
The Role of Nutraceutical Containing Polyphenols in Diabetes Prevention




Abstract
:1. Introduction
Diabetes
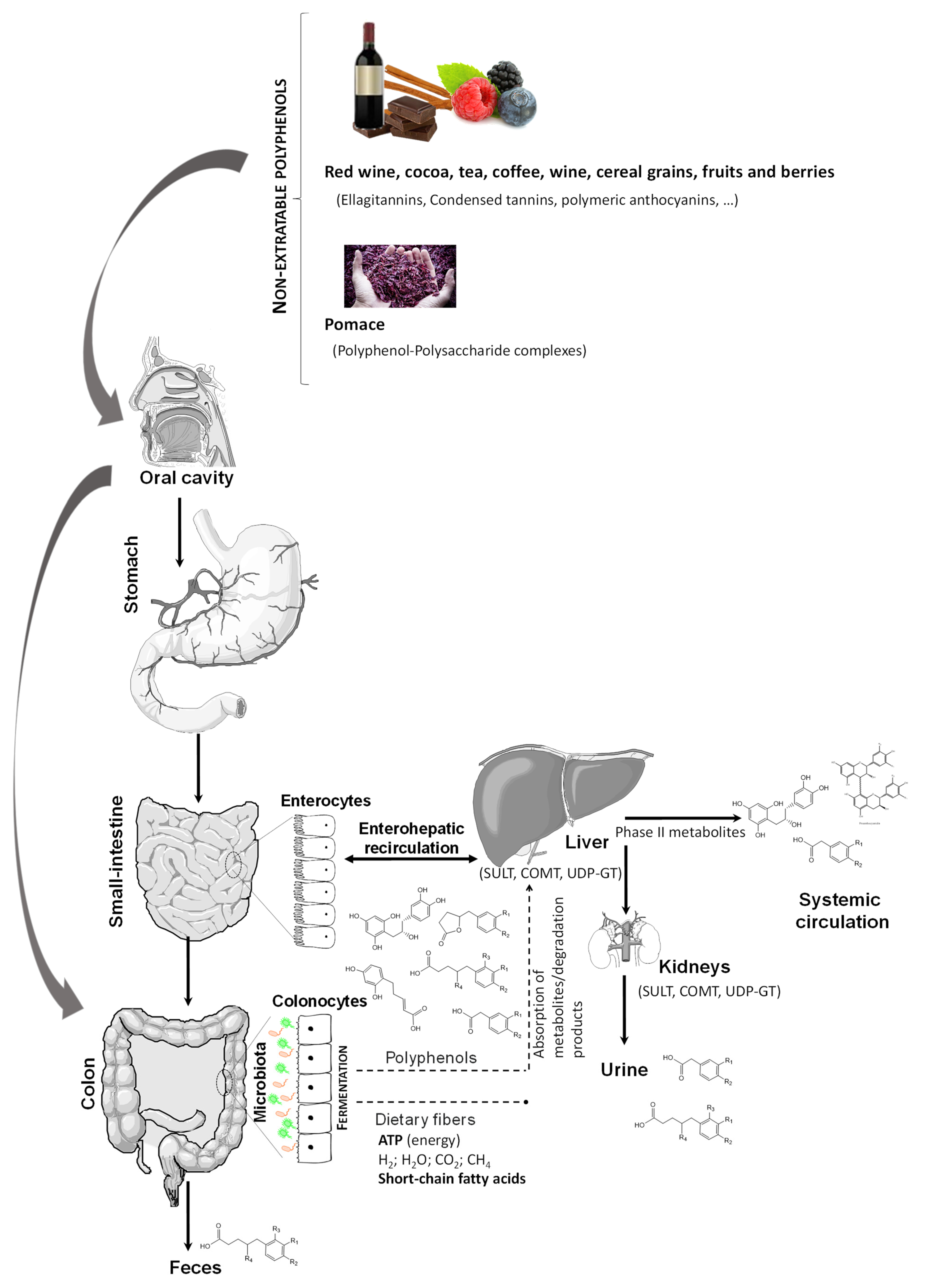
2. Phytochemical Compounds: Structure, Food Sources, Bioavailability, and Metabolism
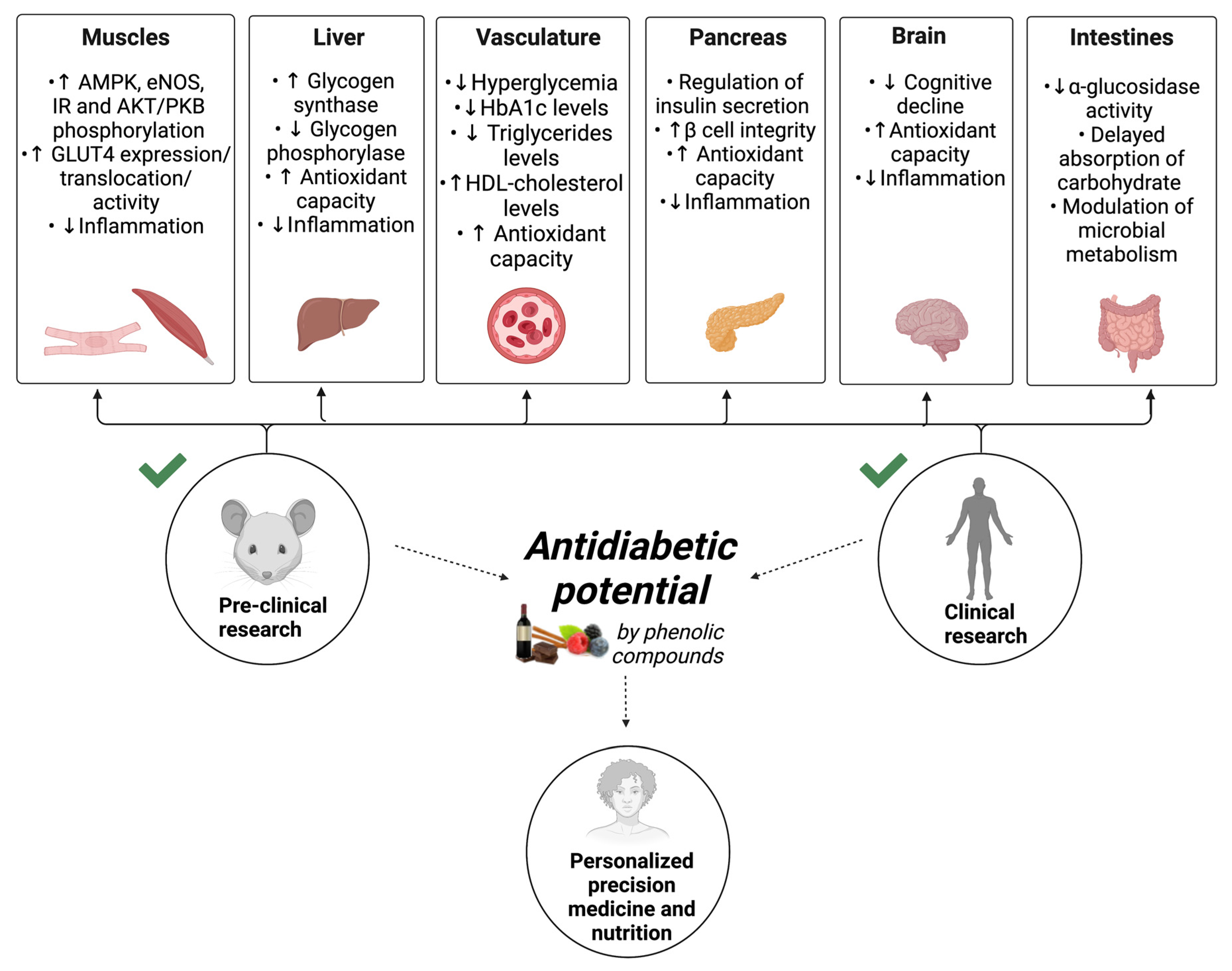
3. Intracellular Metabolites and Biomarkers Altered in Response to Nutraceutical Containing Polyphenol Intake
4. General Summary and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diabetes Around the World in 2021. Available online: https://diabetesatlas.org (accessed on 15 December 2021).
