
Investigating Potential GLP-1 Receptor Agonists in Cyclopeptides from Pseudostellaria heterophylla, Linum usitatissimum, and Drymaria diandra, and Peptides Derived from Heterophyllin B for the Treatment of Type 2 Diabetes: An In Silico Study


Abstract
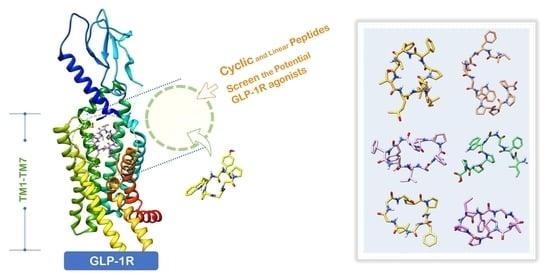
1. Introduction
1.1. GLP-1, GLP-1R Agonists, and Type 2 Diabetes Treatment
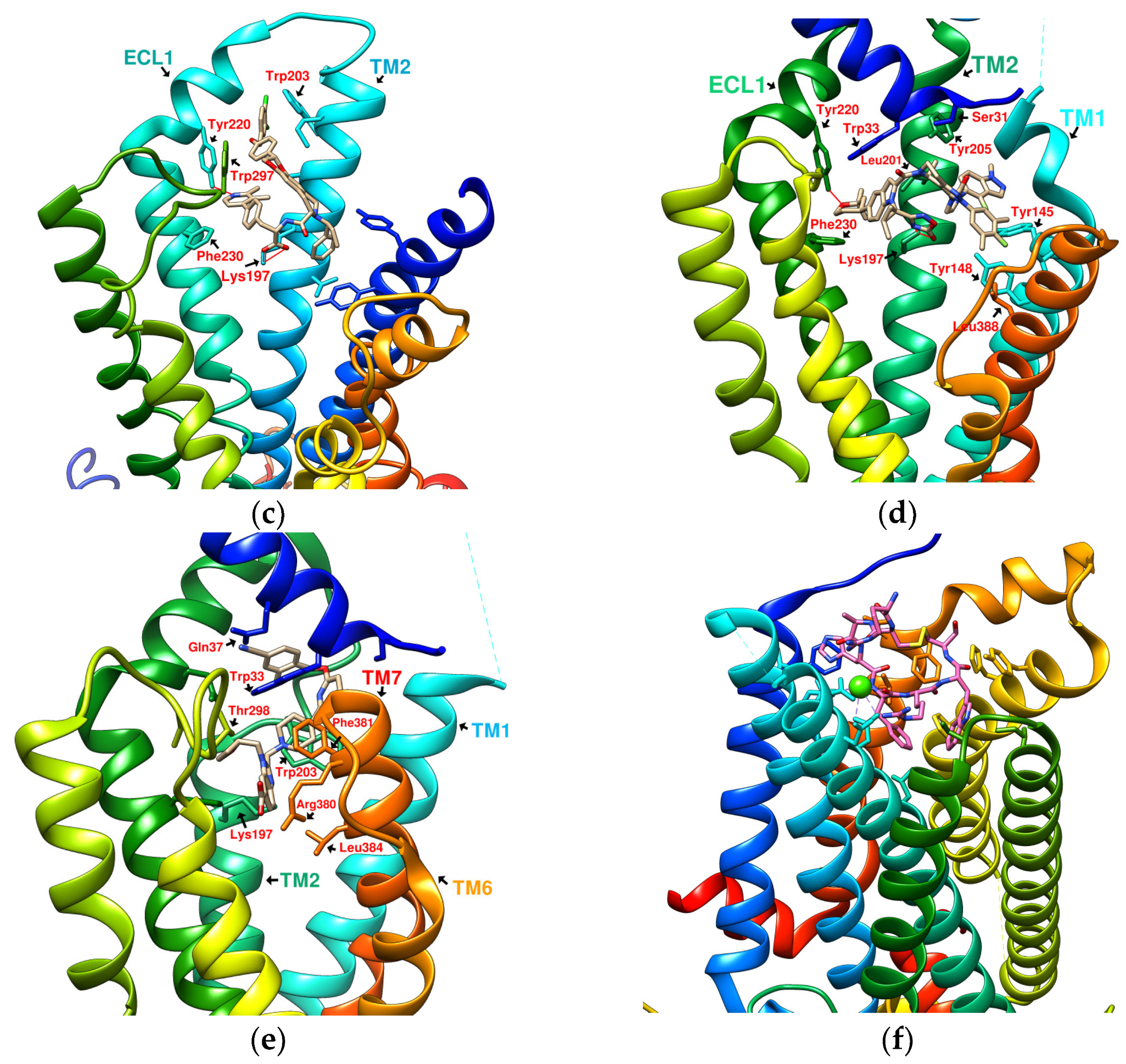
1.2. Development of Small-Molecule Nonpeptide GLP-1R Agonists
1.3. Reference Case of Cyclic Peptides (Cyclopeptides) as GPCR Agonists/Antagonists
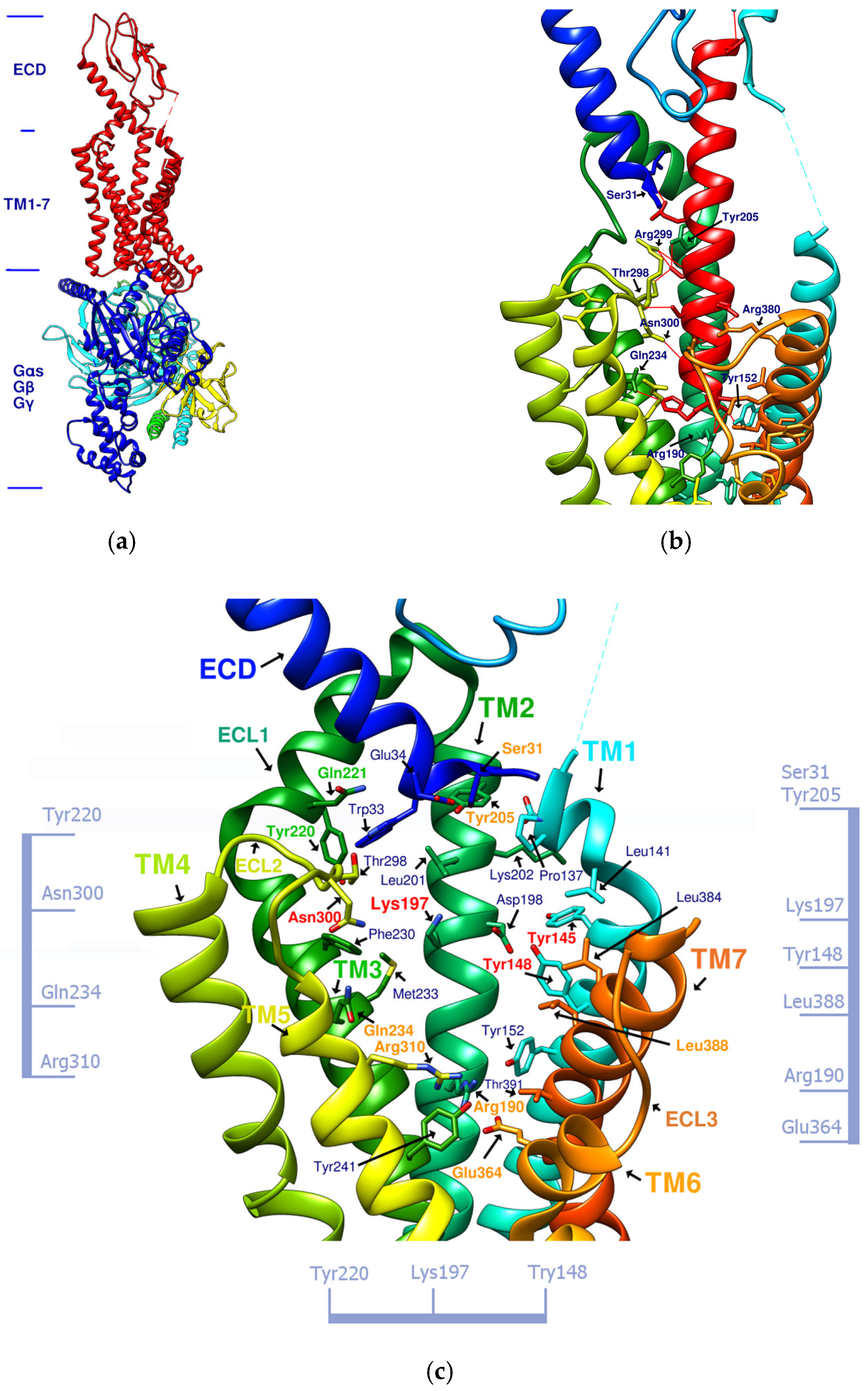
1.4. Correlation between GLP-1R Active Site and Accommodated Molecular Size
1.5. The Research Potential of Pseudostellaria heterophylla, Linum usitatissimum, and Drymaria diandra as Natural Hypoglycemic Products and GLP-1R Agonists
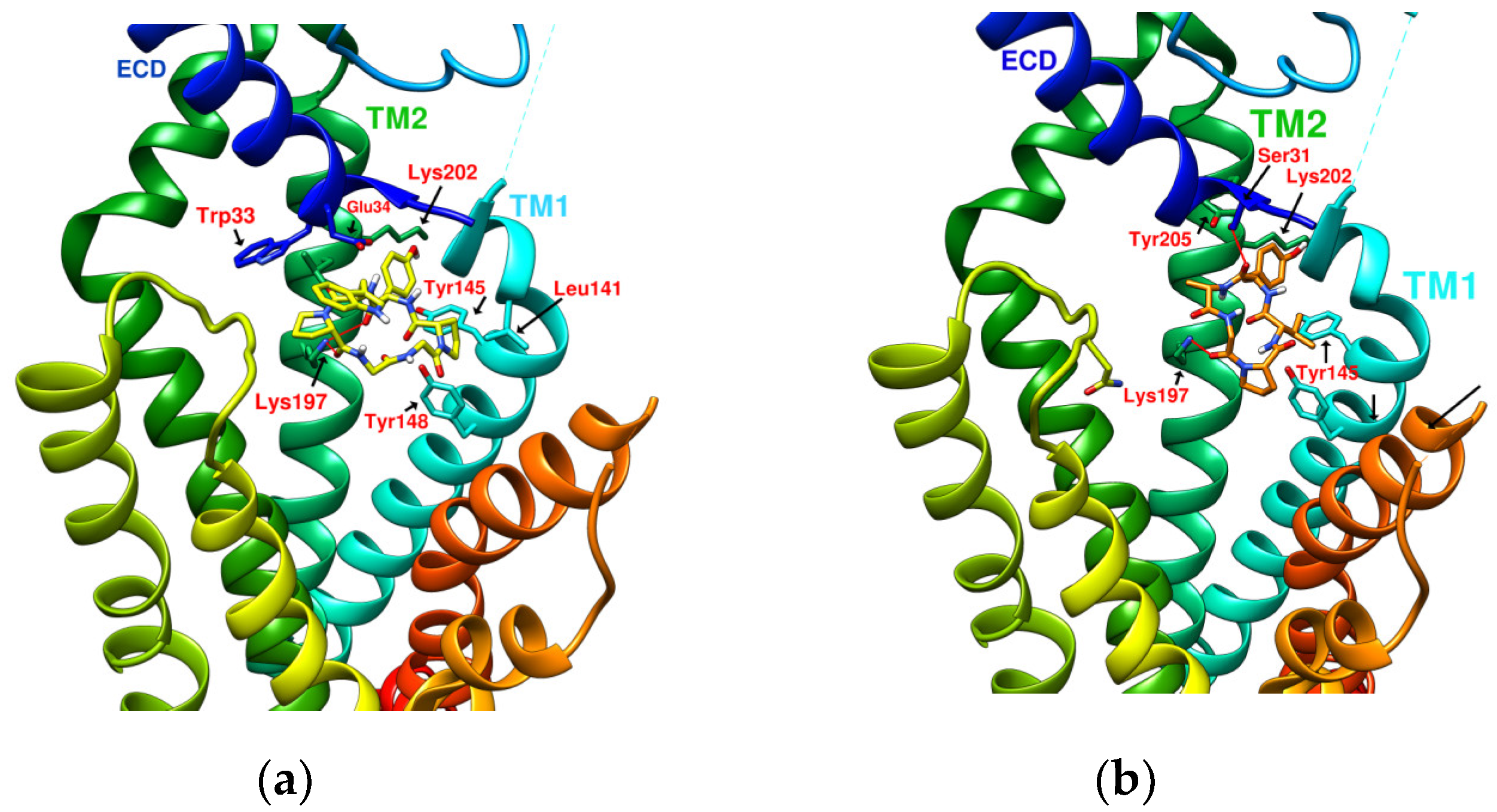
1.6. Exploring the Possibility of HB Linear Precursor “IFGGLPPP” Derivatives Acting on GLP-1R
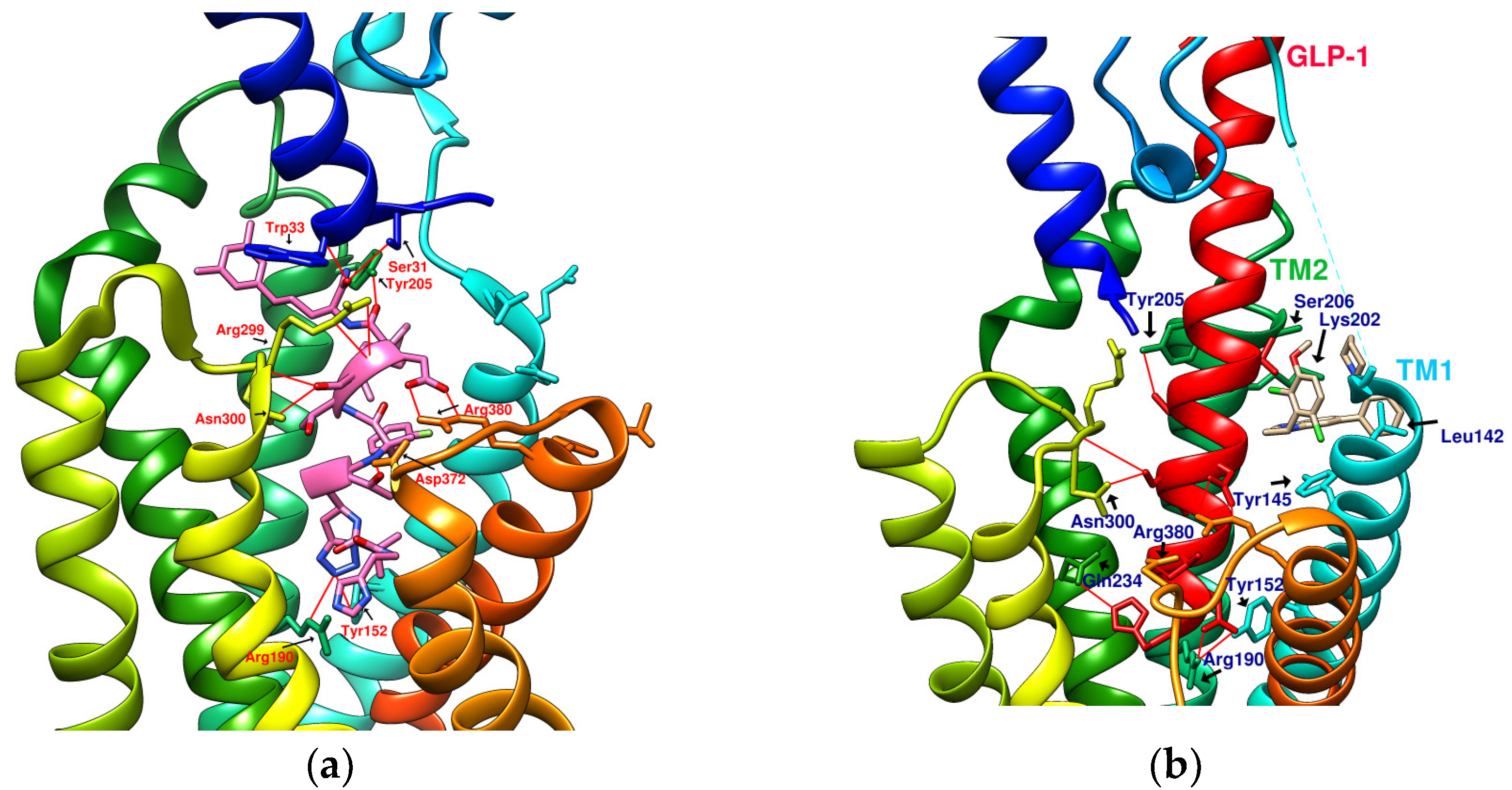
2. Results and Discussion
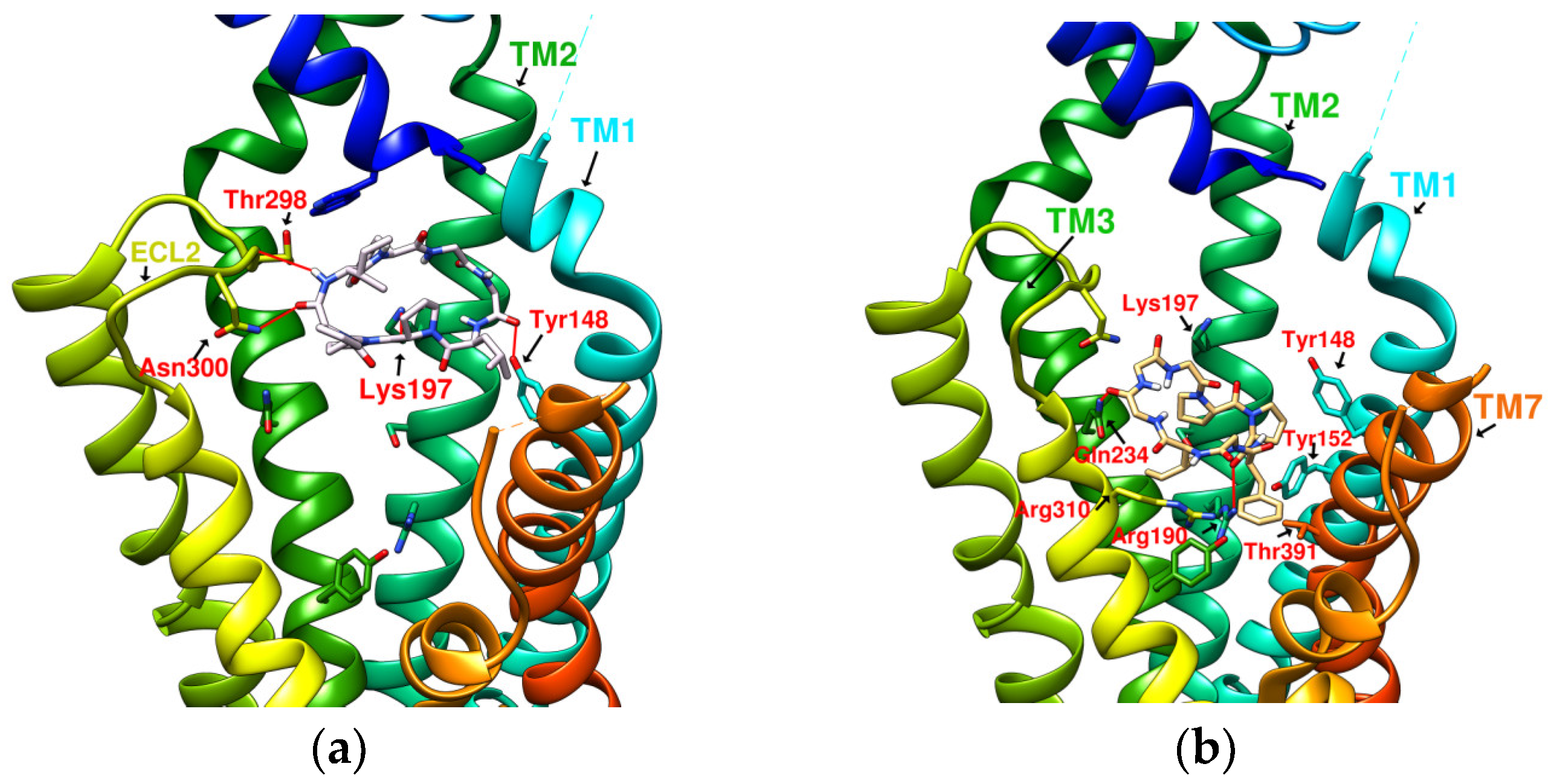
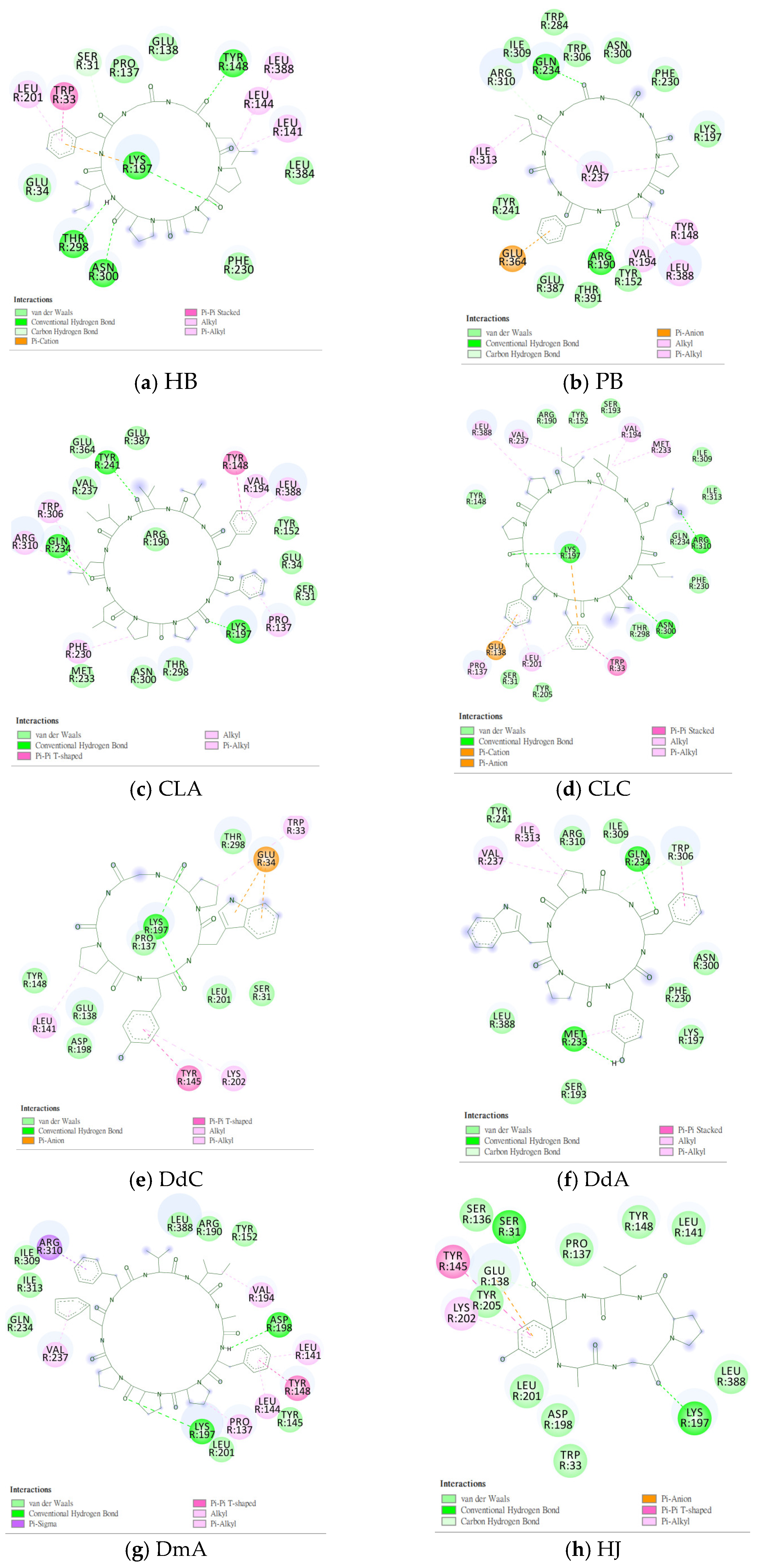
2.1. Prediction of the Binding Affinity of Plant-Derived Cyclopeptides Docking with GLP-1R
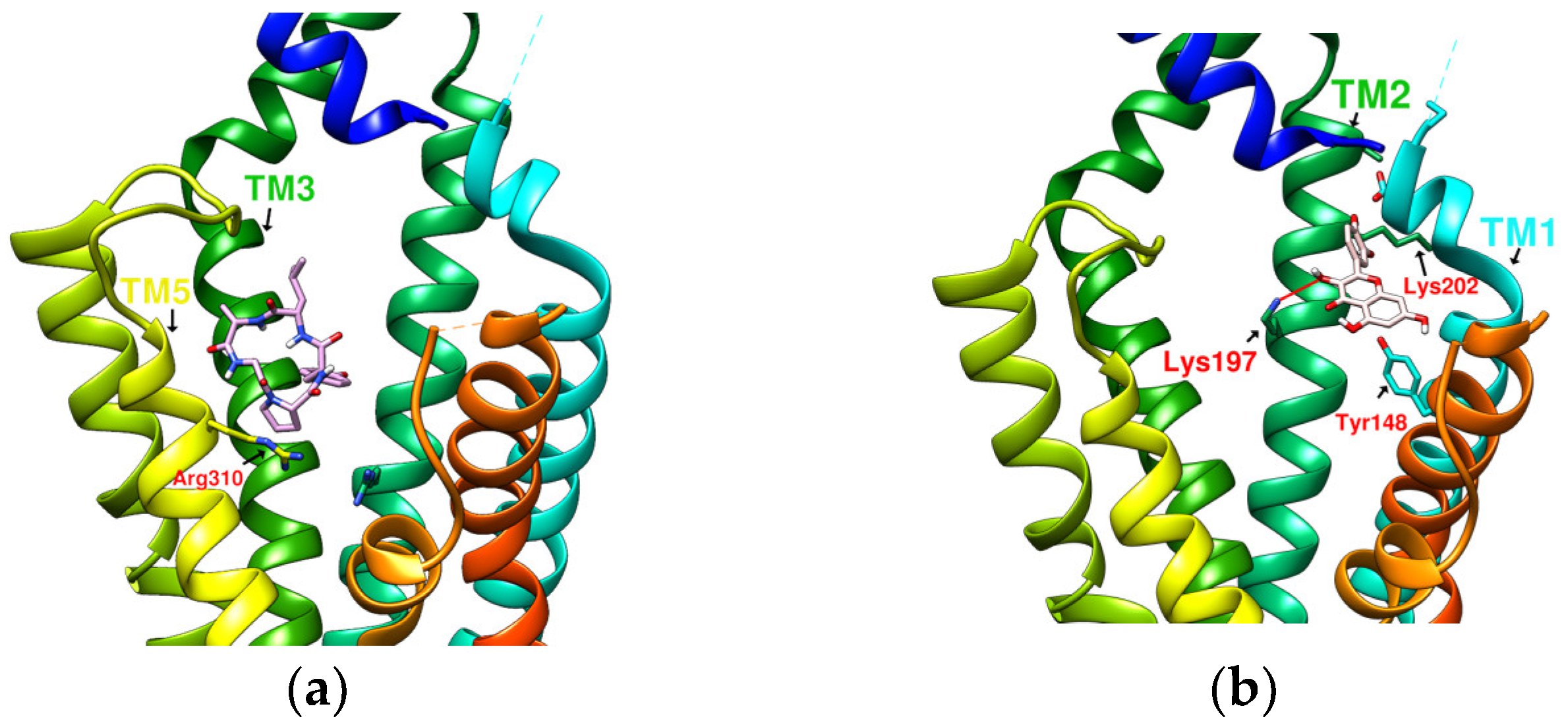
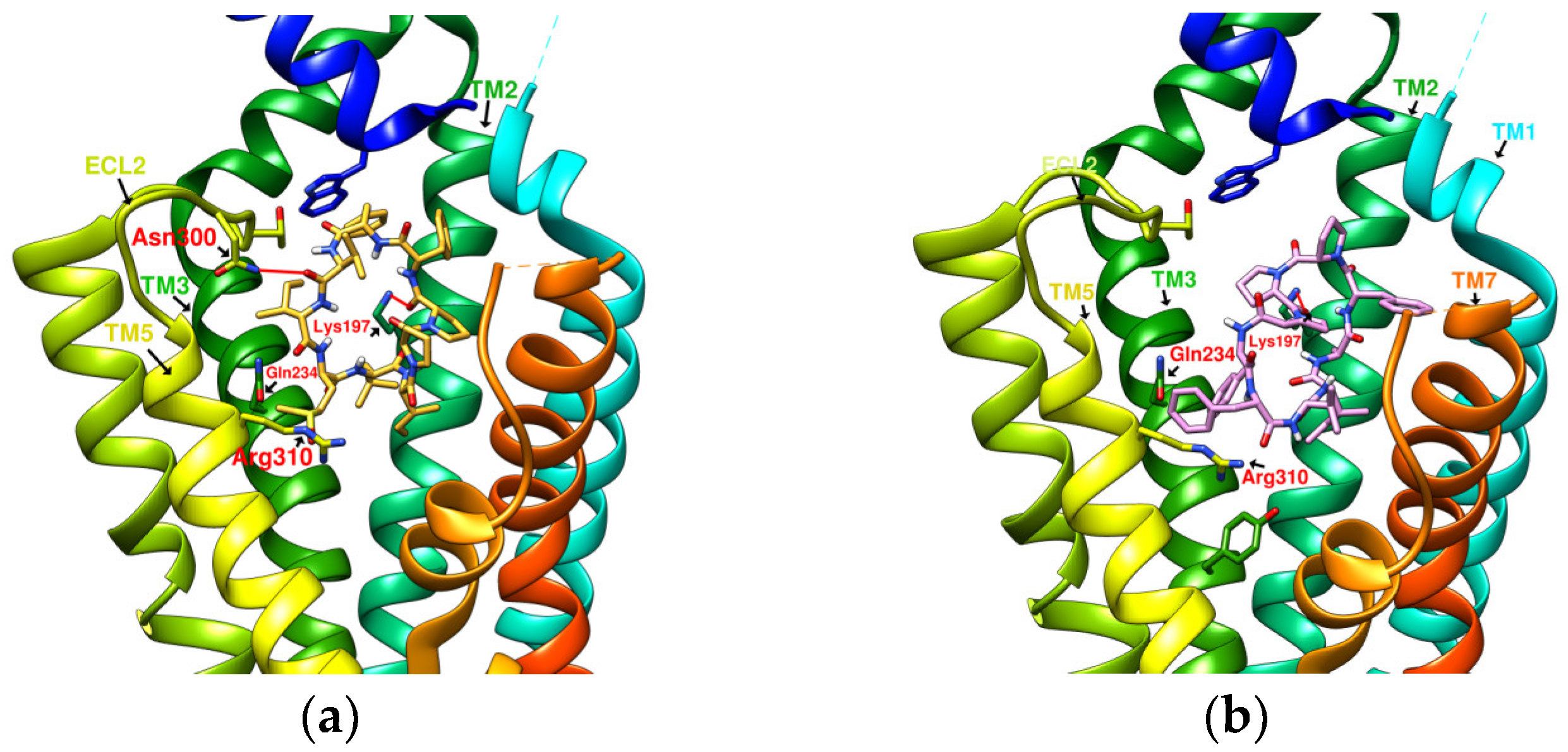
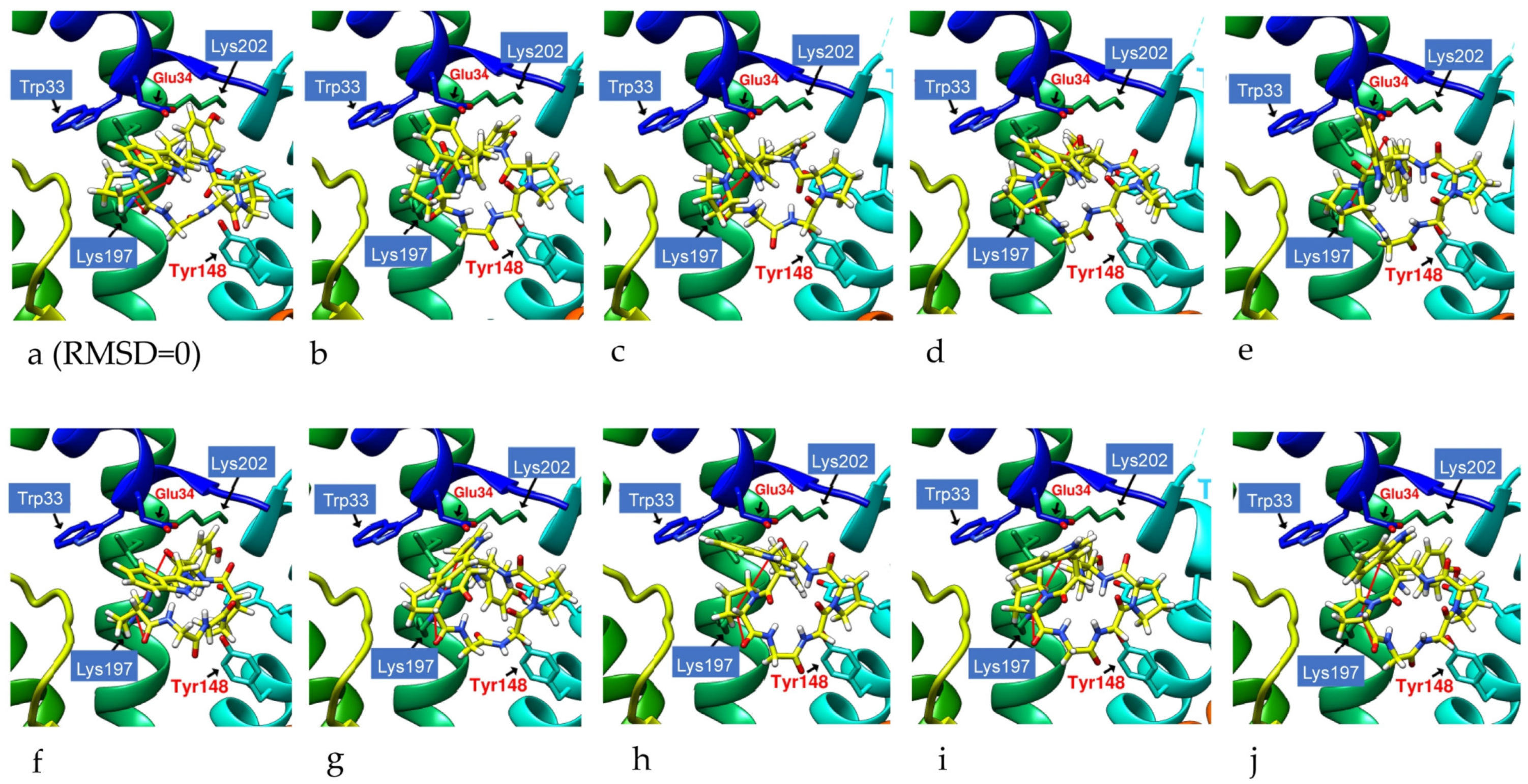
2.2. The Research Potential of DdC as GLP-1RA and Its Configuration Characteristics in Docking
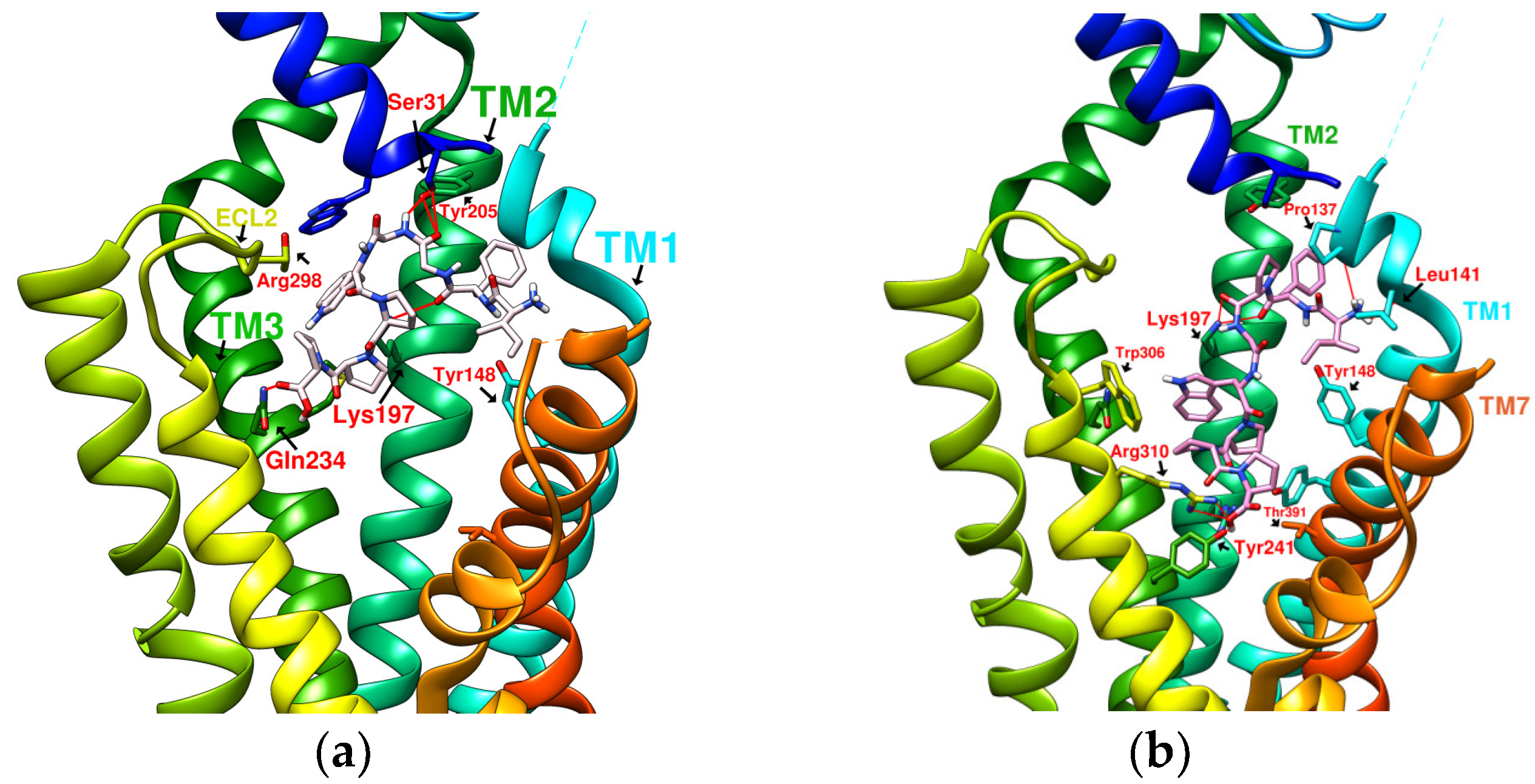
2.3. The Predicted Configuration of Other Cyclopeptides Besides DdC in the Active Site of GLP-1R
2.4. Comparing Configurations of DdC, LSN3160440, Quercetin, HJ, and PA on GLP-1R
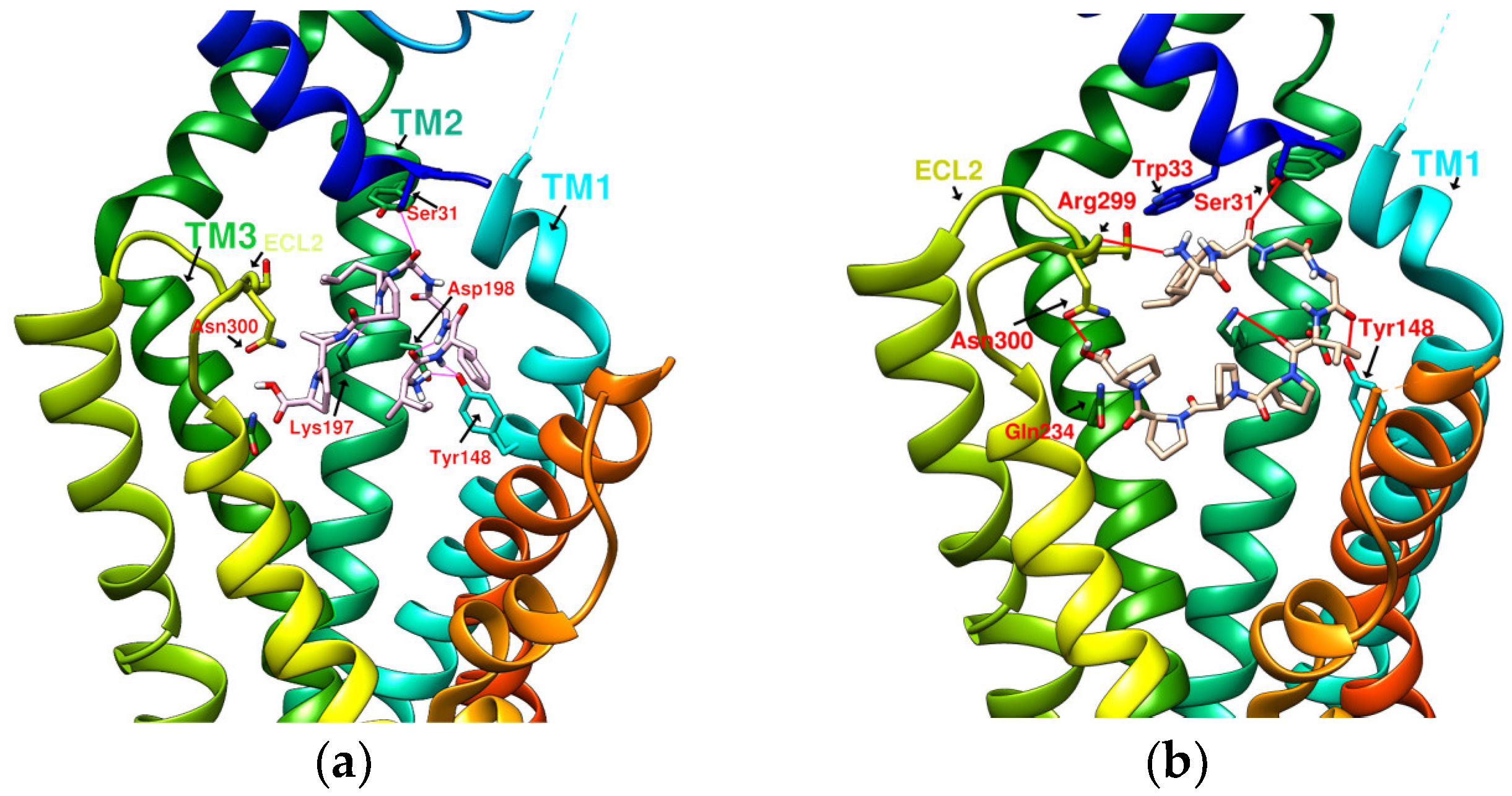
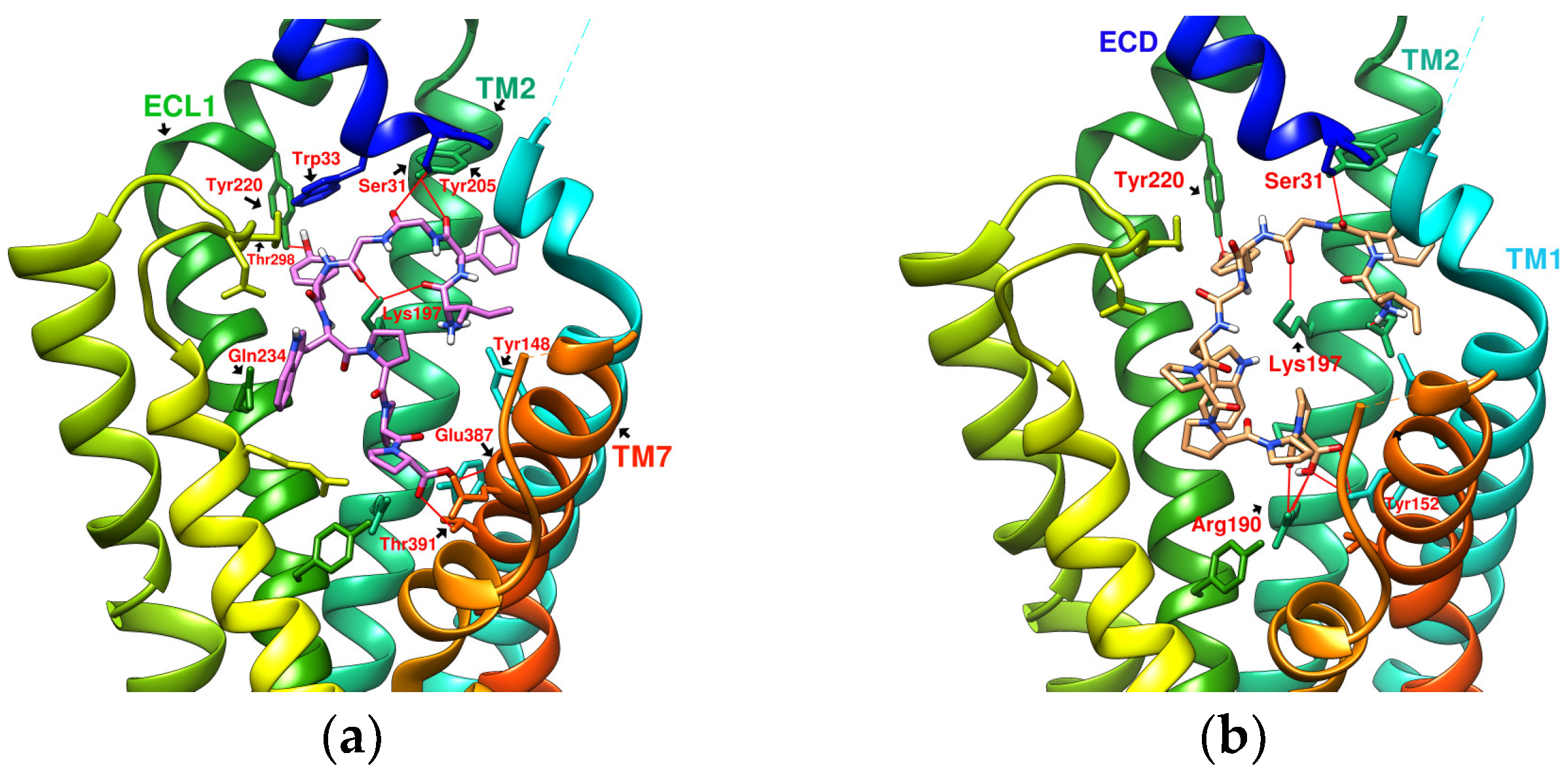
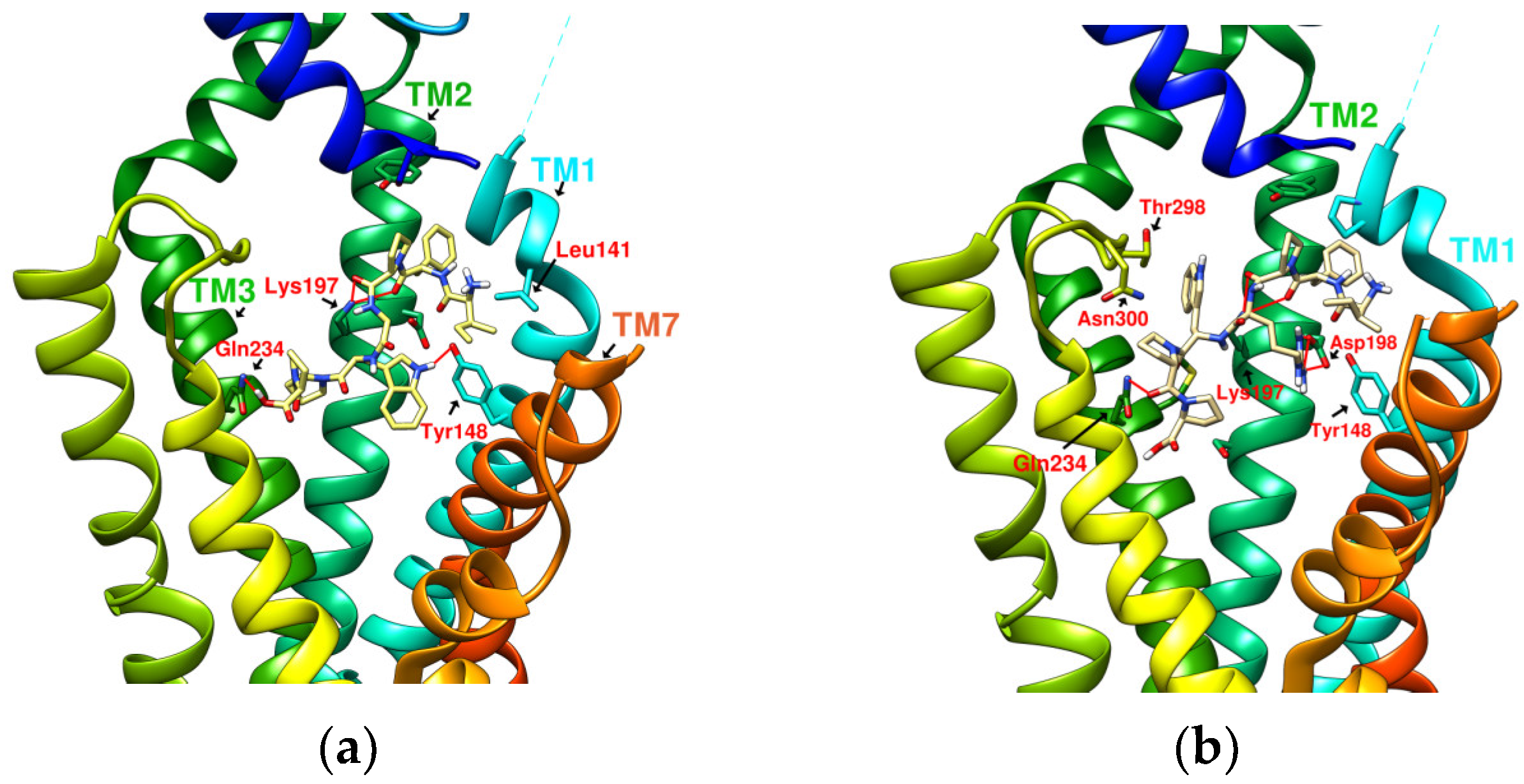
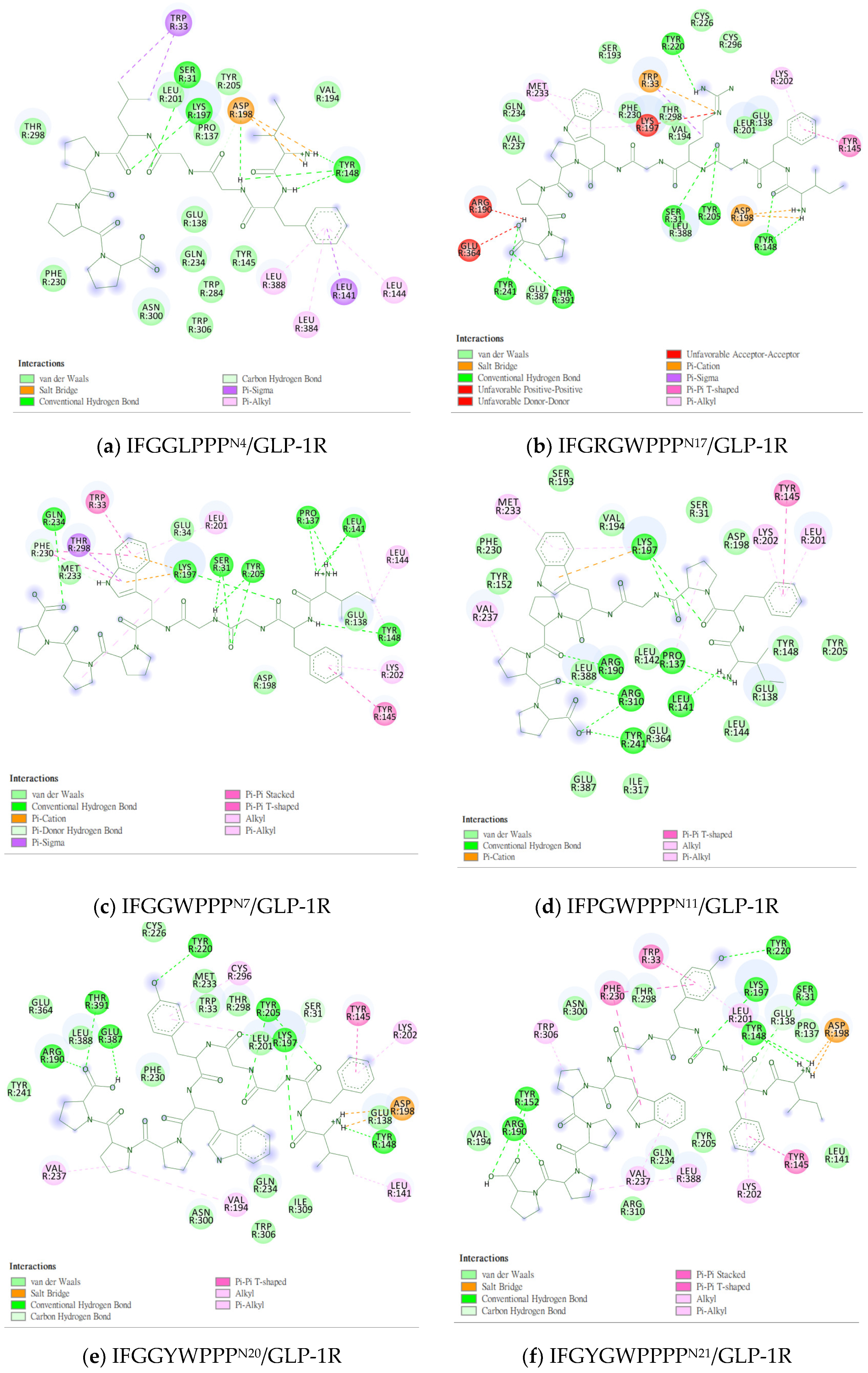
2.5. Prediction of Binding Affinity of IFGGLPPP Peptide Derivatives to GLP-1R
2.6. The Configuration of IFGGWPPP(N7) and IFPGWPPP(N11) in Docking with GLP-1R
2.7. The Configuration of IFGGYWPPP(N20) and IFGYGWPPPP(N21) in Docking with GLP-1R
2.8. Summary of the Interaction Mode between IFGGLPPP Derivatives and GLP-1R in Docking
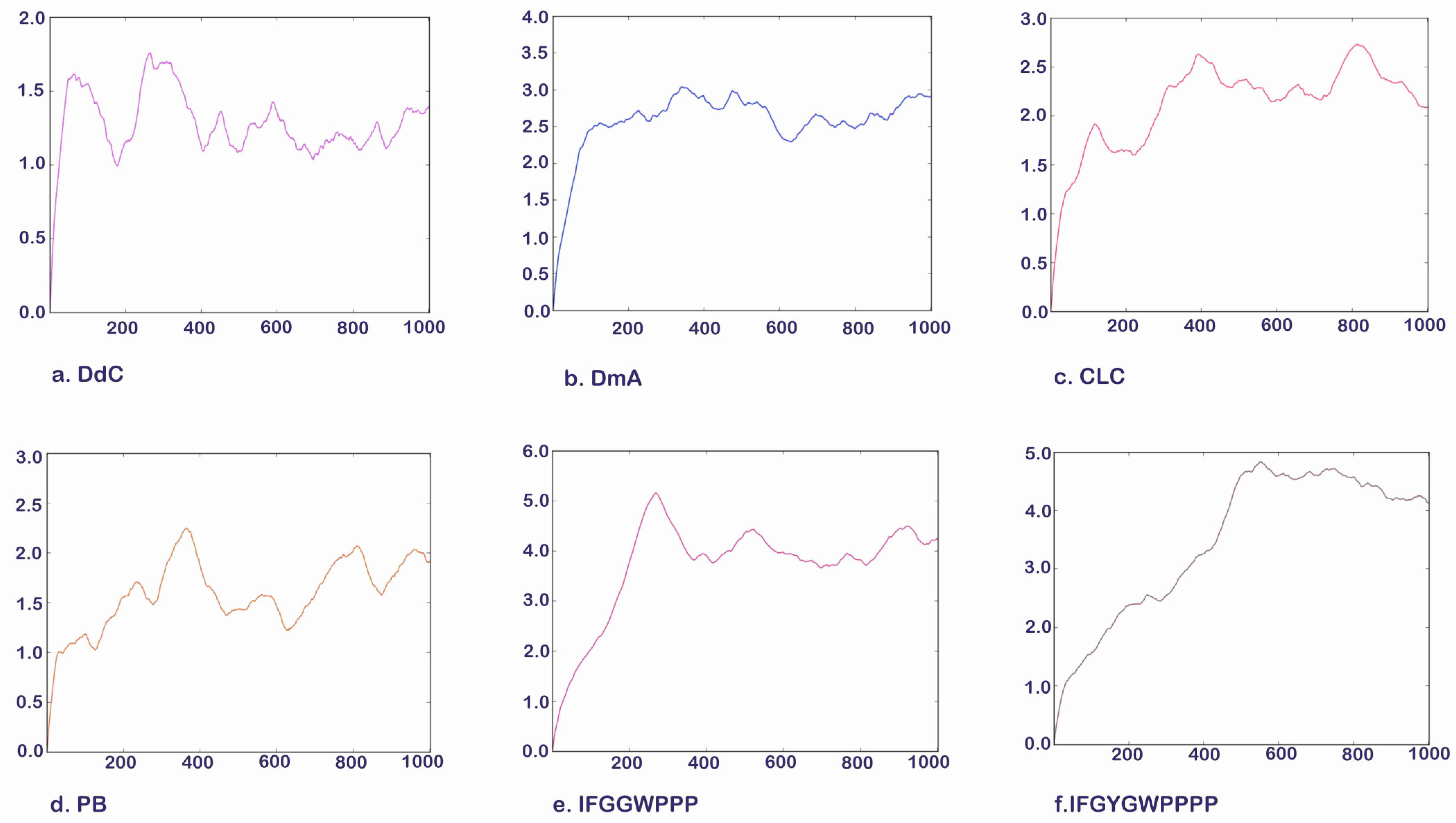
2.9. Molecular Dynamics Simulation of Potential Cyclic and Linear Peptides on GLP-1R
2.10. The Potential of D. diandra as a Nutritional Supplement for Blood Sugar and Weight Control
