

Rhizophora mucronata Lam. (Mangrove) Bark Extract Reduces Ethanol-Induced Liver Cell Death and Oxidative Stress in Swiss Albino Mice: In Vivo and In Silico Studies

 , , ,
, , ,  , , , and
, , , and
Abstract
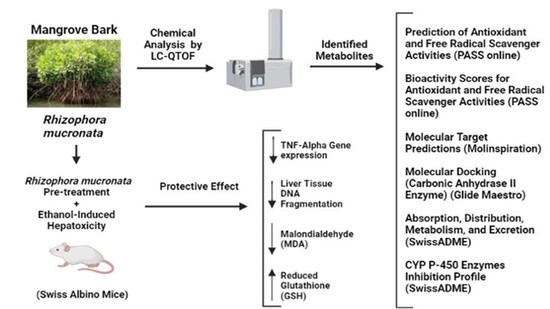
1. Introduction
2. Results
2.1. BERM Prevents Ethanol Intoxication-Induced Liver Necrosis and Injury
2.2. BERM Reduces Ethanol-Induced TNF-α Gene Expression Level and Upregulates an Ethanol-Suppressed NFR2 Gene Expression Level
2.3. BERM Reduces Ethanol-Induced Nuclear DNA Fragmentation
2.4. BERM Lowers Ethanol-Induced MDA Levels and Increases Ethanol-Decreased GSH Levels in Liver Tissues
2.5. Prediction of Antioxidant and Free Radical Scavenger Activities of BERM-Derived Metabolites Using Prediction of Activity Sprectra for Substances PASS Online Web Server
2.6. Molecular Target Predictions of BERM-Derived Metabolites Using the Molinspiration Web Server
2.7. Molecular Docking of BERM-Derived Metabolites into Carbonic Anhydrase (CA) II Enzyme
2.8. Predictions of Absorption, Distribution, Metabolism and Excretion (ADME) Properties for Identified Bioactive BERM-Derived Metabolites
2.9. Cytochrome P450 (CYP) Enzyme Inhibition Profile for the Identified Bioactive BERM-Derived Metabolites
2.10. Organ Toxicity Predictions for the Identified Bioactive BERM-Derived Metabolites
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Sample Collection and Extract Preparation
4.3. Animal Procurement and Maintenance
4.4. Animal Study Design and Sample Preparation
4.5. Histopathological Analysis
4.6. RNA Extraction and RT-PCR
4.7. TUNEL Assay
4.8. Oxidative Stress-Related Biochemical Assays
4.8.1. Estimation of Reduced Glutathione
4.8.2. Estimation of Malondialdehyde
4.9. Identification of BERM Metabolites Using LC-Q-TOF
4.10. Prediction of Antioxidant and Free Radical Scavenger Activity
4.11. Prediction of Molecular Target
4.12. Molecular Docking into CA II Enzyme
4.13. Predictions of ADME Properties
4.14. CYP Enzyme Inhibition Profiling for the Identified Bioactive Metabolites
4.15. Organ Toxicity and Safety Predictions
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Julien, J.; Ayer, T.; Bethea, E.D.; Tapper, E.B.; Chhatwal, J. Projected prevalence and mortality associated with alcohol-related liver disease in the USA, 2019–40: A modelling study. Lancet Public Heal. 2020, 5, e316–e323. [Google Scholar] [CrossRef]
- Vento, S.; Cainelli, F. Chronic liver diseases must be reduced worldwide: It is time to act. Lancet Glob. Health 2022, 10, e471–e472. [Google Scholar] [CrossRef]
- Crabb, D.W.; Im, G.Y.; Szabo, G.; Mellinger, J.L.; Lucey, M.R. Diagnosis and Treatment of Alcohol-Associated Liver Diseases: 2019 Practice Guidance From the American Association for the Study of Liver Diseases. Hepatology 2020, 71, 306–333. [Google Scholar] [CrossRef] [PubMed]
- Mellinger, J.L. Epidemiology of alcohol use and alcoholic liver disease. Clin. Liver Dis. (Hoboken) 2019, 13, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Arab, J.P.; Roblero, J.P.; Altamirano, J.; Bessone, F.; Araujo, R.C.; la Tijera, F.H.-D.; Restrepo-Gutiérrez, J.C.; Torre, A.; Urzua, A.; Simonetto, D.A.; et al. Alcohol-related liver disease: Clinical practice guidelines by the Latin American Association for the Study of the Liver (ALEH). Ann. Hepatol. 2019, 18, 518–535. [Google Scholar] [CrossRef]
