
Intracellularly Released Cholesterol from Polymer-Based Delivery Systems Alters Cellular Responses to Pneumolysin and Promotes Cell Survival

 ,
,  , ,
, ,  , ,
, , 


{kind=link}
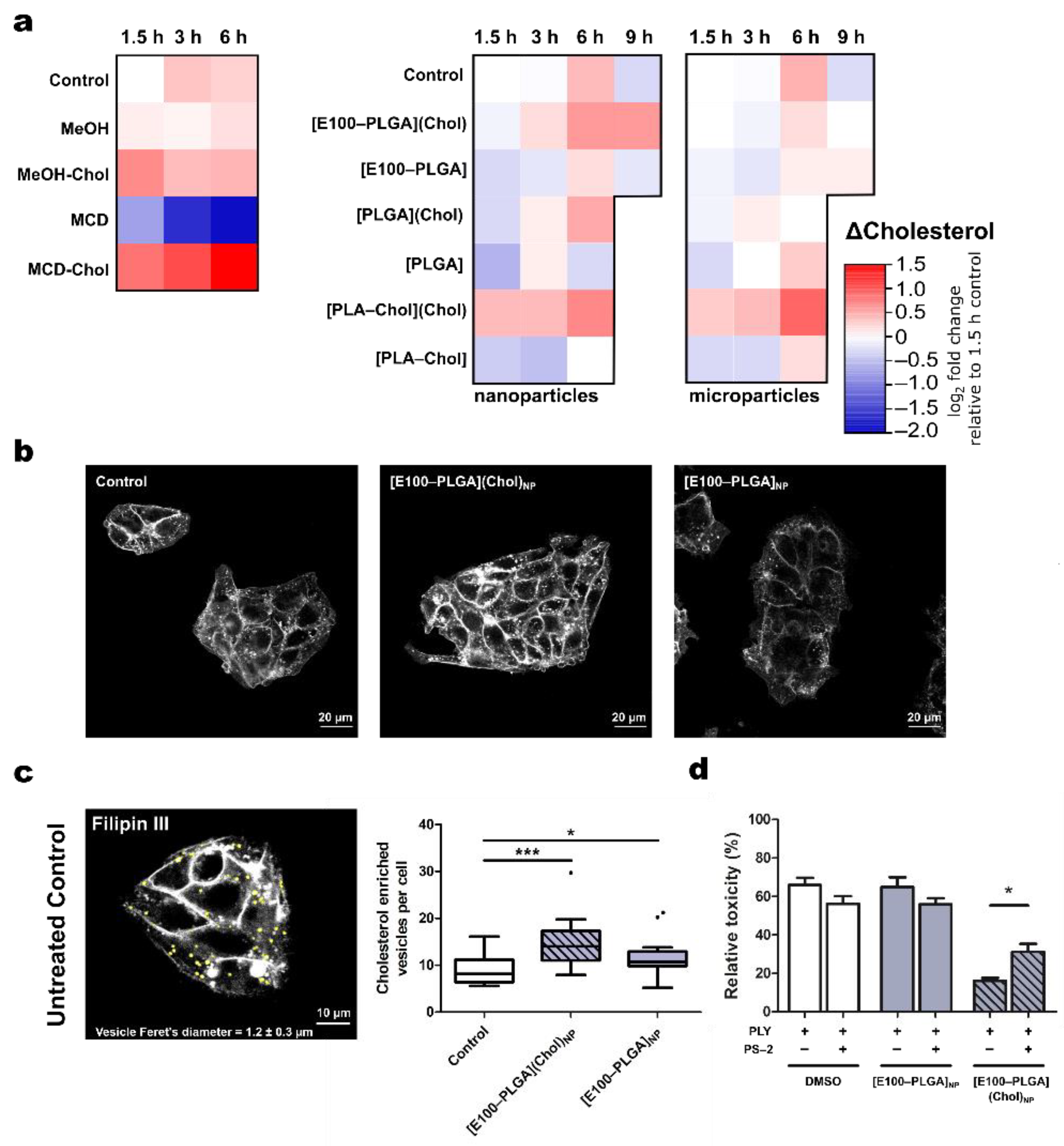{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
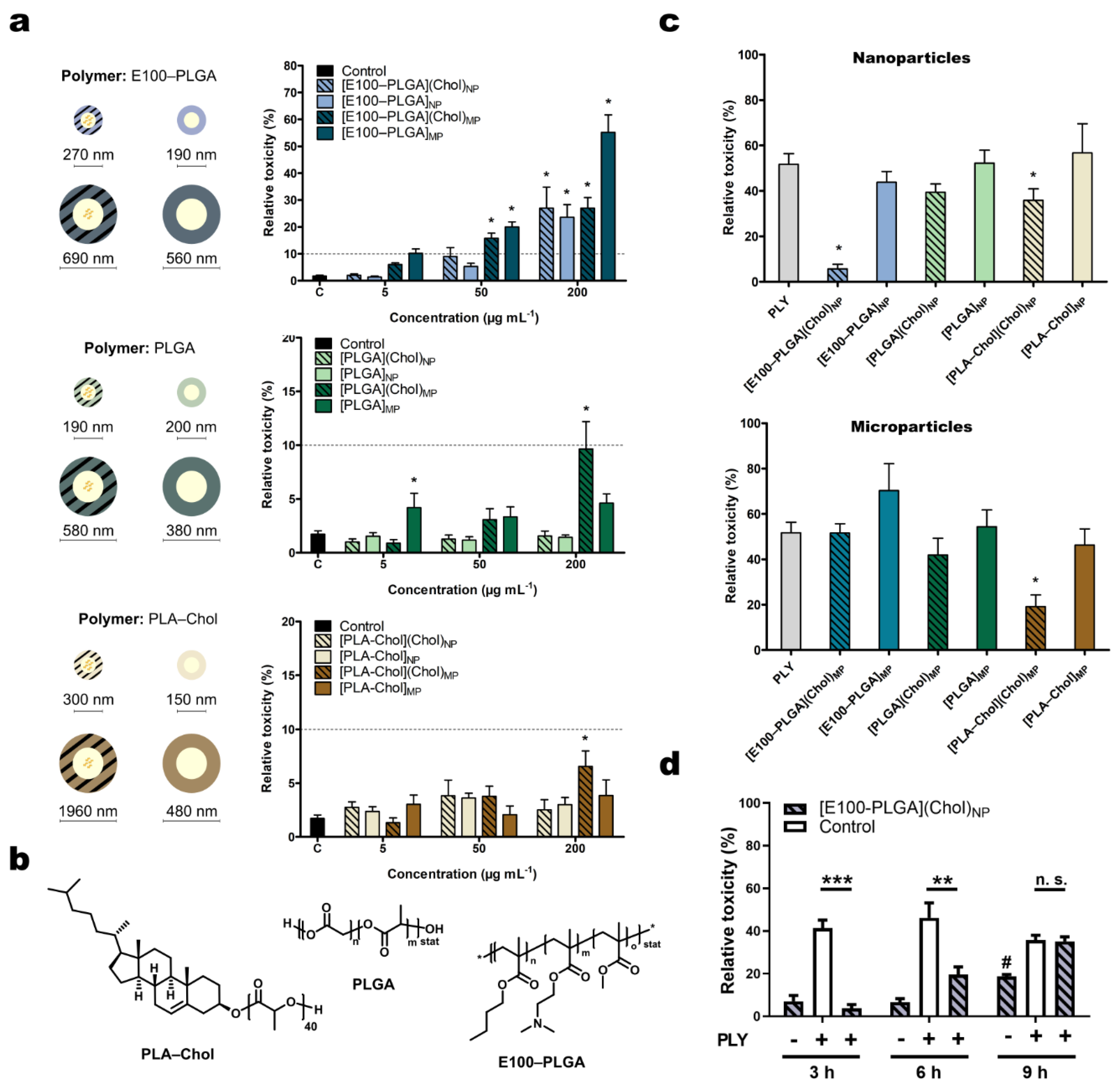
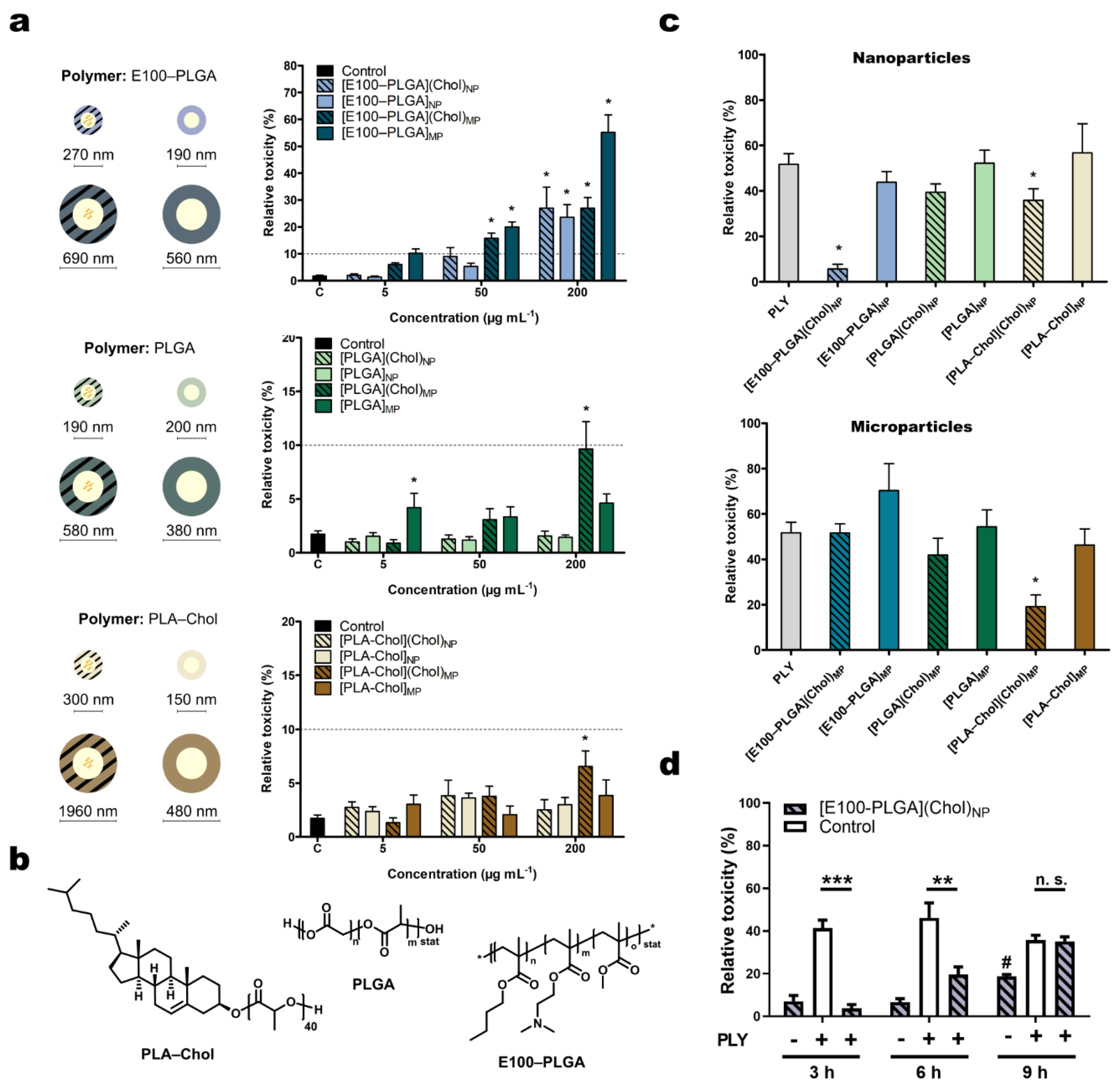
2.1. Characterization of Polymer Particles
2.2. Effects of the Cholesterol Loaded Nano- and Microcarrier on Pneumolysin Toxicity
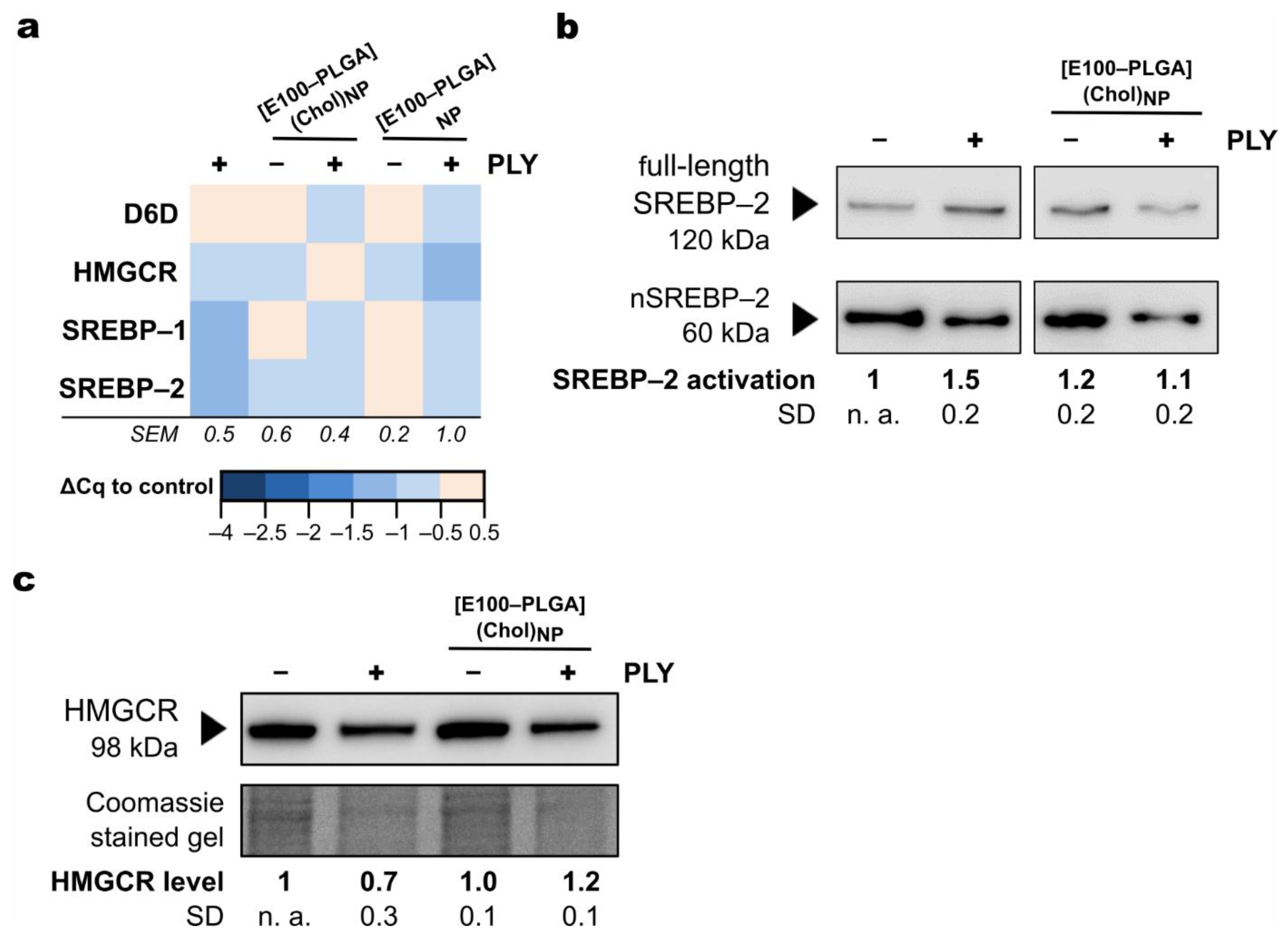
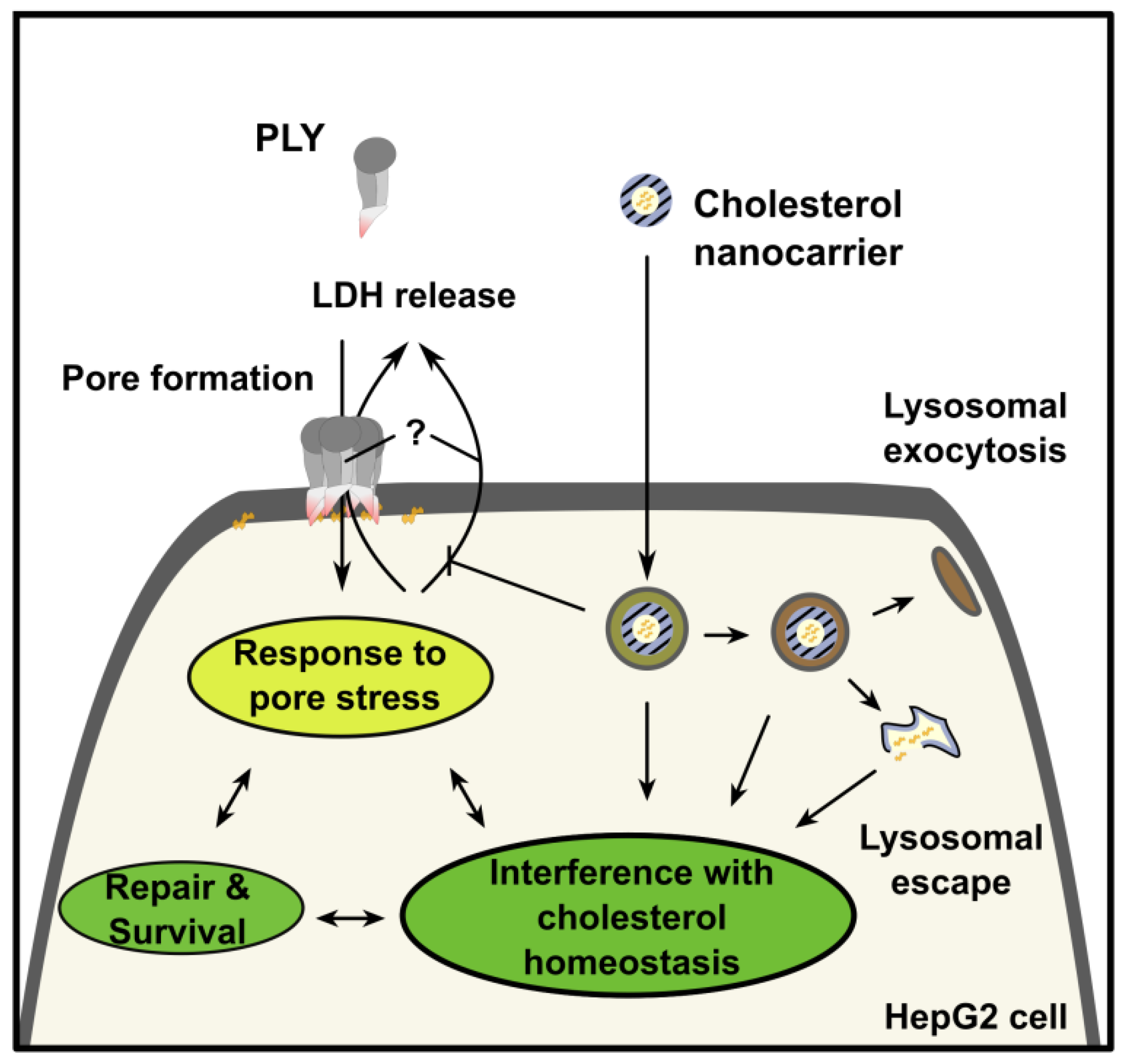
2.3. Mechanism of Protection from Pneumolysin by Nanoformulated Cholesterol
3. Materials and Methods
3.1. Synthesis and Characterization of Cholesterol Terminated Polylactide (PLA–Chol)
3.2. Synthesis and Characterization of Cholesterol-Containing Particles
3.3. Cell Culture
3.4. Stimulations
3.5. Toxicity
3.6. Cholesterol Quantification
3.7. Gene Expression Analysis
3.8. Cholesterol Staining
3.9. Western Blotting
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Howe, V.; Sharpe, L.J.; Alexopoulos, S.J.; Kunze, S.V.; Chua, N.K.; Li, D.; Brown, A.J. Cholesterol homeostasis: How do cells sense sterol excess? Chem. Phys. Lipids 2016, 199, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Ikonen, E. Cellular cholesterol trafficking and compartmentalization. Nat. Rev. Mol. Cell Biol. 2008, 9, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Hupp, S.; Ribes, S.; Seele, J.; Bischoff, C.; Förtsch, C.; Maier, E.; Benz, R.; Mitchell, T.J.; Nau, R.; Iliev, A.I. Magnesium therapy improves outcome in Streptococcus pneumoniae meningitis by altering pneumolysin pore formation. Br. J. Pharmacol. 2017, 174, 4295–4307. [Google Scholar] [CrossRef] [PubMed]