2.11. Effects of Cyclopeptides on DPP4 and GLP-1R, Two Targets of the Incretin System
2.12. Limitations of the Study
3. Materials and Methods
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed]
- Baggio, L.L.; Drucker, D.J. Biology of incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.D.; Finan, B.; Bloom, S.; D’Alessio, D.; Drucker, D.J.; Flatt, P.; Fritsche, A.; Gribble, F.; Grill, H.; Habener, J. Glucagon-like peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef] [PubMed]
- Godinho, R.; Mega, C.; Teixeira-de-Lemos, E.; Carvalho, E.; Teixeira, F.; Fernandes, R.; Reis, F. The Place of Dipeptidyl Peptidase-4 Inhibitors in Type 2 Diabetes Therapeutics: A “Me Too” or “the Special One” Antidiabetic Class? J. Diabetes Res. 2015, 2015, 806979. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J.; Bianchi, C.; Del Prato, S. Use of incretin-based medications: What do current international recommendations suggest with respect to GLP-1 receptor agonists and DPP-4 inhibitors? Metabolism 2020, 107, 154242. [Google Scholar] [CrossRef] [PubMed]
- Aroda, V.R.; Henry, R.R.; Han, J.; Huang, W.; DeYoung, M.B.; Darsow, T.; Hoogwerf, B.J. Efficacy of GLP-1 receptor agonists and DPP-4 inhibitors: Meta-analysis and systematic review. Clin. Ther. 2012, 34, 1247–1258.E22. [Google Scholar] [CrossRef]
- Jazayeri, A.; Rappas, M.; Brown, A.J.H.; Kean, J.; Errey, J.C.; Robertson, N.J.; Fiez-Vandal, C.; Andrews, S.P.; Congreve, M.; Bortolato, A.; et al. Crystal structure of the GLP-1 receptor bound to a peptide agonist. Nature 2017, 546, 254–258. [Google Scholar] [CrossRef]
- Sharma, D.; Verma, S.; Vaidya, S.; Kalia, K.; Tiwari, V. Recent updates on GLP-1 agonists: Current advancements & challenges. Biomed. Pharm. 2018, 108, 952–962. [Google Scholar] [CrossRef]
- Dailey, M.J.; Moran, T.H. Glucagon-like peptide 1 and appetite. Trends Endocrinol. Metab. 2013, 24, 85–91. [Google Scholar] [CrossRef]
- Nuffer, W.A.; Trujillo, J.M. Liraglutide: A New Option for the Treatment of Obesity. Pharmacotherapy 2015, 35, 926–934. [Google Scholar] [CrossRef]
- O’Neil, P.M.; Birkenfeld, A.L.; McGowan, B.; Mosenzon, O.; Pedersen, S.D.; Wharton, S.; Carson, C.G.; Jepsen, C.H.; Kabisch, M.; Wilding, J.P.H. Efficacy and safety of semaglutide compared with liraglutide and placebo for weight loss in patients with obesity: A randomised, double-blind, placebo and active controlled, dose-ranging, phase 2 trial. Lancet 2018, 392, 637–649. [Google Scholar] [CrossRef]
