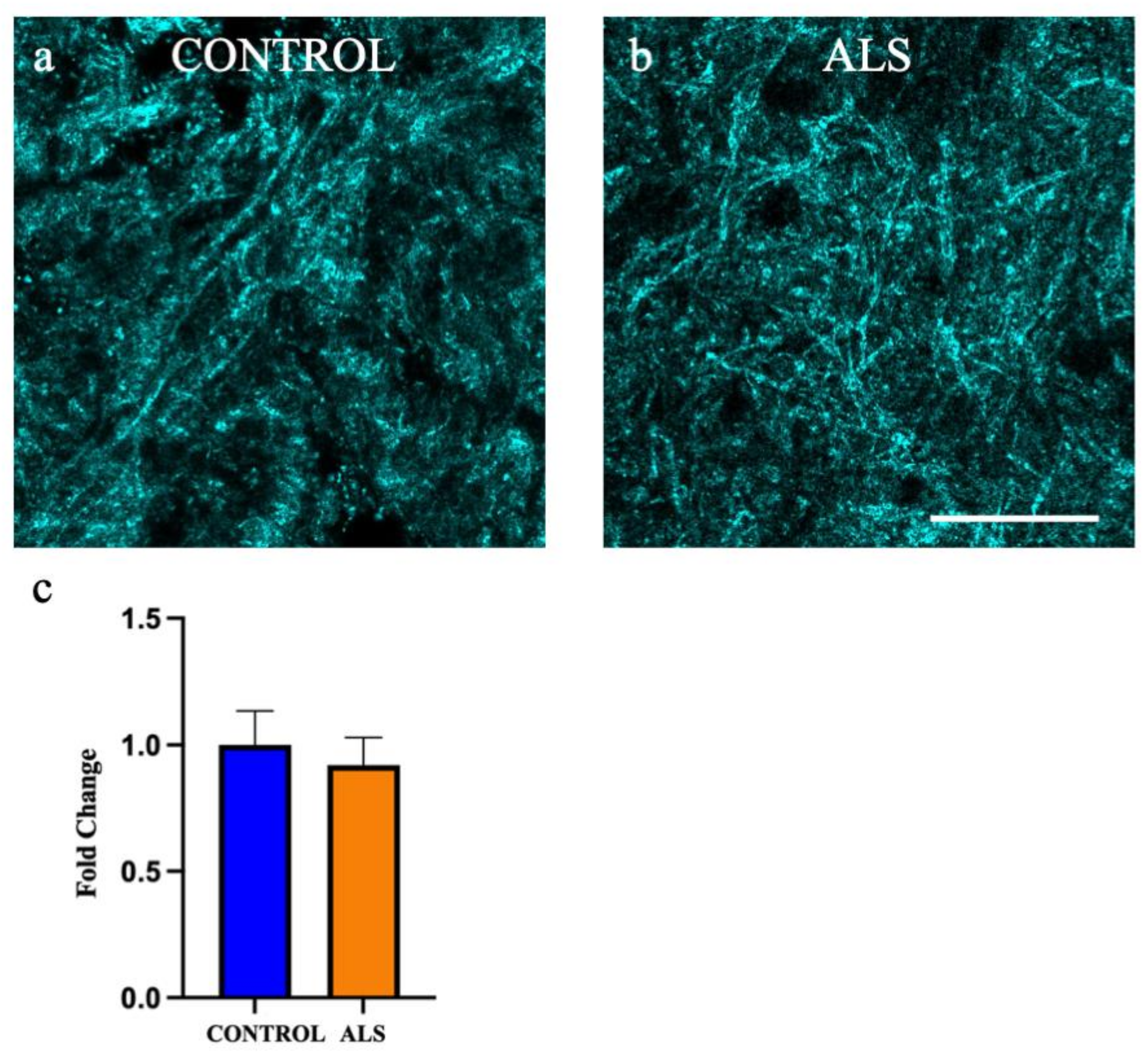
2.4. SCoRe Microscopy Shows No Change to Myelin Density in the White Matter of the Motor Cortex in ALS
3. Discussion
4. Materials and Methods
4.1. Tissue Preparation
4.2. Targeted Lipidomic Analyses of the Motor Cortex White Matter
4.3. Quantitative RT-PCR for Gene Expression of Metabolic Enzymes
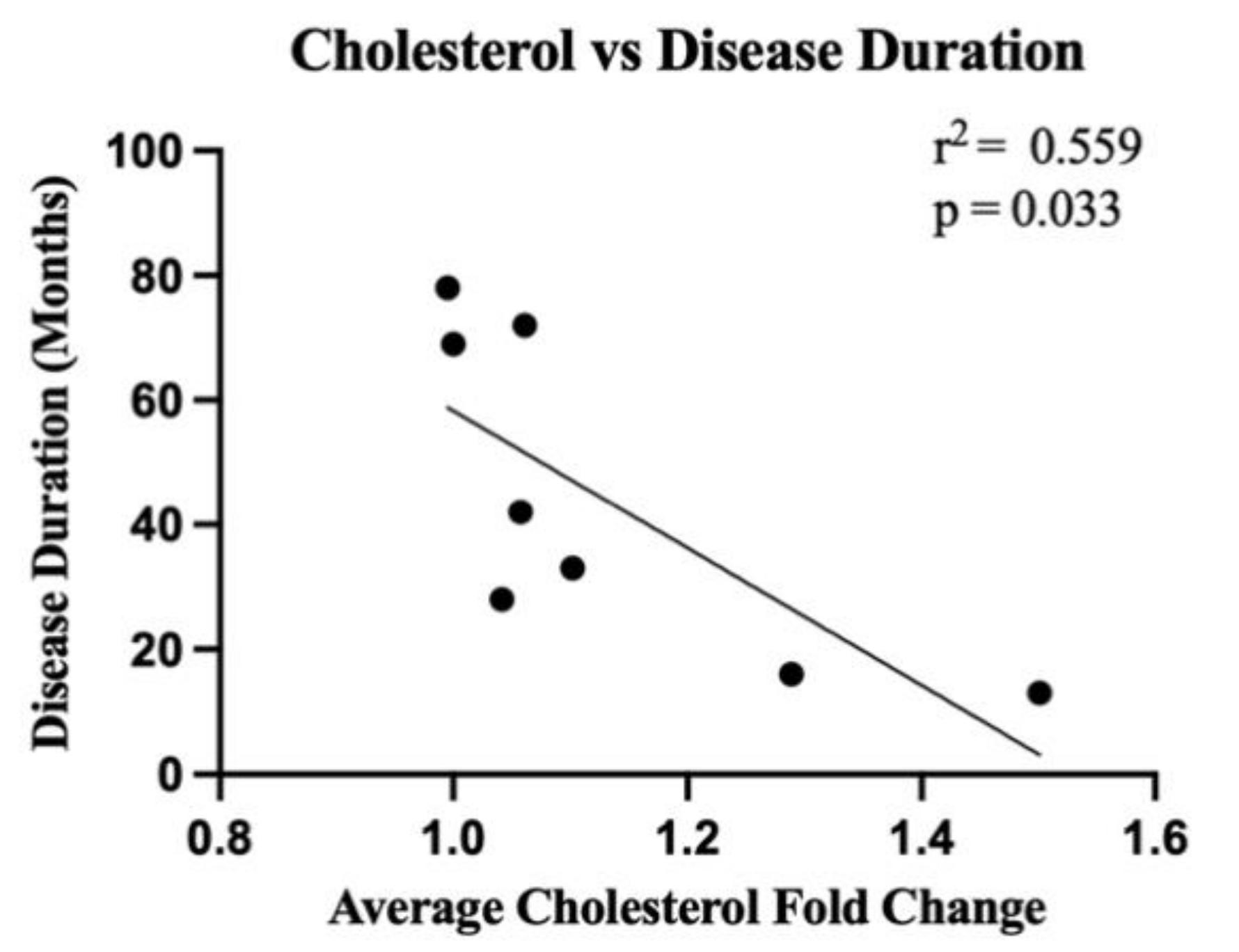
4.4. Correlation of Lipid Expression with Enzymatic Gene Expression and Disease Duration
4.5. Western Blotting for Myelin Proteins
4.6. SCoRe Imaging
4.7. Fluorescent Immunocytochemistry for OLIG2
4.8. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Reilly, E.J.; Wang, H.; Weisskopf, M.G.; Fitzgerald, K.; Falcone, G.; McCullough, M.L.; Thun, M.; Park, Y.; Kolonel, L.N.; Ascherio, A. Premorbid body mass index and risk of amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. Front. Degener. 2013, 14, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Wills, A.-M.; Hubbard, J.; Macklin, E.; Glass, J.; Tandan, R.; Simpson, E.P.; Brooks, B.; Gelinas, D.; Mitsumoto, H.; Mozaffar, T.; et al. Hypercaloric enteral nutrition in patients with amyotrophic lateral sclerosis: A randomised, double-blind, placebo-controlled phase 2 trial. Lancet 2014, 383, 2065–2072. [Google Scholar] [CrossRef]
- Fünfschilling, U.; Supplie, L.M.; Mahad, D.; Boretius, S.; Saab, A.S.; Edgar, J.; Brinkmann, B.G.; Kassmann, C.M.; Tzvetanova, I.D.; Möbius, W.; et al. Glycolytic oligodendrocytes maintain myelin and long-term axonal integrity. Nature 2012, 485, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Morrison, B.M.; Li, Y.; Lengacher, S.; Farah, M.H.; Hoffman, P.N.; Liu, Y.; Tsingalia, A.; Jin, L.; Zhang, P.-W.; et al. Oligodendroglia metabolically support axons and contribute to neurodegeneration. Nature 2012, 487, 443–448. [Google Scholar] [CrossRef]
- Rinholm, J.E.; Bergersen, L.H. Neuroscience: The wrap that feeds neurons. Nature 2012, 487, 435. [Google Scholar] [CrossRef]
- Saab, A.S.; Tzvetanova, I.D.; Nave, K.-A. The role of myelin and oligodendrocytes in axonal energy metabolism. Curr. Opin. Neurobiol. 2013, 23, 1065–1072. [Google Scholar] [CrossRef]
