
Targeted Metabolomic Assessment of the Sub-Lethal Toxicity of Halogenated Acetic Acids (HAAs) to Daphnia magna


Abstract
1. Introduction
2. Results
2.1. Multivariate Analysis of Metabolic Changes with HAA Exposure
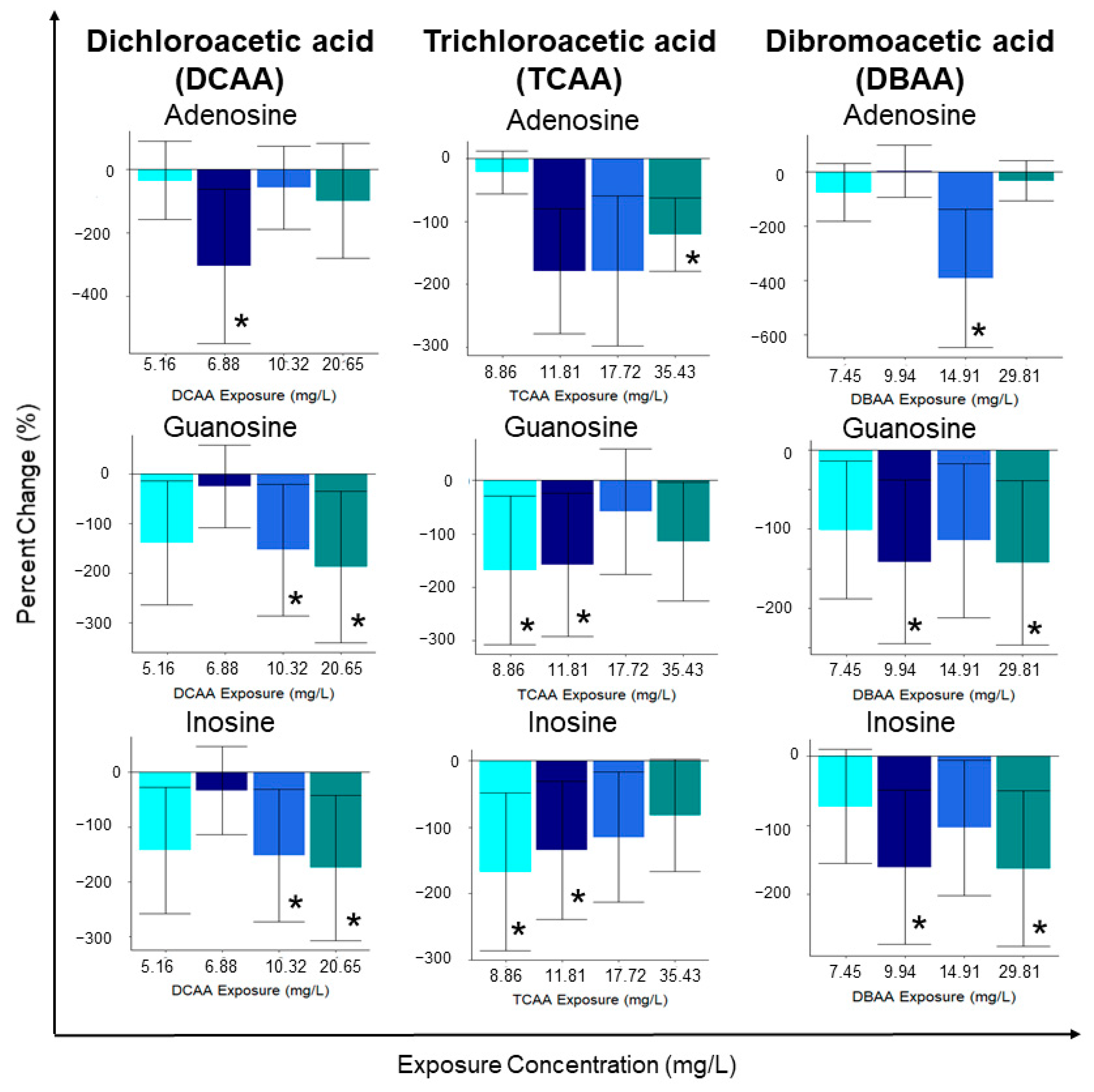
2.2. Changes in Specific Metabolites with HAA Exposure
2.2.1. Dichloroacetic Acid (DCAA) Metabolite Changes
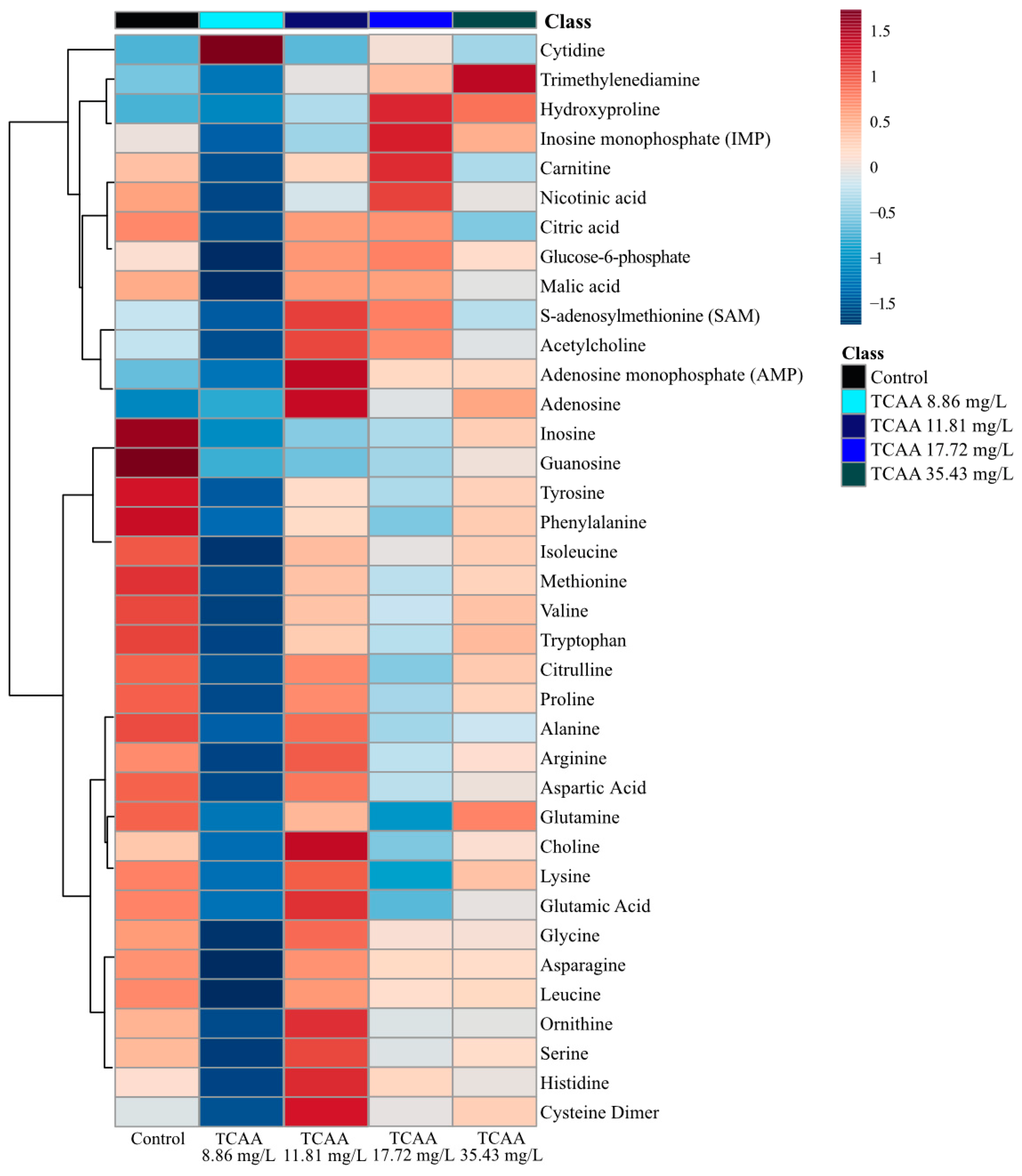
2.2.2. Trichloroacetic Acid (TCAA) Metabolite Changes
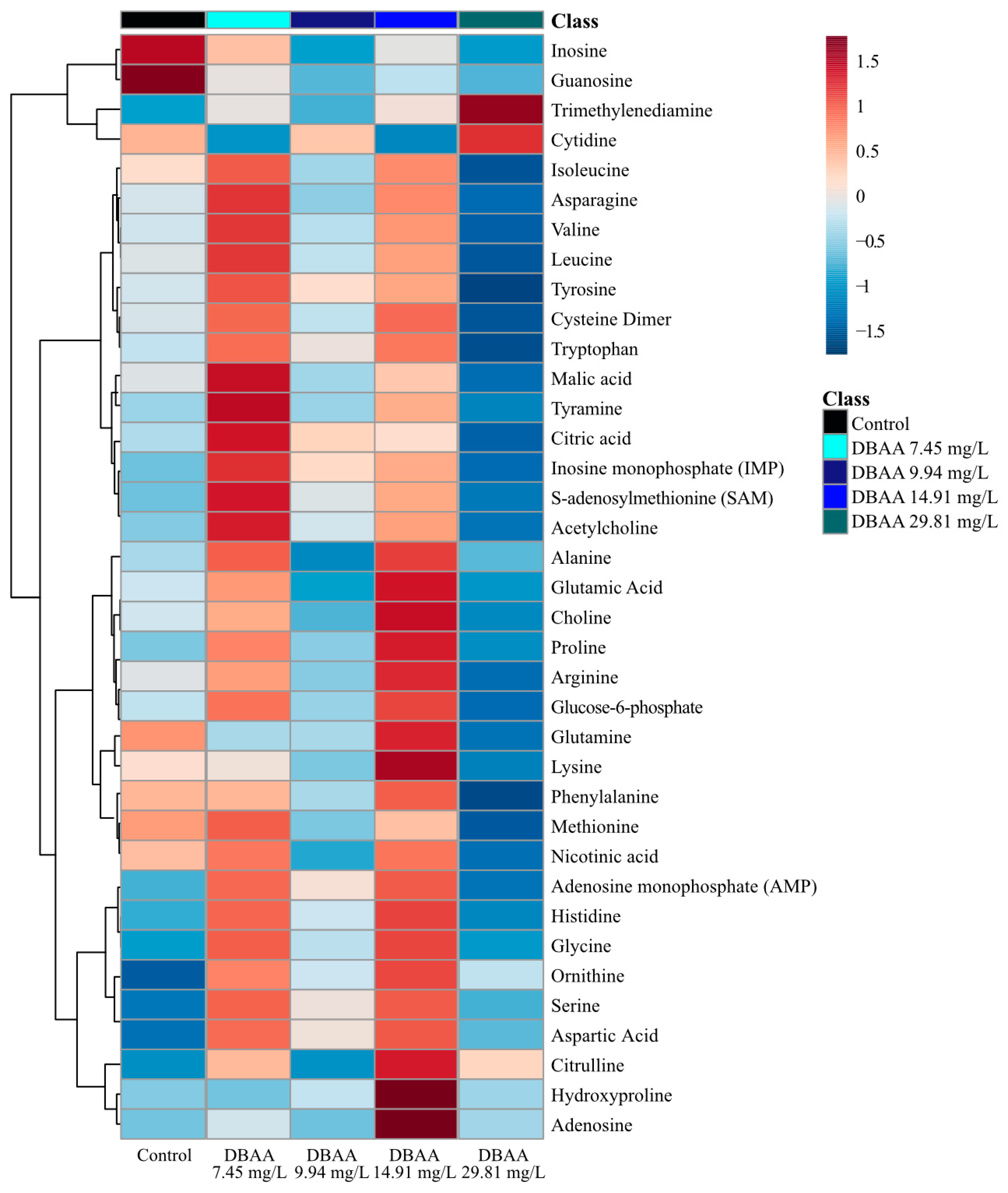
2.2.3. Dibromoacetic Acid (DBAA) Metabolite Changes
2.3. Biochemical Pathway Analysis
3. Discussion
4. Materials and Methods
4.1. Culturing of Daphnia Magna
4.2. Lethal Concentration (LC50) Determination of Select Halogenated Acetic Acids (HAAs)
4.3. Sub-Lethal Exposure of DCAA, TCAA and DBAA to D. magna
4.4. Metabolite Extraction and LC-MS/MS Analysis
4.5. Data Processing and Pathway Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krasner, S.W.; Weinberg, H.S.; Richardson, S.D.; Pastor, S.J.; Chinn, R.; Sclimenti, M.J.; Onstad, G.D.; Thruston, A.D. Occurrence of a New Generation of Disinfection Byproducts†. Environ. Sci. Technol. 2006, 40, 7175–7185. [Google Scholar] [CrossRef]
- Diehl, A.C.; Speitel, G.E.; Symons, J.M.; Krasner, S.W.; Hwang, C.J.; Barrett, S.E. DBP formation during chloramination. J. Am. Water Work. Assoc. 2000, 92, 76–90. [Google Scholar] [CrossRef]
- Selçuk, H.; Meric, C.B.O.A.S.; Nikolaou, A.D.; Bekbolet, M. A comparative study on the control of disinfection by-products (DBPs) and toxicity in drinking water. Desalination Water Treat. 2011, 26, 165–171. [Google Scholar] [CrossRef]
- Park, K.-Y.; Choi, S.-Y.; Lee, S.-H.; Kweon, J.-H.; Song, J.-H. Comparison of formation of disinfection by-products by chlorination and ozonation of wastewater effluents and their toxicity to Daphnia magna. Environ. Pollut. 2016, 215, 314–321. [Google Scholar] [CrossRef]
- Richardson, S.D.; Plewa, M.J.; Wagner, E.D.; Schoeny, R.; DeMarini, D.M. Occurrence, genotoxicity, and carcinogenicity of regulated and emerging disinfection by-products in drinking water: A review and roadmap for research. Mutat. Res. Mutat. Res. 2007, 636, 178–242. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Cheung, L.-M.; Yang, X.; Shang, C. THM, HAA and CNCl formation from UV irradiation and chlor(am)ination of selected organic waters. Water Res. 2006, 40, 2033–2043. [Google Scholar] [CrossRef]
- Liu, W.; Zhao, Y.; Chow, C.W.; Wang, D. Formation of disinfection byproducts in typical Chinese drinking water. J. Environ. Sci. 2011, 23, 897–903. [Google Scholar] [CrossRef]