- Birková, A.; Hubková, B.; Čižmárová, B.; Bolerázska, B. Current view on the mechanisms of alcohol-mediated toxicity. Int. J. Mol. Sci. 2021, 22, 9686. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, M.; Burnham, E.L.; DeVasure, J.M.; Sweeter, J.M.; Hunter, C.D.; Duryee, M.J.; Klassen, L.W.; Kharbanda, K.K.; Sisson, J.H.; Thiele, G.M.; et al. Malondialdehyde-Acetaldehyde (MAA) Protein Adducts Are Found Exclusively in the Lungs of Smokers with Alcohol Use Disorders and Are Associated with Systemic Anti-MAA Antibodies. Alcohol. Clin. Exp. Res. 2017, 41, 2093–2099. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, M.; Kharbanda, K.K.; Wyatt, T.A. Malondialdehyde-acetaldehyde-adducted surfactant protein alters macrophage functions through scavenger receptor A. Alcohol. Clin. Exp. Res. 2016, 40, 2563–2572. [Google Scholar] [CrossRef]
- Tan, H.K.; Yates, E.; Lilly, K.; Dhanda, A.D. Oxidative stress in alcohol-related liver disease. World J. Hepatol. 2020, 12, 332–349. [Google Scholar] [CrossRef]
- Chen, Y.; Singh, S.; Matsumoto, A.; Manna, S.K.; Abdelmegeed, M.A.; Golla, S.; Murphy, R.C.; Dong, H.; Song, B.-J.; Gonzalez, F.J.; et al. Chronic Glutathione Depletion Confers Protection against Alcohol-induced Steatosis: Implication for Redox Activation of AMP-activated Protein Kinase Pathway. Sci. Rep. 2016, 6, 29743. [Google Scholar] [CrossRef]
- Pérez-Hernández, O.; González-Reimers, E.; Quintero-Platt, G.; Abreu-González, P.; De La Vega-Prieto, M.J.; Sánchez-Pérez, M.J.; Martín-González, C.; Martínez-Riera, A.; Santolaria-Fernández, F. Malondialdehyde as a Prognostic Factor in Alcoholic Hepatitis. Alcohol Alcohol. 2016, 52, 305–310. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Miyata, T.; Nagy, L.E. Programmed cell death in alcohol-associated liver disease. Clin. Mol. Hepatol. 2020, 26, 618–625. [Google Scholar] [CrossRef]
- Li, C.; Li, J.; Xu, G.; Sun, H. Influence of chronic ethanol consumption on apoptosis and autophagy following transient focal cerebral ischemia in male mice. Sci. Rep. 2020, 10, 6164. [Google Scholar] [CrossRef]
- Zięba, S.; Maciejczyk, M.; Zalewska, A. Ethanol- and cigarette smoke-related alternations in oral redox homeostasis. Front. Physiol. 2022, 12, 793028. [Google Scholar] [CrossRef]
- Hamad, S.R.; Hamad Mohamed, H.R. Amelioration of ethanol-induced apoptotic DNA damage and ulcerative injuries in the mice gastric tissues by starch oral administration. Toxicol. Mech. Methods 2018, 28, 130–139. [Google Scholar] [CrossRef]
- Abdallah, M.A.; Singal, A.K. Mitochondrial dysfunction and alcohol-associated liver disease: A novel pathway and therapeutic target. Signal Transduct. Target. Ther. 2020, 5, 26. [Google Scholar] [CrossRef] [PubMed]
