Abstract
Among the phospholipase A2 (PLA2) superfamily, the secreted PLA2 (sPLA2) family contains 11 mammalian isoforms that exhibit unique tissue or cellular distributions and enzymatic properties. Current studies using sPLA2-deficient or -overexpressed mouse strains, along with mass spectrometric lipidomics to determine sPLA2-driven lipid pathways, have revealed the diverse pathophysiological roles of sPLA2s in various biological events. In general, individual sPLA2s exert their specific functions within tissue microenvironments, where they are intrinsically expressed through hydrolysis of extracellular phospholipids. Recent studies have uncovered a new aspect of group IIA sPLA2 (sPLA2-IIA), a prototypic sPLA2 with the oldest research history among the mammalian PLA2s, as a modulator of the gut microbiota. In the intestine, Paneth cell-derived sPLA2-IIA acts as an antimicrobial protein to shape the gut microbiota, thereby secondarily affecting inflammation, allergy, and cancer in proximal and distal tissues. Knockout of intestinal sPLA2-IIA in BALB/c mice leads to alterations in skin cancer, psoriasis, and anaphylaxis, while overexpression of sPLA2-IIA in Pla2g2a-null C57BL/6 mice induces systemic inflammation and exacerbates arthritis. These phenotypes are associated with notable changes in gut microbiota and fecal metabolites, are variable in different animal facilities, and are abrogated after antibiotic treatment, co-housing, or fecal transfer. These studies open a new mechanistic action of this old sPLA2 and add the sPLA2 family to the growing list of endogenous factors capable of affecting the microbe–host interaction and thereby systemic homeostasis and diseases.
1. Introduction
The phospholipase A2 (PLA2) superfamily comprises a group of lipolytic enzymes that typically hydrolyze the sn-2 position of glycerophospholipids (hereafter phospholipids) to release free fatty acids (FFAs) and lysophospholipids (LPLs). The mammalian genome encodes more than 50 PLA2s or related enzymes, which are classified into several families on the basis of their structures and functions [1]. Historically, PLA2 enzymes have long attracted attention as a regulator of the production of lipid mediators (especially arachidonic acid (AA)-derived metabolites called eicosanoids), since they hydrolyze membrane phospholipids to release polyunsaturated fatty acids (PUFAs; including AA) and LPLs, which can act by themselves as bioactive lipids or act as precursors of a wide variety of lipid mediators. It is now obvious that PLA2s also participate in membrane homeostasis by regulating phospholipid composition, in energy homeostasis by fueling FFAs for β-oxidation, in microenvironmental regulation of the balance of saturated versus unsaturated FFAs, and so on. Moreover, several enzymes in the PLA2 superfamily have phospholipase A1, phospholipase B, lysophospholipase, triglyceride lipase, or acyltransferase/transacylase activity in addition to, or rather than, PLA2 activity. Thus, one should be careful that the term “PLA2” (or the gene name “PLA2 (in human)” or “Pla2” (in mouse)) does not always mean that a given PLA2 does act as a canonical PLA2 to liberate sn-2 FFAs and LPLs from phospholipids.
The secreted PLA2 (sPLA2) family involves structurally related, low-molecular-mass enzymes with a conserved His-Asp catalytic dyad, which ensures a strict Ca2+-dependent “PLA2” reaction. The mammalian genome encodes 11 sPLA2s (catalytically active IB, IIA, IIC, IID, IIE, IIF, III, V, X, and XIIA and inactive XIIB), which are classified structurally into group I/II/V/X, group III, and group XII branches [2]. Individual sPLA2s exhibit distinct tissue and cellular expression profiles and exert specific functions in lipid mediator-dependent or possibly lipid mediator-independent fashions. In general, individual sPLA2s exert their specific functions within tissue microenvironments where they are locally expressed. Since mammalian cells are relatively resistant to hydrolysis by mammalian sPLA2s, it has been proposed that sPLA2s act on the plasma membrane of activated, damaged, or dying cells, rather than that of resting cells in a paracrine manner [3,4,5]. In addition, non-cellular phospholipid components, such as dietary food, lipoproteins, lung surfactant, extracellular vesicles (EVs), and membranes of invading microorganisms such as bacteria and possibly fungi and parasites, act as bona fide hydrolytic targets of sPLA2s [6,7,8,9,10,11]. In certain situations, sPLA2-binding proteins such as PLA2R1 (sPLA2 receptor) modulate the functions of sPLA2s [12,13]. Various pathophysiological roles of individual sPLA2s, as demonstrated by studies employing sPLA2 gene-manipulated mice over the past decades, are summarized in our current reviews [14,15,16,17,18].
Historically, sPLA2-IB and -IIA are two classical (prototypic) sPLA2s originally identified by protein purification in the late 1980s. While sPLA2-IB is secreted from the pancreas into the intestinal lumen and acts as a digestive enzyme [19], sPLA2-IIA, initially purified from platelets and inflammatory fluids, is the only sPLA2 that is abundantly detected in the circulation of patients with inflammation or infection and has been considered to participate in systemic inflammation and antibacterial defense [20]. Recently, we and Boilard’s group have uncovered a novel aspect of sPLA2-IIA: this sPLA2 expressed in the intestinal Paneth cells contributes to the shaping of the gut microbiota, thereby secondarily affecting systemic events including immunity, allergy, and cancer in proximal and distal tissues. In this article, we summarize a new aspect of this old sPLA2 via the gut microbiota, thus providing an additional insight into the sPLA2 research.
2. A Long-Lasting Question: Does sPLA2-IIA Act as a Regulator of the Gut Microbiota?
The gut microbiota on the epithelial barriers comprises approximately 3 × 1013 microbial cells, whose balance can influence the physiological functions of the host [21,22,23,24]. Dysbiosis caused by various environmental factors, such as a high consumption of sugar, fat, or salt, a low consumption of dietary fiber, abuse of alcohol, or medication or antibiotics [25,26], as well as genetic factors such as mutations in host genes that are involved in the epithelial barrier or innate/adaptive immunity [27,28], leads to an increased prevalence of colitis, allergy, metabolic diseases, neurodegeneration, and cancer.
sPLA2-IIA is a prototypic sPLA2 that is highly induced in various human tissues during inflammation such as rheumatoid arthritis, sepsis, and COVID-19 infection [29,30,31]. It has been well documented that, in addition to its role as an “inflammatory sPLA2” that promotes sterile inflammation by mobilizing lipid mediators [32], sPLA2-IIA degrades bacterial membranes as a “bactericidal sPLA2”, thereby playing a protective role against bacterial infection [33] (Figure 1A,B). Because sPLA2-IIA shows high substrate selectivity toward phosphatidylethanolamine (PE), phosphatidylserine (PS), and phosphatidylglycerol (PG) over phosphatidylcholine (PC), phospholipids comprising the plasma membrane of quiescent mammalian cells, where PC is enriched in the outer leaflet, are relatively resistant to sPLA2-IIA-mediated hydrolysis [34,35]. When overexpressed or added exogenously at high concentrations, sPLA2-IIA is able to hydrolyze phospholipids in activated, damaged, or apoptotic cells, in which PE and PS are exposed on the outer plasma membrane [36,37]. Recently, it has become obvious that sPLA2-IIA, as well as several other sPLA2s, efficiently hydrolyzes phospholipids in EVs (microparticles and exosomes), which could explain the sPLA2-driven generation of lipid mediators during sterile inflammation or other biological events [10,11,32]. In the context of infection, bacterial membranes, which are rich in PE and PG, are superior hydrolytic targets of this enzyme. Indeed, by degrading bacterial membrane phospholipids, sPLA2-IIA efficiently kills Gram-positive bacteria, as well as Gram-negative bacteria in the presence of co-factors such as bacterial permeability-increasing protein (BPI) and lysozyme, at physiological concentrations [20,33,38]. Through its potent bactericidal activity, transgenic mice overexpressing human sPLA2-IIA (PLA2G2ATGN) on a C57BL/6 background are protected from infection with Gram-positive bacteria (e.g., Bacillus anthracis and Staphylococcus aureus) or Gram–negative bacteria (e.g., Escherichia coli and Helicobacter pylori) [39,40]. Thus, it has been thought that the primary biological role of sPLA2-IIA is to protect against bacterial infection (Figure 1B).

Figure 1.
Functions of sPLA2-IIA in sterile inflammation, anti-bacterial defense, and microbiota regulation: (A) In human rheumatoid arthritis, sPLA2-IIA induced in leukocytes and synoviocytes by inflammatory cytokines acts on phospholipids in EVs to release AA, which is further metabolized to eicosanoids such as 12-hydroxyeicosaenoic acid (HEPE) and leukotriene B4 (LTB4) to promote neutrophil recruitment and activation. sPLA2-IIA also releases mitochondrial DNA, one of the danger-associated molecular patterns (DAMPs), from the EVs. As such, sPLA2-IIA amplifies sterile inflammation as an “inflammatory sPLA2” [10,32]. (B) During bacterial infection, sPLA2-IIA induced in various cells (such as epithelial cells and leukocytes) by pathogen-associated molecular patterns (PAMPs) (such as peptidoglycan and lipopolysaccharide) kills invading bacteria by degrading bacterial membranes. As such, sPLA2-IIA plays a role in host defense against bacterial infection as a “bactericidal sPLA2” [20,33]. (C) In the intestinal lumen, sPLA2-IIA secreted from Paneth cells acts as an antimicrobial protein to shape the gut microbiota, thereby secondarily affecting host responses including cancer, psoriasis, allergy, and arthritis (see text).