- Koska, J.; Sands, M.; Burciu, C.; D’Souza, K.M.; Raravikar, K.; Liu, J.; Truran, S.; Franco, D.A.; Schwartz, E.A.; Schwenke, D.C.; et al. Exenatide Protects Against Glucose- and Lipid-Induced Endothelial Dysfunction: Evidence for Direct Vasodilation Effect of GLP-1 Receptor Agonists in Humans. Diabetes 2015, 64, 2624–2635. [Google Scholar] [CrossRef] [PubMed]
- Sloan, L.A. Review of glucagon-like peptide-1 receptor agonists for the treatment of type 2 diabetes mellitus in patients with chronic kidney disease and their renal effects. J. Diabetes 2019, 11, 938–948. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Ratner, R.E.; Han, J.; Kim, D.D.; Fineman, M.S.; Baron, A.D. Effects of exenatide (exendin-4) on glycemic control and weight over 30 weeks in metformin-treated patients with type 2 diabetes. Diabetes Care 2005, 28, 1092–1100. [Google Scholar] [CrossRef]
- Knudsen, L.B.; Lau, J. The Discovery and Development of Liraglutide and Semaglutide. Front. Endocrinol. 2019, 10, 155. [Google Scholar] [CrossRef]
- Manandhar, B.; Ahn, J.M. Glucagon-like peptide-1 (GLP-1) analogs: Recent advances, new possibilities, and therapeutic implications. J. Med. Chem. 2015, 58, 1020–1037. [Google Scholar] [CrossRef]
- Zhang, X.; Belousoff, M.J.; Zhao, P.; Kooistra, A.J.; Truong, T.T.; Ang, S.Y.; Underwood, C.R.; Egebjerg, T.; Šenel, P.; Stewart, G.D.; et al. Differential GLP-1R Binding and Activation by Peptide and Non-peptide Agonists. Mol. Cell 2020, 80, 485–500.e487. [Google Scholar] [CrossRef]
- Coopman, K.; Wallis, R.; Robb, G.; Brown, A.J.; Wilkinson, G.F.; Timms, D.; Willars, G.B. Residues within the transmembrane domain of the glucagon-like peptide-1 receptor involved in ligand binding and receptor activation: Modelling the ligand-bound receptor. Mol. Endocrinol. 2011, 25, 1804–1818. [Google Scholar] [CrossRef][Green Version]
- Wootten, D.; Reynolds, C.A.; Smith, K.J.; Mobarec, J.C.; Koole, C.; Savage, E.E.; Pabreja, K.; Simms, J.; Sridhar, R.; Furness, S.G.B.; et al. The Extracellular Surface of the GLP-1 Receptor Is a Molecular Trigger for Biased Agonism. Cell 2016, 165, 1632–1643. [Google Scholar] [CrossRef]