- Oliveira, A.; Araújo, J.; Severo, M.; Correia, D.; Ramos, E.; Torres, D.; Lopes, C. Prevalence of general and abdominal obesity in Portugal: Comprehensive results from the National Food, nutrition and physical activity survey 2015–2016. BMC Public Health 2018, 18, 614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreto, M.; Kislaya, I.; Gaio, V.; Rodrigues, A.P.; Santos, A.J.; Namorado, S.; Antunes, L.; Gil, A.P.; Boavida, J.M.; Ribeiro, R.T.; et al. Prevalence, awareness, treatment and control of diabetes in Portugal: Results from the first National Health examination Survey (INSEF 2015). Diabetes Res. Clin. Pract. 2018, 140, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.K.; Sharma, V. World diabetes day 2018: Battling the Emerging Epidemic of Diabetic Retinopathy. Indian J. Ophthalmol. 2018, 66, 1652–1653. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, S.; Vaz, M.; Kurpad, A.V. Protein intakes in India. Br. J. Nutr. 2012, 108 (Suppl. S2), S50–S58. [Google Scholar] [CrossRef] [Green Version]
- Mahalle, N.; Kulkarni, M.V.; Naik, S.S.; Garg, M.K. Association of dietary factors with insulin resistance and inflammatory markers in subjects with diabetes mellitus and coronary artery disease in Indian population. J. Diabetes Complicat. 2014, 28, 536–541. [Google Scholar] [CrossRef]
- Geetha, L.; Deepa, M.; Anjana, R.M.; Mohan, V. Prevalence and clinical profile of metabolic obesity and phenotypic obesity in Asian Indians. J. Diabetes Sci. Technol. 2011, 5, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Indulekha, K.; Surendar, J.; Anjana, R.M.; Geetha, L.; Gokulakrishnan, K.; Pradeepa, R.; Mohan, V. Metabolic obesity, adipocytokines, and inflammatory markers in Asian Indians--CURES-124. Diabetes Technol. Ther. 2015, 17, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Burchfield, J.G.; Kebede, M.A.; Meoli, C.C.; Stöckli, J.; Whitworth, P.T.; Wright, A.L.; Hoffman, N.J.; Minard, A.Y.; Ma, X.; Krycer, J.R.; et al. High dietary fat and sucrose results in an extensive and time-dependent deterioration in health of multiple physiological systems in mice. J. Biol. Chem. 2018, 293, 5731–5745. [Google Scholar] [CrossRef] [Green Version]
- Burgeiro, A.; Cerqueira, M.G.; Varela-Rodríguez, B.M.; Nunes, S.; Neto, P.; Pereira, F.C.; Reis, F.; Carvalho, E. Glucose and Lipid Dysmetabolism in a Rat Model of Prediabetes Induced by a High-Sucrose Diet. Nutrients 2017, 9, 638. [Google Scholar] [CrossRef]
- Association, A.D. 1. Improving Care and Promoting Health in Populations: Standards of Medical Care in Diabetes—2019. Diabetes Care 2019, 42, S7–S12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, D.P.; Duvuuri, V. Diabetes mellitus. Clin. Podiatr. Med. Surg. 2002, 19, 79–107. [Google Scholar] [CrossRef]
- O’Rahilly, S.; Hattersley, A.; Vaag, A.; Gray, H. Insulin resistance as the major cause of impaired glucose tolerance: A self-fulfilling prophesy? Lancet 1994, 344, 585–589. [Google Scholar] [CrossRef]
- Roberts, C.K.; Hevener, A.L.; Barnard, R.J. Metabolic syndrome and insulin resistance: Underlying causes and modification by exercise training. Compr. Physiol. 2013, 3, 1–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, E.; Jansson, P.A.; Axelsen, M.; Eriksson, J.W.; Huang, X.; Groop, L.; Rondinone, C.; Sjostrom, L.; Smith, U. Low cellular IRS 1 gene and protein expression predict insulin resistance and NIDDM. FASEB J. 1999, 13, 2173–2178. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, E.; Jansson, P.A.; Nagaev, I.; Wenthzel, A.M.; Smith, U. Insulin resistance with low cellular IRS-1 expression is also associated with low GLUT4 expression and impaired insulin-stimulated glucose transport. FASEB J. 2001, 15, 1101–1103. [Google Scholar] [CrossRef] [Green Version]
- Fritz, J.; Bjorge, T.; Nagel, G.; Manjer, J.; Engeland, A.; Haggstrom, C.; Concin, H.; Teleka, S.; Tretli, S.; Gylling, B.; et al. The triglyceride-glucose index as a measure of insulin resistance and risk of obesity-related cancers. Int. J. Epidemiol. 2020, 49, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, P.; Melnyk, S.; Bennuri, S.C.; Delhey, L.; Reis, A.; Moura, G.R.; Børsheim, E.; Rose, S.; Carvalho, E. Redox Imbalance and Methylation Disturbances in Early Childhood Obesity. Oxid. Med. Cell. Longev. 2021, 2021, 2207125. [Google Scholar] [CrossRef]
- Graf, C.; Ferrari, N. Metabolic Syndrome in Children and Adolescents. Visc. Med. 2016, 32, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Song, S.H.; Hardisty, C.A. Early onset type 2 diabetes mellitus: A harbinger for complications in later years—Clinical observation from a secondary care cohort. QJM 2009, 102, 799–806. [Google Scholar] [CrossRef] [Green Version]
- Daniels, S.R. Complications of obesity in children and adolescents. Int. J. Obes. 2009, 33 (Suppl. S1), S60–S65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Methnani, J.; Amor, D.; Yousfi, N.; Bouslama, A.; Omezzine, A.; Bouhlel, E. Sedentary behavior, exercise and COVID-19: Immune and metabolic implications in obesity and its comorbidities. J. Sports Med. Phys. Fit. 2020, 61, 1538–1547. [Google Scholar] [CrossRef] [PubMed]
- Azar, W.S.; Njeim, R.; Fares, A.H.; Azar, N.S.; Azar, S.T.; El Sayed, M.; Eid, A.A. COVID-19 and diabetes mellitus: How one pandemic worsens the other. Rev. Endocr. Metab. Disord. 2020, 21, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Stefan, N.; Birkenfeld, A.L.; Schulze, M.B. Global pandemics interconnected—obesity, impaired metabolic health and COVID-19. Nat. Rev. Endocrinol. 2021, 17, 135–149. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, P.J.; Ismail-Beigi, F. Near-Normalization of Glucose and Microvascular Diabetes Complications: Data from ACCORD and ADVANCE. Ther. Adv. Endocrinol. Metab. 2011, 2, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, C.; Del Prato, S. Metabolic memory and individual treatment aims in type 2 diabetes--outcome-lessons learned from large clinical trials. Rev. Diabet Stud. 2011, 8, 432–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.M.; Patterson, S.J.; Speck, M.; Ehses, J.A.; Levings, M.K. Insulin inhibits IL-10-mediated regulatory T cell function: Implications for obesity. J. Immunol. 2014, 192, 623–629. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.Y.; Mei Wong, J.L.; Sim, Y.J.; Wong, S.S.; Mohamed Elhassan, S.A.; Tan, S.H.; Ling Lim, G.P.; Rong Tay, N.W.; Annan, N.C.; Bhattamisra, S.K.; et al. Type 1 and 2 diabetes mellitus: A review on current treatment approach and gene therapy as potential intervention. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 364–372. [Google Scholar] [CrossRef]
- Sarmento, B.; Ribeiro, A.; Veiga, F.; Sampaio, P.; Neufeld, R.; Ferreira, D. Alginate/chitosan nanoparticles are effective for oral insulin delivery. Pharm. Res. 2007, 24, 2198–2206. [Google Scholar] [CrossRef] [Green Version]