Although sPLA2-IIA is expressed in various tissues (inflammatory and epithelial cells in particular) of humans and rats, its expression in mice is limited to the intestine (BALB/c and C3H) or not expressed at all (C57BL/6 and 129/Sv) due to a natural mutation [41,42], making the functional analysis of this enzyme using mouse models difficult. Nevertheless, regardless of animal species, sPLA2-IIA is commonly and abundantly expressed in intestinal Paneth cells, which secrete a cocktail of antimicrobial peptides such as defensins, cathelicidins, S100 proteins, RNases, and RegIII [43,44]. Antibiotics treatment decreases sPLA2-IIA expression in Paneth cells [45], while the colonization of germ-free C3H mice with conventionally housed mice increases its expression [46], suggesting that intestinal sPLA2-IIA expression is induced by gut microbiota. The amount of sPLA2-IIA in the small intestine of BALB/c mice is estimated to be ~2000 ng/mg tissue [47], a concentration that is sufficient enough to kill Gram-positive bacteria and even Gram-negative bacteria in the presence of other antimicrobial peptides. Because of these facts, it had been hypothesized that sPLA2-IIA secreted from Paneth cells into the intestinal lumen may function as a regulator of the gut microbiota, although solid experimental evidence for this hypothesis had been lacking. Recently, our and Boilard’s groups have independently addressed this long-standing question using gene-manipulated mice (knockout and transgenic) for this sPLA2 in a back-to-back way [48,49]. As detailed below, the shaping of the gut microbiota by intestinal sPLA2-IIA affects several pathologies in distal tissues, including skin cancer, psoriasis, allergy, and arthritis (Figure 1C).
3. Lessons from Pla2g2a−/− BALB/c Mice
Since the Pla2g2a gene is naturally disrupted in C57BL/6 and 129 strains due to a frameshift mutation as mentioned above [42], it had been difficult to assess the precise in vivo functions of endogenous sPLA2-IIA using a standard knockout strategy. Taking advantages of BALB/c mice, which have an intact Pla2g2a gene [42], we generated Pla2g2a−/− mice on this genetic background by backcrossing the mutated Pla2g2a allele in C57BL/6 mice onto a BALB/c background. Unlike the situation in humans and rats, where sPLA2-IIA is expressed or induced in many tissues, its expression in BALB/c mice is highly restricted to the intestine [43,50]. Beyond this species difference, the Pla2g2a−/− BALB/c strain appears to be a good animal model with which to analyze the physiological function of sPLA2-IIA endogenously expressed in the intestine. Using this new knockout mouse model, we have recently obtained compelling evidence that intestinal sPLA2-IIA is indeed involved in the modulation of gut microbiota, thereby secondarily affecting the pathology of distal skin.
We found that, despite the restricted expression of sPLA2-IIA in the intestine of BALB/c mice, its gene targeting unexpectedly resulted in notable skin phenotypes [48]. In a model of carcinogen-induced skin cancer, the multiplicity and incidence of skin cancer were markedly reduced, with alterations in the immune responses, in Pla2g2a−/− mice relative to Pla2g2a+/+ mice (note that the two genotypes were housed in separate cages; see below). In a model of psoriasis, the ectopic application of imiquimod elicited more severe ear edema, with a greater expression of several psoriasis markers, in Pla2g2a−/− mice than in Pla2g2a+/+ mice. Expression of sPLA2-IIA in the small intestine was decreased after treatment with broad spectrum antibiotics or housing in a germ-free facility, suggesting that its expression is induced by gut microbiota, most likely by bacterial-derived, pathogen-associated molecular patterns (PAMPs; e.g., peptideglycan and lipopolysaccharide) which are known to induce sPLA2-IIA expression through the NF-κB pathway [20]. Notably, although the abundance of fecal microbiota at the order and family levels was not different between Pla2g2a+/+ and Pla2g2a−/− mice, hierarchical clustering showed clear separation of the gut microbiota into two groups (Pla2g2a+/+ cluster and Pla2g2a−/− cluster) at the genus level. In fact, several bacteria including Gram-positive (Ruminococcaceae and Lachnospiraceae) and Gram-negative (Prevotellaceae and Helicobacteraceae) genera were distinctly present in Pla2g2a+/+ and Pla2g2a−/− mice. Importantly, the phenotypes of skin cancer and psoriasis in Pla2g2a−/− mice were absent when both genotypes were co-housed in the same cages (which resulted in mixing of the microbiota through coprophagia) or when these mice were housed in a more stringent pathogen-free facility, in which the expression of Pla2g2a in WT mice was reduced and the composition of gut microbiota in the two genotypes became largely even if not solely identical. Among several hit bacteria, as noted above, particular bacterial species belonging to the Helicobacter and Ruminococcaceae showed better correlation with the skin phenotypes in Pla2g2a−/− mice. Although the functional linkage between these specific bacteria and the skin phenotypes in Pla2g2a−/− mice needs further elucidation, these results provide insight into the notion that the regulation of gut microbiota by sPLA2-IIA is associated with the altered sensitivity to skin cancer and psoriasis.
Transcriptome analysis of the intestine, a proximal tissue where sPLA2-IIA is expressed, revealed notable alterations in the expression of genes related to epithelial barrier and immunity in Pla2g2a−/− mice in comparison with Pla2g2a+/+ mice. Particularly, a number of genes encoding the variable regions of immunoglobulins were markedly changed in Pla2g2a−/− mice compared to Pla2g2a+/+ mice, probably reflecting the distinct antibody responses to commensal microbiota between the genotypes. The reduced expression of several genes related to the anti-inflammatory PPARγ signaling pathway and the increased expression of several proinflammatory genes could partly explain the exacerbated psoriasis and the increased antitumor immunity in Pla2g−/− mice. Overall, the lack of sPLA2-IIA allows the intestine to remain in a mildly proinflammatory state, which appears to be consistent with the increased proportion of proinflammatory Helicobacter and the decreased proportion of anti-inflammatory Ruminococcaceae in Pla2g2−/− mice. While Helicobacter infection is tightly linked to gastric inflammation and cancer [51,52], there is ample evidence that it is protective against asthma and allergy [53,54]. The increase in Helicobacter in Pla2g2a−/− mice can explain why mouse strains with a mutated Pla2g2a gene are more susceptible to intestinal tumorigenesis than those having an intact Pla2g2a gene [42] and also accounts for an inverse correlation between PLA2G2A expression and gastric cancer in humans [55]. Moreover, Helicobacter infection and gastrointestinal inflammation are associated with psoriasis severity [56]. Thus, the effect of sPLA2-IIA on Helicobacter might be a key determinant that affects disease susceptibility in proximal (intestine) and distal (skin) tissues. In support of this, Pla2g2a−/− mice housed in the Helicobacter-free animal facility did not display a psoriasis phenotype.
Metabolome analysis revealed that 19 out of 511 plasma metabolites were significantly altered in Pla2g2a−/− mice compared with Pla2g2a+/+ mice (Figure 2A). These metabolites were classified into several groups such as those related to the urea cycle, reactive oxygen species (ROS), and choline metabolism. The urea cycle is associated with T cell immunity [57], and its dysregulation leads to tumor promotion [58]. Plasma levels of various metabolites in the urea cycle were reduced in Pla2g2a−/− mice compared to Pla2g2a+/+ mice (Figure 2A). Increased production of ROS is often associated with inflammation, metabolic diseases, cancer, and aging [59,60], and the Pla2g2a−/− plasma contained reduced levels of several ROS-related metabolites (Figure 2A). Furthermore, plasma levels of choline-related metabolites, whose unusual accumulation often causes cellular transformation [61], were reduced in Pla2g2a−/− mice (Figure 2A). In addition, several bacteria-specific metabolites such as trigonelline and ectoine, which can affect inflammation and cancer [62], and dicarboxylic acids such as pimelate, sebacate, and azelate, whose levels are associated with the abundance of Ruminococcaceae [63], were reduced in Pla2g2a−/− mice relative to Pla2g2a+/+ mice (Figure 2A). Furthermore, the lipidomic profiling of fecal lipids showed notable reductions in several bacteria-specific lipids with anti-inflammatory potential in Pla2g2a−/− mice (Figure 2B,C). These bacteria-derived lipids included a class of hydroxyl-, oxo-, and conjugated forms of linoleic acid (LA) metabolites, such as KetoB (10-oxo-octadecanoic acid), KetoC (10-oxo-trans-11-octadecenoic acid), and CLA1/3 (cis-9, trans-11- and trans-9, trans-11-octadecadienoic acids), and that of branched or linear fatty acid esters of hydroxy fatty acids (FAHFAs and OAHFAs). Reportedly, long-chain FAHFAs act on the fatty acid receptors GPR40 and GPR120 to exert anti-inflammatory, -oxidant, and -diabetic functions [64,65], and short-chain FAHFAs with acyl α-hydroxy fatty acids (AAHFAs) show inverse correlation with metabolic disease [66]. Although bacteria species generating these unique lipids are currently unknown, the decrease in these anti-inflammatory bacterial lipids may account, at least in part, for the increased antitumor immunity and exacerbated psoriasis in Pla2g2a−/− mice. On the other hand, host-derived lipid mediators, mostly produced by fatty acid oxygenation by lipoxygenases or cytochrome P450s, were unchanged or modestly elevated (rather than decreased) in Pla2g2a−/− mice than in Pla2g2a+/+ mice, probably because of the altered inflammatory state in the gut. Collectively, sPLA2-IIA deficiency alters the circulating and fecal levels of various hydrophilic and hydrophobic metabolites that could affect cancer and immunity. Because of the alterations of multiple bacteria and metabolites, it is presently difficult to conclusively define the specific bacteria species and metabolite(s) that would be truly responsible for the skin phenotypes in Pla2g2a−/− mice. It is likely that the combined actions of these multiple bacteria and metabolites on host immunity and metabolism may underlie the skin cancer and psoriasis phenotypes by the absence of sPLA2-IIA.