- Neumann, M.; Kwong, L.K.; Truax, A.C.; Vanmassenhove, B.; Kretzschmar, H.A.; Van Deerlin, V.M.; Clark, C.M.; Grossman, M.; Miller, B.L.; Trojanowski, J.Q.; et al. TDP-43-Positive White Matter Pathology in Frontotemporal Lobar Degeneration with Ubiquitin-Positive Inclusions. J. Neuropathol. Exp. Neurol. 2007, 66, 177–183. [Google Scholar] [CrossRef]
- Wang, J.; Ho, W.Y.; Lim, K.; Feng, J.; Tucker-Kellogg, G.; Nave, K.-A.; Ling, S.-C. Cell-autonomous requirement of TDP-43, an ALS/FTD signature protein, for oligodendrocyte survival and myelination. Proc. Natl. Acad. Sci. USA 2018, 115, E10941–E10950. [Google Scholar] [CrossRef]
- Ferraiuolo, L.; Meyer, K.; Sherwood, T.W.; Vick, J.; Likhite, S.; Frakes, A.; Miranda, C.J.; Braun, L.; Heath, P.R.; Pineda, R.; et al. Oligodendrocytes contribute to motor neuron death in ALS via SOD1-dependent mechanism. Proc. Natl. Acad. Sci. USA 2016, 113, E6496–E6505. [Google Scholar] [CrossRef]
- Barton, S.K.; Gregory, J.M.; Selvaraj, B.T.; McDade, K.; Henstridge, C.M.; Spires-Jones, T.L.; James, O.G.; Mehta, A.R.; Story, D.; Burr, K.; et al. Dysregulation in Subcellular Localization of Myelin Basic Protein mRNA Does Not Result in Altered Myelination in Amyotrophic Lateral Sclerosis. Front. Neurosci. 2021, 15, 1032. [Google Scholar] [CrossRef]
- Barton, S.K.; Magnani, D.; James, O.G.; Livesey, M.R.; Selvaraj, B.T.; James, O.T.; Perkins, E.M.; Gregory, J.M.; Cleary, E.; Ausems, C.R.M.; et al. Transactive response DNA-binding protein-43 proteinopathy in oligodendrocytes revealed using an induced pluripotent stem cell model. Brain Commun. 2021, 3, fcab255. [Google Scholar] [CrossRef] [PubMed]
- Pons, A.L.; Higginbottom, A.; Cooper-Knock, J.; Alrafiah, A.; Alofi, E.; Kirby, J.; Shaw, P.; Wood, J.D.; Highley, J.R. Oligodendrocyte pathology exceeds axonal pathology in white matter in human amyotrophic lateral sclerosis. J. Pathol. 2020, 251, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.H.; Li, Y.; Fukaya, M.; Lorenzini, I.; Cleveland, D.; Ostrow, L.; Rothstein, J.D.; Bergles, D.E. Degeneration and impaired regeneration of gray matter oligodendrocytes in amyotrophic lateral sclerosis. Nat. Neurosci. 2013, 16, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Thornton, M.A.; Hughes, E.G. Neuron-oligodendroglia interactions: Activity-dependent regulation of cellular signaling. Neurosci. Lett. 2020, 727, 134916. [Google Scholar] [CrossRef]