- Chaudhury, A.; Duvoor, C.; Reddy Dendi, V.S.; Kraleti, S.; Chada, A.; Ravilla, R.; Marco, A.; Shekhawat, N.S.; Montales, M.T.; Kuriakose, K.; et al. Clinical Review of Antidiabetic Drugs: Implications for Type 2 Diabetes Mellitus Management. Front. Endocrinol. 2017, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Kastorini, C.M.; Milionis, H.J.; Esposito, K.; Giugliano, D.; Goudevenos, J.A.; Panagiotakos, D.B. The effect of Mediterranean diet on metabolic syndrome and its components: A meta-analysis of 50 studies and 534,906 individuals. J. Am. Coll. Cardiol. 2011, 57, 1299–1313. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Ou, J.; Chen, L.; Zhang, Y.; Szkudelski, T.; Delmas, D.; Daglia, M.; Xiao, J. Dietary polyphenols and type 2 diabetes: Human Study and Clinical Trial. Crit. Rev. Food Sci. Nutr. 2019, 59, 3371–3379. [Google Scholar] [CrossRef]
- Cieślik, E.; Gręda, A.; Adamus, W. Contents of polyphenols in fruit and vegetables. Food Chem. 2006, 94, 135–142. [Google Scholar] [CrossRef]
- Collin, S.; Jerkovic, V.; Bröhan, M.; Callemien, D. Polyphenols and Beer Quality. In Natural Products: Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 2333–2359. [Google Scholar] [CrossRef]
- Khan, N.; Mukhtar, H. Tea polyphenols for health promotion. Life Sci. 2007, 81, 519–533. [Google Scholar] [CrossRef] [Green Version]
- Gião, M.S.; González-Sanjosé, M.L.; Rivero-Pérez, M.D.; Pereira, C.I.; Pintado, M.E.; Malcata, F.X. Infusions of Portuguese medicinal plants: Dependence of final antioxidant capacity and phenol content on extraction features. J. Sci. Food Agric. 2007, 87, 2638–2647. [Google Scholar] [CrossRef]
- Vinson, J.A.; Proch, J.; Zubik, L. Phenol antioxidant quantity and quality in foods: Cocoa, dark chocolate, and milk chocolate. J. Agric. Food Chem. 1999, 47, 4821–4824. [Google Scholar] [CrossRef] [PubMed]
- Frankel, E.N.; Waterhouse, A.L.; Teissedre, P.L. Principal Phenolic Phytochemicals in Selected California Wines and Their Antioxidant Activity in Inhibiting Oxidation of Human Low-Density Lipoproteins. J. Agric. Food Chem. 1995, 43, 890–894. [Google Scholar] [CrossRef]
- Beckman, C.H. Phenolic-storing cells: Keys to programmed cell death and periderm formation in wilt disease resistance and in general defence responses in plants? Physiol. Mol. Plant. Pathol. 2000, 57, 101–110. [Google Scholar] [CrossRef]
- Papuc, C.; Goran, G.V.; Predescu, C.N.; Nicorescu, V.; Stefan, G. Plant Polyphenols as Antioxidant and Antibacterial Agents for Shelf-Life Extension of Meat and Meat Products: Classification, Structures, Sources, and Action Mechanisms. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1243–1268. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Singh, S.K.; Chowdhury, I.; Singh, R. Understanding the Mechanism of Bacterial Biofilms Resistance to Antimicrobial Agents. Open Microbiol. J. 2017, 11, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Del Bo, C.; Bernardi, S.; Marino, M.; Porrini, M.; Tucci, M.; Guglielmetti, S.; Cherubini, A.; Carrieri, B.; Kirkup, B.; Kroon, P.; et al. Systematic Review on Polyphenol Intake and Health Outcomes: Is there Sufficient Evidence to Define a Health-Promoting Polyphenol-Rich Dietary Pattern? Nutrients 2019, 11, 1355. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.; Kelm, M.A.; Hammerstone, J.F.; Beecher, G.; Holden, J.; Haytowitz, D.; Gebhardt, S.; Prior, R.L. Concentrations of proanthocyanidins in common foods and estimations of normal consumption. J. Nutr. 2004, 134, 613–617. [Google Scholar] [CrossRef]
- Sánchez-González, I.; Jiménez-Escrig, A.; Saura-Calixto, F. In vitro antioxidant activity of coffees brewed using different procedures (Italian, espresso and filter). Food Chem. 2005, 90, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Cheynier, V. Polyphenols in foods are more complex than often thought. Am. J. Clin. Nutr. 2005, 81, 223S–229S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, K.; Nagel, C.W. Occurrence and content of hydroxycinnamic and hydroxybenzoic acid compounds in foods. Crit. Rev. Food Sci. Nutr. 1989, 28, 315–347. [Google Scholar] [CrossRef] [PubMed]
- El-Seedi, H.R.; El-Said, A.M.A.; Khalifa, S.A.M.; Göransson, U.; Bohlin, L.; Borg-Karlson, A.-K.; Verpoorte, R. Biosynthesis, Natural Sources, Dietary Intake, Pharmacokinetic Properties, and Biological Activities of Hydroxycinnamic Acids. J. Agric. Food Chem. 2012, 60, 10877–10895. [Google Scholar] [CrossRef] [PubMed]
- Mølgaard, P.; Ravn, H. Evolutionary aspects of caffeoyl ester distribution In Dicotyledons. Phytochemistry 1988, 27, 2411–2421. [Google Scholar] [CrossRef]
- Vestergaard, M.; Ingmer, H. Antibacterial and antifungal properties of resveratrol. Int. J. Antimicrob. Agents 2019, 53, 716–723. [Google Scholar] [CrossRef]
- Pezzuto, J.M. Resveratrol: Twenty Years of Growth, Development and Controversy. Biomol. Ther. 2019, 27, 1–14. [Google Scholar] [CrossRef]
- Sanders, T.H.; McMichael, R.W., Jr.; Hendrix, K.W. Occurrence of resveratrol in edible peanuts. J. Agric. Food Chem. 2000, 48, 1243–1246. [Google Scholar] [CrossRef]
- Karadeniz, F.; Durst, R.W.; Wrolstad, R.E. Polyphenolic Composition of Raisins. J. Agric. Food Chem. 2000, 48, 5343–5350. [Google Scholar] [CrossRef] [PubMed]
- Ehala, S.; Vaher, M.; Kaljurand, M. Characterization of Phenolic Profiles of Northern European Berries by Capillary Electrophoresis and Determination of their Antioxidant Activity. J. Agric. Food Chem. 2005, 53, 6484–6490. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Xian, D.; Xiong, X.; Lai, R.; Song, J.; Zhong, J. Proanthocyanidins against Oxidative Stress: From Molecular Mechanisms to Clinical Applications. BioMed Res. Int. 2018, 2018, 8584136. [Google Scholar] [CrossRef] [Green Version]
- Giuliani, A.; Cerretani, L.; Cichelli, A. Colors: Properties and Determination of Natural Pigments. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; pp. 273–283. [Google Scholar] [CrossRef]
- Kontoudakis, N.; Esteruelas, M.; Fort, F.; Canals, J.M.; De Freitas, V.; Zamora, F. Influence of the heterogeneity of grape phenolic maturity on wine composition and quality. Food Chem. 2011, 124, 767–774. [Google Scholar] [CrossRef]
- Sun, B.; Neves, A.C.; Fernandes, T.A.; Fernandes, A.L.; Mateus, N.; De Freitas, V.; Leandro, C.; Spranger, M.I. Evolution of Phenolic Composition of Red Wine during Vinification and Storage and Its Contribution to Wine Sensory Properties and Antioxidant Activity. J. Agric. Food Chem. 2011, 59, 6550–6557. [Google Scholar] [CrossRef]
- Agudo, A. Measuring intake of fruits and vegetables. In Proceedings of the Joint FAO/WHO Workshop on Fruits and Vegetables for Health, Kobe, Japan, 1–3 September 2004. [Google Scholar]
- Kapolou, A.; Karantonis, H.C.; Rigopoulos, N.; Koutelidakis, A.E. Association of Mean Daily Polyphenols Intake with Mediterranean Diet Adherence and Anthropometric Indices in Healthy Greek Adults: A Retrospective Study. Appl. Sci. 2021, 11, 4664. [Google Scholar] [CrossRef]
- Visvanathan, R.; Williamson, G. Citrus polyphenols and risk of type 2 diabetes: Evidence from mechanistic studies. Crit. Rev. Food Sci. Nutr. 2021, 1–25. [Google Scholar] [CrossRef]