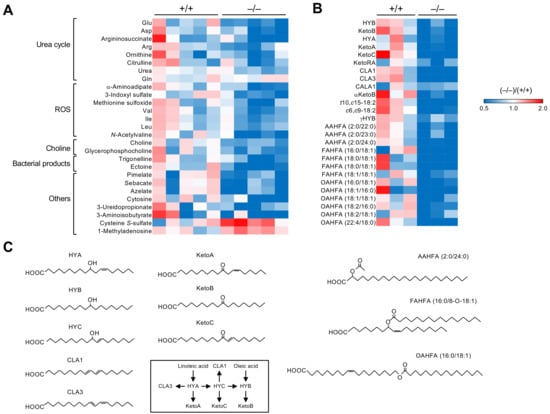
Figure 2.
Alterations in the levels of plasma and fecal metabolites in Pla2g2a−/− BALB/c mice. The heatmaps indicate fold changes of hydrophilic metabolites in plasma (A) and bacterial lipids in stools (B) of individual Pla2g2a−/− mice compared to Pla2g2a+/+ mice. (C) Structures of representative bacterial lipids listed in (B). Biosynthetic routes of hydroxy-, oxo-, and conjugated linoleic acid metabolites are also shown. The mice were housed in a pathogen-free facility (TMIMS) with a 12 h light–dark cycle at 23 °C, with free access to water and food, and all animal experiments were carried out according to protocols approved by the Institutional Animal Care and Use Committees under the Japanese Guide for the Care and Use of Laboratory Animals [48].
Allergic responses are known to be profoundly affected by the gut microbiota [67,68,69]. Therefore, we examined the impact of Pla2g2a deficiency on passive cutaneous anaphylaxis (PCA), an immediate-type allergic reaction that depends on mast cell degranulation. To this end, Pla2g2a+/+ and Pla2g2a−/− mice co-housed (+) or not co-housed (−) in two different animal facilities (TMIMS and UTokyo) were sensitized intradermally with anti-dinitrophenyl (DNP) IgE and then challenged intravenously with DNP-conjugated albumin as an antigen (Ag) together with Evans blue dye. Strikingly, under the co-housing (−) conditions, IgE/Ag-induced PCA reaction was lower at the TMIMS facility (Figure 3A,B), while it was conversely greater at the UTokyo facility, in Pla2g2a−/− mice than in Pla2g2a+/+ mice (Figure 3C,D). In agreement, IgE/Ag-treated and even IgE/Ag-untreated Pla2g2a−/− mice had more degranulated mast cells than the replicate Pla2g2a+/+ mice in the latter facility, although the total mast cell number was unaffected (Figure 3E,F). In both facilities, however, co-housed Pla2g2a+/+ and Pla2g2a−/− mice exhibited a comparable PCA response (Figure 3G,H). Thus, the allergic response in Pla2g2a−/− mice is greatly influenced by housing conditions, implying again the involvement of the gut microbiota. Metagenome analysis of the stool revealed that various bacteria genera were commonly increased (Figure 4A) or decreased (Figure 4B) in Pla2g2a−/− mice relative to Pla2g2a+/+ mice in both TMIMS and UTokyo facilities. Notably, there were also several bacteria genera whose abundance showed reciprocal patterns (i.e., increased or decreased in one facility and vice versa in the other facility) (Figure 4C,D), suggesting that one or more of these bacteria might be responsible for the contrasting PCA phenotypes between the two facilities (Figure 3). In particular, Lachnospiraceae FCS020 group and UCG-008, as well as Ruminococcaceae UCG-013, were more abundant in the TMIMS facility than in the UTokyo facility; moreover, in the TMIMS facility, Lachnospiraceae FCS020 group and Ruminococcaceae UCG-013 were decreased, while Lachnospiraceae UCG-008 was increased, in Pla2g2a−/− mice relative to Pla2g2a+/+ mice (Figure 4E). Reportedly, the abundance of Fimicutes (including Ruminococcaceae and Lachnospiraceae) can be positively or negatively associated with the severity of allergic diseases [70,71,72,73]. In contrast, Mucispirillum, a bacteria genus positively correlated with Th2-related factors in asthma [74], was uniquely abundant only in Pla2g2−/− mice at the UTokyo facility (Figure 4E), which might be related to the elevated PCA response (Figure 3C,D). Although further studies are needed to clarify whether the changes in these specific bacteria would be indeed responsible for the variable PCA phenotypes in Pla2g2a−/− mice, these results nonetheless lend additional support to the view that sPLA2-IIA shapes the gut microbiota, thereby secondarily affecting mast cell fate and the associated allergic reaction in distal skin and probably in other tissues.

Figure 3.
Altered PCA responses in Pla2g2a−/− BALB/c mice under distinct housing conditions. Ears of Pla2g2a+/+ and Pla2g2a−/− mice (8–12 weeks of age, male) were sensitized by subcutaneous injection of anti-DNP IgE monoclonal antibody (30 ng) and then challenged by intravenous injection of a mixture of DNP-conjugated human serum albumin (60 µg) as an antigen (Ag) together with 1 mg Evans blue, as described previously [75]: (A–D) Representative photos of the ears (A,C) and quantification of dye extravasation (B,D) in IgE/Ag-treated (+) or -untreated (−) Pla2g2a+/+ and PLa2g2a−/− mice housed in the TMIMS (A,B) and UTokyo (C,D) facilities. (E,F) Histology of the skin (E) and quantification of total and degranulated mast cells (F) in IgE/Ag-treated or -untreated Pla2g2a+/+ and Pla2g2a−/− mice housed in the UTokyo facility. The ear pinnae were fixed with 10% (v/v) formalin, embedded in paraffin, sectioned (4-µm thickness), and stained with toluidine blue. A total of 55 views for each group (n = 5). Black and red arrows indicate non-degranulated and degranulated mast cells, respectively. Scale bar, 25 µm. (G,H) IgE/Ag-induced PCA reaction in Pla2g2a+/+ and Pla2g2a−/− mice with (+) or without (−) co-housing in the TMIMS (G) and UTokyo (H) facilities. Values are mean ± SEM. *, p < 0.05; **, p < 0.01; NS, not significant. Statistical analysis was performed using Graph Pad PRISM with Brown–Forsythe test and then Kruskal–Wallis and Dunn’s post hoc test (B,D,F,G,H). The numbers of mice used for the analysis are indicated in each panel.
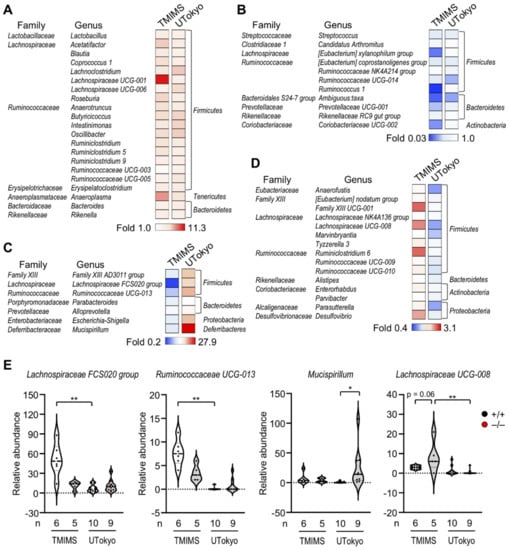
Figure 4.
The effects of sPLA2-IIA deficiency on the gut microbiota in the TMIMS and UTokyo animal facilities. Metagenome analysis of bacterial 16S RNA in the stool was performed as described previously [48]: (A,B) Heatmap representation of bacterial genera commonly increased (A) or decreased (B) in Pla2g2a−/− mice relative to Pla2g2a+/+ mice in both facilities. (C,D) Heatmap representation of bacterial genera that were increased in the UTokyo facility and decreased in the TMIMS facility (C) and vice versa (D) in Pla2g2a−/− mice relative to Pla2g2a+/+ mice. (E) Violin plots of the abundance of representative bacteria in Pla2g2a+/+ and Pla2g2a−/− mice. Values are mean ± SEM. *, p < 0.05; **, p < 0.01. Statistical analysis was performed with one-way ANOVA followed by Šidák multiple comparison test, or Kruskal–Wallis test followed by Dunn’s multiple comparison test (E). The numbers of mice used for the analysis are indicated in each panel.