- Kamal, H.; Tan, G.C.; Ibrahim, S.F.; Shaikh, M.F.; Mohamed, I.N.; Mohamed, R.M.P.; Hamid, A.A.; Ugusman, A.; Kumar, J. Alcohol use disorder, neurodegeneration, Alzheimer’s and Parkinson’s disease: Interplay between oxidative stress, neuroimmune response and excitotoxicity. Front. Cell. Neurosci. 2020, 14, 282. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.K.; Meena, A.S.; Dalal, K.; Canelas, C.; Samak, G.; Pierre, J.F.; Rao, R. Chronic stress and cortisone exacerbate alcohol-induced tissue injury in the gut-liver-brain axis. Sci. Rep. 2021, 11, 826. [Google Scholar] [CrossRef]
- Mudd, T.W.; Guddati, A.K. Management of hepatotoxicity of chemotherapy and targeted agents. Am. J. Cancer Res. 2021, 11, 3461–3474. [Google Scholar] [PubMed]
- Anwanwan, D.; Singh, S.K.; Singh, S.; Saikam, V.; Singh, R. Challenges in liver cancer and possible treatment approaches. Biochim. Biophys. Acta Rev. Cancer 2020, 1873, 188314. [Google Scholar] [CrossRef]
- Ali, M.; Khan, T.; Fatima, K.; Ali, Q.U.A.; Ovais, M.; Khalil, A.T.; Ullah, I.; Raza, A.; Shinwari, Z.K.; Idrees, M. Selected hepatoprotective herbal medicines: Evidence from ethnomedicinal applications, animal models, and possible mechanism of actions. Phytother. Res. 2017, 32, 199–215. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, M.; Kannathasan, K.; Venkatesalu, V.; Prabhakar, K. Antibacterial activity of some salt marsh halophytes and mangrove plants against methicillin resistant Staphylococcus aureus. World J. Microbiol. Biotechnol. 2009, 25, 155–160. [Google Scholar] [CrossRef]
- Jairaman, C.; Yacoob, S.A.M.; Venkatraman, A.; Nagarajan, Y.; Murugesan, A.G. Propugnating Effect of Bark of Rhizophora mucronata Against Different Toxicants Viz Carbon Tetrachloride, Ethanol and Paracetamol on HepG2 Cell Lines. J. Pharmacopunct. 2019, 22, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Jiang, J.; Jing, Y.; Liu, W.; Yang, X.; Hou, X.; Gao, L.; Wei, L. The concentration of tumor necrosis factor-α determines its protective or damaging effect on liver injury by regulating Yap activity. Cell Death Dis. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Hong, Z.; Shao, S.; Li, L.; Yang, B.; Hou, Y.; Wang, H.; Xu, Y.; Zhang, Q.; Pi, J.; et al. Liver-specific Nrf2 deficiency accelerates ethanol-induced lethality and hepatic injury in vivo. Toxicol. Appl. Pharmacol. 2021, 426, 115617. [Google Scholar] [CrossRef]
- Wang, S.; Pacher, P.; De Lisle, R.C.; Huang, H.; Ding, W.-X. A Mechanistic Review of Cell Death in Alcohol-Induced Liver Injury. Alcohol. Clin. Exp. Res. 2016, 40, 1215–1223. [Google Scholar] [CrossRef]
- Joel, E.L.; Bhimba, V. Isolation and characterization of secondary metabolites from the mangrove plant Rhizophora mucronata. Asian Pacif. J Tropic. Med. 2010, 3, 602–604. [Google Scholar] [CrossRef]
- Mitra, S.; Naskar, N.; Chaudhuri, P. A review on potential bioactive phytochemicals for novel therapeutic applications with emphasis on mangrove species. Phytomedicine Plus 2021, 1, 100107. [Google Scholar] [CrossRef]
- Eswaraiah, G.; Peele, K.A.; Krupanidhi, S.; Kumar, R.B.; Venkateswarulu, T. Studies on phytochemical, antioxidant, antimicrobial analysis and separation of bioactive leads of leaf extract from the selected mangroves. J. King Saud Univ. Sci. 2020, 32, 842–847. [Google Scholar] [CrossRef]