- Chen, T.-J.; Kula, B.; Nagy, B.; Barzan, R.; Gall, A.; Ehrlich, I.; Kukley, M. In Vivo Regulation of Oligodendrocyte Precursor Cell Proliferation and Differentiation by the AMPA-Receptor Subunit GluA2. Cell Rep. 2018, 25, 852–861.e7. [Google Scholar] [CrossRef] [PubMed]
- Morell, P.; Quarles, R.H. Characteristic Composition of Myelin. In Basic Neurochemistry: Molecular, Cellular and Medical Aspects, 6th ed.; Lippincott-Raven: Philadelphia, PA, USA, 1999. [Google Scholar]
- Filho, A.D.B.C.; Pinto, I.F.D.; Dantas, L.S.; Xavier, A.M.; Inague, A.; Faria, R.L.; Medeiros, M.H.G.; Glezer, I.; Yoshinaga, M.Y.; Miyamoto, S. Alterations in lipid metabolism of spinal cord linked to amyotrophic lateral sclerosis. Sci. Rep. 2019, 9, 11642. [Google Scholar] [CrossRef] [PubMed]
- Blasco, H.; Veyrat-Durebex, C.; Bocca, C.; Patin, F.; Vourc’h, P.; Nzoughet, J.K.; Lenaers, G.; Andres, C.; Simard, G.; Corcia, P.; et al. Lipidomics Reveals Cerebrospinal-Fluid Signatures of ALS. Sci. Rep. 2017, 7, 17652. [Google Scholar] [CrossRef]
- Philips, T.; Bento-Abreu, A.; Nonneman, A.; Haeck, W.; Staats, K.; Geelen, V.; Hersmus, N.; Küsters, B.; Van Den Bosch, L.; Van Damme, P.; et al. Oligodendrocyte dysfunction in the pathogenesis of amyotrophic lateral sclerosis. Brain 2013, 136 Pt 2, 471–482. [Google Scholar] [CrossRef]
- Chang, K.-J.; Agrawal, I.; Vainshtein, A.; Ho, W.Y.; Xin, W.; Tucker-Kellogg, G.; Susuki, K.; Peles, E.; Ling, S.-C.; Chan, J.R. TDP-43 maximizes nerve conduction velocity by repressing a cryptic exon for paranodal junction assembly in Schwann cells. eLife 2021, 10, e64456. [Google Scholar] [CrossRef]
- Williams, K.A.; Deber, C.M.; Klrschner, O.A. The Structure and Function of Central Nervous System Myelin. Crit. Rev. Clin. Lab. Sci. 1993, 30, 29–64. [Google Scholar] [CrossRef]