- van Pee, K.; Mulvihill, E.; Müller, D.J.; Yildiz, Ö. Unraveling the pore-forming steps of pneumolysin from streptococcus pneumoniae. Nano Lett. 2016, 16, 7915–7924. [Google Scholar] [CrossRef]
- Witzenrath, M.; Gutbier, B.; Hocke, A.C.; Schmeck, B.; Hippenstiel, S.; Berger, K.; Mitchell, T.J.; de los Toyos, J.R.; Rosseau, S.; Suttorp, N.; et al. Role of pneumolysin for the development of acute lung injury in pneumococcal pneumonia. Crit. Care Med. 2006, 34, 1947–1954. [Google Scholar] [CrossRef] [PubMed]
- Sligl, W.I.; Asadi, L.; Eurich, D.T.; Tjosvold, L.; Marrie, T.J.; Majumdar, S.R. Macrolides and mortality in critically ill patients with community-acquired pneumonia: A systematic review and meta-analysis. Crit. Care Med. 2014, 42, 420–432. [Google Scholar] [CrossRef]
- Brown, L.A.; Mitchell, A.M.; Mitchell, T.J. Streptococcus pneumoniae and lytic antibiotic therapy: Are we adding insult to injury during invasive pneumococcal disease and sepsis? J. Med. Microbiol. 2017, 66, 1253–1256. [Google Scholar] [CrossRef]
- Anderson, R.; Feldman, C. Pneumolysin as a potential therapeutic target in severe pneumococcal disease. J. Infect. 2017, 74, 527–544. [Google Scholar] [CrossRef] [Green Version]
- Domon, H.; Maekawa, T.; Yonezawa, D.; Nagai, K.; Oda, M.; Yanagihara, K.; Terao, Y. Mechanism of macrolide-induced inhibition of pneumolysin release involves impairment of autolysin release in macrolide-resistant streptococcus pneumoniae. Antimicrob. Agents Chemother. 2018, 62, e00161-18. [Google Scholar] [CrossRef] [Green Version]