- Chitra, J.; Yacoob, S.A.M.; Kumar, S.S.; Venkataraman, A.; Vijayaraghavan, R.; Nagarajan, Y. HPLC characterization, acute and sub-acute toxicity evaluation of bark extract of Rhizophora mucronata in Swiss Albino mice. Heliyon 2019, 6, e03108. [Google Scholar] [CrossRef] [PubMed]
- Unsal, V.; Cicek, M.; Sabancilar, I. Toxicity of carbon tetrachloride, free radicals and role of antioxidants. Rev. Environ. Health 2020, 36, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tan, H.-Y.; Wang, N.; Zhang, Z.-J.; Lao, L.; Wong, C.-W.; Feng, Y. The role of oxidative stress and antioxidants in liver diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Arellanes, M.A.; Gutiérrez-Rebolledo, G.A.; Meckes-Fischer, M.; León-Díaz, R. Medical plant extracts and natural compounds with a hepatoprotective effect against damage caused by antitubercular drugs: A review. Asian Pac. J. Trop. Med. 2016, 9, 1141–1149. [Google Scholar] [CrossRef]
- Gillessen, A.; Schmidt, H.H. Silymarin as supportive treatment in liver diseases: A narrative review. Adv. Ther. 2020, 37, 1279–1301. [Google Scholar] [CrossRef]
- Padmanabhan, P.; Jangle, S.N. Hepatoprotective activity of herbal preparation (Hp-4) against alcohol-induced hepatotoxicity in mice. Int. J. Appl. Sci. Biotechnol. 2014, 2, 50–58. [Google Scholar] [CrossRef]
- Nowak, A.J.; Relja, B. The impact of acute or chronic alcohol intake on the NF-κB signaling pathway in alcohol-related liver disease. Int. J. Mol. Sci. 2020, 21, 9407. [Google Scholar] [CrossRef] [PubMed]
- McClain, C.J.; Cohen, D.A. Increased tumor necrosis factor production by monocytes in alcoholic hepatitis. Hepatology 1989, 9, 349–351. [Google Scholar] [CrossRef]
- McClain, C.J.; Barve, S.; Deaciuc, I.; Kugelmas, M.; Hill, D. Cytokines in alcoholic liver disease. Semin. Liver Dis. 1999, 19, 205–219. [Google Scholar] [CrossRef]
- Wilson, G.K.; Tennant, D.A.; McKeating, J.A. Hypoxia inducible factors in liver disease and hepatocellular carcinoma: Current understanding and future directions. J. Hepatol. 2014, 61, 1397–1406. [Google Scholar] [CrossRef]
- Fouad, D.; Badr, A.; Attia, H.A. Hepatoprotective activity of raspberry ketone is mediated via inhibition of the NF-κB/TNF-α/caspase axis and mitochondrial apoptosis in chemically induced acute liver injury. Toxicol. Res. (Camb) 2019, 8, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Laissue, J.A.; Zimmermann, A. TUNEL-positive hepatocytes in alcoholic liver disease. A retrospective biopsy study using DNA nick end-labelling. Virchows Arch. 1997, 431, 337–344. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Sehrawat, T.S.; Verma, V.K.; Navarro-Corcuera, A.; Sidhu, G.; Mauer, A.; Luo, X.; Katsumi, T.; Chen, J.; Shah, S.; et al. XIAP knockdown in alcohol-associated liver disease models exhibits divergent in vitro and in vivo phenotypes owing to a potential zonal inhibitory role of SMAC. Front. Physiol. 2021, 12, 664222. [Google Scholar] [CrossRef]
- Saha, P.; Talukdar, A.D.; Nath, R.; Sarker, S.D.; Nahar, L.; Sahu, J.; Choudhury, M.D. Role of natural phenolics in hepatoprotection: A mechanistic review and analysis of regulatory network of associated genes. Front. Pharmacol. 2019, 10, 509. [Google Scholar] [CrossRef] [PubMed]