- Mattei, D.; Cataudella, S.; Mancini, L.; Tancioni, L.; Migliore, L. Tiber River Quality in the Stretch of a Sewage Treatment Plant: Effects of River Water or Disinfectants to Daphnia and Structure of Benthic Macroinvertebrates Community. Water Air Soil Pollut. 2006, 177, 441–455. [Google Scholar] [CrossRef]
- Delinsky, A.D.; Delinsky, D.C.; Muralidhara, S.; Fisher, J.W.; Bruckner, J.V.; Bartlett, M. Analysis of dichloroacetic acid in rat blood and tissues by hydrophilic interaction liquid chromatography with tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2005, 19, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Uden, P.C.; Miller, J.W. Chlorinated acids and chloral in drinking water. J. Am. Water Work. Assoc. 1983, 75, 524–527. [Google Scholar] [CrossRef]
- DeAngelo, A.B. The carcinogenicity of dichloroacetic acid in the male fischer 344 rat. Toxicology 1996, 114, 207–221. [Google Scholar] [CrossRef]
- Karnes, J.B.; Usatine, R.P. Management of External Genital Warts. Am. Fam. Physician 2014, 90, 312–318. [Google Scholar]
- Beutner, K.R.; Spruance, S.L.; Hougham, A.J.; Fox, T.L.; Owens, M.L.; Douglas, J.M. Treatment of genital warts with an immune-response modifier (imiquimod). J. Am. Acad. Dermatol. 1998, 38, 230–239. [Google Scholar] [CrossRef]
- Bowden, D.J.; Clegg, S.L.; Brimblecombe, P. The Henry’s Law Constant of Trichloroacetic Acid. Water Air Soil Pollut. 1998, 101, 197–215. [Google Scholar] [CrossRef]
- Bowden, D.J.; Clegg, S.L.; Brimblecombe, P. The Henry’s Law Constants of the Haloacetic Acids. J. Atmos. Chem. 1998, 29, 85–107. [Google Scholar] [CrossRef]
- Hanson, M.L.; Sibley, P.K.; Mabury, S.A.; Solomon, K.R.; Muir, D.C. Trichloroacetic acid (TCA) and trifluoroacetic acid (TFA) mixture toxicity to the macrophytes Myriophyllum spicatum and Myriophyllum sibiricum in aquatic microcosms. Sci. Total Environ. 2002, 285, 247–259. [Google Scholar] [CrossRef]
- Ellis, D.A.; Hanson, M.L.; Sibley, P.K.; Shahid, T.; Fineberg, N.A.; Solomon, K.R.; Muir, D.C.G.; Mabury, S.A. The fate and persistence of trifluoroacetic and chloroacetic acids in pond waters. Chemosphere 2001, 42, 309–318. [Google Scholar] [CrossRef]
- Fetzner, S. Bacterial dehalogenation. Appl. Microbiol. Biotechnol. 1998, 50, 633–657. [Google Scholar] [CrossRef]
- Bayless, W.; Andrews, R.C. Biodegradation of six haloacetic acids in drinking water. J. Water Health 2007, 6, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Scott, B.F.; MacTavish, D.; Spencer, C.; Strachan, W.M.J.; Muir, D.C.G. Haloacetic Acids in Canadian Lake Waters and Precipitation. Environ. Sci. Technol. 2000, 34, 4266–4272. [Google Scholar] [CrossRef]
- Melo, A.; Ferreira, C.; Ferreira, I.M.; Mansilha, C.R. Acute and chronic toxicity assessment of haloacetic acids using Daphnia magna. J. Toxicol. Environ. Health Part A 2019, 82, 977–989. [Google Scholar] [CrossRef]
