
Collection and Storage of Human Plasma for Measurement of Oxylipins

 ,
,  and
and
Abstract
1. Introduction
2. Results
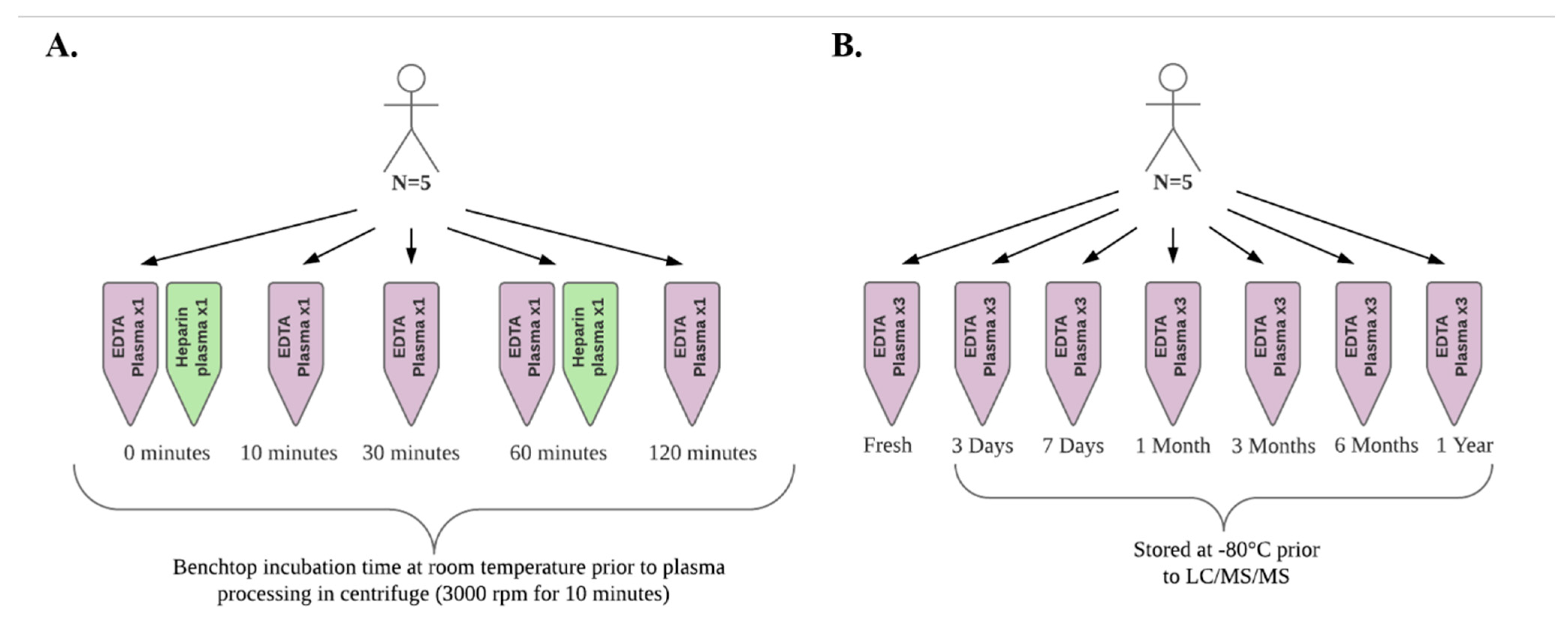
2.1. Sample Collection and Delayed Processing Experiment
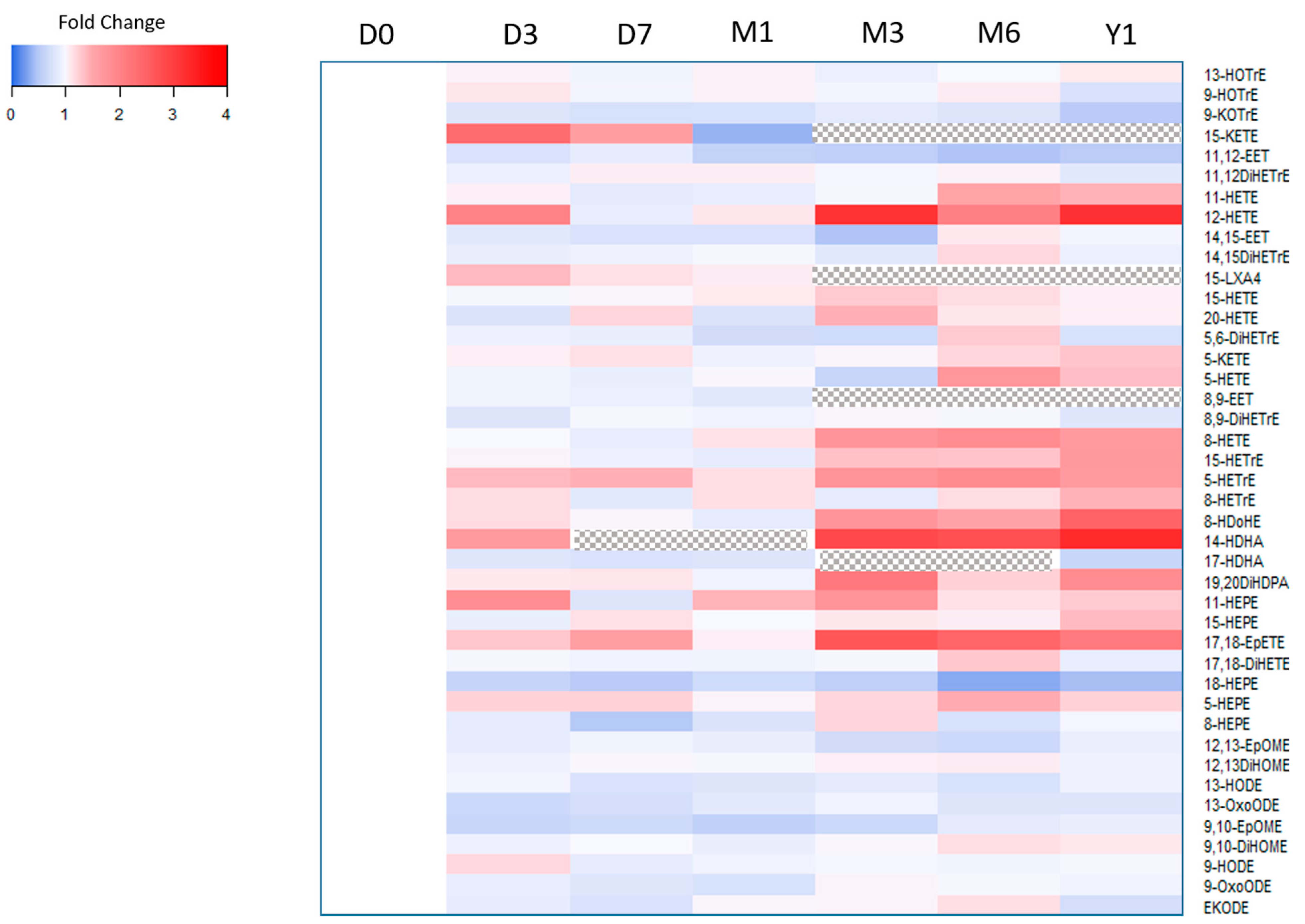
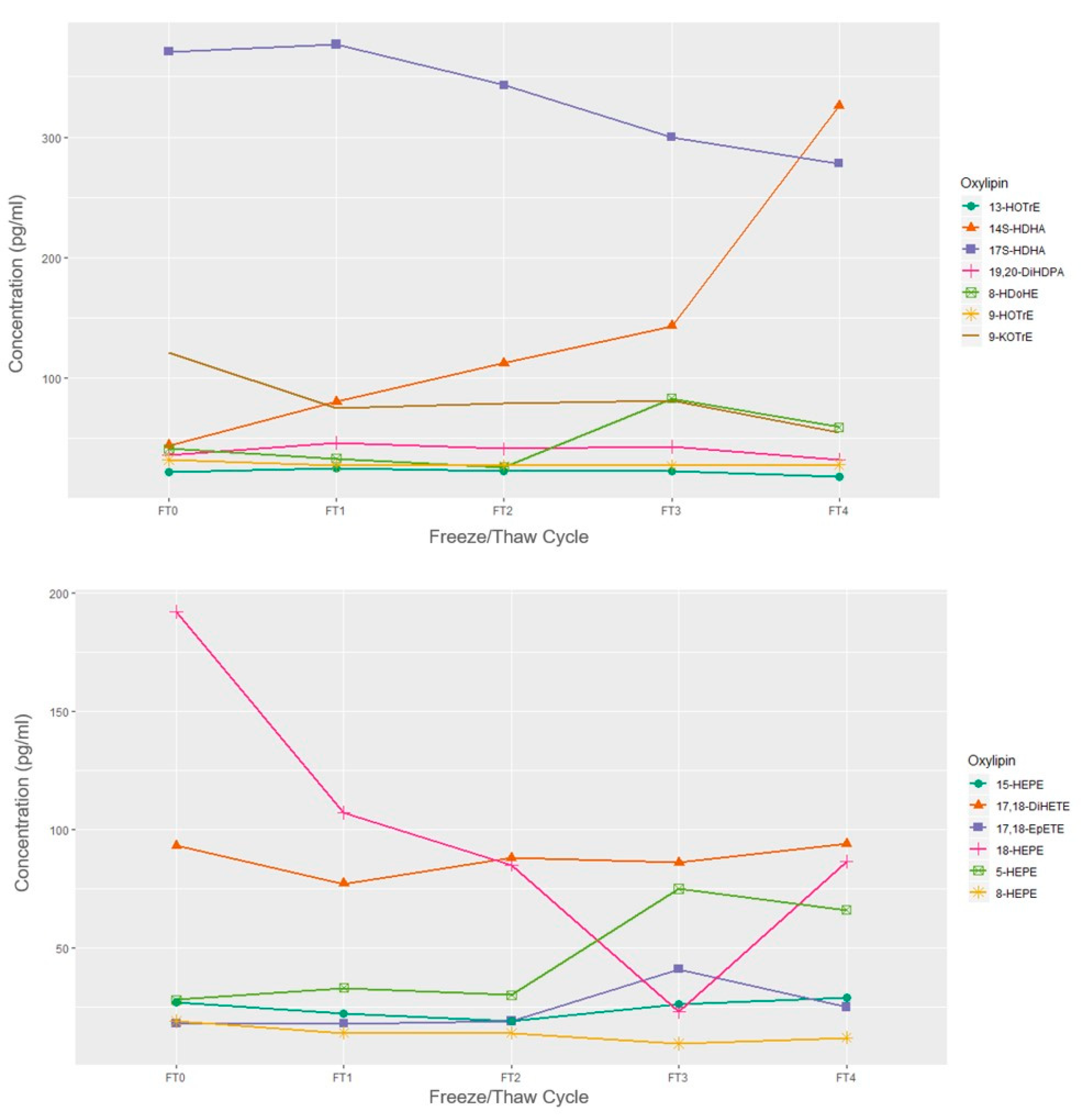
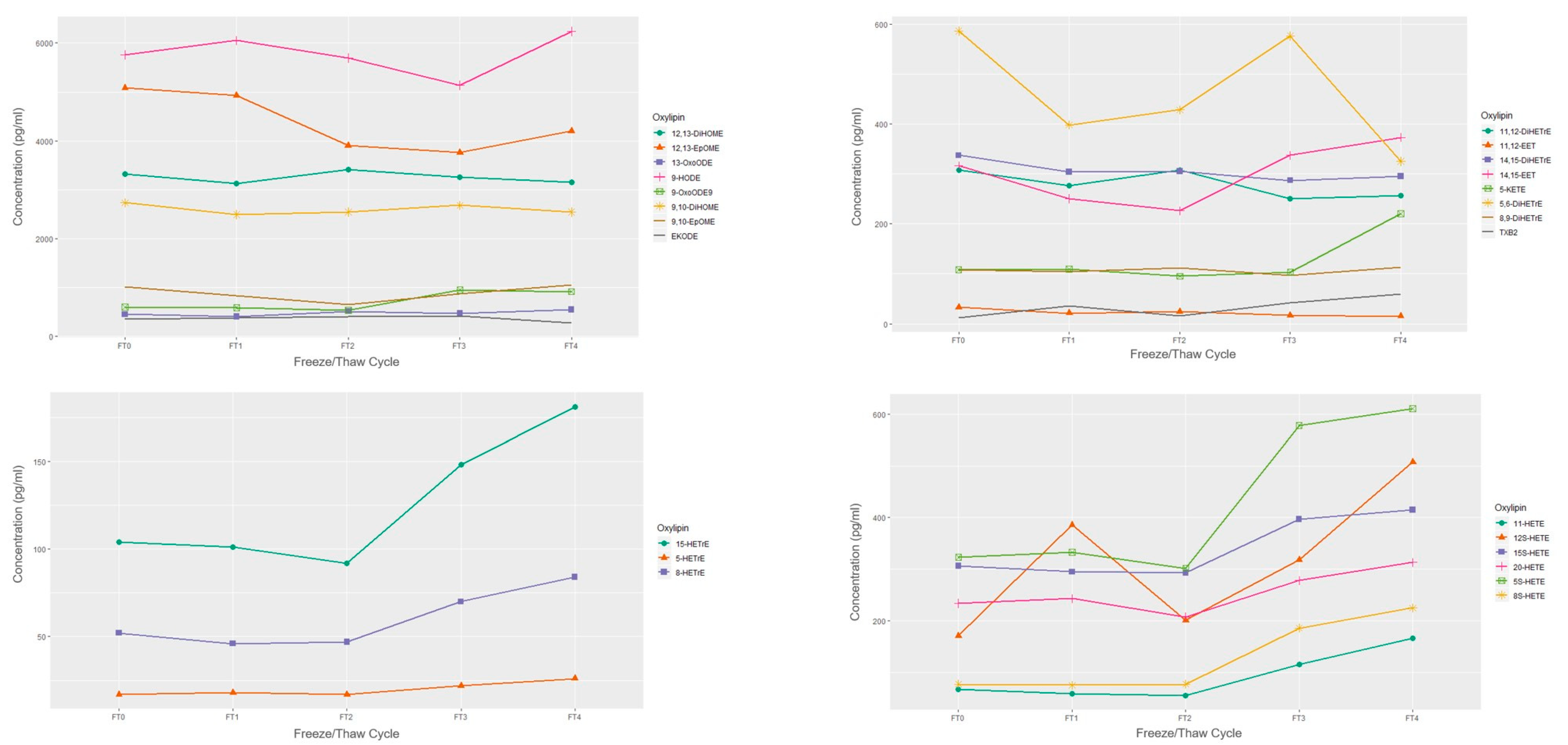
2.2. Sample Storage Experiment
Freeze/Thaw
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Delayed Processing Experiment
4.2. Sample Storage Experiment
4.2.1. Freeze/Thaw
4.2.2. Spiked Plasma
4.3. Oxylipin Sample Preparation
4.4. Liquid Chromatography-Tandem Mass Spectrometry
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gabbs, M.; Leng, S.; Devassy, J.G.; Monirujjaman; Aukema, H.M. Advances in Our Understanding of Oxylipins Derived from Dietary PUFAs. Adv. Nutr. 2015, 6, 513–540. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. Pro-resolving lipid mediators are leads for resolution physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Dalli, J.; Colas, R.A.; Walker, M.E.; Serhan, C.N. Lipid Mediator Metabolomics Via LC-MS/MS Profiling and Analysis. Methods Mol. Biol. 2018, 1730, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, M.; Liu, A.H.; Harbeck, R.; Reisdorph, R.; Rabinovitch, N.; Reisdorph, N. Leukotriene-E4 in human urine: Comparison of on-line purification and liquid chromatography–tandem mass spectrometry to affinity purification followed by enzyme immunoassay. J. Chromatogr. B 2009, 877, 3169–3174. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gladine, C.; Ostermann, A.I.; Newman, J.W.; Schebb, N.H. MS-based targeted metabolomics of eicosanoids and other oxylipins: Analytical and inter-individual variabilities. Free. Radic. Biol. Med. 2019, 144, 72–89. [Google Scholar] [CrossRef] [PubMed]
- Willenberg, I.; Ostermann, A.I.; Schebb, N.H. Targeted metabolomics of the arachidonic acid cascade: Current state and challenges of LC–MS analysis of oxylipins. Anal. Bioanal. Chem. 2015, 407, 2675–2683. [Google Scholar] [CrossRef] [PubMed]
- Kilburg-Basnyat, B.; Reece, S.W.; Crouch, M.J.; Luo, B.; Boone, A.D.; Yaeger, M.; Hodge, M.; Psaltis, C.; Hannan, J.L.; Manke, J.; et al. Specialized Pro-Resolving Lipid Mediators Regulate Ozone-Induced Pulmonary and Systemic Inflammation. Toxicol. Sci. 2018, 163, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Özdemir, R.B.; Gündüz, Ö.S.; Özdemir, A.T.; Akgül, Ö. Low levels of pro-resolving lipid mediators lipoxin-A4, resolvin-D1 and resolvin-E1 in patients with rheumatoid arthritis. Immunol. Lett. 2020, 227, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Dorow, J.; Becker, S.; Kortz, L.; Thiery, J.; Hauschildt, S.; Ceglarek, U. Preanalytical Investigation of Polyunsaturated Fatty Acids and Eicosanoids in Human Plasma by Liquid Chromatography–Tandem Mass Spectrometry. Biopreservation Biobanking 2016, 14, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Jonasdottir, H.S.; Brouwers, H.; Toes, R.E.; Ioan-Facsinay, A.; Giera, M. Effects of anticoagulants and storage conditions on clinical oxylipid levels in human plasma. Biochim. et Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2018, 1863, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Ramsden, C.E.; Yuan, Z.