- Bozzetto, L.; Annuzzi, G.; Pacini, G.; Costabile, G.; Vetrani, C.; Vitale, M.; Griffo, E.; Giacco, A.; De Natale, C.; Cocozza, S.; et al. Polyphenol-rich diets improve glucose metabolism in people at high cardiometabolic risk: A controlled randomised intervention trial. Diabetologia 2015, 58, 1551–1560. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, T.; Fukui, M.; Tanaka, M.; Senmaru, T.; Iwase, H.; Yamazaki, M.; Aoi, W.; Inui, T.; Nakamura, N.; Marunaka, Y. Effect of Brazilian green propolis in patients with type 2 diabetes: A double-blind randomized placebo-controlled study. Biomed. Rep. 2015, 3, 355–360. [Google Scholar] [CrossRef]
- Rakvaag, E.; Dragsted, L.O. Acute effects of light and dark roasted coffee on glucose tolerance: A randomized, controlled crossover trial in healthy volunteers. Eur. J. Nutr. 2016, 55, 2221–2230. [Google Scholar] [CrossRef]
- Santangelo, C.; Filesi, C.; Varì, R.; Scazzocchio, B.; Filardi, T.; Fogliano, V.; D’Archivio, M.; Giovannini, C.; Lenzi, A.; Morano, S.; et al. Consumption of extra-virgin olive oil rich in phenolic compounds improves metabolic control in patients with type 2 diabetes mellitus: A possible involvement of reduced levels of circulating visfatin. J. Endocrinol. Investig. 2016, 39, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Yang, B.; Tan, J.; Jiang, J.; Li, D. Associations of dietary intakes of anthocyanins and berry fruits with risk of type 2 diabetes mellitus: A systematic review and meta-analysis of prospective cohort studies. Eur. J. Clin. Nutr. 2016, 70, 1360–1367. [Google Scholar] [CrossRef] [PubMed]
- Rienks, J.; Barbaresko, J.; Nöthlings, U. Association of isoflavone biomarkers with risk of chronic disease and mortality: A systematic review and meta-analysis of observational studies. Nutr. Rev. 2017, 75, 616–641. [Google Scholar] [CrossRef] [PubMed]
- Dias, R.; Oliveira, H.; Fernandes, I.; Simal-Gandara, J.; Perez-Gregorio, R. Recent advances in extracting phenolic compounds from food and their use in disease prevention and as cosmetics. Crit. Rev. Food Sci. Nutr. 2021, 61, 1130–1151. [Google Scholar] [CrossRef]
- Brenes, A.; Viveros, A.; Chamorro, S.; Arija, I. Use of polyphenol-rich grape by-products in monogastric nutrition. A review. Anim. Feed Sci. Technol. 2016, 211, 1–17. [Google Scholar] [CrossRef]
- Marín, L.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Bioavailability of dietary polyphenols and gut microbiota metabolism: Antimicrobial properties. BioMed Res. Int. 2015, 2015, 905215. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, I.; Faria, A.; Calhau, C.; de Freitas, V.; Mateus, N. Bioavailability of anthocyanins and derivatives. J. Funct. Foods 2014, 7, 54–66. [Google Scholar] [CrossRef]
- Czank, C.; Cassidy, A.; Zhang, Q.; Morrison, D.J.; Preston, T.; Kroon, P.A.; Botting, N.P.; Kay, C.D. Human metabolism and elimination of the anthocyanin, cyanidin-3-glucoside: A 13C-tracer study. Am. J. Clin. Nutr. 2013, 97, 995–1003. [Google Scholar] [CrossRef] [Green Version]
- Marques, C.; Fernandes, I.; Norberto, S.; Sá, C.; Teixeira, D.; de Freitas, V.; Mateus, N.; Calhau, C.; Faria, A. Pharmacokinetics of blackberry anthocyanins consumed with or without ethanol: A randomized and crossover trial. Mol. Nutr. Food Res. 2016, 60, 2319–2330. [Google Scholar] [CrossRef]
- Campos, F.; Peixoto, A.F.; Fernandes, P.A.R.; Coimbra, M.A.; Mateus, N.; de Freitas, V.; Fernandes, I.; Fernandes, A. The Antidiabetic Effect of Grape Pomace Polysaccharide-Polyphenol Complexes. Nutrients 2021, 13, 4495. [Google Scholar] [CrossRef]
- Gonçalves, R.; Mateus, N.; de Freitas, V. Inhibition of α-amylase activity by condensed tannins. Food Chem. 2011, 125, 665–672. [Google Scholar] [CrossRef]
- Xiao, D.; Sandhu, A.; Huang, Y.; Park, E.; Edirisinghe, I.; Burton-Freeman, B.M. The effect of dietary factors on strawberry anthocyanins oral bioavailability. Food Funct. 2017, 8, 3970–3979. [Google Scholar] [CrossRef] [PubMed]
- Ayua, E.O.; Nkhata, S.G.; Namaumbo, S.J.; Kamau, E.H.; Ngoma, T.N.; Aduol, K.O. Polyphenolic inhibition of enterocytic starch digestion enzymes and glucose transporters for managing type 2 diabetes may be reduced in food systems. Heliyon 2021, 7, e06245. [Google Scholar] [CrossRef]
- Muraki, I.; Imamura, F.; Manson, J.E.; Hu, F.B.; Willett, W.C.; van Dam, R.M.; Sun, Q. Fruit consumption and risk of type 2 diabetes: Results from three prospective longitudinal cohort studies. BMJ Br. Med. J. 2013, 347, f5001. [Google Scholar] [CrossRef] [Green Version]
- Prpa, E.J.; Corpe, C.P.; Atkinson, B.; Blackstone, B.; Leftley, E.S.; Parekh, P.; Philo, M.; Kroon, P.A.; Hall, W.L. Apple polyphenol-rich drinks dose-dependently decrease early-phase postprandial glucose concentrations following a high-carbohydrate meal: A randomized controlled trial in healthy adults and in vitro studies. J. Nutr. Biochem. 2020, 85, 108466. [Google Scholar] [CrossRef]
- Castro-Acosta, M.L.; Stone, S.G.; Mok, J.E.; Mhajan, R.K.; Fu, C.I.; Lenihan-Geels, G.N.; Corpe, C.P.; Hall, W.L. Apple and blackcurrant polyphenol-rich drinks decrease postprandial glucose, insulin and incretin response to a high-carbohydrate meal in healthy men and women. J. Nutr. Biochem. 2017, 49, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Hsia, D.S.; Zhang, D.J.; Beyl, R.S.; Greenway, F.L.; Khoo, C. Effect of daily consumption of cranberry beverage on insulin sensitivity and modification of cardiovascular risk factors in adults with obesity: A pilot, randomised, placebo-controlled study. Br. J. Nutr. 2020, 124, 577–585. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, F.; Yeo, J.D. Insoluble-Bound Phenolics in Food. Molecules 2016, 21, 1216. [Google Scholar] [CrossRef]
- Le Bourvellec, C.; Renard, C.M.G.C. Interactions between Polyphenols and Macromolecules: Quantification Methods and Mechanisms. Crit. Rev. Food Sci. Nutr. 2012, 52, 213–248. [Google Scholar] [CrossRef]
- Shi, Y.; Williamson, G. Quercetin lowers plasma uric acid in pre-hyperuricaemic males: A randomised, double-blinded, placebo-controlled, cross-over trial. Br. J. Nutr. 2016, 115, 800–806. [Google Scholar] [CrossRef] [Green Version]
- Jafarpour-Sadegh, F.; Montazeri, V.; Adili, A.; Esfehani, A.; Rashidi, M.-R.; Pirouzpanah, S. Consumption of Fresh Yellow Onion Ameliorates Hyperglycemia and Insulin Resistance in Breast Cancer Patients during Doxorubicin-Based Chemotherapy: A Randomized Controlled Clinical Trial. Integr. Cancer Ther. 2017, 16, 276–289. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.Y.; Fang, J.C.; Gao, Z.H.; Zhang, C.; Xie, S.Y. Higher intake of fruits, vegetables or their fiber reduces the risk of type 2 diabetes: A meta-analysis. J. Diabetes Investig. 2016, 7, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Pinto, P.; Santos, C.N. Worldwide (poly)phenol intake: Assessment methods and identified gaps. Eur. J. Nutr. 2017, 56, 1393–1408. [Google Scholar] [CrossRef] [PubMed]
- Mateos-Martín, M.L.; Pérez-Jiménez, J.; Fuguet, E.; Torres, J.L. Non-extractable proanthocyanidins from grapes are a source of bioavailable (epi)catechin and derived metabolites in rats. Br. J. Nutr. 2012, 108, 290–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Jiménez, J.; Serrano, J.; Tabernero, M.; Arranz, S.; Díaz-Rubio, M.E.; García-Diz, L.; Goñi, I.; Saura-Calixto, F. Bioavailability of phenolic antioxidants associated with dietary fiber: Plasma antioxidant capacity after acute and long-term intake in humans. Plant Foods Hum. Nutr. 2009, 64, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Houghton, M.J.; Kerimi, A.; Mouly, V.; Tumova, S.; Williamson, G. Gut microbiome catabolites as novel modulators of muscle cell glucose metabolism. FASEB J. 2019, 33, 1887–1898. [Google Scholar] [CrossRef] [Green Version]