- Hanson, M.L.; Solomon, K.R. Haloacetic acids in the aquatic environment. Part I: Macrophyte toxicity. Environ. Pollut. 2004, 130, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Berends, A.; Boutonnet, J.C.; De Rooij, C.G.; Thompson, R.S. Toxicity of trifluoroacetate to aquatic organisms. Environ. Toxicol. Chem. 1999, 18, 1053–1059. [Google Scholar] [CrossRef]
- Fitzsimmons, P.N.; Hoffman, A.D.; Lien, G.J.; Hammermeister, D.E.; Nichols, J.W. Kinetics and effects of dichloroacetic acid in rainbow trout. Aquat. Toxicol. 2009, 94, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.L.; Sibley, P.K.; A Mabury, S.; Muir, D.C.; Solomon, K.R. Field level evaluation and risk assessment of the toxicity of dichloroacetic acid to the aquatic macrophytes Lemna gibba, Myriophyllum spicatum, and Myriophyllum sibiricum. Ecotoxicol. Environ. Saf. 2003, 55, 46–63. [Google Scholar] [CrossRef]
- Toussaint, M.W.; Brennan, L.M.; Rosencrance, A.B.; Dennis, W.E.; Hoffmann, F.J.; Gardner, H.S., Jr. Acute Toxicity of Four Drinking Water Disinfection By-Products to Japanese Medaka Fish. Bull. Environ. Contam. Toxicol. 2001, 66, 255–262. [Google Scholar] [CrossRef]
- Williams, F.E.; Sickelbaugh, T.J.; Hassoun, E. Modulation by ellagic acid of DCA-induced developmental toxicity in the zebrafish (Danio rerio). J. Biochem. Mol. Toxicol. 2006, 20, 183–190. [Google Scholar] [CrossRef]
- Hassoun, E.; Kariya, C.; Williams, F.E. Dichloroacetate-induced developmental toxicity and production of reactive oxygen species in zebrafish embryos. J. Biochem. Mol. Toxicol. 2005, 19, 52–58. [Google Scholar] [CrossRef]
- Fisher, D.; Yonkos, L.; Ziegler, G.; Friedel, E.; Burton, D. Acute and chronic toxicity of selected disinfection byproducts to Daphnia magna, Cyprinodon variegatus, and Isochrysis galbana. Water Res. 2014, 55, 233–244. [Google Scholar] [CrossRef]
- Giller, S.; Le Curieux, F.; Erb, F.; Marzin, D. Comparative genotoxicity of halogenated acetic acids found in drinking water. Mutagenesis 1997, 12, 321–328. [Google Scholar] [CrossRef]
- Rizzo, L.; Belgiorno, V.; Gallo, M.; Meric, S. Removal of THM precursors from a high-alkaline surface water by enhanced coagulation and behaviour of THMFP toxicity on D. magna. Desalination 2005, 176, 177–188. [Google Scholar] [CrossRef]
- Andrews, J.; Nichols, H.; Schmid, J.; Mole, L.; Hunter, E.; Klinefelter, G. Developmental toxicity of mixtures: The water disinfection by-products dichloro-, dibromo- and bromochloro acetic acid in rat embryo culture. Reprod. Toxicol. 2004, 19, 111–116. [Google Scholar] [CrossRef]
- Smith, M.K.; Randall, J.L.; Read, E.J.; Stober, J.A. Developmental toxicity of dichloroacetate in the rat. Teratology 1992, 46, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.K.; Randall, J.L.; Read, E.J.; Stober, J.A. Teratogenic activity of trichloroacetic acid in the rat. Teratology 1989, 40, 445–451. [Google Scholar] [CrossRef]