-X.; Horowitz, M.S.; Zamora, D.; Majchrzak-Hong, S.F.; Muhlhausler, B.S.; Taha, A.Y.; Makrides, M.; Gibson, R.A. Temperature and time-dependent effects of delayed blood processing on oxylipin concentrations in human plasma. Prostaglandins, Leukot. Essent. Fat. Acids 2019, 150, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Rund, K.M.; Nolte, F.; Doricib, J.; Greite, R.; Schott, S.; Lichtinghagen, R.; Gueler, F.; Schebb, N.H. Clinical blood sampling for oxylipin analysis—Effect of storage and pneumatic tube transport of blood on free and total oxylipin profile in human plasma and serum. Anal. 2020, 145, 2378–2388. [Google Scholar] [CrossRef] [PubMed]
- Koch, E.; Mainka, M.; Dalle, C.; Ostermann, A.I.; Rund, K.M.; Kutzner, L.; Froehlich, L.-F.; Bertrand-Michel, J.; Gladine, C.; Schebb, N.H. Stability of oxylipins during plasma generation and long-term storage. Talanta 2020, 217, 121074. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.S.; Ng’Alla, L.S. Heparin Therapy Leads to Platelet Activation and Prolongation of PFA-100 Closure Time. J. Cardiovasc. Pharmacol. Ther. 2005, 10, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Yeung, J.; Hawley, M.; Holinstat, M. The expansive role of oxylipins on platelet biology. J. Mol. Med. 2017, 95, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, M.; Manke, J.; Nkrumah-Elie, Y.; Shaikh, S.R.; Reisdorph, N. Improved quantification of lipid mediators in plasma and tissues by liquid chromatography tandem mass spectrometry demonstrates mouse strain specific differences. Prostaglandins Other Lipid Mediat. 2020, 151, 106483. [Google Scholar] [CrossRef] [PubMed]
- Kosaraju, R.; Guesdon, W.; Crouch, M.J.; Teague, H.L.; Sullivan, E.M.; Karlsson, E.A.; Schultz-Cherry, S.; Gowdy, K.; Bridges, L.C.; Reese, L.R.; et al. B Cell Activity Is Impaired in Human and Mouse Obesity and Is Responsive to an Essential Fatty Acid upon Murine Influenza Infection. J. Immunol. 2017, 198, 4738–4752. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author, Year | n | Quantification of Oxylipins (n) 1 | Storage Conditions | Delayed Plasma Processing Conditions | Freeze/Thaw Cycles | ||||
|---|---|---|---|---|---|---|---|---|---|
| Temperature | Duration | Sample Type(s) | Temperature | Duration | Sample Type(s) | ||||
| Present Study | 5 | non-esterified plasma oxylipins (46) | −80 °C | 3 days, 7 days, 1 month, 3 months, 6 months, and 1 year | EDTA plasma (no additives) | Room Temperature | 0, 10, 30, 60, and 120 min | pre-processed EDTA and heparin plasma (i.e., whole blood) | up to 4 cycles |
| Willenberg 2015 [6] | 1 | non-esterified plasma oxylipins (8) | Room temperature or ice | 5, 30, 60, and 120 min | pre-processed EDTA plasma (i.e., whole blood) | ||||
| Dorow, 2016 [9] | 6 | non-esterified plasma and serum oxylipins (28) | 4 °C or room temperature | 30, 60, 90, 120 min | EDTA plasma (with and without additives) | 4 ℃ or room temperature | 30, 60, 90, and 120 min | pre-processed EDTA plasma (i.e., whole blood) | up to 5 cycles |