- Martemucci, G.; Costagliola, C.; Mariano, M. Free radical properties, sources and targets, antioxidant consumption and health. Oxygen 2022, 2, 48–78. [Google Scholar] [CrossRef]
- Chang, B.; Kim, H.; Kim, T.; Kim, S. Enzyme-treated Zizania latifolia extract protects against alcohol-induced liver injury by regulating the NRF2 pathway. Antioxidants 2021, 10, 960. [Google Scholar] [CrossRef]
- Brocardo, P.S.; Gil-Mohapel, J.; Wortman, R.; Noonan, A.; McGinnis, E.; Patten, A.R.; Christie, B.R. The effects of ethanol exposure during distinct periods of brain development on oxidative stress in the adult rat brain. Alcohol. Clin. Exp. Res. 2017, 41, 26–37. [Google Scholar] [CrossRef]
- Contreras-Zentella, M.; Villalobos-Garcia, D.; Hernández-Muñoz, R. Ethanol metabolism in the liver, the induction of oxidant stress, and the antioxidant defense system. Antioxidants 2022, 11, 1258. [Google Scholar] [CrossRef]
- Quinteros, D.A.; Hansen, A.W.; Bellaver, B.; Bobermin, L.D.; Pulcinelli, R.R.; Bandiera, S.; Caletti, G.; Bitencourt, P.E.R.; Quincozes-Santos, A.; Gomez, R. Combined exposure to alcohol and tobacco smoke changes oxidative, inflammatory, and neurotrophic parameters in different areas of the brain of rats. ACS Chem. Neurosci. 2019, 10, 1336–1346. [Google Scholar] [CrossRef]
- Şentürk, M.; Gülçin, I.; Beydemir, S.; Küfrevioğlu, O.I.; Supuran, C.T. In vitro inhibition of human carbonic anhydrase I and II isozymes with natural phenolic compounds. Chem. Biol. Drug Des. 2011, 77, 494–499. [Google Scholar] [CrossRef]
- Koz, O.; Ekinci, D.; Perrone, A.; Piacente, S.; Alankuş-Çalışkan, Ö.; Bedir, E.; Supuran, C.T. Analysis of saponins and phenolic compounds as inhibitors of α-carbonic anhydrase isoenzymes. J. Enzyme Inhib. Med. Chem. 2013, 28, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Alver, A.; Şentürk, A.; Çakirbay, H.; Menteşe, A.; Gökmen, F.; Keha, E.; Uçar, F. Carbonic anhydrase II autoantibody and oxidative stress in rheumatoid arthritis. Clin. Biochem. 2011, 44, 1385–1389. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Lu, X.; Feng, X.; Xuan, J.; Liu, F.; Feng, Z. Carbonic anhydrase inhibition, antioxidant activity against alveolar epithelial cells and antibacterial effect against Klebsiella pneumoniae enabled by synthesized silica nanoparticles through laser ablation technique. Life Sci. 2021, 278, 119032. [Google Scholar] [CrossRef] [PubMed]
- Göcer, H.; Akıncıoğlu, A.; Göksu, S.; Gülçin, I. Carbonic anhydrase inhibitory properties of phenolic sulfonamides derived from dopamine related compounds. Arab. J. Chem. 2017, 10, 398–402. [Google Scholar] [CrossRef]
- Imran, M.; Irfan, A.; Ibrahim, M.; Assiri, M.A.; Khalid, N.; Ullah, S.; Al-Sehemi, A.G. Carbonic anhydrase and cholinesterase inhibitory activities of isolated flavonoids from Oxalis corniculata L. and their first-principles investigations. Ind. Crops Prod. 2020, 148, 112285. [Google Scholar] [CrossRef]
- Honório, M.K.; Moda, T.L.; Andricopulo, A.D. Pharmacokinetic properties and in silico ADME modeling in drug discovery. Med. Chem. 2013, 9, 163–176. [Google Scholar] [CrossRef]