- O’Brien, J.S. Stability of the Myelin Membrane. Science 1965, 147, 1099–1107. [Google Scholar] [CrossRef] [PubMed]
- Poitelon, Y.; Kopec, A.M.; Belin, S. Myelin Fat Facts: An Overview of Lipids and Fatty Acid Metabolism. Cells 2020, 9, 812. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, Q. Cholesterol metabolism and homeostasis in the brain. Protein Cell 2015, 6, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Pennetta, G.; Welte, M. Emerging Links between Lipid Droplets and Motor Neuron Diseases. Dev. Cell 2018, 45, 427–432. [Google Scholar] [CrossRef]
- Smolič, T.; Zorec, R.; Vardjan, N. Pathophysiology of Lipid Droplets in Neuroglia. Antioxidants 2021, 11, 22. [Google Scholar] [CrossRef]
- Abdel-Khalik, J.; Yutuc, E.; Crick, P.J.; Gustafsson, J.; Warner, M.; Roman, G.; Talbot, K.; Gray, E.; Griffiths, W.J.; Turner, M.R.; et al. Defective cholesterol metabolism in amyotrophic lateral sclerosis. J. Lipid Res. 2017, 58, 267–278. [Google Scholar] [CrossRef]
- Cutler, R.G.; Pedersen, W.A.; Camandola, S.; Rothstein, J.D.; Mattson, M.P. Evidence that accumulation of ceramides and cholesterol esters mediates oxidative stress-induced death of motor neurons in amyotrophic lateral sclerosis. Ann. Neurol. 2002, 52, 448–457. [Google Scholar] [CrossRef]
- Don, A.S.; Hsiao, J.-H.T.; Bleasel, J.M.; Couttas, T.A.; Halliday, G.M.; Kim, W.S. Altered lipid levels provide evidence for myelin dysfunction in multiple system atrophy. Acta Neuropathol. Commun. 2014, 2, 150. [Google Scholar] [CrossRef]
- Henriques, A.; Croixmarie, V.; Bouscary, A.; Mosbach, A.; Keime, C.; Boursier-Neyret, C.; Walter, B.; Spedding, M.; Loeffler, J.-P. Sphingolipid Metabolism Is Dysregulated at Transcriptomic and Metabolic Levels in the Spinal Cord of an Animal Model of Amyotrophic Lateral Sclerosis. Front. Mol. Neurosci. 2018, 10, 433. [Google Scholar] [CrossRef]
- Stallings, N.R.; Puttaparthi, K.; Dowling, K.J.; Luther, C.M.; Burns, D.K.; Davis, K.; Elliott, J.L. TDP-43, an ALS Linked Protein, Regulates Fat Deposition and Glucose Homeostasis. PLoS ONE 2013, 8, e71793. [Google Scholar] [CrossRef]