| −20 °C, −80 °C, and −150 °C | 1 day, 1, 2, and 6 months | ||||||||
| Jonasdottir, 2018 [10] | 4 | non-esterified plasma oxylipins (18) | Room temperature | 2 and 8 h | EDTA and heparin plasma (with and without additives) | ||||
| 6 °C | 2, 8, and 24 h | ||||||||
| −20 °C | 24 h and 1 week or 1, 4, 12, 26, and 52 weeks | ||||||||
| −80 °C | 4, 12, 26, and 52 weeks | ||||||||
| Ramsden, 2019 [11] | 1 (on three separate occasions) | non-esterified plasma oxylipins (16) | Room Temperature or ice | 0, 10, 20, 30, 60, and 120 min | pre-processed EDTA plasma (i.e., whole blood) | ||||
| Gladine, 2019 [5] | 5 | non-esterified plasma oxylipins (4) | −80 °C | 2.5 years | EDTA plasma (no additives) | ||||
| Rund, 2020 [12] | 6 | non-esterified and total plasma and serum oxylipins (23) | 4 ℃ 2 | 4 h, 24 h, or after pneumatic tube system transport | pre-processed EDTA plasma and serum (i.e., whole blood) 2 | ||||
| Koch, 2020 [13] | 4 | non-esterified and total plasma oxylipins (133) | −80 °C | 1−6, 9, 12, and 15 months | EDTA plasma | 4 ℃ | 24 h | pre-processed EDTA plasma (i.e., whole blood) | |
| 20 ℃ | 4 h | processed EDTA plasma | |||||||
| −20 ℃ | 5 days | (with and without additives) | |||||||
| Oxylipin | T0 | T10 | T30 | T60 | T120 | p-Value * |
|---|---|---|---|---|---|---|
| 11,12-EET | 20.1 (11.4) | 26 (26.7) | 14.9 (9.8) | 17.7 (3.9) | 21.9 (15.5) | 0.92 |
| 11,12-DiHETrE | 222.4 (81.9) | 243.3 (10) | 226.3 (18.4) | 237.5 (39.8) | 250.4 (27.2) | 0.74 |
| 11-HETE | 103.0 (46) | 93.6 (27.3) | 92.7 (46.7) | 94.4 (62.6) | 90.9 (57.6) | 0.99 |
| 12-HETE | 491.7 (155.4) | 326.9 (28.7) | 281 (73.9) | 249.9 (152.9) | 177.3 (198.1) | 0.32 |
| 14,15-EET | 259.3 (100.1) | 218.6 (31.4) | 219.9 (33.3) | 273.1 (11.5) | 252.4 (27.5) | 0.65 |
| 14,15-DiHETrE | 199.0 (61) | 217.1 (24.9) | 206.5 (18.5) | 215.8 (39.8) | 224.9 (44.5) | 0.86 |
| 15-LXA4 | 11.5 (4.1) | 8.7 (1.8) | 8.95 (1.35) | 7.1 (0.6) | 8.5 (3.3) | 0.37 |
| 15-HETE | 394.4 (119.9) | 370.5 (246.3) | 431.1 (203.7) | 373.6 (135.1) | 422.1 (247.3) | 0.98 |
| 20-HETE | 152.8 (76) | 175.4 (18.2) | 209.4 (110.8) | 239.6 (80) | 220.4 (148.2) | 0.85 |
| 5,6-DiHETrE | 618.7 (177.1) | 740 (126.6) | 625.4 (196) | 641.4 (314.6) | 676.5 (38.9) | 0.90 |
| 5-KETE | 120.8 (54.8) | 114 (31.7) | 66.2 (21.3) | 100.9 (23.2) | 62 (34.1) | 0.54 |
| 5-HETE | 396.3 (230.6) | 307.2 (38.5) | 307.2 (23.2) | 336.7 (83.8) | 343.2 (83.8) | 0.88 |
| 8,9-DiHETrE | 115.6 (32.6) | 129.3 (19.8) | 129.6 (32.8) | 114.3 (34) | 136.1 (43.6) | 0.73 |
| 8-HETE | 135.2 (30.3) | 135.8 (22.7) | 107.7 (13.5) | 129 (27.9) | 131.3 (35.6) | 0.85 |
| 9-HETE | 91.2 (19.4) | 70.6 (45.3) | 103.2 (39) | 82.2 (46.6) | 109.7 (90.3) | 0.92 |
| LXA4 | 41.6 (11.6) | 31.4 (3.2) | 26.9 (4.6) | 31.3 (10.4) | 32 (7.2) | 0.16 |
| PGJ2 | 22.3 (6) | 16.5 (1.4) | 19.6 (4.1) | 17.9 (11) | 19.6 (11.7) | 0.94 |
| 12,13-EPOME | 3240.8 (2539) | 4255.7 (3280.1) | 2422.2 (3977.1) | 3827.6 (2512.7) | 3860.2 (1480.3) | 0.91 |