- Epstein, D.L.; Nolen, G.A.; Randall, J.L.; Christ, S.A.; Read, E.J.; Stober, J.A.; Smith, M.K. Cardiopathic effects of dichloroacetate in the fetal long-evans rat. Teratology 1992, 46, 225–235. [Google Scholar] [CrossRef]
- Bull, R.J.; Orner, G.A.; Cheng, R.S.; Stillwell, L.; Stauber, A.J.; Sasser, L.B.; Lingohr, M.K.; Thrall, B.D. Contribution of Dichloroacetate and Trichloroacetate to Liver Tumor Induction in Mice by Trichloroethylene. Toxicol. Appl. Pharmacol. 2002, 182, 55–65. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Echigo, S.; Itoh, S.; Natsui, T.; Araki, T.; Ando, R. Contribution of brominated organic disinfection by-products to the mutagenicity of drinking water. Water Sci. Technol. 2004, 50, 321–328. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, X. Comparative Developmental Toxicity of New Aromatic Halogenated DBPs in a Chlorinated Saline Sewage Effluent to the Marine PolychaetePlatynereis dumerilii. Environ. Sci. Technol. 2013, 47, 10868–10876. [Google Scholar] [CrossRef]
- Lambert, V.; Hansen, S.; Schoumacher, M.; LeComte, J.; Leenders, J.; Hubert, P.; Herfs, M.; Blacher, S.; Carnet, O.; Yip, C.; et al. Pyruvate dehydrogenase kinase/lactate axis: A therapeutic target for neovascular age-related macular degeneration identified by metabolomics. J. Mol. Med. 2020, 98, 1737–1751. [Google Scholar] [CrossRef]
- Pettit, F.H.; Pelley, J.W.; Reed, L.J. Regulation of pyruvate dehydrogenase kinase and phosphatase by acetyl-CoA/CoA and NADH/NAD ratios. Biochem. Biophys. Res. Commun. 1975, 65, 575–582. [Google Scholar] [CrossRef]
- Law, J.M.; Lopez, L.; DeAngelo, A.B. Hepatotoxicity of the drinking water disinfection by-product, dichloroacetic acid, in the medaka small fish model. Toxicol. Lett. 1998, 94, 19–27. [Google Scholar] [CrossRef]
- Lee, B.-Y.; Choi, B.-S.; Kim, M.-S.; Park, J.C.; Jeong, C.-B.; Han, J.; Lee, J.-S. The genome of the freshwater water flea Daphnia magna: A potential use for freshwater molecular ecotoxicology. Aquat. Toxicol. 2019, 210, 69–84. [Google Scholar] [CrossRef]
- Smirnov, N.N. Physiology of the Cladocera; Elsevier: Amsterdam, The Netherlands, 2014; Volume 1. [Google Scholar]
- Sarma, S.S.S.; Nandini, S. Review of Recent Ecotoxicological Studies on Cladocerans. J. Environ. Sci. Health Part B 2006, 41, 1417–1430. [Google Scholar] [CrossRef]
- Han, G.H.; Hur, H.G.; Kim, S.D. Ecotoxicological Risk of Pharmaceuticals from Wastewater Treatment Plants in Korea: Occurrence and Toxicity to Daphnia Magna. Environ. Toxicol. Chem. 2006, 25, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.; Coors, A.; Stoks, R.; De Meester, L. Evolutionary ecotoxicology of pesticide resistance: A case study in Daphnia. Ecotoxicology 2011, 20, 543–551. [Google Scholar] [CrossRef]
