Metabolomic Analysis of Response to Nitrogen-Limiting Conditions in Yarrowia spp.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
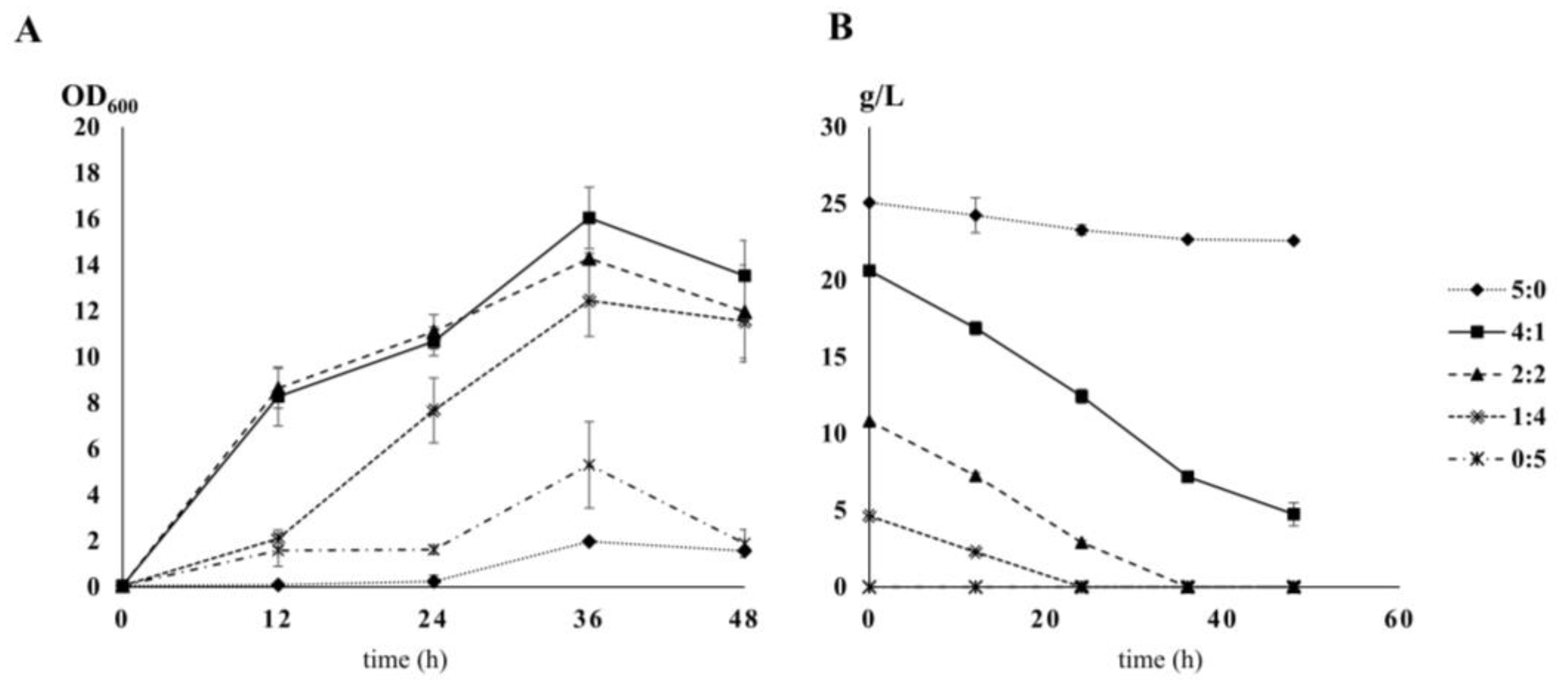