- Yang, D.; de Graaf, C.; Yang, L.; Song, G.; Dai, A.; Cai, X.; Feng, Y.; Reedtz-Runge, S.; Hanson, M.A.; Yang, H.; et al. Structural Determinants of Binding the Seven-transmembrane Domain of the Glucagon-like Peptide-1 Receptor (GLP-1R). J. Biol. Chem. 2016, 291, 12991–13004. [Google Scholar] [CrossRef]
- Kawai, T.; Sun, B.; Yoshino, H.; Feng, D.; Suzuki, Y.; Fukazawa, M.; Nagao, S.; Wainscott, D.B.; Showalter, A.D.; Droz, B.A.; et al. Structural basis for GLP-1 receptor activation by LY3502970, an orally active nonpeptide agonist. Proc. Natl. Acad. Sci. USA 2020, 117, 29959–29967. [Google Scholar] [CrossRef] [PubMed]
- Bueno, A.B.; Sun, B.; Willard, F.S.; Feng, D.; Ho, J.D.; Wainscott, D.B.; Showalter, A.D.; Vieth, M.; Chen, Q.; Stutsman, C.; et al. Structural insights into probe-dependent positive allosterism of the GLP-1 receptor. Nat. Chem. Biol. 2020, 16, 1105–1110. [Google Scholar] [CrossRef] [PubMed]
- Cong, Z.; Chen, L.N.; Ma, H.; Zhou, Q.; Zou, X.; Ye, C.; Dai, A.; Liu, Q.; Huang, W.; Sun, X.; et al. Molecular insights into ago-allosteric modulation of the human glucagon-like peptide-1 receptor. Nat. Commun. 2021, 12, 3763. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Liang, Y.L.; Belousoff, M.J.; Deganutti, G.; Fletcher, M.M.; Willard, F.S.; Bell, M.G.; Christe, M.E.; Sloop, K.W.; Inoue, A.; et al. Activation of the GLP-1 receptor by a non-peptidic agonist. Nature 2020, 577, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Quoyer, J.; Longuet, C.; Broca, C.; Linck, N.; Costes, S.; Varin, E.; Bockaert, J.; Bertrand, G.; Dalle, S. GLP-1 mediates antiapoptotic effect by phosphorylating Bad through a beta-arrestin 1-mediated ERK1/2 activation in pancreatic beta-cells. J. Biol. Chem. 2010, 285, 1989–2002. [Google Scholar] [CrossRef] [PubMed]
- Swedberg, J.E.; Schroeder, C.I.; Mitchell, J.M.; Durek, T.; Fairlie, D.P.; Edmonds, D.J.; Griffith, D.A.; Ruggeri, R.B.; Derksen, D.R.; Loria, P.M. Cyclic alpha-conotoxin peptidomimetic chimeras as potent GLP-1R agonists. Eur. J. Med. Chem. 2015, 103, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Collet, T.H.; Dubern, B.; Mokrosinski, J.; Connors, H.; Keogh, J.M.; Mendes de Oliveira, E.; Henning, E.; Poitou-Bernert, C.; Oppert, J.M.; Tounian, P.; et al. Evaluation of a melanocortin-4 receptor (MC4R) agonist (Setmelanotide) in MC4R deficiency. Mol. Metab. 2017, 6, 1321–1329. [Google Scholar] [CrossRef]