- Escajadillo, T.; Nizet, V. Pharmacological targeting of pore-forming toxins as adjunctive therapy for invasive bacterial infection. Toxins 2018, 10, 542. [Google Scholar] [CrossRef] [Green Version]
- Roveran Genga, K.; Lo, C.; Cirstea, M.; Zhou, G.; Walley, K.R.; Russell, J.A.; Levin, A.; Boyd, J.H. Two-year follow-up of patients with septic shock presenting with low HDL: The effect upon acute kidney injury, death and estimated glomerular filtration rate. J. Intern. Med. 2017, 281, 518–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, M.; Lambeck, S.; Ding, N.; Henken, S.; Kohl, M.; Deigner, H.P.; Enot, D.P.; Igwe, E.I.; Frappart, L.; Kiehntopf, M.; et al. Hepatic induction of cholesterol biosynthesis reflects a remote adaptive response to pneumococcal pneumonia. FASEB J. 2012, 26, 2424–2436. [Google Scholar] [CrossRef] [PubMed]
- Wade, K.R.; Hotze, E.M.; Briles, D.E.; Tweten, R.K. Mouse, but not human, ApoB-100 lipoprotein cholesterol is a potent innate inhibitor of Streptococcus pneumoniae pneumolysin. PLoS Pathog. 2014, 10, e1004353. [Google Scholar] [CrossRef] [Green Version]
- Henry, B.D.; Neill, D.R.; Becker, K.A.; Gore, S.; Bricio-Moreno, L.; Ziobro, R.; Edwards, M.J.; Mühlemann, K.; Steinmann, J.; Kleuser, B.; et al. Engineered liposomes sequester bacterial exotoxins and protect from severe invasive infections in mice. Nat. Biotechnol. 2014, 33, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Laterre, P.