- Woermann, M.; Sures, B. Ecotoxicological effects of micropollutant-loaded powdered activated carbon emitted from wastewater treatment plants on Daphnia magna. Sci. Total Environ. 2020, 746, 141104. [Google Scholar] [CrossRef]
- Taylor, N.S.; Weber, R.J.M.; Southam, A.D.; Payne, T.G.; Hrydziuszko, O.; Arvanitis, T.N.; Viant, M.R. A new approach to toxicity testing in Daphnia magna: Application of high throughput FT-ICR mass spectrometry metabolomics. Metabolomics 2009, 5, 44–58. [Google Scholar] [CrossRef]
- Taylor, N.S.; Weber, R.J.M.; White, T.A.; Viant, M.R. Discriminating between Different Acute Chemical Toxicities via Changes in the Daphnid Metabolome. Toxicol. Sci. 2010, 118, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Nagato, E.G.; Simpson, A.J.; Simpson, M.J. Metabolomics reveals energetic impairments in Daphnia magna exposed to diazinon, malathion and bisphenol-A. Aquat. Toxicol. 2016, 170, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Jeong, T.-Y.; Simpson, M.J. Daphnia magna metabolic profiling as a promising water quality parameter for the biological early warning system. Water Res. 2019, 166, 115033. [Google Scholar] [CrossRef]
- Jeong, T.-Y.; Simpson, M.J. Time-dependent biomolecular responses and bioaccumulation of perfluorooctane sulfonate (PFOS) in Daphnia magna. Comp. Biochem. Physiol. Part D Genom. Proteom. 2020, 35, 100701. [Google Scholar] [CrossRef] [PubMed]
- Jeong, T.-Y.; Simpson, M.J. Reproduction Stage Differentiates the Time-Course Regulation of Metabolites in Daphnia magna. Environ. Sci. Technol. 2019, 53, 12764–12773. [Google Scholar] [CrossRef] [PubMed]
- Jeong, T.-Y.; Simpson, M.J. Reproduction stage specific dysregulation of Daphnia magna metabolites as an early indicator of reproductive endocrine disruption. Water Res. 2020, 184, 116107. [Google Scholar] [CrossRef]
- Saccenti, E.; Hoefsloot, H.C.J.; Smilde, A.K.; Westerhuis, J.A.; Hendriks, M.M.W.B. Reflections on univariate and multi-variate analysis of metabolomics data. Metabolomics 2014, 10, 361–374. [Google Scholar] [CrossRef]
- Worley, B.; Powers, R. Multivariate Analysis in Metabolomics. Curr. Metab. 2012, 1, 92–107. [Google Scholar] [CrossRef][Green Version]
- Kapoore, R.V.; Vaidyanathan, S. Towards quantitative mass spectrometry-based metabolomics in microbial and mammalian systems. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2016, 374, 20150363. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more trans-parent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- Machado, A.A.D.S.; Zarfl, C.; Rehse, S.; Kloas, W. Low-Dose Effects: Nonmonotonic Responses for the Toxicity of a Bacillus thuringiensis Biocide to Daphnia magna. Environ. Sci. Technol. 2017, 51, 1679–1686. [Google Scholar] [CrossRef]