- Marques, C.; Fernandes, I.; Meireles, M.; Faria, A.; Spencer, J.P.E.; Mateus, N.; Calhau, C. Gut microbiota modulation accounts for the neuroprotective properties of anthocyanins. Sci. Rep. 2018, 8, 11341. [Google Scholar] [CrossRef] [Green Version]
- Saura-Calixto, F. The Story of the Introduction of Non-extractable Polyphenols into Polyphenol Research: Origin, Development and Perspectives. In Non-Extractable Polyphenols and Carotenoids: Importance in Human Nutrition and Health; The Royal Society of Chemistry: London, UK, 2018; Chapter 1; pp. 1–16. [Google Scholar] [CrossRef]
- Crous-Bou, M.; Fung, T.T.; Prescott, J.; Julin, B.; Du, M.; Sun, Q.; Rexrode, K.M.; Hu, F.B.; De Vivo, I. Mediterranean diet and telomere length in Nurses’ Health Study: Population based cohort study. BMJ 2014, 349, g6674. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Jiménez, J.; Díaz-Rubio, M.E.; Saura-Calixto, F. Non-extractable polyphenols, a major dietary antioxidant: Occurrence, metabolic fate and health effects. Nutr. Res. Rev. 2013, 26, 118–129. [Google Scholar] [CrossRef] [Green Version]
- González-Sarrías, A.; Espín, J.C.; Tomás-Barberán, F.A. Non-extractable polyphenols produce gut microbiota metabolites that persist in circulation and show anti-inflammatory and free radical-scavenging effects. Trends Food Sci. Technol. 2017, 69, 281–288. [Google Scholar] [CrossRef]
- Bao, Y.; Wang, W.; Zhou, Z.; Sun, C. Benefits and risks of the hormetic effects of dietary isothiocyanates on cancer prevention. PLoS ONE 2014, 9, e114764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petkovic, M.; Leal, E.C.; Alves, I.; Bose, C.; Palade, P.T.; Singh, P.; Awasthi, S.; Børsheim, E.; Dalgaard, L.T.; Singh, S.P.; et al. Dietary supplementation with sulforaphane ameliorates skin aging through activation of the Keap1-Nrf2 pathway. J. Nutr. Biochem. 2021, 98, 108817. [Google Scholar] [CrossRef]
- L Suraweera, T.; Rupasinghe, H.P.V.; Dellaire, G.; Xu, Z. Regulation of Nrf2/ARE Pathway by Dietary Flavonoids: A Friend or Foe for Cancer Management? Antioxidants 2020, 9, 973. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, A.; Manda, G.; Hassan, A.; Alcaraz, M.J.; Barbas, C.; Daiber, A.; Ghezzi, P.; León, R.; López, M.G.; Oliva, B.; et al. Transcription Factor NRF2 as a Therapeutic Target for Chronic Diseases: A Systems Medicine Approach. Pharm. Rev. 2018, 70, 348–383. [Google Scholar] [CrossRef] [Green Version]
- Afsar, B.; Afsar, R.E.; Ertuglu, L.A.; Covic, A.; Kanbay, M. Nutrition, Immunology, and Kidney: Looking Beyond the Horizons. Curr. Nutr. Rep. 2022. [Google Scholar] [CrossRef] [PubMed]
- Jubaidi, F.F.; Zainalabidin, S.; Taib, I.S.; Hamid, Z.A.; Budin, S.B. The Potential Role of Flavonoids in Ameliorating Diabetic Cardiomyopathy via Alleviation of Cardiac Oxidative Stress, Inflammation and Apoptosis. Int. J. Mol. Sci. 2021, 22, 5094. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-R.; Li, G.-H.; Zhou, M.-X.; Xiang, L.; Ren, D.-M.; Lou, H.-X.; Wang, X.-N.; Shen, T. Discovery of natural flavonoids as activators of Nrf2-mediated defense system: Structure-activity relationship and inhibition of intracellular oxidative insults. Bioorganic Med. Chem. 2018, 26, 5140–5150. [Google Scholar] [CrossRef]
- Oliveira, H.; Fernandes, A.; F Brás, N.; Mateus, N.; de Freitas, V.; Fernandes, I. Anthocyanins as Antidiabetic Agents—In Vitro and In Silico Approaches of Preventive and Therapeutic Effects. Molecules 2020, 25, 3813. [Google Scholar] [CrossRef]
- Carrizzo, A.; Izzo, C.; Forte, M.; Sommella, E.; Di Pietro, P.; Venturini, E.; Ciccarelli, M.; Galasso, G.; Rubattu, S.; Campiglia, P.; et al. A Novel Promising Frontier for Human Health: The Beneficial Effects of Nutraceuticals in Cardiovascular Diseases. Int. J. Mol. Sci. 2020, 21, 8706. [Google Scholar] [CrossRef]
- Wang, L.; Waltenberger, B.; Pferschy-Wenzig, E.M.; . Blunder, M.; Liu, X.; Malainer, C.; Blazevic, T.; Schwaiger, S.; Rollinger, J.M.; Heiss, E.H.; et al. Natural product agonists of peroxisome proliferator-activated receptor gamma (PPARγ): A review. Biochem. Pharm. 2014, 92, 73–89. [Google Scholar] [CrossRef] [Green Version]
- Yasmin, S.; Jayaprakash, V. Thiazolidinediones and PPAR orchestra as antidiabetic agents: From past to present. Eur. J. Med. Chem. 2017, 126, 879–893. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, H.C.; Hung, L.M.; Chen, J.K. Resveratrol, a red wine antioxidant, possesses an insulin-like effect in streptozotocin-induced diabetic rats. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E1339–E1346. [Google Scholar] [CrossRef] [Green Version]
- Penumathsa, S.V.; Thirunavukkarasu, M.; Zhan, L.; Maulik, G.; Menon, V.P.; Bagchi, D.; Maulik, N. Resveratrol enhances GLUT-4 translocation to the caveolar lipid raft fractions through AMPK/Akt/eNOS signalling pathway in diabetic myocardium. J. Cell. Mol. Med. 2008, 12, 2350–2361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palsamy, P.; Subramanian, S. Modulatory effects of resveratrol on attenuating the key enzymes activities of carbohydrate metabolism in streptozotocin-nicotinamide-induced diabetic rats. Chem. Biol. Interact. 2009, 179, 356–362. [Google Scholar] [CrossRef]
- Sharma, S.; Anjaneyulu, M.; Kulkarni, S.K.; Chopra, K. Resveratrol, a polyphenolic phytoalexin, attenuates diabetic nephropathy in rats. Pharmacology 2006, 76, 69–75. [Google Scholar] [CrossRef]
- Tian, Z.; Wang, J.; Xu, M.; Wang, Y.; Zhang, M.; Zhou, Y. Resveratrol Improves Cognitive Impairment by Regulating Apoptosis and Synaptic Plasticity in Streptozotocin-Induced Diabetic Rats. Cell Physiol. Biochem. 2016, 40, 1670–1677. [Google Scholar] [CrossRef]
- Wong, R.H.; Raederstorff, D.; Howe, P.R. Acute Resveratrol Consumption Improves Neurovascular Coupling Capacity in Adults with Type 2 Diabetes Mellitus. Nutrients 2016, 8, 425. [Google Scholar] [CrossRef] [Green Version]
- Hoseini, A.; Namazi, G.; Farrokhian, A.; Reiner, Ž.; Aghadavod, E.; Bahmani, F.; Asemi, Z. The effects of resveratrol on metabolic status in patients with type 2 diabetes mellitus and coronary heart disease. Food Funct. 2019, 10, 6042–6051. [Google Scholar] [CrossRef]
- Bashmakov, Y.K.; Assaad-Khalil, S.H.; Abou Seif, M.; Udumyan, R.; Megallaa, M.; Rohoma, K.H.; Zeitoun, M.; Petyaev, I.M. Resveratrol promotes foot ulcer size reduction in type 2 diabetes patients. ISRN Endocrinol. 2014, 2014, 816307. [Google Scholar] [CrossRef]