| 12,13-DiHOME | 3928.0 (2801.4) | 3670.7 (2915.3) | 3316.1 (3834.4) | 3861.5 (3385.7) | 3792 (3574.4) | 1.00 |
| 13-HODE | 35,458 (29.457) | 43,784 (52,306) | 29,965 (52,477) | 30,773(50,510) | 36,191 (39,458) | 1.00 |
| 13-OxoODE | 492.7 (447) | 646 (135.1) | 555.8 (353.5) | 451.2 (259.4) | 690.6 (191.6) | 0.82 |
| 9,10-EPOME | 515.8 (547.4) | 886.9 (636.4) | 427.7 (853.1) | 516.8 (549.4) | 891.6 (578.4) | 0.93 |
| 9,10-DiHOME | 2393.7 (1901.4) | 2463.7 (1799.4) | 2093.1 (1969.4) | 2379.6 (1748) | 2413 (1708.4) | 1.00 |
| 9-HODE | 4218.5 (1796.3) | 5098.4 (4391.3) | 3795.1 (5510.5) | 3748.4 (3828.9) | 3901.7 (5749.5) | 1.00 |
| 9-OxoODE | 1011.0 (399) | 1033 (238.7) | 756.2 (206.3) | 885.2 (250.7) | 895.3 (369.1) | 0.70 |
| EKODE | 429.1 (373.1) | 512.4 (448.8) | 395.7 (179.7) | 323.1 (413) | 406.5 (297.2) | 0.77 |
| 15-HETrE | 145.8 (86.3) | 142.2 (43.1) | 142.4 (48.2) | 135.3 (60.6) | 179.5 (76) | 0.99 |
| 5-HETrE | 42.5 (11.5) | 41.8 (10.7) | 38.7 (19.6) | 35.7 (13.5) | 42.1(19.8) | 0.99 |
| 8-HETrE | 35.3 (54.4) | 37.3 (33.3) | 41.6 (37.9) | 37.9 (38.3) | 34 (45.9) | 0.90 |
| 13-HOTrE | 23.4 (8) | 25.2 (16.3) | 21.2 (13.2) | 27.9 (15.5) | 24.3 (17.8) | 0.99 |
| 9-HOTrE | 22.6 (29.1) | 18.3 (18.7) | 17 (32.7) | 20.2 (29.6) | 24.6 (33) | 0.92 |
| 9-KOTrE | 46.0 (49.9) | 67.8 (11.9) | 61.6 (13.2) | 63.9 (61.6) | 59.05 (41.9) | 0.99 |
| 11-HDoHE | 106.3 (43.6) | 83.5 (88) | 84.3 (62.7) | 62.3 (29.8) | 50.1 (8.3) | 0.07 |
| 14-HDHA | 114.9 (42.9) | 105.4 (99.4) | 84.85 (30.3) | 103.1 (54.8) | 90.9 (42.1) | 0.67 |
| 17-HDHA | 447.8 (200.7) | 392.8 (143.8) | 378 (155.6) | 431.8 (172.9) | 427.6 (93.2) | 0.75 |
| 19,20-DiHDPA | 48.4 (33) | 54 (12.4) | 48.7 (27.4) | 34.8 (28.2) | 55.3 (51) | 0.70 |
| 8-HDoHE | 81.6 (23.5) | 57.2 (36.6) | 79.2 (19.3) | 72.2 (38.2) | 84 (7.8) | 0.83 |
| 15-HEPE | 37.3 (5.1) | 39.1 (4.3) | 32.5 (17.1) | 48.1 (17.6) | 37.8 (13.9) | 0.89 |
| 17,18-EpETE | 62.1 (11) | 60.7 (28.8) | 63.5 (15.5) | 63 (27.8) | 56.9 (14.8) | 0.98 |
| 17,18-DiHETE | 70.2 (43.4) | 83.3 (35.1) | 68.7 (16.4) | 84.5 (43.6) | 72.7 (36.7) | 0.95 |
| 18-HEPE | 189.2 (40.8) | 191.9 (22.2) | 162 (54.7) | 182.6 (40.6) | 164.2 (41.8) | 0.86 |
| 5-HEPE | 47.6 (21.2) | 81.1 (36) | 72.2 (32.6) | 51.4 (37.5) | 73.8 (31.1) | 0.96 |
| 8-HEPE | 25.0 (21.3) | 30.8 (17.5) | 33.7 (11.1) | 23 (26.3) | 27.5 (1.6) | 0.77 |
| Oxylipin | T0 | T60 | p-Value * | Oxylipin | T0 | T60 | p-Value * |
|---|---|---|---|---|---|---|---|
| 13-HOTrE | 18 (8.5) | 20.7 (9.4) | 0.60 | 5-HETrE | 53.7 (18.8) | 49.2 (21.9) | 0.92 |
| 9-HOTrE | 20.7 (9.4) | 14.9 (19.8) | 0.60 | 8-HETrE | 65.8 (34) | 56.4 (37.4) | 0.92 |
| 11-HDoHE | 472.2 (387.7) | 579.5 (334.5) | 0.92 | 11,12-EET | 44.5 (14.8) | 40.8 (22.5) | 0.75 |
| 14-HDHA | 400.3 (297.6) | 483.5 (292.8) | 0.92 | 11,12-DiHETrE | 215.6 (46.5) | 214.5 (17.9) | 0.92 |
| 17-HDHA | 426.3 (135.9) | 429.8 (77.1) | 0.92 | 11-HETE | 161.9 (21.4 | 125.1 (29.8) | 0.17 |
| 19,20-DiHDPA | 30.5 (17.6) | 27.7 (7.3) | 0.83 | 12-HETE | 1831.3 (502.7) | 1521.5 (148.8) | 0.25 |
| 8-HDoHE | 121.9 (63.5) | 137.4 (9.6) | 0.46 | 12-HHTrE | 223.6 (285.3) | 224 (374.3) | 0.62 |