-F.; Colin, G.; Dequin, P.-F.; Dugernier, T.; Boulain, T.; Azeredo da Silveira, S.; Lajaunias, F.; Perez, A.; François, B. CAL02, a novel antitoxin liposomal agent, in severe pneumococcal pneumonia: A first-in-human, double-blind, placebo-controlled, randomised trial. Lancet Infect. Dis. 2019, 19, 620–630. [Google Scholar] [CrossRef]
- Yildirim, I.; Crotty, S.; Loh, C.H.; Festag, G.; Weber, C.; Caponi, P.-F.; Gottschaldt, M.; Westerhausen, M.; Schubert, U.S. End-functionalized polylactides using a calcium-based precatalyst: Synthesis and insights by mass spectrometry. J. Polym. Sci. Part A Polym. Chem. 2016, 54, 437–448. [Google Scholar] [CrossRef]
- Yildirim, I.; Yildirim, T.; Kalden, D.; Festag, G.; Fritz, N.; Weber, C.; Schubert, S.; Westerhausen, M.; Schubert, U.S. Retinol initiated poly(lactide)s: Stability upon polymerization and nanoparticle preparation. Polym. Chem. 2017, 8, 4378–4387. [Google Scholar] [CrossRef]
- Leiske, M.N.; Trützschler, A.-K.; Armoneit, S.; Sungur, P.; Hoeppener, S.; Lehmann, M.; Traeger, A.; Schubert, U.S. Mission ImPOxable—Or the unknown utilization of non-toxic poly(2-oxazoline)s as cryoprotectants and surfactants at the same time. J. Mater. Chem. B 2017, 5, 9102–9113. [Google Scholar] [CrossRef]
- Wall, E.C.; Gordon, S.B.; Hussain, S.; Goonetilleke, U.R.S.; Gritzfeld, J.; Scarborough, M.; Kadioglu, A. Persistence of pneumolysin in the cerebrospinal fluid of patients with pneumococcal meningitis is associated with mortality. Clin. Infect. Dis. 2012, 54, 701–705. [Google Scholar] [CrossRef]