- Rungseesantivanon, S.; Thenchaisri, N.; Ruangvejvorachai, P.; Patumraj, S. Curcumin supplementation could improve diabetes-induced endothelial dysfunction associated with decreased vascular superoxide production and PKC inhibition. BMC Complement. Altern. Med. 2010, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- Hodaei, H.; Adibian, M.; Nikpayam, O.; Hedayati, M.; Sohrab, G. The effect of curcumin supplementation on anthropometric indices, insulin resistance and oxidative stress in patients with type 2 diabetes: A randomized, double-blind clinical trial. Diabetol. Metab. Syndr. 2019, 11, 41. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Chowdhury, S.; Sarkar, P.; Sil, P.C. Ameliorative role of ferulic acid against diabetes associated oxidative stress induced spleen damage. Food Chem. Toxicol. 2018, 118, 272–286. [Google Scholar] [CrossRef]
- Obafemi, T.O.; Jaiyesimi, K.F.; Olomola, A.A.; Olasehinde, O.R.; Olaoye, O.A.; Adewumi, F.D.; Afolabi, B.A.; Adewale, O.B.; Akintayo, C.O.; Ojo, O.A. Combined effect of metformin and gallic acid on inflammation, antioxidant status, endoplasmic reticulum (ER) stress and glucose metabolism in fructose-fed streptozotocin-induced diabetic rats. Toxicol. Rep. 2021, 8, 1419–1427. [Google Scholar] [CrossRef]
- Draganescu, D.; Andritoiu, C.; Hritcu, D.; Dodi, G.; Popa, M.I. Flaxseed Lignans and Polyphenols Enhanced Activity in Streptozotocin-Induced Diabetic Rats. Biology 2021, 10, 43. [Google Scholar] [CrossRef]
- Liu, Y.; Tian, X.; Gou, L.; Sun, L.; Ling, X.; Yin, X. Luteolin attenuates diabetes-associated cognitive decline in rats. Brain Res. Bull. 2013, 94, 23–29. [Google Scholar] [CrossRef]
- Sandireddy, R.; Yerra, V.G.; Komirishetti, P.; Areti, A.; Kumar, A. Fisetin Imparts Neuroprotection in Experimental Diabetic Neuropathy by Modulating Nrf2 and NF-κB Pathways. Cell. Mol. Neurobiol. 2016, 36, 883–892. [Google Scholar] [CrossRef]
- Coskun, O.; Kanter, M.; Korkmaz, A.; Oter, S. Quercetin, a flavonoid antioxidant, prevents and protects streptozotocin-induced oxidative stress and beta-cell damage in rat pancreas. Pharm. Res. 2005, 51, 117–123. [Google Scholar] [CrossRef]
- Sharifzadeh, M.; Ranjbar, A.; Hosseini, A.; Khanavi, M. The Effect of Green Tea Extract on Oxidative Stress and Spatial Learning in Streptozotocin-diabetic Rats. Iran. J. Pharm. Res 2017, 16, 201–209. [Google Scholar]
- Rostami, A.; Khalili, M.; Haghighat, N.; Eghtesadi, S.; Shidfar, F.; Heidari, I.; Ebrahimpour-Koujan, S.; Eghtesadi, M. High-cocoa polyphenol-rich chocolate improves blood pressure in patients with diabetes and hypertension. ARYA Atheroscler. 2015, 11, 21–29. [Google Scholar]
- El-Marasy, S.A.; Abdallah, H.M.; El-Shenawy, S.M.; El-Khatib, A.S.; El-Shabrawy, O.A.; Kenawy, S.A. Anti-depressant effect of hesperidin in diabetic rats. Can. J. Physiol. Pharm. 2014, 92, 945–952. [Google Scholar] [CrossRef]
- Rajput, M.S.; Sarkar, P.D. Modulation of neuro-inflammatory condition, acetylcholinesterase and antioxidant levels by genistein attenuates diabetes associated cognitive decline in mice. Chem. Biol. Interact. 2017, 268, 93–102. [Google Scholar] [CrossRef]
- Li, D.; Zhang, Y.; Liu, Y.; Sun, R.; Xia, M. Purified anthocyanin supplementation reduces dyslipidemia, enhances antioxidant capacity, and prevents insulin resistance in diabetic patients. J. Nutr. 2015, 145, 742–748. [Google Scholar] [CrossRef]
- Nizamutdinova, I.T.; Jin, Y.C.; Chung, J.I.; Shin, S.C.; Lee, S.J.; Seo, H.G.; Lee, J.H.; Chang, K.C.; Kim, H.J. The anti-diabetic effect of anthocyanins in streptozotocin-induced diabetic rats through glucose transporter 4 regulation and prevention of insulin resistance and pancreatic apoptosis. Mol. Nutr. Food Res. 2009, 53, 1419–1429. [Google Scholar] [CrossRef]
- Sarikaphuti, A.; Nararatwanchai, T.; Hashiguchi, T.; Ito, T.; Thaworanunta, S.; Kikuchi, K.; Oyama, Y.; Maruyama, I.; Tancharoen, S. Preventive effects of Morus alba L. anthocyanins on diabetes in Zucker diabetic fatty rats. Exp. Ther. Med. 2013, 6, 689–695. [Google Scholar] [CrossRef] [Green Version]
- Rojo, L.E.; Ribnicky, D.; Logendra, S.; Poulev, A.; Rojas-Silva, P.; Kuhn, P.; Dorn, R.; Grace, M.H.; Lila, M.A.; Raskin, I. In Vitro and in Vivo Anti-Diabetic Effects of Anthocyanins from Maqui Berry (Aristotelia chilensis). Food Chem. 2012, 131, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.J.; Rimando, A.M.; Fish, W.; Mentreddy, S.R.R.; Mathews, S.T. Serviceberry [Amelanchier alnifolia (Nutt.) Nutt. ex. M. Roem (Rosaceae)] leaf extract inhibits mammalian α-glucosidase activity and suppresses postprandial glycemic response in a mouse model of diet-induced obesity and hyperglycemia. J. Ethnopharmacol. 2012, 143, 481–487. [Google Scholar] [CrossRef]
- Movahed, A.; Nabipour, I.; Lieben Louis, X.; Thandapilly, S.J.; Yu, L.; Kalantarhormozi, M.; Rekabpour, S.J.; Netticadan, T. Antihyperglycemic effects of short term resveratrol supplementation in type 2 diabetic patients. Evid.-Based Complement. Altern. Med. 2013, 2013, 851267. [Google Scholar] [CrossRef] [Green Version]
- Brasnyó, P.; Molnár, G.A.; Mohás, M.; Markó, L.; Laczy, B.; Cseh, J.; Mikolás, E.; Szijártó, I.A.; Mérei, A.; Halmai, R.; et al. Resveratrol improves insulin sensitivity, reduces oxidative stress and activates the Akt pathway in type 2 diabetic patients. Br. J. Nutr. 2011, 106, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Zhou, R.; Wang, B.; Mi, M.T. Effect of resveratrol on glucose control and insulin sensitivity: A meta-analysis of 11 randomized controlled trials. Am. J. Clin. Nutr. 2014, 99, 1510–1519. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.; Xia, M.; Ma, J.; Hao, Y.; Liu, J.; Mou, H.; Cao, L.; Ling, W. Anthocyanin supplementation improves serum LDL- and HDL-cholesterol concentrations associated with the inhibition of cholesteryl ester transfer protein in dyslipidemic subjects. Am. J. Clin. Nutr. 2009, 90, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, D.W.; Tallent, R.; Navalta, J.W.; Salazar, A.; Lyons, T.J.; Basu, A. Effects of Acute Cocoa Supplementation on Postprandial Apolipoproteins, Lipoprotein Subclasses, and Inflammatory Biomarkers in Adults with Type 2 Diabetes after a High-Fat Meal. Nutrients 2020, 12, 1902. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, A.; Sandhu, A.K.; Edirisinghe, I.; Burton-Freeman, B.M. Functional Deficits in Gut Microbiome of Young and Middle-Aged Adults with Prediabetes Apparent in Metabolizing Bioactive (Poly)phenols. Nutrients 2020, 12, 3595. [Google Scholar] [CrossRef] [PubMed]
- Paquette, M.; Medina Larqué, A.S.; Weisnagel, S.J.; Desjardins, Y.; Marois, J.; Pilon, G.; Dudonné, S.; Marette, A.; Jacques, H. Strawberry and cranberry polyphenols improve insulin sensitivity in insulin-resistant, non-diabetic adults: A parallel, double-blind, controlled and randomised clinical trial. Br. J. Nutr. 2017, 117, 519–531. [Google Scholar] [CrossRef] [Green Version]
- Hoggard, N.; Cruickshank, M.; Moar, K.M.; Bestwick, C.; Holst, J.J.; Russell, W.; Horgan, G. A single supplement of a standardised bilberry (Vaccinium myrtillus L.) extract (36 % wet weight anthocyanins) modifies glycaemic response in individuals with type 2 diabetes controlled by diet and lifestyle. J. Nutr. Sci. 2013, 2, e22. [Google Scholar] [CrossRef] [Green Version]
- Borges, C.M.; Papadimitriou, A.; Duarte, D.A.; Lopes de Faria, J.M.; Lopes de Faria, J.B. The use of green tea polyphenols for treating residual albuminuria in diabetic nephropathy: A double-blind randomised clinical trial. Sci. Rep. 2016, 6, 28282. [Google Scholar] [CrossRef] [Green Version]