| RVD1 | 3.7 (5.3) | 3.9 (6.9) | 0.71 | 14,15-EET | 800.7 (142.7) | 710.1 (80.9) | 0.25 |
| 15-HEPE | 30.8 (30) | 30.2 (17) | 0.46 | 14,15-DiHETrE | 217.8 (69.9) | 203.9 (77.3) | 0.75 |
| 17,18-EpETE | 112.6 (78.5) | 101.9 (29.2) | 0.92 | 15-LXA4 | 10.9 (3.6) | 6.9 (0.7) | 0.05 |
| 17,18-DiHETE | 52 (18.3) | 47.9 (30) | 0.92 | 15-HETE | 524.4 (144.2) | 446.4 (53) | 0.46 |
| 18-HEPE | 205.7 (14.7) | 205 (42.6) | 0.92 | 20-HETE | 116.2 (57.4) | 145.7 (156.6) | 0.35 |
| 5-HEPE | 141.3 (41.6) | 140 (18) | 0.46 | 5,6-DiHETrE | 1222.9 (301.4) | 1073.8 (151.6) | 0.75 |
| 8-HEPE | 30.7 (6.9) | 29.3 (7.8) | 0.92 | 5-KETE | 216.7 (37.3) | 136.9 (36.7) | 0.35 |
| 12,13-EPOME | 3720.8 (2129.7) | 3034.4 (2417.7) | 0.75 | 5-HETE | 1107.2 (305.1) | 1004.6 (216.6) | 0.46 |
| 12,13-DiHOME | 3244.4 (1975.2) | 3419.2 (2042) | 0.92 | 8,9-DiHETrE | 125.9 (32.3) | 133 (34.5) | 0.25 |
| 13-HODE | 35,908.6 (3337.9) | 32,104.2 (35,691.9) | 0.75 | 8-HETE | 161.9 (9) | 150.3 (24.5) | 0.75 |
| 13-OxoODE | 346.8 (138.1) | 389.9 (94.5) | 0.60 | 9-HETE | 109.3 (33.8) | 104.7 (2.7) | 0.92 |
| 9,10-EPOME | 478.3 (326.6) | 447.5 (389.2) | 0.35 | LXA4 | 32 (16.7) | 24.7 (1.8) | 0.60 |
| 9,10-DiHOME | 1915.6 (1331.8) | 2001.8 (1535.1) | 0.92 | LTB4 | 11.6 (17.7) | 34.8 (16.8) | 0.08 |
| 9-HODE | 4387.7 (1927.6) | 3572.2 (1589.7) | 0.60 | PGJ2 | 19.4 (5.1) | 16.8 (2.7) | 0.75 |
| 9-OxoODE | 940.9 (339) | 648 (158.8) | 0.17 | TXB2 | 88.7 (84.9) | 45.5 (49.5) | 0.60 |
| EKODE | 306.7 (99) | 166.7 (69.4) | 0.25 | 9-HEPE | 31.1 (23.8) | 44.1 (17.2) | 0.58 |
| Oxylipin | Fresh | Day 3 | Day 7 | Month 1 | Month 3 | Month 6 | Month 12 | p-Value * |
|---|---|---|---|---|---|---|---|---|
| 8-HDoHE | 29 (14) | 34 (12) | 29.5 (14) | 25 (12) | 51 (9) | 45 (15) | 72 (29) | 0.03 |
| 12-HHTrE | 58 (39) | 278 (309) | 100.75 (111) | 59 (85) | 359.5 (462) | 502 (581) | 293 (255) | 0.17 |
| 14-HDHA | 50 (22.5) | 84 (26) | 143 (25) | 138 (32) | 164.5 (60) | 0.04 | ||
| 15-KETE | 120.5 (63.5) | 285 (122) | 195 (52) | 46 (26) | NA | |||
| 11-HEPE | 7 (0) | 13 (6) | 5.5 (2.5) | 10 (3.5) | 12.5 (1) | 8 (2) | 9 (8) | 0.76 |
| 15-HEPE | 27 (17) | 24 (15) | 31 (12) | 27 (22) | 30 (8) | 29 (11) | 37 (13) | 0.68 |
| TXB2 | 12 (2) | 90.5 (148.5) | 17.5 (12.75) | 19 (10) | 49 (58) | 46 (81) | 87.25 (64) | 0.04 |
| 11,12-EET | 30 (10) | 23 (12) | 26 (5) | 18 (1) | 17 (11) | 14 (3) | 16 (3) | 0.03 |
| 11,12-DiHETrE | 234 (51) | 212 (79) | 251 (90) | 251 (83) | 226 (62) | 243 (23) | 193 (56) | 0.42 |
| 11-HETE | 60 (23) | 63 (15) | 51 (8) | 53 (17) | 59 (10) | 92 (46) | 86 (8) | 0.09 |
| 12,13-EpOME | 5082 (1910) | 4390 (1038) | 4806 (2415) | 4499 (1814) | 3526 (1163) | 3326 (81) | 4573 (1033) | 0.83 |
| 12,13-DiHOME | 2574 (2310) | 2390 (2479) | 2589 (2180) | 2546 (2276) | 2724 (2448) | 2787 (2155) | 2403 (2655) | 0.83 |
| 12-HETE | 196 (20) | 398 (143) | 172 (20) | 219 (47) | 623 (283) | 409 (54) | 627 (258) | 0.03 |
| 13-HODE | 39,497 (24,524) | 38,047 (21,533) | 30,455 (22,657) | 31,351 (27,704) | 33,346 (21,538) | 29,678 (15,572) | 36,535 (27,286) | 0.99 |