4. Lessons from PLA2G2ATGN C57BL/6 Mice
Along with the study using Pla2g2a−/− BALB/c mice as described above, Boilard and colleagues performed a complementary study using PLA2G2ATGN mice in which the human PLA2G2GA gene was overexpressed in Pla2g2a-null (naturally mutated) C57BL/6 mice [49]. Generally speaking, the results using transgenic mice should be interpreted with caution, as the expression of a given transgene at a super-physiological level, even in tissues or cells where the enzyme is not intrinsically expressed, could result in an artificial phenotype. An advantage of the PLA2G2ATGN strain is that the transgene contains a part of the promoter region of human PLA2G2A gene, which could partially allow the expression profile of sPLA2-IIA similar, even if not identical, to that in humans. Although the levels of sPLA2-IIA detected in the blood [250 to 2300 ng/mL (up to 8700 ng/mL)] of PLA2G2ATGN mice are much greater than those measured in healthy individuals [76], they are comparable to those present in patients with severe bacterial infection or sepsis [77,78]. Hence, the use of PLA2G2ATGN mice has permitted an insight into pathophysiological manifestations that might be relevant to human diseases.
Boilard and colleagues found that aged PLA2G2ATGN mice spontaneously developed systemic inflammation, with enlargement of the spleen and lymph nodes, increased counts of T and B cells, plasmablasts and granulocytes, and elevated levels of serum IgG and IgA [49]. These events were similarly observed in PLA2G2ATGN mice lacking Pla2g4a or Alox12, ruling out the involvement of cytosolic PLA2 (cPLA2α) and/or 12-lipoxygenase that potentially act downstream of sPLA2-IIA in the context of lipid mediator signaling [28]. However, these phenotypes became milder when PLA2G2ATGN mice were housed in a different facility with a more stringent pathogen-free standard, suggesting the involvement of the gut microbiota. Indeed, a whole-genome shotgun sequencing of the fecal flora distinguished the PLA2G2ATGN and WT microbiota as a result of sPLA2-IIA overexpression. In particular, Odoribacter, Prevotell, and Helicobacter were more abundant in PLA2G2ATGN mice, while Eubacterium, Lachnoclostridium, and Clostridium were more abundant in Pla2g2a-mutated WT mice. An overall trend is that Gram-negative genera were more abundant in PLA2G2ATGN mice and Gram-positive genera were more abundant in WT mice, consistent with the view that sPLA2-IIA preferentially kills Gram-positive bacteria. Since the increases in Helicobacter and Prevotella are often associated with inflammatory processes [79,80,81,82], these changes in the gut microbiota may explain, at least in part, the increased systemic inflammation in PLA2G2ATGN mice.
The gut microbiota can also influence the severity of rheumatoid arthritis [83]. sPLA2-IIA is highly induced in rheumatoid arthritis [29,84], and PLA2G2ATGN mice have higher susceptibility to the K/B × N serum-induced arthritis model [29,85,86]. Beyond the lipid mediator-dependent, proinflammatory action of sPLA2-IIA within the arthritic joint (Figure 1A), Boilard and colleagues found that the changes in gut microbiota significantly affected arthritis sensitivity in PLA2G2ATGN mice. While the depletion of the gut microflora using broad spectrum antibiotics failed to affect the development of arthritis in WT mice, it abrogated the increased sensitivity of PLA2G2ATGN mice to the arthritis. The concentration of sPLA2-IIA in the serum of PLA2G2ATGN mice was markedly increased in arthritic mice, whereas the microbiota depletion cancelled this event. Furthermore, while the fecal transfer from PLA2G2ATGN mice did not affect the severity of arthritis in WT mice, that from WT mice into PLA2G2ATGN mice reduced the severity, suggesting that the exacerbation of arthritis in PLA2G2ATGN mice depends largely on the changes in the gut microbiota and that some metabolites present in the WT microbiota could be protective against arthritis. Metagenome analysis of fecal microbiota under the arthritis conditions showed that Muribaculum might be associated with the arthritic phenotype of PLA2G2ATGN mice.
Although there was a limited impact of sPLA2-IIA on the gut eicosanoid profile of the host, untargeted lipidomics revealed that sPLA2-IIA expression led to significant alterations in the fecal lipid profile. Among the lipid species detected thus far, cholesteryl ester, ether-linked diacylglycerol, and triacylglycerol, as well as multiple LPLs and phospholipids, were elevated in both arthritic and non-arthritic PLA2G2ATGN mice. The increase in the concentration of total FFAs and LPLs was evident in both arthritic and non-arthritic PLA2G2ATGN mice, which was mitigated upon the depletion of the microbiota by antibiotics. Using machine learning, lipid metabolites belonging to the diacylglycerol, triacylglycerol, FFA, and ceramide families were found to be associated with sPLA2-IIA expression. Overall, these data suggest that sPLA2-IIA offers systemic effects on the immune system through its activity on the microbiota and thereby lipid metabolites.
5. Summary and Future Prospects
In this article, we have made an overview of the new role of an old sPLA2, sPLA2-IIA, as a regulator of gut microbiota, as revealed by its gene targeting and transgenic overexpression recently performed by our and Boilard’s groups, respectively, in a back-to-back methodology [44]. Since sPLA2-IIA has the ability to degrade bacterial membranes at physiological concentrations [20,33,38], and since it is abundantly expressed in intestinal Paneth cells that secrete various antimicrobial peptides [43,44], it had long been speculated that this bactericidal sPLA2 might play a role in controlling gut microbiota, although there had been no solid experimental evidence using proper animal models. It is now evident that several phenotypes in Pla2g2a−/− and PLA2G2ATGN mice are profoundly influenced by forcible manipulations (e.g., co-housing, fecal transfer, and antibiotics) of the gut microbiota. The results obtained from the studies by us and Boilard’s group are not necessarily complementary, since they were carried out using different disease models in knockout versus transgenic mice on different genetic backgrounds (BALB/c versus C57BL/6, respectively) using different approaches under different housing conditions in different facilities [44]. Nonetheless, both studies have reached the same conclusion that intestinal sPLA2-IIA acts as a host factor that contributes to the shaping of the gut microbiota, whose perturbation by knockout or overexpression leads to systemic effects. To further strengthen this concept, it would be important to identify which bacterial species and/or metabolites altered by sPLA2-IIA deficiency or overexpression are responsible for the disease phenotypes in the future study.
Intestinal expression of sPLA2-IIA is markedly reduced by treatment with antibiotics or by housing in a germ-free facility, indicating a feed-forward cycle of the sPLA2-IIA–microbiota interaction, where some bacteria-derived PAMPs induce the expression of sPLA2-IIA, which then shapes the microbial community and thus affects disease severity in distal tissues. It should be noted that the impact of sPLA2-IIA on gut microbiota cannot be simply explained by its ability to directly kill bacteria only, since only limited bacteria genera were changed, while the abundance of most bacteria were barely affected by its deficiency or overexpression. It is thus likely that the alteration of gut microbiota in Pla2g2a−/− or PLA2G2ATGN mice may also rely on complex interactions between different bacterial species and between bacteria and host. It is conceivable that sPLA2-IIA may increase certain bacteria through eradicating competing commensals. Indeed, in the lung of cystic fibrosis patients, Gram-negative Pseudomonas aeruginosa (sPLA2-IIA-resistant) increases the expression of sPLA2-IIA, which in turn kills Gram-positive Staphylococcus aureus (sPLA2-IIA-sensitive), allowing the former bacterium to be dominant [87]. Alternatively, indirect actions of sPLA2-IIA on the microbiota via modulation of the host immunity involving the generation of certain lipid mediators, or via the PLA2R1-driven mechanism, should also be taken into account, even though the overall changes in host lipid mediators in the stool of Pla2g2a−/− or PLA2G2ATGN mice were only modest [48,49]. Using PLA2G2ATGN mice, Schewe et al. reported that sPLA2-IIA modifies the YAP-dependent differentiation of intestinal stem cells through the pathway involving the interaction of sPLA2-IIA with PLA2R1 [44].
Since several sPLA2 isoforms are expressed in the gastrointestinal tract [16,88,89], they may play a general role as a regulator of gut microbiome. Indeed, knockout of pancreatic sPLA2-IB, a digestive sPLA2 that degrades dietary and biliary phospholipids in the intestinal lumen [19], impairs the defense against parasite infection in association with the change in gut microbiota [90], although it remains unclear whether this event would be related to the improved atherosclerotic and metabolic phenotypes in Pla2g1b−/− mice [6,91,92]. sPLA2-V secreted from macrophages increases phagocytosis and promotes bacterial or fungal clearance [93,94]. Therefore, sPLA2-V might also contribute to the clearance of some bacteria or fungi in the gut. Since the deficiency of sPLA2-IID, an immunosuppressive sPLA2 that is expressed in dendritic cells and M2 macrophages, increases adaptive immune responses against coronavirus infection [95], this sPLA2 might also be associated with the intestinal antimicrobial immunity. sPLA2-X is another sPLA2 that is abundantly expressed in the intestine, especially in colonic epithelial cells and goblet cells [88,96]. Although Pla2g10−/− mice independently generated and analyzed by different laboratories have been reported to display inflammatory, cardiovascular, and metabolic phenotypes, the results are not necessarily consistent among the studies; for instance, Pla2g10−/− mice have greater [88] and lower [97] inflammation, increased [98] and decreased [96] body weight, and exacerbated [99] and ameliorated [100] atherosclerosis. Hence, we speculate that the variable phenotypes in Pla2g10−/− mice observed in current studies might also involve the action of sPLA2-X on the gut microbiota. Furthermore, this action mode might be applicable not only to the microbiota in the gut, but also to those in other anatomical sites such as the skin. For instance, sPLA2-IIE and -IIF, which are highly expressed in the hair follicle and epidermis, respectively [101,102], might affect skin homeostasis partly by modulating the skin microbiota.