- Curtis, P.J.; van der Velpen, V.; Berends, L.; Jennings, A.; Feelisch, M.; Umpleby, A.M.; Evans, M.; Fernandez, B.O.; Meiss, M.S.; Minnion, M.; et al. Blueberries improve biomarkers of cardiometabolic function in participants with metabolic syndrome-results from a 6-month, double-blind, randomized controlled trial. Am. J. Clin. Nutr. 2019, 109, 1535–1545. [Google Scholar] [CrossRef] [Green Version]

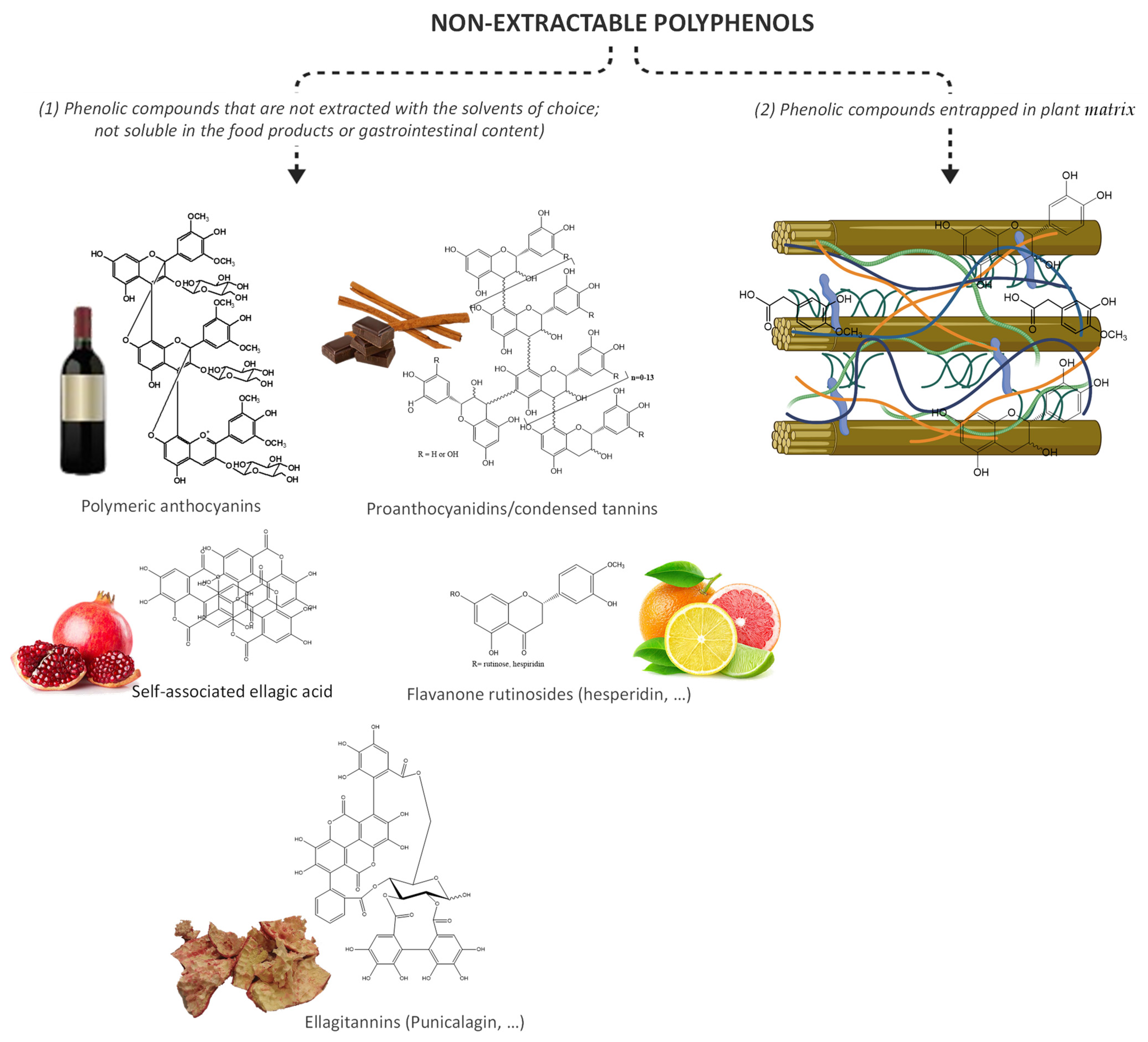


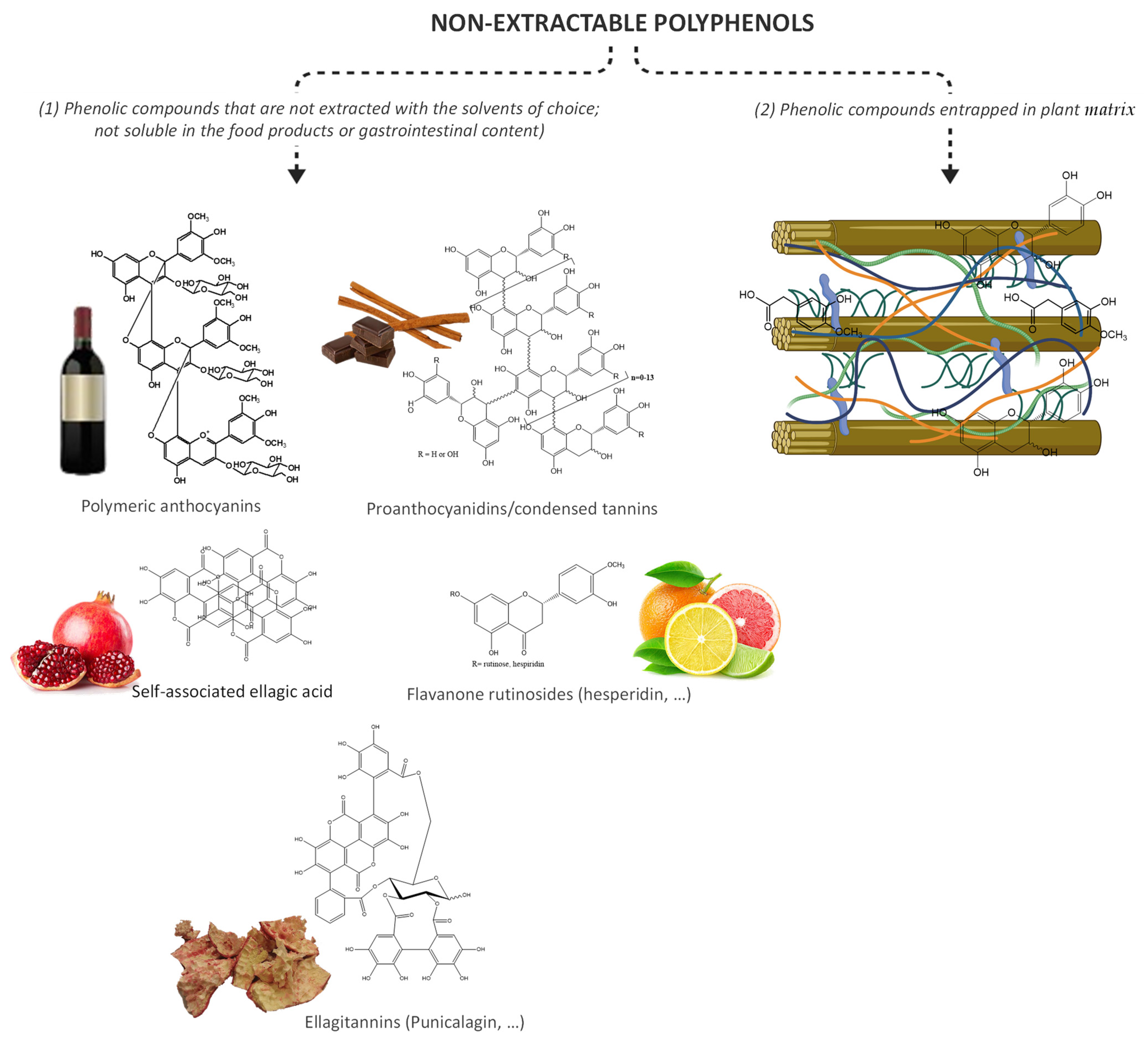
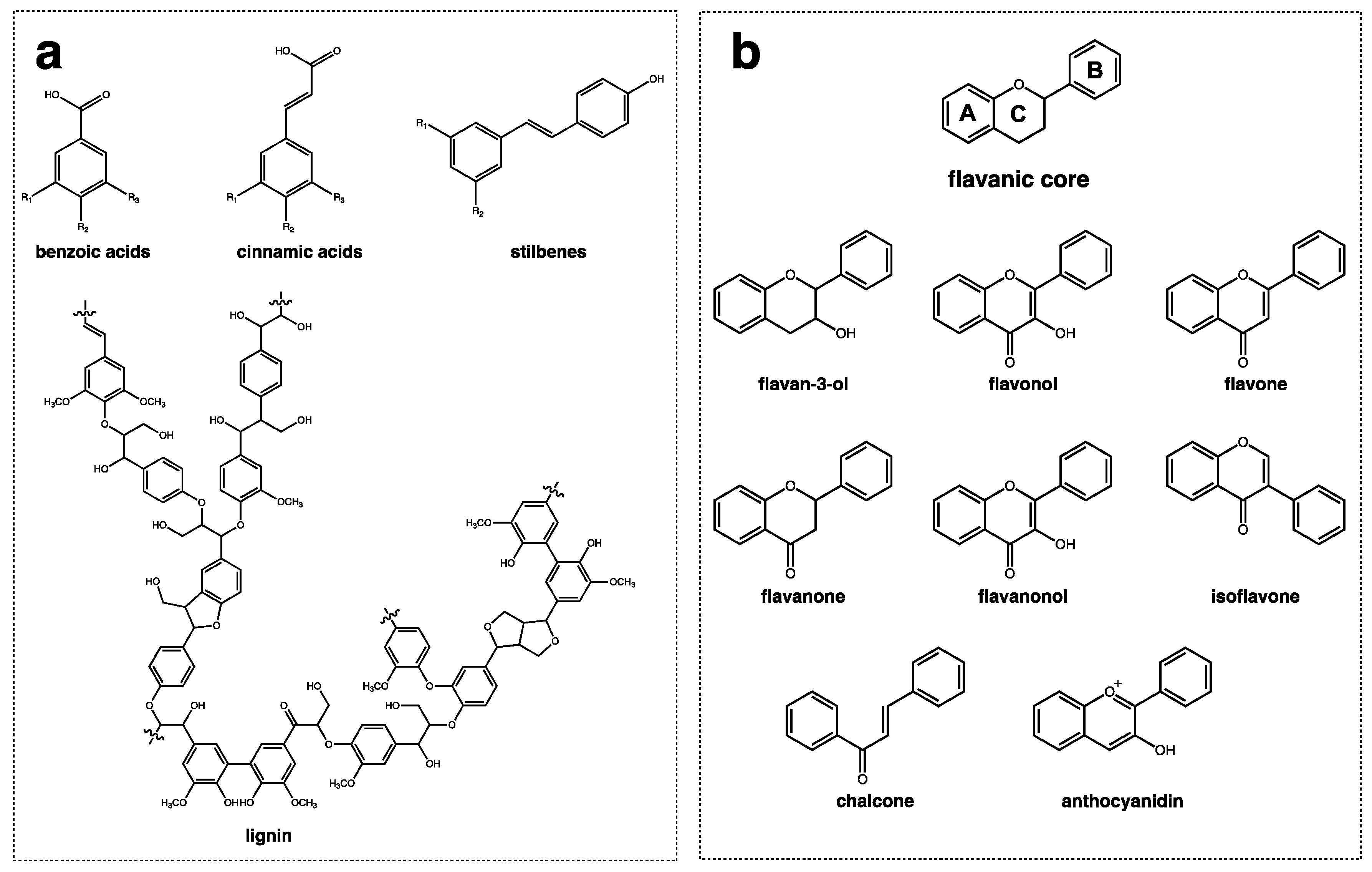{kind=link}
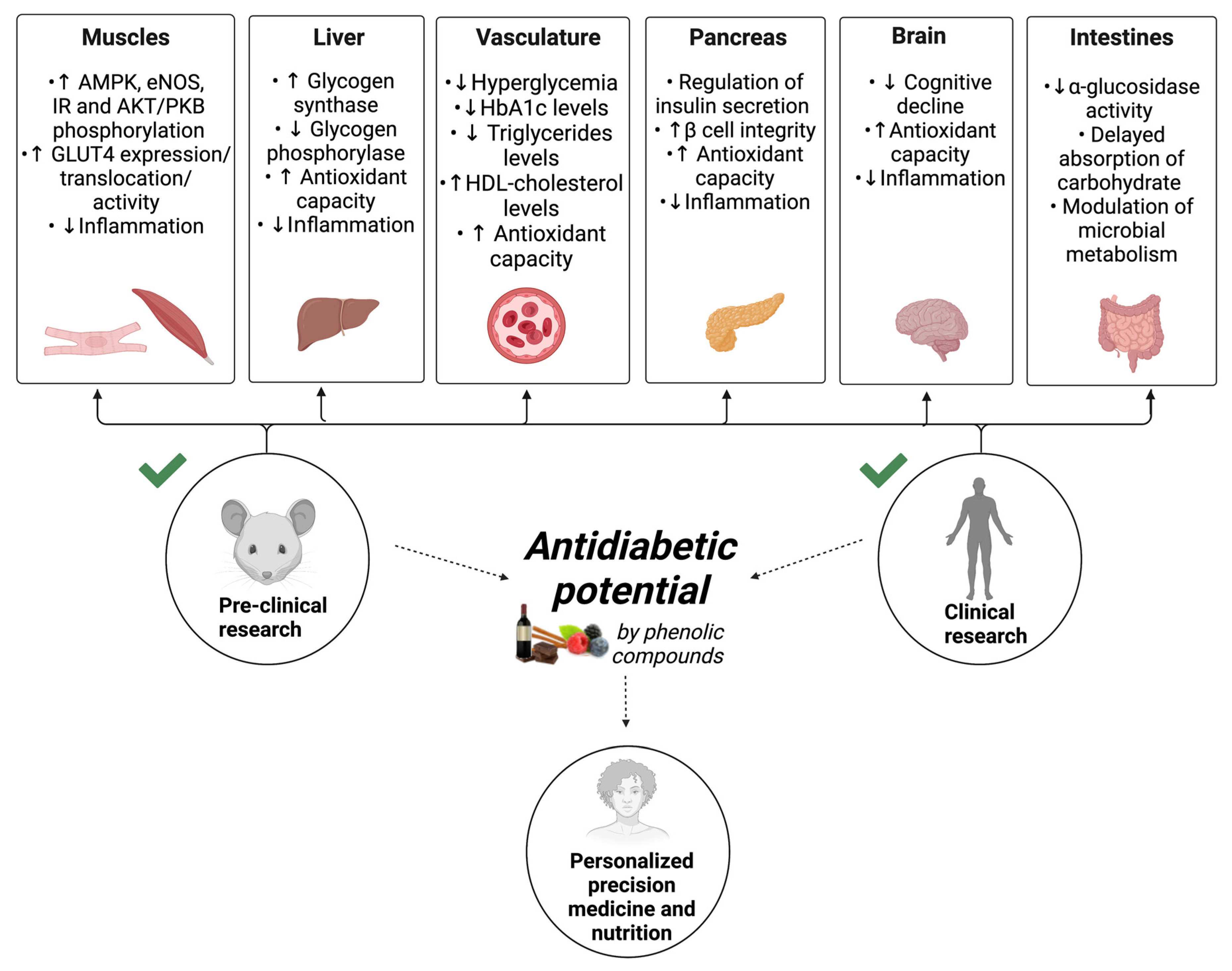
{kind=link}
{kind=link}
{kind=link}
| Phenolic Compounds Class | Compounds | Role in Diabetic Complications | Animal Models/ Diabetic Patients | Dose/Time | References |
|---|---|---|---|---|---|
| Stilbenes | Resveratrol | Reduced fed blood glucose levels, insulin concentration, plasma triglycerides | Streptozotocin– nicotinamide diabetic rats | 0.5 mg/kg body weight; 14 days | [108] |
| Increased phosphorylation of AMPK, eNOS and AKT/PKB, expression of GLUT4 in the myocardium | Streptozotocin- induced diabetic rats | 2.5 mg/kg body weight; 2 weeks | [109] | ||
| Decreased fasting blood glucose, HbA1c, increased insulin, improved hepatic glycogen content, increased glycogen synthase activity and reduced glycogen phosphorylase activity | Streptozotocin–nicotinamide diabetic rats | 5 mg/kg body weight; 30 days | [110] | ||
| Alleviated diabetic nephropathy by reduced renal dysfunction and oxidative stress | Streptozotocin-induced diabetic rats | 5 or 10 mg/kg body weight; 2 weeks | [111] | ||
| Ameliorated cognitive decline by inhibition of hippocampal apoptosis via the Bcl-2/Bax and caspase-3 pathway, improvement of synaptic dysfunction | Streptozotocin-induced diabetic rats | 80 mg/kg body weight; 4 weeks | [112] | ||
| Enhanced cerebral vasodilator function, improved cognitive performance | Patients with T2D | a single dose of 75 mg | [113] | ||
| Reduced fasting blood glucose, increased serum HDL-cholesterol levels, decreased total-/HDL-cholesterol ratio, increased total antioxidant capacity, decreased plasma MDA levels, upregulation of PPAR-γ and SIRT1 in PBMCs | Subjects with T2D and coronary heart disease | 500 mg/day; 4 weeks | [114] | ||
| Reduced foot ulcer size and plasma fibrinogen level | Patients with diabetic foot syndrome | 50 mg/ twice a day; 60 days | [115] | ||
| Phenolic acids | Curcumin (Hydroxycinnamic acid) | Improved diabetes-induced endothelial dysfunction through superoxide reduction and PKC inhibition | Streptozotocin-induced diabetic rats | 30 and 300 mg/kg body weight; 6 weeks | [116] |
| Reduced fasting blood glucose, reduced weight and BMI | Overweight patients with T2D | 1500 mg/3 times in a day; 10 weeks | [117] | ||
| Ferulic acid (Hydroxycinnamic acid) | Decreased fasting blood glucose levels, reduced level of serum insulin and spleen size. Reduced oxidative stress mediated inflammation and apoptosis | Streptozotocin-induced diabetic rats | 50 mg/kg body weight; 8 weeks | [118] | |
| Gallic acid (Hydrobenzoic acid) | Reduced fasting serum glucose and lipids, improved hepatic and pancreatic antioxidant capacity, lowered levels of IL-6 and TNF-α in liver and pancreas | Fructose-fed streptozotocin-induced diabetic rats | 50 mg/kg body weight; 21 days | [119] | |
| Lignans + Phenolic acids | Flaxseed extract (SDG, SECO, LARI, MATA, PINO + ferulic acid, GAE, p-coumaric acid) | Reduced fasting blood glucose, plasma cholesterol, LDL-cholesterol, triglycerides, plasma creatinine, urea and uric acid levels, partially recovers pancreas, liver, and kidney functions | Streptozotocin-induced diabetic rats | 0.774 mg/day of lignans + 0.073 mg/day of phenolic acids; 60 days | [120] |
| Flavonoids | Luteolin (Flavone) | Improved neuronal injury and cognition by attenuating oxidative stress | Streptozotocin-induced diabetic rats | 50 and 100 mg/kg body weight; 8 weeks | [121] |
| Fisetin (Flavonol) | Diabetic neuropathy modulation by improved motor nerve conduction velocity and reduced inflammation in sciatic nerves by NF-κB inhibition and Nrf2-positive modulation | Streptozotocin-induced diabetic rats | 5 and 10 mg/kg body weight; 2 weeks | [122] | |
| Quercetin (Flavonol) | Reduced pancreatic tissue MDA levels, serum NO concentrations, increased SOD, GSHPx, and CAT enzyme activation in pancreatic homogenates, and preserved pancreatic β-cell integrity | Streptozotocin-induced diabetic rats | 15 mg/kg body weight; 4 weeks | [123] | |
| Total green tea extract (Flavanol) | Reduced fasting blood glucose level, increased total antioxidant capacity and thiol groups in blood | Streptozotocin-induced diabetic rats | 3 mg/L through drinking water; 8 weeks | [124] | |
| Cocoa (Flavanol) | Decreased fasting plasma glucose, HbA1c, and blood pressure levels | Individuals with T2D and hypertension | 450 mg/day; 8 weeks | [125] | |
| Hesperidin (Flavanone) | Attenuated streptozotocin-induced neurochemical alterations, increased norepinephrine, dopamine, and serotonin levels, decreased MDA, increased GSH, and decreased IL-6 in brain | Streptozotocin-induced diabetic rats | 25, 50 or 100 mg/kg body mass; 21 consecutive days | [126] | |
| Genistein (Isoflavone) | Reduced hyperglycemia, improved cognition by restoring acetylcholinesterase activity and ameliorated neuroinflammation via decreasing TNF-α, IL-1β, and nitrites in brain | Streptozotocin-induced diabetic mice | 2.5, 5 and 10 mg/kg body weight; 30 days | [127] | |
| Cyanidin, delphinidin, petunidin, peonidin, malvidin extract (Anthocyanidins) | Reduced fasting plasma glucose and HbA1c levels, elevated serum adiponectin and β-hydroxybutyrate concentrations, improved dyslipidemia, enhanced the antioxidant capacity measured in plasma | Patients with T2D | 160 mg/day; 24 weeks | [128] | |
| Cyanidin-3-glucoside, delphinidin-3-glucoside, and petunidin-3-glucoside extract (Anthocyanins) | Decreased fed blood glucose, triglycerides levels, enhanced GLUT4 expression and insulin receptor phosphorylation in heart and skeletal muscle, protected pancreatic tissue of apoptosis through regulation of caspase-3, Bax, and Bcl-2 proteins, suppressed MDA levels, and restored SOD and CAT activities in serum | Streptozotocin- induced diabetic rats. | 50 mg/kg body weight; 30 days | [129] | |
| Cyanidin 3-rutinoside, cyanidin 3-glucoside, pelargonidin 3-glucoside and pelargonidin 3-rutinoside extract (Anthocyanins) | Reduced fasting blood glucose, maintain insulin levels and β cell histology | Zucker diabetic fatty rats | 125 or 250 mg/kg body weight; 5 weeks | [130] | |
| Delphinidin and cyanidin extract (Anthocyanidins) | Decreased fasting blood glucose levels and improved glucose tolerance | Hyperglycemic obese mice fed high fat diet | 50−500 mg/kg body weight; 16 weeks | [131] | |
| Cyanidin extract (Anthocyanidins) | Inhibited intestinal α-glucosidase activity and decreased post-prandial glycemic response, delayed absorption of carbohydrates | Diet-induced obese and hyperglycemic mice | 50 or 100 mg/kg body weight 60 min prior to an oral gavage of sucrose, starch or glucose | [132] |