- Spreer, A.; Kerstan, H.; Böttcher, T.; Gerber, J.; Siemer, A.; Zysk, G.; Mitchell, T.J.; Eiffert, H.; Nau, R. Reduced release of pneumolysin by Streptococcus pneumoniae in vitro and in vivo after treatment with nonbacteriolytic antibiotics in comparison to ceftriaxone. Antimicrob. Agents Chemother. 2003, 47, 2649–2654. [Google Scholar] [CrossRef] [Green Version]
- Iliev, A.I.; Djannatian, J.R.; Nau, R.; Mitchell, T.J.; Wouters, F.S. Cholesterol-dependent actin remodeling via RhoA and Rac1 activation by the Streptococcus pneumoniae toxin pneumolysin. Proc. Natl. Acad. Sci. USA 2007, 104, 2897–2902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostadinova, E.; Chaput, C.; Gutbier, B.; Lippmann, J.; Sander, L.E.; Mitchell, T.J.; Suttorp, N.; Witzenrath, M.; Opitz, B. NLRP3 protects alveolar barrier integrity by an inflammasome-independent increase of epithelial cell adherence. Sci. Rep. 2016, 6, 30943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Pee, K.; Neuhaus, A.; D’Imprima, E.; Mills, D.J.; Kühlbrandt, W.; Yildiz, Ö. CryoEM structures of membrane pore and prepore complex reveal cytolytic mechanism of Pneumolysin. Elife 2017, 6, e23644. [Google Scholar] [CrossRef] [PubMed]
- Evavold, C.L.; Ruan, J.; Tan, Y.; Xia, S.; Wu, H.; Kagan, J.C. The pore-forming protein gasdermin D regulates interleukin-1 secretion from living macrophages. Immunity 2018, 48, 35–44.e6. [Google Scholar] [CrossRef] [Green Version]
- Lyons, M.A.; Brown, A.J. Metabolism of an oxysterol, 7-ketocholesterol, by sterol 27-hydroxylase in HepG2 cells. Lipids 2001, 36, 701–711. [Google Scholar] [CrossRef]