Taken together, the sPLA2/microbiota axis described here has opened a new insight into the mechanistic actions of the sPLA2 family. We speculate that some of the functions of sPLA2s that have been reported to date might be mediated, at least in part, through their actions on the gut microbiota in either a direct (i.e., sPLA2 kills bacteria) or indirect (i.e., sPLA2 regulates host immunity that in turn affects the microbiota) way. Translation of these findings using mouse models to human pathology is not so simple, since, besides the species difference in sPLA2 expression profiles, we need to consider the tissue-intrinsic effects of sPLA2s and the extrinsic effects of the gut sPLA2/microbiota axis, both of which can be diversely affected by environmental factors. Nonetheless, the findings reported herein have raised the possibility that the increased levels of sPLA2-IIA or possibly other sPLA2s in human stool could have a predictive value for several diseases. Moreover, oral application of sPLA2 inhibitors could be a potential therapy to treat or prevent allergy, arthritis, and cancer. In this regard, oral application of methyl indoxam, a pan-sPLA2 inhibitor, efficiently suppressed diet-induced obesity and glucose intolerance in mice [103]. In the context of this review, it is possible that this beneficial effect of the sPLA2 inhibitor on systemic metabolism may rely partly on the suppression of the sPLA2–microbiota interaction in the gastrointestinal tract, although other mechanisms, such as the inhibition of dietary phospholipid digestion by digestive sPLA2s [6,7] or lipoprotein modification by metabolic sPLA2s [8], should also be considered.
Author Contributions
M.M. wrote the manuscript. Figures were prepared by Y.T. and Y.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Grants-in-Aid for Scientific Research JP19K22483 and JP20H05691 (to M.M.), JP19K07042 (to Y.M.), and JP18K06624 (to Y.T.) from Japan Society for the Promotion of Science, and AMED-CREST JP21gm1210013 from the Japan Agency for Medical Research and Development (to M.M.).
Acknowledgments
We thank Tomoyoshi Soga (Keio University, Yamagata, Japan), Makoto Arita (RIKEN, Yokohama, Japan) and Jun Kunisawa (National Institutes of Biomedical Innovation, Health and Nutrition, Osaka, Japan) for metabolome, lipidome and microbiome analyses, respectively. We thank Eric Boilard (Centre de Recherche du CHU de Québec-Université Laval, Québec, Canada) for helpful discussions on the role of sPLA2-IIA in the regulation of gut microbiota.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Murakami, M. Novel functions of phospholipase A2s: Overview. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 763–765. [Google Scholar] [CrossRef] [PubMed]
- Lambeau, G.; Gelb, M.H. Biochemistry and physiology of mammalian secreted phospholipases A2. Annu. Rev. Biochem. 2008, 77, 495–520. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Kambe, T.; Shimbara, S.; Kudo, I. Functional coupling between various phospholipase A2s and cyclooxygenases in immediate and delayed prostanoid biosynthetic pathways. J. Biol. Chem. 1999, 274, 3103–3115. [Google Scholar] [CrossRef] [PubMed]
- Bezzine, S.; Koduri, R.S.; Valentin, E.; Murakami, M.; Kudo, I.; Ghomashchi, F.; Sadilek, M.; Lambeau, G.; Gelb, M.H. Exogenously added human group X secreted phospholipase A2 but not the group IB, IIA, and V enzymes efficiently release arachidonic acid from adherent mammalian cells. J. Biol. Chem. 2000, 275, 3179–3191. [Google Scholar] [CrossRef]
- Murakami, M.; Koduri, R.S.; Enomoto, A.; Shimbara, S.; Seki, M.; Yoshihara, K.; Singer, A.; Valentin, E.; Ghomashchi, F.; Lambeau, G.; et al. Distinct arachidonate-releasing functions of mammalian secreted phospholipase A2s in human embryonic kidney 293 and rat mastocytoma RBL-2H3 cells through heparan sulfate shuttling and external plasma membrane mechanisms. J. Biol. Chem. 2001, 276, 10083–10096. [Google Scholar] [CrossRef]
- Huggins, K.W.; Boileau, A.C.; Hui, D.Y. Protection against diet-induced obesity and obesity-related insulin resistance in Group 1B PLA2-deficient mice. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E994–E1001. [Google Scholar] [CrossRef]
- Labonte, E.D.; Kirby, R.J.; Schildmeyer, N.M.; Cannon, A.M.; Huggins, K.W.; Hui, D.Y. Group 1B phospholipase A2-mediated lysophospholipid absorption directly contributes to postprandial hyperglycemia. Diabetes 2006, 55, 935–941. [Google Scholar] [CrossRef]
- Sato, H.; Taketomi, Y.; Ushida, A.; Isogai, Y.; Kojima, T.; Hirabayashi, T.; Miki, Y.; Yamamoto, K.; Nishito, Y.; Kobayashi, T.; et al. The adipocyte-inducible secreted phospholipases PLA2G5 and PLA2G2E play distinct roles in obesity. Cell Metab. 2014, 20, 119–132. [Google Scholar] [CrossRef]
- Ohtsuki, M.; Taketomi, Y.; Arata, S.; Masuda, S.; Ishikawa, Y.; Ishii, T.; Takanezawa, Y.; Aoki, J.; Arai, H.; Yamamoto, K.; et al. Transgenic expression of group V, but not group X, secreted phospholipase A2 in mice leads to neonatal lethality because of lung dysfunction. J. Biol. Chem. 2006, 281, 36420–36433. [Google Scholar] [CrossRef]
- Boudreau, L.H.; Duchez, A.C.; Cloutier, N.; Soulet, D.; Martin, N.; Bollinger, J.; Pare, A.; Rousseau, M.; Naika, G.S.; Levesque, T.; et al. Platelets release mitochondria serving as substrate for bactericidal group IIA-secreted phospholipase A2 to promote inflammation. Blood 2014, 124, 2173–2183. [Google Scholar] [CrossRef]
- Kudo, K.; Miki, Y.; Carreras, J.; Nakayama, S.; Nakamoto, Y.; Ito, M.; Nagashima, E.; Yamamoto, K.; Higuchi, H.; Morita, S.Y.; et al. Secreted phospholipase A2 modifies extracellular vesicles and accelerates B cell lymphoma. Cell Metab. 2022, 34, 615–633.e8. [Google Scholar] [CrossRef] [PubMed]
- Yokota, Y.; Hanasaki, K.; Ono, T.; Nakazato, H.; Kobayashi, T.; Arita, H. Suppression of murine endotoxic shock by sPLA2 inhibitor, indoxam, through group IIA sPLA2-independent mechanisms. Biochim. Biophys. Acta 1999, 1438, 213–222. [Google Scholar] [CrossRef]
- Tamaru, S.; Mishina, H.; Watanabe, Y.; Watanabe, K.; Fujioka, D.; Takahashi, S.; Suzuki, K.; Nakamura, T.; Obata, J.E.; Kawabata, K.; et al. Deficiency of phospholipase A2 receptor exacerbates ovalbumin-induced lung inflammation. J. Immunol. 2013, 191, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Taketomi, Y.; Miki, Y.; Sato, H.; Hirabayashi, T.; Yamamoto, K. Recent progress in phospholipase A2 research: From cells to animals to humans. Prog. Lipid Res. 2011, 50, 152–192. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Sato, H.; Miki, Y.; Yamamoto, K.; Taketomi, Y. A new era of secreted phospholipase A2. J. Lipid Res. 2015, 56, 1248–1261. [Google Scholar] [CrossRef]
- Murakami, M.; Yamamoto, K.; Miki, Y.; Murase, R.; Sato, H.; Taketomi, Y. The roles of the secreted phospholipase A2 gene family in immunology. Adv. Immunol. 2016, 132, 91–134. [Google Scholar] [CrossRef]
- Murakami, M.; Miki, Y.; Sato, H.; Murase, R.; Taketomi, Y.; Yamamoto, K. Group IID, IIE, IIF and III secreted phospholipase A2s. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 803–818. [Google Scholar] [CrossRef]
- Murakami, M.; Sato, H.; Taketomi, Y. Updating phospholipase A2 biology. Biomolecules 2020, 10, 1457. [Google Scholar] [CrossRef]
- Hui, D.Y. Group 1B phospholipase A2 in metabolic and inflammatory disease modulation. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 784–788. [Google Scholar] [CrossRef]