| 13-HOTrE | 22 (4) | 23 (5) | 21 (4) | 23 (3) | 20 (13) | 22 (17) | 24 (11) | 0.76 |
| 13-OxoODE | 452 (181) | 301 (17) | 329 (39) | 378 (116) | 423 (114) | 361 (86) | 361 (223) | 0.99 |
| 14,15-EET | 305 (56) | 254 (52) | 234 (92) | 235 (44) | 144 (27) | 337 (159) | 292 (45) | 0.68 |
| 14,15-DiHETrE | 229 (43) | 203 (93) | 213 (90) | 227 (111) | 190 (123) | 276 (104) | 208 (69) | 0.37 |
| 15-LXA4 | 13 (3) | 18 (3) | 15 (7) | 14 (4) | . | . | . | NA |
| 15-HETrE | 104 (46) | 107 (39) | 96 (46) | 90 (39) | 140 (48) | 138 (75) | 177 (48) | 0.09 |
| 15-HETE | 316 (53) | 310 (47) | 321 (43) | 346 (57) | 406 (51) | 370 (22) | 332 (22) | 0.99 |
| 17,18-EpETE | 20 (18) | 26 (10) | 32 (4) | 21 (9) | 53 (11) | 49 (66) | 43 (12) | 0.32 |
| 17-HDHA | 371 (72) | 299 (82) | 285 (75) | 295 (77) | . | . | 235 (82) | 0.06 |
| 17,18-DiHETE | 93 (30) | 91 (32) | 87 (27) | 88 (25) | 92 (9) | 121 (51) | 82 (5) | 0.48 |
| 18-HEPE | 266 (31) | 165 (80) | 141 (33) | 182 (80) | 149.5 (83) | 88 (29) | 116 (18.5) | 0.03 |
| 19,20-DiHDPA | 33 (13) | 36.5 (37.5) | 37 (17) | 31 (18) | 72 (113) | 41 (27) | 62 (147) | 0.4 |
| 20-HETE | 172 (121) | 134 (64) | 208 (110) | 133 (137) | 249 (76) | 192 (119) | 183 (110) | 0.99 |
| 5,6-DiHETrE | 586 (57) | 540 (205) | 520 (153) | 409 (125) | 400 (115) | 754 (582) | 436 (91) | 0.24 |
| 5-HEPE | 38 (21) | 47 (3) | 47 (18) | 39 (11) | 46 (31) | 56 (29) | 47 (3) | 0.68 |
| 5-HETrE | 18 (32) | 25 (25) | 26 (12) | 21 (19) | 32 (7) | 34 (19) | 30 (21) | 0.99 |
| 5-KETE | 108 (15) | 115 (38) | 124 (28) | 100 (45) | 110 (31) | 131 (38) | 142 (36) | 0.06 |
| 5-HETE | 336 (36) | 320 (39) | 300 (82) | 338 (48) | 211 (51) | 582 (292) | 454 (92) | 0.32 |
| 8,9-EET | 732 (835) | 695 (688) | 663 (927) | 606 (1313) | . | . | . | NA |
| 8,9-DiHETrE | 108 (26) | 85 (34) | 106 (27) | 101 (21) | 110 (31) | 107 (34) | 87 (15) | 0.32 |
| 8-HEPE | 18 (32) | 15.5 (2) | 9 (1.5) | 14 (13) | 22 (6) | 13.5 (6) | 17.5 (19.5) | 0.9 |
| 8-HETrE | 44 (17) | 52 (11) | 37 (16) | 51 (4) | 38 (11) | 52 (23) | 63 (7) | 0.17 |
| 8-HETE | 68 19) | 68 (13) | 60 (8) | 77 (25) | 119 (32) | 127 (42) | 114 (44) | 0.03 |
| 9,10-EpOME | 852 (480) | 542 (375) | 577 (438) | 495 (418) | 566 (352) | 727 (463) | 761 (380) | 0.68 |
| 9,10-DiHOME | 1719 (1681) | 1578 (1584) | 1720 (1275) | 1532 (1582) | 1734 (1784) | 1997 (1228) | 1884 (2036) | 0.99 |
| 9-HODE | 4073 (1849) | 4912 (1390) | 3553 (1839) | 3814 (1856) | 3937 (1474) | 3858 (1546) | 4005 (1986) | 0.99 |
| 9-HOTrE | 25 (10) | 28 (3) | 24 (5) | 26 (3) | 24 (3) | 27 (3) | 19 (8) | 0.68 |
| 9-KOTrE | 84 (25) | 66 (13) | 62 (20) | 63 (12) | 71 (7) | 66.5 (14.5) | 44 (25) | 0.09 |
| 9-OxoODE | 600 (157) | 526 (119) | 485 (56) | 446 (148) | 620 (256) | 593 (228) | 564 (326) | 0.99 |
| EKODE | 360 (220) | 313 (177) | 275 (20) | 367 (84) | 371 (146) | 417 (323) | 260 (108) | 0.32 |
| Oxylipin | Biosynthesis Pathway | Calibration Equation Average 1 | Sample Processing p-Values (Table 2) 2 | Sample Storage p-Values (Table 3) 3 | Intraday CVs 4 | |
|---|---|---|---|---|---|---|
| Average Slope | CV | T0, 10, 30, 60, 120 min | Fresh vs. Frozen | Fresh EDTA Plasma | ||