- Yu, J.; Gimenez, L.E.; Hernandez, C.C.; Wu, Y.; Wein, A.H.; Han, G.W.; McClary, K.; Mittal, S.R.; Burdsall, K.; Stauch, B.; et al. Determination of the melanocortin-4 receptor structure identifies Ca(2+) as a cofactor for ligand binding. Science 2020, 368, 428–433. [Google Scholar] [CrossRef]
- Heyder, N.A.; Kleinau, G.; Speck, D.; Schmidt, A.; Paisdzior, S.; Szczepek, M.; Bauer, B.; Koch, A.; Gallandi, M.; Kwiatkowski, D.; et al. Structures of active melanocortin-4 receptor-Gs-protein complexes with NDP-α-MSH and setmelanotide. Cell Res. 2021, 31, 1176–1189. [Google Scholar] [CrossRef]
- Hu, D.J.; Shakerian, F.; Zhao, J.; Li, S.P. Chemistry, pharmacology and analysis of Pseudostellaria heterophylla: A mini-review. Chin. Med. 2019, 14, 21. [Google Scholar] [CrossRef]
- Su, K.; Zhu, F.; Guo, L.; Zhu, Y.; Li, W.; Xiong, X. Retrospective study on Professor Zhongying Zhou’s experience in Traditional Chinese Medicine treatment on diabetic nephropathy. J. Tradit. Chin. Med. 2013, 33, 262–267. [Google Scholar] [CrossRef]
- Chen, J.; Pang, W.; Shi, W.; Yang, B.; Kan, Y.; He, Z.; Hu, J. Structural Elucidation of a Novel Polysaccharide from Pseudostellaria heterophylla and Stimulating Glucose Uptake in Cells and Distributing in Rats by Oral. Molecules 2016, 21, 1233. [Google Scholar] [CrossRef] [PubMed]
- Mani, U.V.; Mani, I.; Biswas, M.; Kumar, S.N. An open-label study on the effect of flax seed powder (Linum usitatissimum) supplementation in the management of diabetes mellitus. J. Diet. Suppl. 2011, 8, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Pan, A.; Sun, J.; Chen, Y.; Ye, X.; Li, H.; Yu, Z.; Wang, Y.; Gu, W.; Zhang, X.; Chen, X.; et al. Effects of a flaxseed-derived lignan supplement in type 2 diabetic patients: A randomized, double-blind, cross-over trial. PLoS ONE 2007, 2, e1148. [Google Scholar] [CrossRef]
- Patra, S.; Bhattacharya, S.; Bala, A.; Haldar, P.K. Antidiabetic effect of Drymaria cordata leaf against streptozotocin-nicotinamide-induced diabetic albino rats. J. Adv. Pharm. Technol. Res. 2020, 11, 44–52. [Google Scholar] [CrossRef]
- Lu, F.; Yang, H.; Lin, S.D.; Zhao, L.; Jiang, C.; Chen, Z.B.; Liu, Y.Y.; Kan, Y.J.; Hu, J.; Pang, W.S. Cyclic Peptide Extracts Derived From Pseudostellaria heterophylla Ameliorates COPD via Regulation of the TLR4/MyD88 Pathway Proteins. Front. Pharmacol. 2020, 11, 850. [Google Scholar] [CrossRef]