- Watkins, J.A.; Alix, J.J.P.; Shaw, P.J.; Mead, R.J. Extensive phenotypic characterisation of a human TDP-43Q331K transgenic mouse model of amyotrophic lateral sclerosis (ALS). Sci. Rep. 2021, 11, 16659. [Google Scholar] [CrossRef] [PubMed]
- Niebroj-Dobosz, I.; Rafałowska, J.; Fidziańska, A.; Gadamski, R.; Grieb, P. Myelin composition of spinal cord in a model of amyotrophic lateral sclerosis (ALS) in SOD1G93A transgenic rats. Folia Neuropathol. 2007, 45, 236–241. [Google Scholar] [PubMed]
- Raffaele, S.; Boccazzi, M.; Fumagalli, M. Oligodendrocyte Dysfunction in Amyotrophic Lateral Sclerosis: Mechanisms and Therapeutic Perspectives. Cells 2021, 10, 565. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Halliday, G.M.; Kim, W.S. Exploring Myelin Dysfunction in Multiple System Atrophy. Exp. Neurobiol. 2014, 23, 337–344. [Google Scholar] [CrossRef]
- Saugier-Veber, P.; Munnich, A.; Bonneau, D.; Rozet, J.-M.; Le Merrer, M.; Gil, R.; Boespflug-Tanguy, O. X–linked spastic paraplegia and Pelizaeus–Merzbacher disease are allelic disorders at the proteolipid protein locus. Nat. Genet. 1994, 6, 257–262. [Google Scholar] [CrossRef]
- Huynh, K.; Barlow, C.K.; Jayawardana, K.S.; Weir, J.M.; Mellett, N.A.; Cinel, M.; Magliano, D.; Shaw, J.E.; Drew, B.G.; Meikle, P.J. High-Throughput Plasma Lipidomics: Detailed Mapping of the Associations with Cardiometabolic Risk Factors. Cell Chem. Biol. 2019, 26, 71–84.e4. [Google Scholar] [CrossRef]
- Huynh, K.; Pernes, G.; Mellett, N.A.; Meikle, P.J.; Murphy, A.J.; Lancaster, G.I. Lipidomic Profiling of Murine Macrophages Treated with Fatty Acids of Varying Chain Length and Saturation Status. Metabolites 2018, 8, 29. [Google Scholar] [CrossRef]
- Liebisch, G.; Vizcaino, J.A.; Köfeler, H.; Trötzmüller, M.; Griffiths, W.; Schmitz, G.; Spener, F.; Wakelam, M. Shorthand notation for lipid structures derived from mass spectrometry. J. Lipid Res. 2013, 54, 1523–1530. [Google Scholar] [CrossRef]
- Gonsalvez, D.G.; Yoo, S.; Fletcher, J.; Wood, R.J.; Craig, G.A.; Murray, S.S.; Xiao, J. Imaging and Quantification of Myelin Integrity After Injury with Spectral Confocal Reflectance Microscopy. Front. Mol. Neurosci. 2019, 12, 275. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Diagnosis | Age * | Sex | PMI # | Region | Onset | Duration * |