- Kim, J.; Lee, S.; Kim, C.-M.; Seo, J.; Park, Y.; Kwon, D.; Lee, S.-H.; Yoon, T.-H.; Choi, K. Non-monotonic concentration–response relationship of TiO2 nanoparticles in freshwater cladocerans under environmentally relevant UV-A light. Ecotoxicol. Environ. Saf. 2014, 101, 240–247. [Google Scholar] [CrossRef]
- Wagner, N.D.; Helm, P.A.; Simpson, A.J.; Simpson, M.J. Metabolomic responses to pre-chlorinated and final effluent wastewater with the addition of a sub-lethal persistent contaminant in Daphnia magna. Environ. Sci. Pollut. Res. 2019, 26, 9014–9026. [Google Scholar] [CrossRef]
- Kariuki, M.N.; Nagato, E.G.; Lankadurai, B.P.; Simpson, A.J.; Simpson, M.J. Analysis of Sub-Lethal Toxicity of Perfluorooctane Sulfonate (PFOS) to Daphnia magna Using 1H Nuclear Magnetic Resonance-Based Metabolomics. Metabolities 2017, 7, 15. [Google Scholar] [CrossRef]
- Kakuda, T.N. Pharmacology of nucleoside and nucleotide reverse transcriptase inhibitor-induced mitochondrial toxicity. Clin. Ther. 2000, 22, 685–708. [Google Scholar] [CrossRef]
- Oikawa, T.G.; Smith, M. Nucleotides in the Encysted Embryos of Daphnia magna. Biochemistry 1966, 5, 1517–1521. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, S.J.; Warner, A.H. The Presence of Guanosine 5’-Diphospho-5’qguanosine and Guanosine 5’-Triphospho-5’-adenosine in Brine Shrimp Embryos. J. Biol. Chem. 1978, 253, 4960–4965. [Google Scholar] [CrossRef]
- Pedley, A.M.; Benkovic, S.J. A New View into the Regulation of Purine Metabolism: The Purinosome. Trends Biochem. Sci. 2017, 42, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Orgel, L.E. RNA catalysis and the origins of life. J. Theor. Biol. 1986, 123, 127–149. [Google Scholar] [CrossRef]
- Müller-Hill, B. What is life? The paradigm of DNA and protein cooperation at high local concentrations. Mol. Microbiol. 2006, 60, 253–255. [Google Scholar] [CrossRef] [PubMed]
- Michalak, E.M.; Burr, M.L.; Bannister, A.J.; Dawson, M.A. The roles of DNA, RNA and histone methylation in ageing and cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 573–589. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef]
- Campos, B.; Garcia-Reyero, N.; Rivetti, C.; Escalon, L.; Habib, T.; Tauler, R.; Tsakovski, S.; Piña, B.; Barata, C. Identification of Metabolic Pathways in Daphnia magna Explaining Hormetic Effects of Selective Serotonin Reuptake Inhibitors and 4-Nonylphenol Using Transcriptomic and Phenotypic Responses. Environ. Sci. Technol. 2013, 47, 9434–9443. [Google Scholar] [CrossRef]