- Shim, Y.Y.; Song, Z.; Jadhav, P.D.; Reaney, M.J. Orbitides from flaxseed (Linum usitatissimum L.): A comprehensive review. Trends Food Sci. Technol. 2019, 93, 197–211. [Google Scholar] [CrossRef]
- Hsieh, P.W.; Chang, F.R.; Wu, C.C.; Wu, K.Y.; Li, C.M.; Wang, W.Y.; Gu, L.C.; Wu, Y.C. Selective Inhibition of Collagen-Induced Platelet Aggregation by a Cyclic Peptide from Drymaria diandra. Helv. Chim. Acta 2004, 87, 57–66. [Google Scholar] [CrossRef]
- Hsieh, P.-W.; Chang, F.-R.; Lee, K.-H.; Hwang, T.-L.; Chang, S.-M.; Wu, Y.-C. A new anti-HIV alkaloid, drymaritin, and a new C-Glycoside flavonoid, diandraflavone, from Drymaria d iandra. J. Nat. Prod. 2004, 67, 1175–1177. [Google Scholar] [CrossRef]
- Tan, N.H.; Zhou, J. Plant cyclopeptides. Chem. Rev. 2006, 106, 840–895. [Google Scholar] [CrossRef]
- Arnison, P.G.; Bibb, M.J.; Bierbaum, G.; Bowers, A.A.; Bugni, T.S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: Overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2013, 30, 108–160. [Google Scholar] [CrossRef] [PubMed]
- He, J.-H.; Chen, L.-X.; Li, H. Progress in the discovery of naturally occurring anti-diabetic drugs and in the identification of their molecular targets. Fitoterapia 2019, 134, 270–289. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A. Incretin-based therapies for type 2 diabetes mellitus: Properties, functions, and clinical implications. Am. J. Med. 2011, 124, S3–S18. [Google Scholar] [CrossRef] [PubMed]
- Klemann, C.; Wagner, L.; Stephan, M.; von Hörsten, S. Cut to the chase: A review of CD26/dipeptidyl peptidase-4’s (DPP4) entanglement in the immune system. Clin. Exp. Immunol. 2016, 185, 1–21. [Google Scholar] [CrossRef]
- Liao, H.-J.; Tzen, J.T.C. The Potential Role of Cyclopeptides from Pseudostellaria heterophylla, Linum usitatissimum and Drymaria diandra, and Peptides Derived from Heterophyllin B as Dipeptidyl Peptidase IV Inhibitors for the Treatment of Type 2 Diabetes: An In Silico Study. Metabolites 2022, 12, 387. [Google Scholar] [CrossRef]
- Bockus, A.T.; McEwen, C.M.; Lokey, R.S. Form and function in cyclic peptide natural products: A pharmacokinetic perspective. Curr. Top. Med. Chem. 2013, 13, 821–836. [Google Scholar] [CrossRef]
- Ahlbach, C.L.; Lexa, K.W.; Bockus, A.T.; Chen, V.; Crews, P.; Jacobson, M.P.; Lokey, R.S. Beyond cyclosporine A: Conformation-dependent passive membrane permeabilities of cyclic peptide natural products. Future Med. Chem. 2015, 7, 2121–2130. [Google Scholar] [CrossRef]