- Sommer, U.; Herscovitz, H.; Welty, F.K.; Costello, C.E. LC-MS-based method for the qualitative and quantitative analysis of complex lipid mixtures. J. Lipid Res. 2006, 47, 804–814. [Google Scholar] [CrossRef] [Green Version]
- Ioannou, G.N.; Landis, C.S.; Jin, G.-Y.; Haigh, W.G.; Farrell, G.C.; Kuver, R.; Lee, S.P.; Savard, C. Cholesterol crystals in hepatocyte lipid droplets are strongly associated with human nonalcoholic steatohepatitis. Hepatol. Commun. 2019, 3, 776–791. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Williamson, C.D.; Cole, N.B.; Donaldson, J.G. Pitstop 2 is a potent inhibitor of clathrin-independent endocytosis. PLoS ONE 2012, 7, e45799. [Google Scholar] [CrossRef] [Green Version]
- Press, A.T.; Traeger, A.; Pietsch, C.; Mosig, A.; Wagner, M.; Clemens, M.G.; Jbeily, N.; Koch, N.; Gottschaldt, M.; Bézière, N.; et al. Cell type-specific delivery of short interfering RNAs by dye-functionalised theranostic nanoparticles. Nat. Commun. 2014, 5, 5565. [Google Scholar] [CrossRef] [Green Version]
- Solomun, J.I.; Cinar, G.; Mapfumo, P.; Richter, F.; Moek, E.; Hausig, F.; Martin, L.; Hoeppener, S.; Nischang, I.; Traeger, A. Solely aqueous formulation of hydrophobic cationic polymers for efficient gene delivery. Int. J. Pharm. 2021, 593, 120080. [Google Scholar] [CrossRef]
- Kim, S.R.; Ho, M.J.; Lee, E.; Lee, J.W.; Choi, Y.W.; Kang, M.J. Cationic PLGA/Eudragit RL nanoparticles for increasing retention time in synovial cavity after intra-articular injection in knee joint. Int. J. Nanomed. 2015, 10, 5263–5271. [Google Scholar] [CrossRef]