| ALA | ||||||
| 13-HOTrE | 15-LOX | 4.51 | 11.3% | 0.99 | 0.76 | 17.1% |
| 9-HOTrE | NA | 22.40 | 7.4% | 0.92 | 0.68 | 28.2% |
| 9-KOTrE | NA | 0.60 | 13.9% | 0.99 | 0.09 | 14.1% |
| EPA | ||||||
| 11-HEPE 5 | ROS | 1.08 | 7.0% | NA | 0.76 | 13.7% |
| 12-HEPE 6 | 12-LOX | 0.08 | 28.2% | NA | NA | NA |
| 14(15)-EpETE 6 | CYP | 0.49 | 11.4% | NA | NA | NA |
| 14,15-DiHETE 6 | EH | 0.05 | 5.3% | NA | NA | NA |
| 15-HEPE | 15-LOX | 0.78 | 5.5% | NA | 0.68 | 17.2% |
| 17(18)-EpETE | CYP | 0.93 | 8.2% | NA | 0.32 | 34.1% |
| 17,18-DiHETE | EH | 3.23 | 4.1% | NA | 0.48 | 14.0% |
| 18-HEPE | 15-LOX | 0.61 | 19.5% | NA | 0.03 | 25.9% |
| 5-HEPE | 5-LOX | 1.01 | 8.9% | NA | 0.68 | 36.4% |
| 8-HEPE | ROS | 0.78 | 7.5% | NA | 0.90 | 29.3% |
| 9-HEPE6 | ROS | 0.37 | 7.5% | NA | NA | NA |
| LXA5 6 | 5-LOX | 2.44 | 2.3% | NA | NA | NA |
| RVD5 6 | 15-LOX | 0.79 | 1.7% | NA | NA | NA |
| RVE1 5 | CYP/5LOX | 1.81 | 5.2% | NA | NA | NA |
| DHA | ||||||
| 10,17-DiHDoHE 6 | 15-LOX | 1.62 | 1.7% | NA | NA | NA |
| 11-HDoHE 5 | ROS | 0.90 | 7.9% | 0.07 | NA | NA |
| 14-HDHA | 12-LOX | 1.50 | 10.3% | 0.67 | 0.04 | 60.7% |
| 17-HDHA | 15-LOX | 0.43 | 15.0% | 0.75 | 0.06 | 13.3% |
| 17R-RVD1 6 | 15-LOX | 0.60 | 4.6% | NA | NA | NA |
| RVD1 6 | 15-LOX | 1.22 | 6.4% | NA | NA | NA |
| RVD2 6 | 15-LOX | 0.40 | 6.8% | NA | NA | NA |
| RVD3 5 | 15-LOX | 3.07 | 2.3% | NA | NA | NA |
| 19,20-EpDPE 5 | CYP | 0.71 | 10.1% | NA | NA | NA |
| 19,20-DiHDPA | EH | 7.00 | 17.0% | NA | 0.40 | 21.8% |
| 7R Maresin-1 5 | 12-LOX | 1.00 | 4.4% | NA | NA | NA |
| 7S Maresin-1 6 | 12-LOX | 0.26 | 6.8% | NA | NA | NA |
| 8-HDoHE | ROS | 0.80 | 8.3% | NA | 0.03 | 29.1% |
| LA | ||||||
| 12(13)-EpOME | CYP | 0.34 | 20.0% | 0.91 | 0.83 | 15.7% |
| 12,13-DiHOME | EH | 2.20 | 7.7% | 1.00 | 0.83 | 14.5% |
| 13-HODE | 15-LOX | 0.14 | 7.7% | 1.00 | 0.99 | 21.4% |
| 13-OxoODE | 15-LOX | 0.56 | 4.3% | 0.82 | 0.99 | 14.0% |
| 9(10)-EpOME | CYP | 2.09 | 14.0% | 0.93 | 0.68 | 18.9% |
| 9,10-DiHOME | EH | 3.35 | 4.4% | 1.00 | 0.99 | 14.9% |
| 9-HODE | NA | 1.66 | 10.3% | 1.00 | 0.99 | 21.1% |
| 9-OxoODE | NA | 0.50 | 2.6% | 0.70 | 0.99 | 20.7% |
| EKODE | NA | 2.32 | 6.7% | 0.77 | 0.32 | 21.3% |
| DGLA | ||||||
| 15-HETrE | 15-LOX | 3.24 | 8.6% | 0.99 | 0.09 | 16.3% |
| 5-HETrE | 5-LOX | 3.03 | 6.8% | 0.99 | 0.99 | 20.7% |
| 8-HETrE | ROS | 1.25 | 8.7% | 0.9 | 0.17 | 16.7% |
| ARA | ||||||
| 11(12)-EET | CYP | 0.90 | 8.0% | 0.92 | 0.03 | 16.9% |
| 11,12-DiHETrE | EH | 1.97 | 3.8% | 0.74 | 0.42 | 8.2% |
| 11-HETE | ROS | 7.58 | 8.3% | 0.99 | 0.09 | 11.2% |
| 11B-PGF2a 5 | COX | 21.6 | 5.5% | NA | NA | NA |
| 11-dehydro-TBX3 6 | COX | 7.3 | 6.4% | NA | NA | NA |
| 12-HHTrE | COX | 0.31 | 11.1% | NA | 0.17 | 29.2% |
| 12-HETE | 12-LOX | 0.64 | 10.7% | 0.32 | 0.03 | 17.7% |
| 12-KETE 6 | 12-LOX | 0.07 | 19.4% | NA | NA | NA |
| 15-KETE 6 | 15-LOX | 1.5 | 12.2% | NA | NA | NA |
| 14(15)-EET | CYP | 5.93 | 13.8% | 0.65 | 0.68 | 15.2% |
| 14,15-DiHETrE | EH | 3.01 | 3.6% | 0.86 | 0.37 | 9.9% |
| 15R-LXA4 6 | 5-LOX | 1.19 | 5.5% | 0.37 | NA | 14.5% |
| 15-HETE | 15-LOX | 1.05 | 8.6% | 0.98 | 0.99 | 11.2% |