- Dore, E.; Boilard, E. Roles of secreted phospholipase A2 group IIA in inflammation and host defense. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 789–802. [Google Scholar] [CrossRef]
- Baumler, A.J.; Sperandio, V. Interactions between the microbiota and pathogenic bacteria in the gut. Nature 2016, 535, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Littman, D.R. The microbiota in adaptive immune homeostasis and disease. Nature 2016, 535, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Trinchieri, G. Microbiota: A key orchestrator of cancer therapy. Nat. Rev. Cancer 2017, 17, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Balasubramanian, I.; Laubitz, D.; Tong, K.; Bandyopadhyay, S.; Lin, X.; Flores, J.; Singh, R.; Liu, Y.; Macazana, C.; et al. Paneth cell-derived lysozyme defines the composition of mucolytic microbiota and the inflammatory tone of the intestine. Immunity 2020, 53, 398–416.e8. [Google Scholar] [CrossRef]
- Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Cheng, J.; Duncan, A.E.; Kau, A.L.; Griffin, N.W.; Lombard, V.; Henrissat, B.; Bain, J.R.; et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 2013, 341, 1241214. [Google Scholar] [CrossRef]
- Arrieta, M.C.; Stiemsma, L.T.; Dimitriu, P.A.; Thorson, L.; Russell, S.; Yurist-Doutsch, S.; Kuzeljevic, B.; Gold, M.J.; Britton, H.M.; Lefebvre, D.L.; et al. Early infancy microbial and metabolic alterations affect risk of childhood asthma. Sci. Transl. Med. 2015, 7, 307ra152. [Google Scholar] [CrossRef]
- Elinav, E.; Strowig, T.; Kau, A.L.; Henao-Mejia, J.; Thaiss, C.A.; Booth, C.J.; Peaper, D.R.; Bertin, J.; Eisenbarth, S.C.; Gordon, J.I.; et al. NLRP6 inflammasome regulates colonic microbial ecology and risk for colitis. Cell 2011, 145, 745–757. [Google Scholar] [CrossRef]
- Martinez-Lopez, M.; Iborra, S.; Conde-Garrosa, R.; Mastrangelo, A.; Danne, C.; Mann, E.R.; Reid, D.M.; Gaboriau-Routhiau, V.; Chaparro, M.; Lorenzo, M.P.; et al. Microbiota sensing by Mincle-Syk axis in dendritic cells regulates interleukin-17 and -22 production and promotes intestinal barrier integrity. Immunity 2019, 50, 446–461.e9. [Google Scholar] [CrossRef]
- Boilard, E.; Lai, Y.; Larabee, K.; Balestrieri, B.; Ghomashchi, F.; Fujioka, D.; Gobezie, R.; Coblyn, J.S.; Weinblatt, M.E.; Massarotti, E.M.; et al. A novel anti-inflammatory role for secretory phospholipase A2 in immune complex-mediated arthritis. EMBO Mol. Med. 2010, 2, 172–187. [Google Scholar] [CrossRef]
- Pruzanski, W.; Vadas, P. Phospholipase A2—A mediator between proximal and distal effectors of inflammation. Immunol. Today 1991, 12, 143–146. [Google Scholar] [CrossRef]
- Snider, J.M.; You, J.K.; Wang, X.; Snider, A.J.; Hallmark, B.; Zec, M.M.; Seeds, M.C.; Sergeant, S.; Johnstone, L.; Wang, Q.; et al. Group IIA secreted phospholipase A2 is associated with the pathobiology leading to COVID-19 mortality. J. Clin. Investig. 2021, 131, e149236. [Google Scholar] [CrossRef] [PubMed]
- Duchez, A.C.; Boudreau, L.H.; Naika, G.S.; Bollinger, J.; Belleannee, C.; Cloutier, N.; Laffont, B.; Mendoza-Villarroel, R.E.; Levesque, T.; Rollet-Labelle, E.; et al. Platelet microparticles are internalized in neutrophils via the concerted activity of 12-lipoxygenase and secreted phospholipase A2-IIA. Proc. Natl. Acad. Sci. USA 2015, 112, E3564–E3573. [Google Scholar] [CrossRef] [PubMed]
- Weinrauch, Y.; Abad, C.; Liang, N.S.; Lowry, S.F.; Weiss, J. Mobilization of potent plasma bactericidal activity during systemic bacterial challenge. Role of group IIA phospholipase A2. J. Clin. Investig. 1998, 102, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Bezzine, S.; Bollinger, J.G.; Singer, A.G.; Veatch, S.L.; Keller, S.L.; Gelb, M.H. On the binding preference of human groups IIA and X phospholipases A2 for membranes with anionic phospholipids. J. Biol. Chem. 2002, 277, 48523–48534. [Google Scholar] [CrossRef]
- Singer, A.G.; Ghomashchi, F.; Le Calvez, C.; Bollinger, J.; Bezzine, S.; Rouault, M.; Sadilek, M.; Nguyen, E.; Lazdunski, M.; Lambeau, G.; et al. Interfacial kinetic and binding properties of the complete set of human and mouse groups I, II, V, X, and XII secreted phospholipases A2. J. Biol. Chem. 2002, 277, 48535–48549. [Google Scholar] [CrossRef]
- Murakami, M.; Nakatani, Y.; Kudo, I. Type II secretory phospholipase A2 associated with cell surfaces via C-terminal heparin-binding lysine residues augments stimulus-initiated delayed prostaglandin generation. J. Biol. Chem. 1996, 271, 30041–30051. [Google Scholar] [CrossRef]
- Murakami, M.; Shimbara, S.; Kambe, T.; Kuwata, H.; Winstead, M.V.; Tischfield, J.A.; Kudo, I. The functions of five distinct mammalian phospholipase A2S in regulating arachidonic acid release. Type IIa and type V secretory phospholipase A2S are functionally redundant and act in concert with cytosolic phospholipase A2. J. Biol. Chem. 1998, 273, 14411–14423. [Google Scholar] [CrossRef]
- Buckland, A.G.; Heeley, E.L.; Wilton, D.C. Bacterial cell membrane hydrolysis by secreted phospholipases A2: A major physiological role of human group IIa sPLA2 involving both bacterial cell wall penetration and interfacial catalysis. Biochim. Biophys. Acta 2000, 1484, 195–206. [Google Scholar] [CrossRef]
- Laine, V.J.; Grass, D.S.; Nevalainen, T.J. Protection by group II phospholipase A2 against Staphylococcus aureus. J. Immunol. 1999, 162, 7402–7408. [Google Scholar]
- Laine, V.J.; Grass, D.S.; Nevalainen, T.J. Resistance of transgenic mice expressing human group II phospholipase A2 to Escherichia coli infection. Infect. Immun. 2000, 68, 87–92. [Google Scholar] [CrossRef]
- Kennedy, B.P.; Payette, P.; Mudgett, J.; Vadas, P.; Pruzanski, W.; Kwan, M.; Tang, C.; Rancourt, D.E.; Cromlish, W.A. A natural disruption of the secretory group II phospholipase A2 gene in inbred mouse strains. J. Biol. Chem. 1995, 270, 22378–22385. [Google Scholar] [CrossRef] [PubMed]
- MacPhee, M.; Chepenik, K.P.; Liddell, R.A.; Nelson, K.K.; Siracusa, L.D.; Buchberg, A.M. The secretory phospholipase A2 gene is a candidate for the Mom1 locus, a major modifier of ApcMin-induced intestinal neoplasia. Cell 1995, 81, 957–966. [Google Scholar] [CrossRef]
- Mulherkar, R.; Rao, R.S.; Wagle, A.S.; Patki, V.; Deo, M.G. Enhancing factor, a Paneth cell specific protein from mouse small intestines: Predicted amino acid sequence from RT-PCR amplified cDNA and its expression. Biochem. Biophys. Res. Commun. 1993, 195, 1254–1263. [Google Scholar] [CrossRef] [PubMed]
- Schewe, M.; Franken, P.F.; Sacchetti, A.; Schmitt, M.; Joosten, R.; Bottcher, R.; van Royen, M.E.; Jeammet, L.; Payre, C.; Scott, P.M.; et al. Secreted phospholipases A2 are intestinal stem cell niche factors with distinct roles in homeostasis, inflammation, and cancer. Cell Stem Cell 2016, 19, 38–51. [Google Scholar] [CrossRef]
- Reikvam, D.H.; Erofeev, A.; Sandvik, A.; Grcic, V.; Jahnsen, F.L.; Gaustad, P.; McCoy, K.D.; Macpherson, A.J.; Meza-Zepeda, L.A.; Johansen, F.E. Depletion of murine intestinal microbiota: Effects on gut mucosa and epithelial gene expression. PLoS ONE 2011, 6, e17996. [Google Scholar] [CrossRef]
- Brodziak, F.; Meharg, C.; Blaut, M.; Loh, G. Differences in mucosal gene expression in the colon of two inbred mouse strains after colonization with commensal gut bacteria. PLoS ONE 2013, 8, e72317. [Google Scholar] [CrossRef]
- Eerola, L.I.; Surrel, F.; Nevalainen, T.J.; Gelb, M.H.; Lambeau, G.; Laine, V.J. Analysis of expression of secreted phospholipases A2 in mouse tissues at protein and mRNA levels. Biochim. Biophys. Acta 2006, 1761, 745–756. [Google Scholar] [CrossRef]