| Clinical Trial Identifier No. | Objective | Voluntary and Dose | Recruitment Status | Results (References) |
|---|---|---|---|---|
| NCT01886989 | Investigate the postprandial effects of cocoa supplementation in glucose and lipids, and surrogate markers of atherosclerosis in patients with T2D | 18 subjects taking one beverage of cocoa polyphenols (960 mg) reconstituted in water after a high-fat meal challenge | Completed | Cocoa decreased total very LDL and chylomicron particles and increased the concentration of total HDL particles over the 6 h postprandial phase. Serum IL-18 was decreased by cocoa vs. placebo. Polyphenol-rich cocoa lowered dyslipidemia and inflammation following a high-fat dietary challenge in adults with T2D [137]. |
| NCT03049631 | Assess the effects of regular consumption of red raspberries (RRB) with and without fructooligosaccharide (FOS) on the composition of the gut microbiota and characterize plasma and urine metabolite profiles | 20 insulin resistant and 10 healthy subjects were given 125 g/day of RRB or 125 g/d RRB + 8 g/d FOS for 4 weeks | Completed | Individuals with prediabetes and insulin resistance compared to the reference group: (1) enriched Blautia obeum and Blautia wexlerae and depleted Bacteroides dorei and Coprococcus eutactus. Akkermansia muciniphila and Bacteroides spp. were depleted in the lean PreDM-IR subset; and (2) impaired microbial catabolism of select (poly)phenols resulting in lower 3,8-dihydroxy-urolithin (urolithin A), phenyl-γ-valerolactones and phenolic acid concentrations (p < 0.05). Controlling for obesity revealed relationships with microbial species that serve as metagenomic markers of diabetes development and therapeutic targets [138]. |
| NCT01766570 | Measure the beneficial effects of an optimized berries extracts on diabetes and cardiovascular diseases prevention | 60 men and women who were assigned to a 6-week experimental period where they consumed the rich polyphenol berries extract mix (333 mg of polyphenols from strawberries and cranberries) | Completed | Rich polyphenol berries extract mix improved insulin sensitivity in overweight and obese non-diabetic, insulin-resistant human subjects but was not effective in improving other cardiometabolic risk factors [139]. |
| NCT04847999 | Verify glucose levels before and after consumption of Ross Chocolates’ blend of sweeteners dark chocolate and conventional chocolate in people with diabetes | Individuals with T1D or T2D (10 participants each) were given a Ross Chocolate or conventional sugar-sweetened dark chocolate bar | Completed | No result posted No publication |
| NCT02650726 | Investigate the effect of purified anthocyanins on high-density lipoprotein and endothelial function in subjects with T2D | 80 male and female patients were given daily dose of 320 mg anthocyanin for 24 weeks in a randomized double blinded placebo-controlled trial | Completed | No result posted No publication |
| NCT01245270 | Investigated the acute effect of a standardized bilberry extract on glucose metabolism in T2D | 8 male volunteers with T2D were given a single oral capsule of either 0.47 g standardized bilberry extract (36% wet weight anthocyanins) or placebo followed by a polysaccharide drink (equivalent to 75 g glucose) in a double blinded cross over intervention with a 2-week washout period | Completed | The ingestion of a concentrated bilberry extract reduces postprandial glycaemia and insulin in volunteers with T2D. The most likely mechanism for the lower glycemic response involves reduced rates of carbohydrate digestion and/or absorption [140]. |
| NCT01923597 | Determine the safety and effect of green tea polyphenols (epigallocatechin gallate) on residual albuminuria of diabetic patients with nephropathy | Patients received four capsules (one capsule = 200 mg of epigallocatechin gallate) of green tea extract per day or placebo for 3 months | Completed | Green tea polyphenols administration reduces albuminuria in diabetic patients receiving the maximum recommended dose of renin-angiotensin (RAS) inhibition. Reduction in podocyte apoptosis by activation of the WNT pathway may have contributed to this effect [141]. |
| NCT02035592 | Investigate dose-dependent impact of blueberry powder intake on insulin sensitivity and resistance, cardiovascular disease risk factors, and lung and cognitive function in a population with metabolic syndrome | 144 male and female subjects received 26 g of freeze-dried blueberry powder (equivalent to 2 portions of fresh blueberries) per day for 6 months | Completed | A daily intake of 1 cup of blueberries improved endothelial function, systemic arterial stiffness, and attenuated cyclic guanosine monophosphate concentrations. Reduced insulinemia and glucose levels, decreased total cholesterol, and improved HDL-cholesterol, fractions of HDL-particles and Apolipoprotein A1 [142]. |
| NCT04383639 | Evaluate the effects of a single intake of a mixture of cocoa and carob (rich in high-molecular-weight polyphenols) in postprandial metabolism in subjects with T2D | The subjects will receive, after overnight fasting, a high-fat, high-sugar breakfast. In treatment A, they will not receive any additional product; in treatment B, they will receive at the same time a mixture of cocoa and carob; in treatment C, they will receive the mixture of cocoa and carob 10 h before breakfast. A total of 6 blood samples will be collected during each visit: 0–30–60–120–180–240–270 min | Recruiting | |
| NCT02291250 | Investigated the acute affect blackcurrants on glucose metabolism in overweight/obese volunteers | 16 overweight/obese volunteers will give 200 g of blackcurrants (which contain anthocyanins) or greencurrants (which naturally contain no anthocyanins) | Recruiting | |
| NCT04419948 | Investigate the acute effect of oleocanthal rich extra-virgin olive oil on postprandial hyperglycemia and platelet activation of T2D patients | Non-insulin-dependent diabetic patients will be randomly assigned to consume in five different days white bread (50 g CHO) with butter, butter with ibuprofen, refined olive oil, and olive oil with oleocanthal (250 mg/kg 500 mg/kg) | Recruiting |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, I.; Oliveira, J.; Pinho, A.; Carvalho, E. The Role of Nutraceutical Containing Polyphenols in Diabetes Prevention. Metabolites 2022, 12, 184. https://doi.org/10.3390/metabo12020184
Fernandes I, Oliveira J, Pinho A, Carvalho E. The Role of Nutraceutical Containing Polyphenols in Diabetes Prevention. Metabolites. 2022; 12(2):184. https://doi.org/10.3390/metabo12020184
Chicago/Turabian StyleFernandes, Iva, Joana Oliveira, Aryane Pinho, and Eugenia Carvalho. 2022. "The Role of Nutraceutical Containing Polyphenols in Diabetes Prevention" Metabolites 12, no. 2: 184. https://doi.org/10.3390/metabo12020184
APA StyleFernandes, I., Oliveira, J., Pinho, A., & Carvalho, E. (2022). The Role of Nutraceutical Containing Polyphenols in Diabetes Prevention. Metabolites, 12(2), 184. https://doi.org/10.3390/metabo12020184