|---|---|---|---|---|---|---|---|
| 1 | Control | 73.6 | M | 49 | BA4 | / | / |
| 2 | Control | 52.1 | M | 33 | BA4 | / | / |
| 3 | Control | 63.9 | M | 68 | BA4 | / | / |
| 4 | Control | 75.9 | M | 50 | BA4 | / | / |
| 5 | Control | 77.5 | M | 28 | BA4 | / | / |
| 6 | Control | 57.6 | M | 20.5 | BA4 | / | / |
| 7 | Control | 73.5 | M | 22 | BA4 | / | / |
| 8 | Control | 79.6 | M | 31.5 | BA4 | / | / |
| 9 | fALS | 43.8 | M | 77 | BA4 | Limb-lower | 1.6 |
| 10 | fALS | 58.4 | M | 24.5 | BA4 | Limb-upper | 2.5 |
| 11 | fALS | 64.3 | F | 39.5 | BA4 | Limb-upper (left) | 2.5 |
| 12 | fALS | 69.3 | F | 45 | BA4 | Limb-upper & lower (right) | 3.0 |
| 13 | fALS | 72.8 | M | 21.5 | BA4 | Limb-lower (left) | 3.0 |
| 14 | sALS | 41.6 | M | 42 | BA4 | Limb-lower | 2.3 |
| 15 | sALS | 53 | M | 29 | BA4 | Limb-lower (right) | 6.0 |
| 16 | sALS | 64.8 | M | 45 | BA4 | Limb-lower | 6.5 |
| 17 | sALS | 75.7 | M | 26 | BA4 | Limb-upper & lower | 2.8 |
| 18 | sALS | 78.1 | M | 13.5 | BA4 | Bulbar | 3.5 |
| 19 | sALS | 66.9 | M | 21 | BA4 | Limb-lower (left) | 1.3 |
| 20 | sALS | 57.6 | M | 21.5 | BA4 | Limb-upper & lower (left) | 1.1 |
| 21 | sALS | 66.1 | M | 32.5 | BA4 | Limb-lower | 5.75 |
| 22 | fALS | 41.9 | F | 30.5 | BA4 | Limb-upper & lower | 0.1 (7 weeks) |
| Gene Name | Forward Primer | Reverse Primer | Anneal Temp (°C) |
|---|---|---|---|
| ASAH1 | 5′-GCACAAGTTATGAAGGAAGCCAAG | 5′-TCCAATGATTCCTTTCTGTCTCG | 57 |
| CERS1 | 5′-CTGCTCCAGGGAAGCTTCTA | 5′-GGAGGAGACGATGAGGATGA | 59 |
| DEGS1 | 5′-GCAAAGCAATGTGGAATCGC | 5′-ACCAGCCCTCAAAATCGGTA | 57 |
| SGMS1 | 5′-TGAGCCTCTGGAGCATTTCA | 5′-CCGTTCTTGTGTGCTTCCAA | 61 |
| SMPD1 | 5′-CAGACTCGTCAGGACCAACT | 5′-GGGGAGGGAAGCTATTGACA | 64 |
| UGT8 | 5′-ATGGGTAAATGGTGCTAATG | 5′-TCTGGTCATAGTATCATAATGG | 63 |
| GALC | 5′-GGCTCATTATCCTGGAACCC | 5′-GCCTGCACCCATGTCACTAT | 57 |
| HMGCS1 | 5′-TGTACACATCTTCAGTATATGGTTCCC | 5′-AAGAAAACACTCCAATTCTCTTCCCT | 57 |
| HMGCR | 5′-TGCTCGTGGAATGGCAATTT | 5′-ACCAACTCCAATCACAAGACA | 63 |
| SCAP | 5′-TGGGGATGTCACCTCCCTTAC | 5′-TGCTGAATGGAGTAGAACTTGATGC | 59 |