- Turner, P.V.; Hickman, D.L.; Van Luijk, J.; Ritskes-Hoitinga, M.; Sargeant, J.M.; Kurosawa, T.M.; Agui, T.; Baumans, V.; Choi, W.S.; Choi, Y.-K.; et al. Welfare impact of carbon dioxide euthanasia on laboratory mice and rats: A systematic review. Front. Vet. Sci. 2020, 7, 411. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Moron, M.S.; Depierre, J.W.; Mannervik, B. Levels of glutathione, glutathione reductase and glutathione S-transferase activities in rat lung and liver. Biochim. Biophys. Acta 1979, 582, 67–78. [Google Scholar] [CrossRef]
- Högberg, J.; Larson, R.E.; Kristoferson, A.; Orrenius, S. NADPH-dependent reductase solubilized from microsomes by peroxidation and its activity. Biochem. Biophys. Res. Commun. 1974, 56, 836–842. [Google Scholar] [CrossRef]
- Filimonov, D.; Lagunin, A.A.; Gloriozova, T.A.; Rudik, A.; Druzhilovskii, D.S.; Pogodin, P.V.; Poroikov, V.V. Prediction of the biological activity spectra of organic compounds using the PASS online web resource. Chem. Heterocycl. Comp. 2014, 50, 444–457. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, P.; Eckert, A.O.; Schrey, A.K.; Preissner, R. Pro-Tox-II: A webserver for the prediction of toxicity of chemicals. Nucleic Acids Res. 2018, 46, W257–W263. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound/ Metabolite (Antioxidant Activity) | Probability of Being Active (Pa) | Probability of Being Inactive (Pi) |
|---|---|---|
| Silymarin | 0.859 | 0.003 |
| 1 | 0.222 | 0.045 |
| 2 | 0.147 | 0.106 |
| 3 | 0.280 | 0.027 |
| 4 | 0.283 | 0.026 |
| 5 | 0.594 | 0.005 |
| 6 | 0.278 | 0.028 |
| 7 | 0.469 | 0.008 |
| 8 | - | - |
| 9 | 0.181 | 0.068 |
| Compound/ Metabolite (Free radical scavenger) | (Pa) | (Pi) |
| Silymarin | 0.956 | 0.001 |
| 1 | 0.315 | 0.027 |
| 2 | 0.201 | 0.075 |
| 3 | - | - |
| 4 | 0.360 | 0.021 |
| 5 | 0.743 | 0.003 |
| 6 | 0.357 | 0.022 |
| 7 | 0.469 | 0.012 |
| 8 | - | - |
| 9 | - | - |
| Compound/ Metabolite | Molinspiration Scores | |
|---|---|---|
| Silymarin | G protein-coupled receptor (GPCR) ligand | 0.07 |
| Ion channel modulator | −0.05 | |
| Kinase inhibitor | 0.01 | |
| Nuclear receptor ligand | 0.16 | |
| Protease inhibitor | 0.02 | |
| Enzyme inhibitor | 0.23 | |
| 1 | GPCR ligand | 0.02 |
| Ion channel modulator | 0.06 | |
| Kinase inhibitor | −0.33 | |
| Nuclear receptor ligand | 0.08 | |
| Protease inhibitor | −0.04 | |
| Enzyme inhibitor | 0.18 | |
| 2 | GPCR ligand | −3.51 |
| Ion channel modulator | −3.34 | |
| Kinase inhibitor | −3.84 | |
| Nuclear receptor ligand | −2.89 | |
| Protease inhibitor | −3.43 | |
| Enzyme inhibitor | −2.82 | |
| 3 | GPCR ligand | 0.27 |
| Ion channel modulator | 0.11 | |
| Kinase inhibitor | −0.42 | |
| Nuclear receptor ligand | 0.85 | |
| Protease inhibitor | 0.15 | |
| Enzyme inhibitor | 0.52 | |
| 4 | GPCR ligand | 0.17 |
| Ion channel modulator | 0.07 | |
| Kinase inhibitor | −0.22 | |
| Nuclear receptor ligand | 0.23 | |
| Protease inhibitor | 0.07 | |
| Enzyme inhibitor | 0.27 | |
| 5 | GPCR ligand | −0.21 |
| Ion channel modulator | −0.30 | |
| Kinase inhibitor | −0.01 | |
| Nuclear receptor ligand | −0.01 | |
| Protease inhibitor | −0.47 | |
| Enzyme inhibitor | 0.11 | |
| 6 | GPCR ligand | 0.05 |