- Räder, A.F.B.; Reichart, F.; Weinmüller, M.; Kessler, H. Improving oral bioavailability of cyclic peptides by N-methylation. Bioorg Med. Chem. 2018, 26, 2766–2773. [Google Scholar] [CrossRef]
- Dougherty, P.G.; Sahni, A.; Pei, D. Understanding Cell Penetration of Cyclic Peptides. Chem. Rev. 2019, 119, 10241–10287. [Google Scholar] [CrossRef]
- Wang, C.K.; Craik, D.J. Cyclic peptide oral bioavailability: Lessons from the past. Biopolymers 2016, 106, 901–909. [Google Scholar] [CrossRef]
- Zheng, W.; Zhou, T.; Li, J.; Jiang, W.; Zhang, J.; Xiao, C.; Wei, D.; Yang, C.; Xu, R.; Gong, A. The Biosynthesis of Heterophyllin B in Pseudostellaria heterophylla From prePhHB-Encoded Precursor. Front. Plant Sci. 2019, 10, 1259. [Google Scholar] [CrossRef] [PubMed]
- Koole, C.; Wootten, D.; Simms, J.; Valant, C.; Sridhar, R.; Woodman, O.L.; Miller, L.J.; Summers, R.J.; Christopoulos, A.; Sexton, P.M. Allosteric ligands of the glucagon-like peptide 1 receptor (GLP-1R) differentially modulate endogenous and exogenous peptide responses in a pathway-selective manner: Implications for drug screening. Mol. Pharmacol. 2010, 78, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Henninot, A.; Collins, J.C.; Nuss, J.M. The Current State of Peptide Drug Discovery: Back to the Future? J. Med. Chem. 2018, 61, 1382–1414. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, R.; Singh, S.; Varghese Gupta, S.; Sutariya, V.B.; Bhatia, D.; Mourya, R.; Chennupati, S.V.; Sharma, A. First Total Synthesis and Pharmacological Potential of a Plant Based Hexacyclopeptide. Iran. J. Pharm Res. 2019, 18, 938–947. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ma, L.; Yi, D.; Wang, H.; Wang, J.; Zhang, Y.; Guo, Y.; Li, X.; Zhou, J.; Shi, Y.; et al. Novel cyclo-peptides inhibit Ebola pseudotyped virus entry by targeting primed GP protein. Antivir. Res. 2018, 155, 1–11. [Google Scholar] [CrossRef]
- Kounnis, V.; Chondrogiannis, G.; Mantzaris, M.D.; Tzakos, A.G.; Fokas, D.; Papanikolaou, N.A.; Galani, V.; Sainis, I.; Briasoulis, E. Microcystin LR Shows Cytotoxic Activity Against Pancreatic Cancer Cells Expressing the Membrane OATP1B1 and OATP1B3 Transporters. Anticancer. Res. 2015, 35, 5857–5865. Available online: https://ar.iiarjournals.org/content/35/11/5857.long (accessed on 1 March 2022).