| ABCA1 | 5′-TGCAAGGCTACCAGTTACATT | 5′-TTAGTGTTCTCAGGATTGGCT | 59 |
| ApoE | 5′-TGGACGAGGTGAAGGAGCAG | 5′-CTCGAACCAGCTCTTGAGGC | 57 |
| CYP46A1 | 5′-CAGGGAAGAGGAAGCAGCTC | 5′-ATCTGTGTGAGGATGTCGGC | 60 |
| CEPT1 | 5′-GCGGGATCCATGAGTGGGCATCGATCAACA | 5′-GCGGTCGACTTAATGATTAGAATGAGCTGT | 63 |
| LPIN1 | 5′-TGAAAAGGGGCTCTGTGGAC | 5′-ACTACAGAGCTGCTTGACGG | 59 |
| DGAT1 | 5′-TGCAGGATTCTTTATTCAGCTCT | 5′-GCATCACCACACACCAGTTC | 57 |
| MBP | 5′-CTTCTTTGGCGGTGACAGG | 5′-CGGGGTTTTCATCTTGGGTC | 57 |
| ACTB | 5′-GTTACAGGAACTCCCTTGCCATCC | 5′-CACCTCCCCTGTGTGGACTTGGG | 57 |
| 18S | 5′-GTAACCCGTTGAACCCCATT | 5′-CCATCCAATCGGTAGTAGCG | 59 |
| ASAH1 | 5′-GCACAAGTTATGAAGGAAGCCAAG | 5′-TCCAATGATTCCTTTCTGTCTCG | 57 |
| CERS1 | 5′-CTGCTCCAGGGAAGCTTCTA | 5′-GGAGGAGACGATGAGGATGA | 59 |
| DEGS1 | 5′-GCAAAGCAATGTGGAATCGC | 5′-ACCAGCCCTCAAAATCGGTA | 57 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadler, G.L.; Lewis, K.N.; Narayana, V.K.; De Souza, D.P.; Mason, J.; McLean, C.; Gonsalvez, D.G.; Turner, B.J.; Barton, S.K. Lipid Metabolism Is Dysregulated in the Motor Cortex White Matter in Amyotrophic Lateral Sclerosis. Metabolites 2022, 12, 554. https://doi.org/10.3390/metabo12060554
Sadler GL, Lewis KN, Narayana VK, De Souza DP, Mason J, McLean C, Gonsalvez DG, Turner BJ, Barton SK. Lipid Metabolism Is Dysregulated in the Motor Cortex White Matter in Amyotrophic Lateral Sclerosis. Metabolites. 2022; 12(6):554. https://doi.org/10.3390/metabo12060554
Chicago/Turabian StyleSadler, Gemma L., Katherine N. Lewis, Vinod K. Narayana, David P. De Souza, Joel Mason, Catriona McLean, David G. Gonsalvez, Bradley J. Turner, and Samantha K. Barton. 2022. "Lipid Metabolism Is Dysregulated in the Motor Cortex White Matter in Amyotrophic Lateral Sclerosis" Metabolites 12, no. 6: 554. https://doi.org/10.3390/metabo12060554
APA StyleSadler, G. L., Lewis, K. N., Narayana, V. K., De Souza, D. P., Mason, J., McLean, C., Gonsalvez, D. G., Turner, B. J., & Barton, S. K. (2022). Lipid Metabolism Is Dysregulated in the Motor Cortex White Matter in Amyotrophic Lateral Sclerosis. Metabolites, 12(6), 554. https://doi.org/10.3390/metabo12060554