- Yan, L.-J.; Levine, R.L.; Sohal, R.S. Oxidative damage during aging targets mitochondrial aconitase. Proc. Natl. Acad. Sci. USA 1997, 94, 11168–11172. [Google Scholar] [CrossRef]
- Beinert, H.; Kennedy, A.M.C.; Stout§, C.D. Aconitase as Iron−Sulfur Protein, Enzyme, and Iron-Regulatory Protein. Chem. Rev. 1996, 96, 2335–2374. [Google Scholar] [CrossRef]
- Zurita, J.L.; Jos, A.; Cameán, A.M.; Salguero, M.; López-Artíguez, M.; Repetto, G. Ecotoxicological evaluation of sodium fluoroacetate on aquatic organisms and investigation of the effects on two fish cell lines. Chemosphere 2007, 67, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Goyer, A. Thiamine in plants: Aspects of its metabolism and functions. Phytochemistry 2010, 71, 1615–1624. [Google Scholar] [CrossRef]
- Allen, S.; Zilles, J.L.; Downs, D.M. Metabolic Flux in Both the Purine Mononucleotide and Histidine Biosynthetic Pathways Can Influence Synthesis of the Hydroxymethyl Pyrimidine Moiety of Thiamine in Salmonella enterica. J. Bacteriol. 2002, 184, 6130–6137. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rapala-Kozik, M.; Kowalska, E.; Ostrowska, K. Modulation of thiamine metabolism in Zea mays seedlings under conditions of abiotic stress. J. Exp. Bot. 2008, 59, 4133–4143. [Google Scholar] [CrossRef]
- Mayr, J.A.; Freisinger, P.; Schlachter, K.; Rolinski, B.; Zimmermann, F.A.; Scheffner, T.; Haack, T.B.; Koch, J.; Ahting, U.; Prokisch, H.; et al. Thiamine Pyrophosphokinase Deficiency in Encephalopathic Children with Defects in the Pyruvate Oxidation Pathway. Am. J. Hum. Genet. 2011, 89, 806–812. [Google Scholar] [CrossRef]
- Curtius, H.-C.; Völlmin, J.; Baerlocher, K. The use of deuterated phenylalanine for the elucidation of the phenylalanine-tyrosine metabolism. Clin. Chim. Acta 1972, 37, 277–285. [Google Scholar] [CrossRef]
- Gertsman, I.; Gangoiti, J.A.; Nyhan, W.L.; A Barshop, B. Perturbations of tyrosine metabolism promote the indolepyruvate pathway via tryptophan in host and microbiome. Mol. Genet. Metab. 2015, 114, 431–437. [Google Scholar] [CrossRef]
- Ontario Ministry of the Environment. Daphnia Magna Culturing; Ontario Ministry of the Environment: Toronto, ON, Canada, 2012; Volume SOP DM1.v8.
- Ontario Ministry of the Environment. Algae Culturing for Use as Food; Ontario Ministry of the Environment: Toronto, ON, Canada, 2012; Volume SOP AL1.v6.
- Environment Canada. Biological Test Method: Reference Method for Determining Acute Lethality of Effluents to Daphnia Magna; Environment Canada: Gatineau, QC, Canada, 2000; ISBN 0660616157.
- Robertson, J.L.; Russell, R.M.; Savin, N.E. POLO: A User’s Guide to Probit or LOgit Analysis, 1st ed.; U.S. Department of Agriculture: Washington, DC, USA; Pacific Southwest Forest and Range Experiment Station: Berkeley, CA, USA, 1980.