- Miki, Y.; Taketomi, Y.; Kidoguchi, Y.; Yamamoto, K.; Muramatsu, K.; Nishito, Y.; Park, J.; Hosomi, K.; Mizuguchi, K.; Kunisawa, J.; et al. Group IIA secreted phospholipase A2 controls skin carcinogenesis and psoriasis by shaping the gut microbiota. JCI Insight 2022, 7, e152611. [Google Scholar] [CrossRef]
- Dore, E.; Joly-Beauparlant, C.; Morozumi, S.; Mathieu, A.; Levesque, T.; Allaeys, I.; Duchez, A.C.; Cloutier, N.; Leclercq, M.; Bodein, A.; et al. The interaction of secreted phospholipase A2-IIA with the microbiota alters its lipidome and promotes inflammation. JCI Insight 2022, 7, e152638. [Google Scholar] [CrossRef]
- Sawada, H.; Murakami, M.; Enomoto, A.; Shimbara, S.; Kudo, I. Regulation of type V phospholipase A2 expression and function by proinflammatory stimuli. Eur. J. Biochem. 1999, 263, 826–835. [Google Scholar] [CrossRef]
- Fox, J.G.; Wang, T.C. Inflammation, atrophy, and gastric cancer. J. Clin. Investig. 2007, 117, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Nagata, M.; Toyonaga, K.; Ishikawa, E.; Haji, S.; Okahashi, N.; Takahashi, M.; Izumi, Y.; Imamura, A.; Takato, K.; Ishida, H.; et al. Helicobacter pylori metabolites exacerbate gastritis through C-type lectin receptors. J. Exp. Med. 2021, 218, e20200815. [Google Scholar] [CrossRef] [PubMed]
- Blaser, M.J.; Chen, Y.; Reibman, J. Does Helicobacter pylori protect against asthma and allergy? Gut 2008, 57, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Blaser, M.J. Helicobacter pylori colonization is inversely associated with childhood asthma. J. Infect. Dis. 2008, 198, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Leung, S.Y.; Chen, X.; Chu, K.M.; Yuen, S.T.; Mathy, J.; Ji, J.; Chan, A.S.; Li, R.; Law, S.; Troyanskaya, O.G.; et al. Phospholipase A2 group IIA expression in gastric adenocarcinoma is associated with prolonged survival and less frequent metastasis. Proc. Natl. Acad. Sci. USA 2002, 99, 16203–16208. [Google Scholar] [CrossRef] [PubMed]
- Onsun, N.; Arda Ulusal, H.; Su, O.; Beycan, I.; Biyik Ozkaya, D.; Senocak, M. Impact of Helicobacter pylori infection on severity of psoriasis and response to treatment. Eur. J. Dermatol. 2012, 22, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Lercher, A.; Bhattacharya, A.; Popa, A.M.; Caldera, M.; Schlapansky, M.F.; Baazim, H.; Agerer, B.; Gurtl, B.; Kosack, L.; Majek, P.; et al. Type I interferon signaling disrupts the hepatic urea cycle and alters systemic metabolism to suppress T cell function. Immunity 2019, 51, 1074–1087.e9. [Google Scholar] [CrossRef]
- Lee, J.S.; Adler, L.; Karathia, H.; Carmel, N.; Rabinovich, S.; Auslander, N.; Keshet, R.; Stettner, N.; Silberman, A.; Agemy, L.; et al. Urea cycle dysregulation generates clinically relevant genomic and biochemical signatures. Cell 2018, 174, 1559–1570.e22. [Google Scholar] [CrossRef]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach? Nat. Rev. Drug Discov. 2009, 8, 579–591. [Google Scholar] [CrossRef]
- Kuraishy, A.; Karin, M.; Grivennikov, S.I. Tumor promotion via injury- and death-induced inflammation. Immunity 2011, 35, 467–477. [Google Scholar] [CrossRef]
- Glunde, K.; Bhujwalla, Z.M.; Ronen, S.M. Choline metabolism in malignant transformation. Nat. Rev. Cancer 2011, 11, 835–848. [Google Scholar] [CrossRef] [PubMed]
- Arlt, A.; Sebens, S.; Krebs, S.; Geismann, C.; Grossmann, M.; Kruse, M.L.; Schreiber, S.; Schafer, H. Inhibition of the Nrf2 transcription factor by the alkaloid trigonelline renders pancreatic cancer cells more susceptible to apoptosis through decreased proteasomal gene expression and proteasome activity. Oncogene 2013, 32, 4825–4835. [Google Scholar] [CrossRef] [PubMed]
- Brink, L.R.; Mercer, K.E.; Piccolo, B.D.; Chintapalli, S.V.; Elolimy, A.; Bowlin, A.K.; Matazel, K.S.; Pack, L.; Adams, S.H.; Shankar, K.; et al. Neonatal diet alters fecal microbiota and metabolome profiles at different ages in infants fed breast milk or formula. Am. J. Clin. Nutr. 2020, 111, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Syed, I.; Lee, J.; Moraes-Vieira, P.M.; Donaldson, C.J.; Sontheimer, A.; Aryal, P.; Wellenstein, K.; Kolar, M.J.; Nelson, A.T.; Siegel, D.; et al. Palmitic acid hydroxystearic acids activate GPR40, which is involved in their beneficial effects on glucose homeostasis. Cell Metab. 2018, 27, 419–427.e4. [Google Scholar] [CrossRef]
- Yore, M.M.; Syed, I.; Moraes-Vieira, P.M.; Zhang, T.; Herman, M.A.; Homan, E.A.; Patel, R.T.; Lee, J.; Chen, S.; Peroni, O.D.; et al. Discovery of a class of endogenous mammalian lipids with anti-diabetic and anti-inflammatory effects. Cell 2014, 159, 318–332. [Google Scholar] [CrossRef]
- Gowda, S.G.B.; Liang, C.; Gowda, D.; Hou, F.; Kawakami, K.; Fukiya, S.; Yokota, A.; Chiba, H.; Hui, S.P. Identification of short-chain fatty acid esters of hydroxy fatty acids (SFAHFAs) in a murine model by nontargeted analysis using ultra-high-performance liquid chromatography/linear ion trap quadrupole-Orbitrap mass spectrometry. Rapid Commun. Mass Spectrom. 2020, 34, e8831. [Google Scholar] [CrossRef]
- Fujimura, K.E.; Sitarik, A.R.; Havstad, S.; Lin, D.L.; Levan, S.; Fadrosh, D.; Panzer, A.R.; LaMere, B.; Rackaityte, E.; Lukacs, N.W.; et al. Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat. Med. 2016, 22, 1187–1191. [Google Scholar] [CrossRef]
- Kirjavainen, P.V.; Karvonen, A.M.; Adams, R.I.; Taubel, M.; Roponen, M.; Tuoresmaki, P.; Loss, G.; Jayaprakash, B.; Depner, M.; Ege, M.J.; et al. Farm-like indoor microbiota in non-farm homes protects children from asthma development. Nat. Med. 2019, 25, 1089–1095. [Google Scholar] [CrossRef]
- Depner, M.; Taft, D.H.; Kirjavainen, P.V.; Kalanetra, K.M.; Karvonen, A.M.; Peschel, S.; Schmausser-Hechfellner, E.; Roduit, C.; Frei, R.; Lauener, R.; et al. Maturation of the gut microbiome during the first year of life contributes to the protective farm effect on childhood asthma. Nat. Med. 2020, 26, 1766–1775. [Google Scholar] [CrossRef]
- Chen, C.C.; Chen, K.J.; Kong, M.S.; Chang, H.J.; Huang, J.L. Alterations in the gut microbiotas of children with food sensitization in early life. Pediatr. Allergy Immunol. 2016, 27, 254–262. [Google Scholar] [CrossRef]
- Sordillo, J.E.; Zhou, Y.; McGeachie, M.J.; Ziniti, J.; Lange, N.; Laranjo, N.; Savage, J.R.; Carey, V.; O’Connor, G.; Sandel, M.; et al. Factors influencing the infant gut microbiome at age 3–6 months: Findings from the ethnically diverse Vitamin D Antenatal Asthma Reduction Trial (VDAART). J. Allergy Clin. Immunol. 2017, 139, 482–491.e414. [Google Scholar] [CrossRef] [PubMed]
- Chua, H.H.; Chou, H.C.; Tung, Y.L.; Chiang, B.L.; Liao, C.C.; Liu, H.H.; Ni, Y.H. Intestinal dysbiosis featuring abundance of Ruminococcus gnavus associates with allergic diseases in infants. Gastroenterology 2018, 154, 154–167. [Google Scholar] [CrossRef] [PubMed]
- Bao, R.; Hesser, L.A.; He, Z.; Zhou, X.; Nadeau, K.C.; Nagler, C.R. Fecal microbiome and metabolome differ in healthy and food-allergic twins. J. Clin. Investig. 2021, 131, e141935. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Chen, Y.; Li, Q.; Ye, X.; Guo, X.; Sun, L.; Zou, J.; Shen, Y.; Mao, Y.; Li, C.; et al. Tetrahydrocurcumin alleviates allergic airway inflammation in asthmatic mice by modulating the gut microbiota. Food Funct. 2021, 12, 6830–6840. [Google Scholar] [CrossRef] [PubMed]