- Aksungur, P.; Demirbilek, M.; Denkbaş, E.B.; Vandervoort, J.; Ludwig, A.; Ünlü, N. Development and characterization of Cyclosporine A loaded nanoparticles for ocular drug delivery: Cellular toxicity, uptake, and kinetic studies. J. Control. Release 2011, 151, 286–294. [Google Scholar] [CrossRef]
- Gurcel, L.; Abrami, L.; Girardin, S.; Tschopp, J.; van der Goot, F.G. Caspase-1 Activation of Lipid Metabolic Pathways in Response to Bacterial Pore-Forming Toxins Promotes Cell Survival. Cell 2006, 126, 1135–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirst, R.A.; Kadioglu, A.; O’callaghan, C.; Andrew, P.W. The role of pneumolysin in pneumococcal pneumonia and meningitis. Clin. Exp. Immunol. 2004, 138, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.; Nel, J.G.; Feldman, C. Multifaceted role of pneumolysin in the pathogenesis of myocardial injury in community-acquired pneumonia. Int. J. Mol. Sci. 2018, 19, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aden, D.P.; Fogel, A.; Plotkin, S.; Damjanov, I.; Knowles, B.B. Controlled synthesis of HBsAg in a differentiated human liver carcinoma-derived cell line. Nature 1979, 282, 615–616. [Google Scholar] [CrossRef]
- Shewell, L.K.; Harvey, R.M.; Higgins, M.A.; Day, C.J.; Hartley-Tassell, L.E.; Chen, A.Y.; Gillen, C.M.; James, D.B.A.; Alonzo, F., 3rd; Torres, V.J.; et al. The cholesterol-dependent cytolysins pneumolysin and streptolysin O require binding to red blood cell glycans for hemolytic activity. Proc. Natl. Acad. Sci. USA 2014, 111, E5312–E5320. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kammann, T.; Hoff, J.; Yildirim, I.; Shkodra, B.; Müller, T.; Weber, C.; Gräler, M.H.; Maus, U.A.; Paton, J.C.; Singer, M.; et al. Intracellularly Released Cholesterol from Polymer-Based Delivery Systems Alters Cellular Responses to Pneumolysin and Promotes Cell Survival. Metabolites 2021, 11, 821. https://doi.org/10.3390/metabo11120821
Kammann T, Hoff J, Yildirim I, Shkodra B, Müller T, Weber C, Gräler MH, Maus UA, Paton JC, Singer M, et al. Intracellularly Released Cholesterol from Polymer-Based Delivery Systems Alters Cellular Responses to Pneumolysin and Promotes Cell Survival. Metabolites. 2021; 11(12):821. https://doi.org/10.3390/metabo11120821
Chicago/Turabian StyleKammann, Tobias, Jessica Hoff, Ilknur Yildirim, Blerina Shkodra, Tina Müller, Christine Weber, Markus H. Gräler, Ulrich A. Maus, James C. Paton, Mervyn Singer, and et al. 2021. "Intracellularly Released Cholesterol from Polymer-Based Delivery Systems Alters Cellular Responses to Pneumolysin and Promotes Cell Survival" Metabolites 11, no. 12: 821. https://doi.org/10.3390/metabo11120821
APA StyleKammann, T., Hoff, J., Yildirim, I., Shkodra, B., Müller, T., Weber, C., Gräler, M. H., Maus, U. A., Paton, J. C., Singer, M., Traeger, A., Schubert, U. S., Bauer, M., & Press, A. T. (2021). Intracellularly Released Cholesterol from Polymer-Based Delivery Systems Alters Cellular Responses to Pneumolysin and Promotes Cell Survival. Metabolites, 11(12), 821. https://doi.org/10.3390/metabo11120821