- Pang, S.; Zhao, R.; Wang, S.; Wang, J. Cyclopeptides design as blockers against HCV p7 channel in silico. Mol. Simul. 2019, 45, 1419–1425. [Google Scholar] [CrossRef]
- Wang, J.; Wang, W.; Kollman, P.A.; Case, D.A. Automatic atom type and bond type perception in molecular mechanical calculations. J. Mol. Graph. Model 2006, 25, 247–260. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Abbrev. | Molecular Name | Molecular Formula | Binding Affinity | MW |
|---|---|---|---|---|---|
| 1 | HA | Heterophyllin A | (cyclo)-PVIFGIT-(cyclo) | −8.7 | 727.9 |
| 2 | HB | Heterophyllin B | (cyclo)-GGLPPPIF-(cyclo) | −9.5 | 778.9 |
| 3 | HJ | Heterophyllin J | (cyclo)-AGPVY-(cyclo) | −9.7 | 487.5 |
| 4 | PA | Pseudostellarin A | (cyclo)-AGPYL-(cyclo) | −8.1 | 501.6 |
| 5 | PB | Pseudostellarin B | (cyclo)-GGGPPFGI-(cyclo) | −10.4 | 682.8 |
| 6 | PD | Pseudostellarin D | (cyclo)-GPLILGY-(cyclo) | −9.0 | 713.9 |
| 7 | PE | Pseudostellarin E | (cyclo)-GPPLGPVIF-(cyclo) | −9.8 | 878.1 |
| 8 | PH | Pseudostellarin H | (cyclo)-GTPTPLFF-(cyclo) | −10.3 | 861 |
| 9 | CLA | Cyclolinopeptide A | (cyclo)-ILLPPFFLV-(cyclo) | −9.9 | 1040.3 |
| 10 | CLB | Cyclolinopeptide B | (cyclo)-IMLIPPFFV-(cyclo) | −10.3 | 1058.4 |
| 11 | CLC | Cyclolinopeptide C | (cyclo)-IM(O)LIPPFFV-(cyclo) | −10.6 | 1074.4 |
| 12 | CLF | Cyclolinopeptide F | (cyclo)-LM(O)PFFWVM(O)-(cyclo) | −8.8 | 1084.4 |
| 13 | DmA | Drymarin A | (cyclo)-AFPPPFFVI-(cyclo) | −11.2 | 1016.2 |
| 14 | DmB | Drymarin B | (cyclo)-GLPFYP-(cyclo) | −9.8 | 674.8 |
| 15 | DdA | Diandrine A | (cyclo)-GPWPYF-(cyclo) | −9.5 | 747.8 |
| 16 | DdB | Diandrine B | (cyclo)-GPLPLWSS-(cyclo) | −8.8 | 838 |
| 17 | DdC | Diandrine C | (cyclo)-GGPYWP-(cyclo) | −11.9 | 657.7 |
| Compound | ECD | TM1 | TM2 | TM3 | TM7 | ECL1 | ECL2 | TM5 | TM6 |
|---|---|---|---|---|---|---|---|---|---|
| TT-OAD2 | Y145, Y148 | K197, L201 | Y220 | ||||||
| LY3502970 | W33 | Y148, Y145 | Y205 | L388 | Y220 | ||||
| PF-06882961 | S31 | L141 | K197 | F230 M233 | L384 L388 | Q221 | T298 | ||
| HB | W33π | Y148 | K197 | N300 T298 | |||||
| CLC | W33π | K197 | N300 | R310 | |||||
| DdC | E34π | P137, Y145π | K197 | ||||||
| PB | Y148 | R190 | Q234 | E364π | |||||
| DmA | Y148π | K197, D198 | R310π |
| N | Sequence | BA of GLP-1 | MW (g/mol) | H-bond, Attractive Charge and Salt Bridge between Linear Peptide and GLP-1R (Data from DS Visualizer) | π–π Interaction |
|---|---|---|---|---|---|
| 1 | PYWP | −9.1 | 561.644 | K197, L141, Y148 | - |
| 2 | GGPYWP | −9.2 | 675.748 | E34, M233, K197, D198, L141, Y148 | - |
| 3 | AFPPPFFVI | −9.0 | 1034.277 | N300, Q234, K197, R190 | - |
| 4 | IFGGLPPP | −8.9 | 796.973 | S31, K197, D198, Y148 | - |
| 5 | IFGGLPPPP | −9.6 | 894.091 | T298, Q234, S31, Y205, Y148 | W33, F230 |
| 6 | IFGWPPP | −10.3 | 812.975 | Q234, L141 | F230 |
| 7 | IFGGWPPP | −10.7 | 870.027 | Q234, S31, Y205, Y148, K197, P137, L141 | T298, W33, Y145 |
| 8 | IFGGWPFP | −10.2 | 920.087 | Q234, Y148 | F230 |
| 9 | IFGWWPPP | −10.2 | 999.190 | Q234, L141, Y148 | F230 |
| 10 | IFPGWPP | −10.4 | 812.975 | K197, P137, E138 | Y145 |
| 11 | IFPGWPPP | −11.0 | 910.093 | K197, R190, P137, L141, Y241, R310 | Y145 |
| 12 | IFPGWPYP | −10.8 | 976.153 | Y241, K197, R190, P137, L141, Y148 | Y145 |
| 13 | IFPRWPP | −10.4 | 912.112 | Q234, K197, D198, P137, L141, Y148 | W33, F230 |
| 14 | IFPRWPPP | −10.5 | 1009.230 | Y241, K197, R190, P137, L141, Y148 | F230, Y145 |
| 15 | IFPRWPYP | −11.1 | 1075.290 | N300, M233, K197, D198, P137, Y148 | F230, Y145 |
| 16 | IFGRWPPP | −10.0 | 969.164 | W33, D198, Y152 | Y145 |
| 17 | IFGRGWPPP | −10.7 | 1026.216 | Y220, Y241, S31, Y205, Y148, T391 | Y145 |
| 18 | IFPRGWPPP | −11.1 | 1066.282 | Y220, W33, Q234, Y241, D198, R190, Y148, T391 | Y145 |
| 19 | IFGGGWPPPP | −10.3 | 1024.197 | Y205, K197, Y148, E364, T391 | F230, Y145 |
| 20 | IFGGYWPPP | −10.9 | 1033.205 | Y220, Y205, K197, D198, R190, Y148, T391, E387 | Y145 |
| 21 | IFGYGWPPPP | −11.7 | 1130.323 | Y220, S31, K197, D198, R190, Y148, Y152 | W33, F230, Y145 |
| 22 | IFGGYWPPPP | −10.9 | 1130.323 | Y241, K197, R190, Y148, E364, T391 | F230, Y145 |
| Compound | DdC | DmA | CLC | PB | IFGGWPPP | IFGYGWPPPP |
|---|---|---|---|---|---|---|
| Number of atoms | 48 | 74 | 76 | 49 | 63 | 82 |
| Average RMSD (Å) over 1000 frames | 1.392 | 2.898 | 2.083 | 1.903 | 4.231 | 4.121 |
| Average potential energy | 736.384 | 1143.682 | 995.403 | 729.980 | 856.631 | 1066.311 |
| Average kinetic energy | 296.459 | 557.515 | 551.055 | 354.734 | 412.130 | 591.659 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, H.-J.; Tzen, J.T.C. Investigating Potential GLP-1 Receptor Agonists in Cyclopeptides from Pseudostellaria heterophylla, Linum usitatissimum, and Drymaria diandra, and Peptides Derived from Heterophyllin B for the Treatment of Type 2 Diabetes: An In Silico Study. Metabolites 2022, 12, 549. https://doi.org/10.3390/metabo12060549
Liao H-J, Tzen JTC. Investigating Potential GLP-1 Receptor Agonists in Cyclopeptides from Pseudostellaria heterophylla, Linum usitatissimum, and Drymaria diandra, and Peptides Derived from Heterophyllin B for the Treatment of Type 2 Diabetes: An In Silico Study. Metabolites. 2022; 12(6):549. https://doi.org/10.3390/metabo12060549
Chicago/Turabian StyleLiao, Hui-Jun, and Jason T. C. Tzen. 2022. "Investigating Potential GLP-1 Receptor Agonists in Cyclopeptides from Pseudostellaria heterophylla, Linum usitatissimum, and Drymaria diandra, and Peptides Derived from Heterophyllin B for the Treatment of Type 2 Diabetes: An In Silico Study" Metabolites 12, no. 6: 549. https://doi.org/10.3390/metabo12060549
APA StyleLiao, H.-J., & Tzen, J. T. C. (2022). Investigating Potential GLP-1 Receptor Agonists in Cyclopeptides from Pseudostellaria heterophylla, Linum usitatissimum, and Drymaria diandra, and Peptides Derived from Heterophyllin B for the Treatment of Type 2 Diabetes: An In Silico Study. Metabolites, 12(6), 549. https://doi.org/10.3390/metabo12060549