| 15-deoxy-Δ12,14-PGJ2 5 | COX | 3.86 | 5.4% | NA | NA | NA |
| 20-HETE | CYP | 0.57 | 6.0% | 0.85 | 0.99 | 21.6% |
| 20-hydroxy-LTB4 6 | 5-LOX | 3.94 | 6.5% | NA | NA | NA |
| 5,6-EpETrE 5 | CYP | 0.03 | 12.7% | NA | NA | NA |
| 5,6-DiHETrE | EH | 1.15 | 8.0% | 0.9 | 0.24 | 20.8% |
| 5-KETE | 5-LOX | 0.62 | 7.6% | 0.54 | 0.06 | 24.9% |
| 5-HETE | 5-LOX | 1.26 | 5.0% | 0.88 | 0.32 | 14.7% |
| 5,15-DiHETE 5 | 5-LOX | 0.8 | 2.4% | NA | NA | NA |
| 6-α-PG 6 | COX | 11.4 | 4.5% | NA | NA | NA |
| 6-keto-PGF1α 6 | COX | 3.81 | 4.2% | NA | NA | NA |
| 6-trans-LTB4 6 | 5-LOX | 2.05 | 61.1% | NA | NA | NA |
| 8(9)-EET | CYP | 0.69 | 30.0% | NA | NA | 19.2% |
| 8,9-DiHETrE | EH | 0.93 | 5.0% | 0.73 | 0.32 | 16.3% |
| 8,15-DiHETE 5 | 5-LOX | 0.63 | 5.0% | NA | NA | NA |
| 8-iso-15-PGF2α 5 | ROS | 8.67 | 25.3% | NA | NA | NA |
| 8-iso-PGF2α 5 | ROS | 14.82 | 5.9% | NA | NA | NA |
| 8-HETE | ROS | 0.97 | 11.4% | 0.85 | 0.03 | 20.2% |
| 9-HETE | ROS | 1.03 | 6.7% | 0.92 | NA | NA |
| TXA2 5 | COX | 0.59 | 14.1% | NA | NA | NA |
| TXB2 | COX | 12.62 | 4.7% | NA | 0.04 | 42.2% |
| dinor-11B-PGF2α 6 | COX | 2.68 | 4.1% | NA | NA | NA |
| dinor-6-keto- PGF2α 6 | COX | 1.03 | 6.3% | NA | NA | NA |
| dinor-8-iso- PGF2α 6 | COX | 5.36 | 4.9% | NA | NA | NA |
| LXA4 6 | 5-LOX | 0.73 | 14.8% | NA | NA | NA |
| LXB4 6 | 5-LOX | 0.31 | 5.4% | NA | NA | NA |
| LTB4 6 | 5-LOX | 2.83 | 3.4% | NA | NA | NA |
| LTD4 6 | 5-LOX/GST | 2.59 | 36.6% | NA | NA | NA |
| LTE4 6 | 5-LOX/GST | 2.45 | 43.4% | NA | NA | NA |
| PGB2 6 | COX | 0.98 | 4.1% | NA | NA | NA |
| PGE2 6 | COX | 1.28 | 3.4% | NA | NA | NA |
| PGF2α 5 | COX | 40.23 | 5.8% | NA | NA | NA |
| PGJ2 5 | COX | 2.74 | 5.3% | NA | NA | NA |
| PGD2 6 | COX | 4.73 | 4.3% | NA | NA | NA |
| tetranor-PGEM 5 | COX | 4.20 | 11.2% | NA | NA | NA |
| tetranor-PGFM 6 | COX | 4.61 | 11.9% | NA | NA | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polinski, K.J.; Armstrong, M.; Manke, J.; Seifert, J.; Crume, T.; Yang, F.; Clare-Salzler, M.; Holers, V.M.; Reisdorph, N.; Norris, J.M. Collection and Storage of Human Plasma for Measurement of Oxylipins. Metabolites 2021, 11, 137. https://doi.org/10.3390/metabo11030137
Polinski KJ, Armstrong M, Manke J, Seifert J, Crume T, Yang F, Clare-Salzler M, Holers VM, Reisdorph N, Norris JM. Collection and Storage of Human Plasma for Measurement of Oxylipins. Metabolites. 2021; 11(3):137. https://doi.org/10.3390/metabo11030137
Chicago/Turabian StylePolinski, Kristen J., Michael Armstrong, Jonathan Manke, Jennifer Seifert, Tessa Crume, Fan Yang, Michael Clare-Salzler, V. Michael Holers, Nichole Reisdorph, and Jill M. Norris. 2021. "Collection and Storage of Human Plasma for Measurement of Oxylipins" Metabolites 11, no. 3: 137. https://doi.org/10.3390/metabo11030137
APA StylePolinski, K. J., Armstrong, M., Manke, J., Seifert, J., Crume, T., Yang, F., Clare-Salzler, M., Holers, V. M., Reisdorph, N., & Norris, J. M. (2021). Collection and Storage of Human Plasma for Measurement of Oxylipins. Metabolites, 11(3), 137. https://doi.org/10.3390/metabo11030137