- Taketomi, Y.; Ueno, N.; Kojima, T.; Sato, H.; Murase, R.; Yamamoto, K.; Tanaka, S.; Sakanaka, M.; Nakamura, M.; Nishito, Y.; et al. Mast cell maturation is driven via a group III phospholipase A2-prostaglandin D2-DP1 receptor paracrine axis. Nat. Immunol. 2013, 14, 554–563. [Google Scholar] [CrossRef]
- Nevalainen, T.J.; Eerola, L.I.; Rintala, E.; Laine, V.J.; Lambeau, G.; Gelb, M.H. Time-resolved fluoroimmunoassays of the complete set of secreted phospholipases A2 in human serum. Biochim. Biophys. Acta 2005, 1733, 210–223. [Google Scholar] [CrossRef]
- Green, J.A.; Smith, G.M.; Buchta, R.; Lee, R.; Ho, K.Y.; Rajkovic, I.A.; Scott, K.F. Circulating phospholipase A2 activity associated with sepsis and septic shock is indistinguishable from that associated with rheumatoid arthritis. Inflammation 1991, 15, 355–367. [Google Scholar] [CrossRef]
- Rintala, E.M.; Nevalainen, T.J. Group II phospholipase A2 in sera of febrile patients with microbiologically or clinically documented infections. Clin. Infect. Dis. 1993, 17, 864–870. [Google Scholar] [CrossRef]
- Pratt, J.S.; Sachen, K.L.; Wood, H.D.; Eaton, K.A.; Young, V.B. Modulation of host immune responses by the cytolethal distending toxin of Helicobacter hepaticus. Infect. Immun. 2006, 74, 4496–4504. [Google Scholar] [CrossRef]
- Falsafi, T.; Mahboubi, M. Helicobacter hepaticus, a new pathogenic species of the Helicobacter genus: Similarities and differences with H. pylori. Iran. J. Microbiol. 2013, 5, 185–194. [Google Scholar]
- Scher, J.U.; Sczesnak, A.; Longman, R.S.; Segata, N.; Ubeda, C.; Bielski, C.; Rostron, T.; Cerundolo, V.; Pamer, E.G.; Abramson, S.B.; et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. eLife 2013, 2, e01202. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.M. The immune response to Prevotella bacteria in chronic inflammatory disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Scher, J.U.; Littman, D.R.; Abramson, S.B. Microbiome in inflammatory arthritis and human rheumatic diseases. Arthritis Rheumatol. 2016, 68, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Pruzanski, W.; Scott, K.; Smith, G.; Rajkovic, I.; Stefanski, E.; Vadas, P. Enzymatic activity and immunoreactivity of extracellular phospholipase A2 in inflammatory synovial fluids. Inflammation 1992, 16, 451–457. [Google Scholar] [CrossRef]
- Kouskoff, V.; Korganow, A.S.; Duchatelle, V.; Degott, C.; Benoist, C.; Mathis, D. Organ-specific disease provoked by systemic autoimmunity. Cell 1996, 87, 811–822. [Google Scholar] [CrossRef]
- Matsumoto, I.; Staub, A.; Benoist, C.; Mathis, D. Arthritis provoked by linked T and B cell recognition of a glycolytic enzyme. Science 1999, 286, 1732–1735. [Google Scholar] [CrossRef]
- Pernet, E.; Guillemot, L.; Burgel, P.R.; Martin, C.; Lambeau, G.; Sermet-Gaudelus, I.; Sands, D.; Leduc, D.; Morand, P.C.; Jeammet, L.; et al. Pseudomonas aeruginosa eradicates Staphylococcus aureus by manipulating the host immunity. Nat. Commun. 2014, 5, 5105. [Google Scholar] [CrossRef]
- Murase, R.; Sato, H.; Yamamoto, K.; Ushida, A.; Nishito, Y.; Ikeda, K.; Kobayashi, T.; Yamamoto, T.; Taketomi, Y.; Murakami, M. Group X secreted phospholipase A2 releases ω3 polyunsaturated fatty acids, suppresses colitis, and promotes sperm fertility. J. Biol. Chem. 2016, 291, 6895–6911. [Google Scholar] [CrossRef]
- Murase, R.; Taketomi, Y.; Miki, Y.; Nishito, Y.; Saito, M.; Fukami, K.; Yamamoto, K.; Murakami, M. Group III phospholipase A2 promotes colitis and colorectal cancer. Sci. Rep. 2017, 7, 12261. [Google Scholar] [CrossRef]
- Entwistle, L.J.; Pelly, V.S.; Coomes, S.M.; Kannan, Y.; Perez-Lloret, J.; Czieso, S.; Silva Dos Santos, M.; MacRae, J.I.; Collinson, L.; Sesay, A.; et al. Epithelial-cell-derived phospholipase A2 group 1B is an endogenous anthelmintic. Cell Host Microbe 2017, 22, 484–493.e5. [Google Scholar] [CrossRef]
- Hollie, N.I.; Hui, D.Y. Group 1B phospholipase A2 deficiency protects against diet-induced hyperlipidemia in mice. J. Lipid Res. 2011, 52, 2005–2011. [Google Scholar] [CrossRef] [PubMed]
- Hollie, N.I.; Konaniah, E.S.; Goodin, C.; Hui, D.Y. Group 1B phospholipase A2 inactivation suppresses atherosclerosis and metabolic diseases in LDL receptor-deficient mice. Atherosclerosis 2014, 234, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Balestrieri, B.; Maekawa, A.; Xing, W.; Gelb, M.H.; Katz, H.R.; Arm, J.P. Group V secretory phospholipase A2 modulates phagosome maturation and regulates the innate immune response against Candida albicans. J. Immunol. 2009, 182, 4891–4898. [Google Scholar] [CrossRef] [PubMed]
- Degousee, N.; Kelvin, D.J.; Geisslinger, G.; Hwang, D.M.; Stefanski, E.; Wang, X.H.; Danesh, A.; Angioni, C.; Schmidt, H.; Lindsay, T.F.; et al. Group V phospholipase A2 in bone marrow-derived myeloid cells and bronchial epithelial cells promotes bacterial clearance after Escherichia coli pneumonia. J. Biol. Chem. 2011, 286, 35650–35662. [Google Scholar] [CrossRef]
- Zheng, J.; Meyerholz, D.; Wong, L.R.; Gelb, M.; Murakami, M.; Perlman, S. Coronavirus-specific antibody production in middle-aged mice requires phospholipase A2G2D. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef]
- Sato, H.; Isogai, Y.; Masuda, S.; Taketomi, Y.; Miki, Y.; Kamei, D.; Hara, S.; Kobayashi, T.; Ishikawa, Y.; Ishii, T.; et al. Physiological roles of group X-secreted phospholipase A2 in reproduction, gastrointestinal phospholipid digestion, and neuronal function. J. Biol. Chem. 2011, 286, 11632–11648. [Google Scholar] [CrossRef]
- Shridas, P.; Bailey, W.M.; Talbott, K.R.; Oslund, R.C.; Gelb, M.H.; Webb, N.R. Group X secretory phospholipase A2 enhances TLR4 signaling in macrophages. J. Immunol. 2011, 187, 482–489. [Google Scholar] [CrossRef]
- Li, X.; Shridas, P.; Forrest, K.; Bailey, W.; Webb, N.R. Group X secretory phospholipase A2 negatively regulates adipogenesis in murine models. FASEB J. 2010, 24, 4313–4324. [Google Scholar] [CrossRef]
- Ait-Oufella, H.; Herbin, O.; Lahoute, C.; Coatrieux, C.; Loyer, X.; Joffre, J.; Laurans, L.; Ramkhelawon, B.; Blanc-Brude, O.; Karabina, S.; et al. Group X secreted phospholipase A2 limits the development of atherosclerosis in LDL receptor-null mice. Arter. Thromb. Vasc. Biol. 2013, 33, 466–473. [Google Scholar] [CrossRef]
- Zack, M.; Boyanovsky, B.B.; Shridas, P.; Bailey, W.; Forrest, K.; Howatt, D.A.; Gelb, M.H.; de Beer, F.C.; Daugherty, A.; Webb, N.R. Group X secretory phospholipase A2 augments angiotensin II-induced inflammatory responses and abdominal aortic aneurysm formation in apoE-deficient mice. Atherosclerosis 2011, 214, 58–64. [Google Scholar] [CrossRef]
- Yamamoto, K.; Miki, Y.; Sato, M.; Taketomi, Y.; Nishito, Y.; Taya, C.; Muramatsu, K.; Ikeda, K.; Nakanishi, H.; Taguchi, R.; et al. The role of group IIF-secreted phospholipase A2 in epidermal homeostasis and hyperplasia. J. Exp. Med. 2015, 212, 1901–1919. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Miki, Y.; Sato, H.; Nishito, Y.; Gelb, M.H.; Taketomi, Y.; Murakami, M. Expression and function of group IIE phospholipase A2 in mouse skin. J. Biol. Chem. 2016, 291, 15602–15613. [Google Scholar] [CrossRef] [PubMed]
- Hui, D.Y.; Cope, M.J.; Labonte, E.D.; Chang, H.T.; Shao, J.; Goka, E.; Abousalham, A.; Charmot, D.; Buysse, J. The phospholipase A2 inhibitor methyl indoxam suppresses diet-induced obesity and glucose intolerance in mice. Br. J. Pharmacol. 2009, 157, 1263–1269. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).