- Xie, Y. Analyzing Haloacetic Acids Using Gas Chromatography/Mass Spectrometry. Water Res. 2001, 35, 1599–1602. [Google Scholar] [CrossRef]
- Meng, L.; Wu, S.; Ma, F.; Jia, A.; Hu, J. Trace determination of nine haloacetic acids in drinking water by liquid chromatography–electrospray tandem mass spectrometry. J. Chromatogr. A 2010, 1217, 4873–4876. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Wang, D.; Wei, Z.; Wang, Z. Optimized chromatographic conditions for separation of halogenated acetic acids by ultra-performance liquid chromatography–electrospray ionization-mass spectrometry. J. Chromatogr. A 2013, 1277, 26–34. [Google Scholar] [CrossRef]
- Wu, Y.; Li, L. Sample normalization methods in quantitative metabolomics. J. Chromatogr. A 2016, 1430, 80–95. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Li, S.; Xia, J. MetaboAnalystR 3.0: Toward an Optimized Workflow for Global Metabolomics. Metabolities 2020, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Castillo, S.; Gopalacharyulu, P.; Yetukuri, L.; Orešič, M. Algorithms and tools for the preprocessing of LC–MS metabolomics data. Chemom. Intell. Lab. Syst. 2011, 108, 23–32. [Google Scholar] [CrossRef]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Sun, X.; Weckwerth, W. COVAIN: A toolbox for uni- and multivariate statistics, time-series and correlation network analysis and inverse estimation of the differential Jacobian from metabolomics covariance data. Metabolomics 2012, 8, 81–93. [Google Scholar] [CrossRef]
- Chong, J.; Xia, J. MetaboAnalystR: An R package for flexible and reproducible analysis of metabolomics data. Bioinformatics 2018, 34, 4313–4314. [Google Scholar] [CrossRef]
- Rusilowicz, M.; Dickinson, M.; Charlton, A.; O’Keefe, S.; Wilson, J. A batch correction method for liquid chromatography–mass spectrometry data that does not depend on quality control samples. Metabolomics 2016, 12, 1–11. [Google Scholar] [CrossRef]
- Flores-Valverde, A.M.; Horwood, J.; Hill, E.M. Disruption of the Steroid Metabolome in Fish Caused by Exposure to the Environmental Estrogen 17α-Ethinylestradiol. Environ. Sci. Technol. 2010, 44, 3552–3558. [Google Scholar] [CrossRef]
- Southam, A.D.; Lange, A.; Hines, A.; Hill, E.M.; Katsu, Y.; Iguchi, T.; Tyler, C.R.; Viant, M.R. Metabolomics Reveals Target and Off-Target Toxicities of a Model Organophosphate Pesticide to Roach (Rutilus Rutilus): Implications for Biomonitoring. Environ. Sci. Technol. 2011, 45, 3759–3767. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- R Core Team. R: A Language Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Kovacevic, V.; Simpson, A.J.; Simpson, M.J. 1H NMR-based metabolomics of Daphnia magna responses after sub-lethal exposure to triclosan, carbamazepine and ibuprofen. Comp. Biochem. Physiol. Part D Genom. Proteom. 2016, 19, 199–210. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Exposure Concentration (mg/L) | Impacted Pathway | p Value |
|---|---|---|---|
| DCAA | 20.65 | The citric acid (TCA) cycle | 0.0761 |
| Purine metabolism | 0.0828 | ||
| TCAA | 8.86 | Nicotinate and nicotinamide metabolism Ubiquinone and other terpenoid-quinone biosynthesis Phenylalanine, tyrosine and tryptophan biosynthesis Phenylalanine metabolism Valine, leucine and isoleucine degradation Valine, leucine and isoleucine biosynthesis Pantothenate and CoA biosynthesis Tyrosine metabolism The citric acid (TCA) cycle Histidine metabolism Glycerophospholipid metabolism Thiamine metabolism Aminoacyl-tRNA biosynthesis Butanoate metabolism Cysteine and methionine metabolism | ≤0.10 |
| DBAA | 7.45 | Thiamine metabolism | 0.0536 |
| Tyrosine metabolism | 0.0571 | ||
| The citric acid (TCA) cycle | 0.0840 | ||
| 14.91 | Purine metabolism | 0.0810 |
| Contaminant | Calculated 48 h LC50 (mg/L) | Literature LC50 (mg/L) |
|---|---|---|
| Dichloroacetic acid (DCAA) | 206.5 ± 25.1 | − |
| Trichloroacetic acid (TCAA) | 354.3 ± 22.7 | 249.5 1 |
| Dibromoacetic acid (DBAA) | 298.1 ± 24.7 | 254.4 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Labine, L.M.; Simpson, M.J. Targeted Metabolomic Assessment of the Sub-Lethal Toxicity of Halogenated Acetic Acids (HAAs) to Daphnia magna. Metabolites 2021, 11, 100. https://doi.org/10.3390/metabo11020100
Labine LM, Simpson MJ. Targeted Metabolomic Assessment of the Sub-Lethal Toxicity of Halogenated Acetic Acids (HAAs) to Daphnia magna. Metabolites. 2021; 11(2):100. https://doi.org/10.3390/metabo11020100
Chicago/Turabian StyleLabine, Lisa M., and Myrna J. Simpson. 2021. "Targeted Metabolomic Assessment of the Sub-Lethal Toxicity of Halogenated Acetic Acids (HAAs) to Daphnia magna" Metabolites 11, no. 2: 100. https://doi.org/10.3390/metabo11020100
APA StyleLabine, L. M., & Simpson, M. J. (2021). Targeted Metabolomic Assessment of the Sub-Lethal Toxicity of Halogenated Acetic Acids (HAAs) to Daphnia magna. Metabolites, 11(2), 100. https://doi.org/10.3390/metabo11020100