| Ion channel modulator | −0.04 | |
| Kinase inhibitor | −0.24 | |
| Nuclear receptor ligand | 0.14 | |
| Protease inhibitor | 0.02 | |
| Enzyme inhibitor | 0.12 | |
| 7 | GPCR ligand | −0.30 |
| Ion channel modulator | −0.21 | |
| Kinase inhibitor | −0.12 | |
| Nuclear receptor ligand | −0.18 | |
| Protease inhibitor | −0.52 | |
| Enzyme inhibitor | 0.03 | |
| 8 | GPCR ligand | −0.63 |
| Ion channel modulator | −0.15 | |
| Kinase inhibitor | −0.85 | |
| Nuclear receptor ligand | −0.66 | |
| Protease inhibitor | −0.35 | |
| Enzyme inhibitor | 0.20 | |
| 9 | GPCR ligand | 0.17 |
| Ion channel modulator | 0.21 | |
| Kinase inhibitor | −0.38 | |
| Nuclear receptor ligand | 0.65 | |
| Protease inhibitor | 0.18 | |
| Enzyme inhibitor | 0.58 | |
| Compound/Metabolite | Docking Score | Interactions with Amino Acid Residues |
|---|---|---|
| Silymarin | −6.267 | Asp72, Glu 69, Asn 67, Ile 91, Gln 92, His 94, Phe 131, and Zinc coordination |
| 1 | - | - |
| 2 | −4.381 | Zinc coordination |
| 3 | −2.857 | - |
| 4 | - | - |
| 5 | −5.520 | His94, Thr199, and Zinc coordination |
| 6 | −1.662 | Zinc coordination |
| 7 | −4.881 | Zinc coordination |
| 8 | −3.947 | Thr199, Thr200, and Zinc coordination |
| 9 | −3.085 | Asn62, Thr199, and Zinc coordination |
| Compound/Metabolite | Molecular Weight (g/mol) | Log Po/w | Log S | BBB Permeant | GI Absorption | Rule of Five (ROF) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SWISS ADME | QikProp | SWISS ADME | QikProp | SWISS ADME | QikProp | SWISS ADME | QikProp | SWISS ADME | QikProp (%) | SwissADME | |
| Silymarin | 482.44 | 482.443 | 1.71 | 1.855 | −4.50 Moderately soluble | −5.354 | No | −2 | Low | 63.029 | Yes; 0 violation |
| 1 | 256.42 | 256.428 | 5.55 | 5.271 | −5.31 Moderately soluble | −5.436 | Yes | −2 | High | 88.223 | Yes; 1 violation: MLOGP > 4.15 |
| 2 | 96.13 | 96.129 | 1.30 | 0.737 | −0.88 soluble | −0.055 | Yes | 1 | High | 92.904 | Yes; 0 violation |
| 3 | 426.72 | 426.724 | 8.02 | 7.025 | −6.74 Poorly soluble | −7.801 | No | 1 | Low | 100 | Yes; 1 violation: MLOGP > 4.15 |
| 4 | 282.46 | 282.465 | 6.11 | 6.003 | −5.39 Moderately soluble | −6.587 | No | −2 | High | 91.435 | Yes; 1 violation: MLOGP > 4.15 |
| 5 | 268.26 | 268.268 | 3.17 | 2.682 | −5.68 Moderately soluble | −3.874 | Yes | 0 | High | 100 | Yes; 0 violation |
| 6 | 324.54 | 324.546 | 6.98 | 7.307 | −6.51 Poorly soluble | −8.374 | No | −1 | Low | 100 | Yes; 1 violation: MLOGP > 4.15 |
| 7 | 222.24 | 222.243 | 3.46 | 3.55 | −6.13 Poorly soluble | −3.521 | Yes | 1 | High | 100 | Yes; 0 violation |
| 8 | 194.18 | 194.184 | −2.72 | −1.918 | 1.74 Soluble | −0.337 | No | −2 | Low | 50.691 | Yes; 0 violation |
| 9 | 436.62 | 436.631 | 3.93 | 3.809 | −3.39 soluble | −5.759 | No | −2 | High | 95.213 | Yes; 0 violation |
| Compound/Metabolite | CYP1A2 | CYP2C19 | CYP2C9 | CYP2D6 | CYP3A4 |
|---|---|---|---|---|---|
| Silymarin | No | No | No | No | Yes |
| 1 | Yes | No | Yes | No | No |
| 2 | No | No | No | No | No |
| 3 | No | No | No | No | No |
| 4 | Yes | No | Yes | No | No |
| 5 | Yes | Yes | No | Yes | Yes |
| 6 | Yes | No | No | No | No |
| 7 | Yes | Yes | No | No | No |
| 8 | No | No | No | No | No |
| 9 | No | No | No | No | No |
| Compound Name | Classification | |||||
|---|---|---|---|---|---|---|
| Organ Toxicity (% Probability) | Toxicity Endpoint (% Probability) | Oral Toxicity Prediction (Predicted Toxicity Class) | ||||
| Hepatotoxicity | Carcinogenicity | Immunotoxicity | Mutagenicity | Cytotoxicity |  | |
| Silymarin | 0.78 (inactive) | 0.72 (inactive) | 0.97 (active) | 0.69 (inactive) | 0.77 (inactive) | Class IV |
| 1 | 0.52 (inactive) | 0.63 (inactive) | 0.99 (inactive) | 1.0 (inactive) | 0.74 (inactive) | Class IV |
| 2 | 0.71 (inactive) | 0.75 (inactive) | 0.98 (inactive) | 0.92 (inactive) | 0.75 (inactive) | Class III |
| 3 | 0.91 (inactive) | 0.63 (inactive) | 0.57 (active) | 0.95 (inactive) | 0.97 (inactive) | Class IV |
| 4 | 0.55 (inactive) | 0.64 (inactive) | 0.99 (inactive) | 1.0 (inactive) | 0.71 (inactive) | Class II |
| 5 | 0.71 (inactive) | 0.53 (inactive) | 0.72 (inactive) | 0.68 (inactive) | 0.95 (inactive) | Class V |
| 6 | 0.59 (inactive) | 0.61 (inactive) | 0.91 (inactive) | 0.97 (inactive) | 0.59 (inactive) | Class V |
| 7 | 0.70 (inactive) | 0.69 (active) | 0.99 (inactive) | 0.54 (inactive) | 0.75 (active) | Class V |
| 8 | 0.96 (inactive) | 0.83 (inactive) | 0.99 (inactive) | 0.68 (inactive) | 0.84 (inactive) | Class VI |
| 9 | 0.60 (inactive) | 0.75 (inactive) | 0.57 (active) | 0.73 (inactive) | 0.75 (inactive) | Class V |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jairaman, C.; Alehaideb, Z.I.; Yacoob, S.A.M.; Alghamdi, S.S.; Suliman, R.S.; Venkataraman, A.; Alghanem, B.; Sivanesan, S.; Vijayaraghavan, R.; Rameshbabu, S.; et al. Rhizophora mucronata Lam. (Mangrove) Bark Extract Reduces Ethanol-Induced Liver Cell Death and Oxidative Stress in Swiss Albino Mice: In Vivo and In Silico Studies. Metabolites 2022, 12, 1021. https://doi.org/10.3390/metabo12111021
Jairaman C, Alehaideb ZI, Yacoob SAM, Alghamdi SS, Suliman RS, Venkataraman A, Alghanem B, Sivanesan S, Vijayaraghavan R, Rameshbabu S, et al. Rhizophora mucronata Lam. (Mangrove) Bark Extract Reduces Ethanol-Induced Liver Cell Death and Oxidative Stress in Swiss Albino Mice: In Vivo and In Silico Studies. Metabolites. 2022; 12(11):1021. https://doi.org/10.3390/metabo12111021
Chicago/Turabian StyleJairaman, Chitra, Zeyad I. Alehaideb, Syed Ali Mohamed Yacoob, Sahar S. Alghamdi, Rasha S. Suliman, Anuradha Venkataraman, Bandar Alghanem, Senthilkumar Sivanesan, Rajagopalan Vijayaraghavan, Saranya Rameshbabu, and et al. 2022. "Rhizophora mucronata Lam. (Mangrove) Bark Extract Reduces Ethanol-Induced Liver Cell Death and Oxidative Stress in Swiss Albino Mice: In Vivo and In Silico Studies" Metabolites 12, no. 11: 1021. https://doi.org/10.3390/metabo12111021
APA StyleJairaman, C., Alehaideb, Z. I., Yacoob, S. A. M., Alghamdi, S. S., Suliman, R. S., Venkataraman, A., Alghanem, B., Sivanesan, S., Vijayaraghavan, R., Rameshbabu, S., Pari, S. M., & Matou-Nasri, S. (2022). Rhizophora mucronata Lam. (Mangrove) Bark Extract Reduces Ethanol-Induced Liver Cell Death and Oxidative Stress in Swiss Albino Mice: In Vivo and In Silico Studies. Metabolites, 12(11), 1021. https://doi.org/10.3390/metabo12111021