2.1. Growth of Y. Lipolytica under Different Carbon-to-Nitrogen Ratios
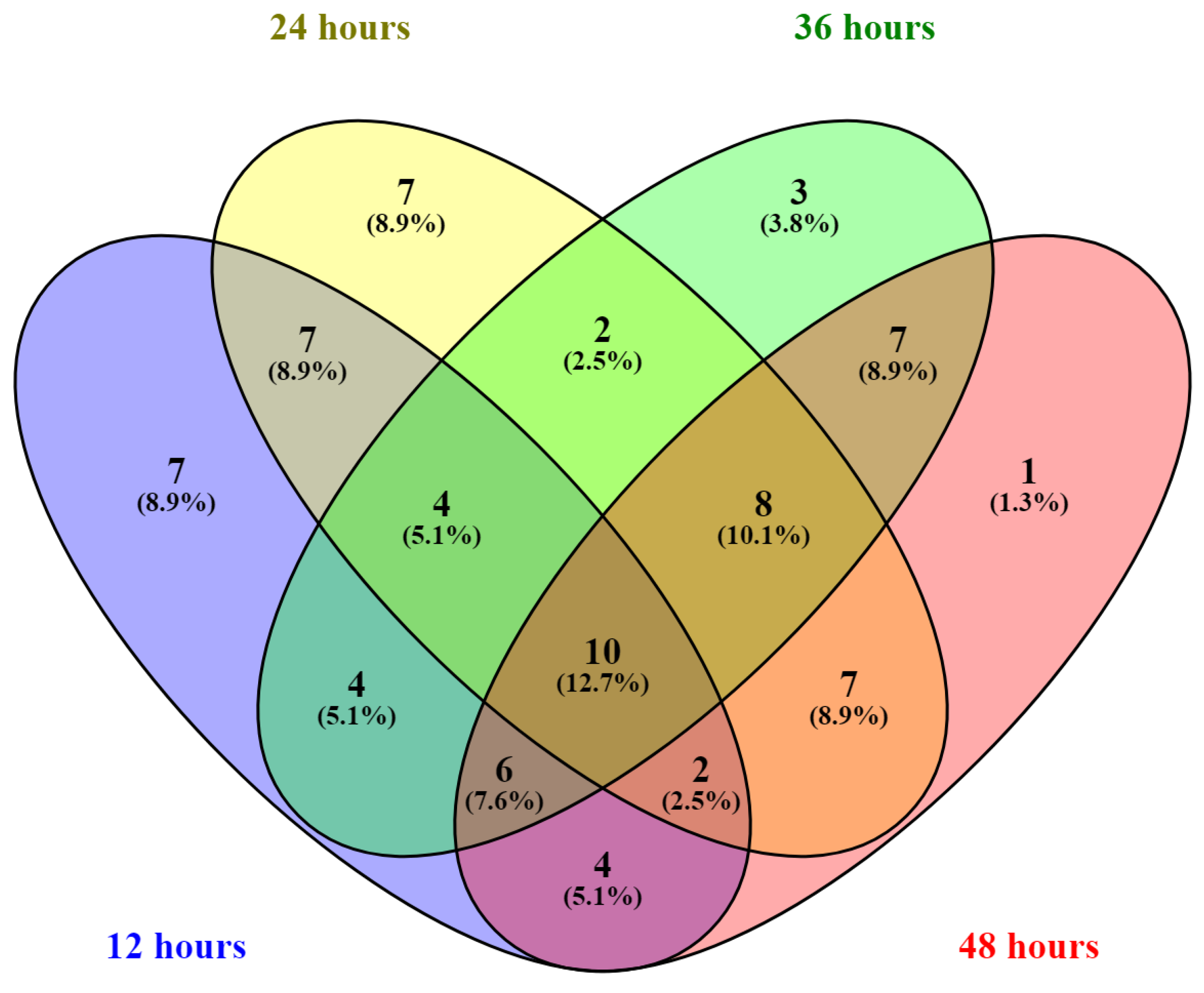
2.2. Intracellular Metabolome Analysis of Y. Lipolytica for Different Carbon-to-Nitrogen Ratios
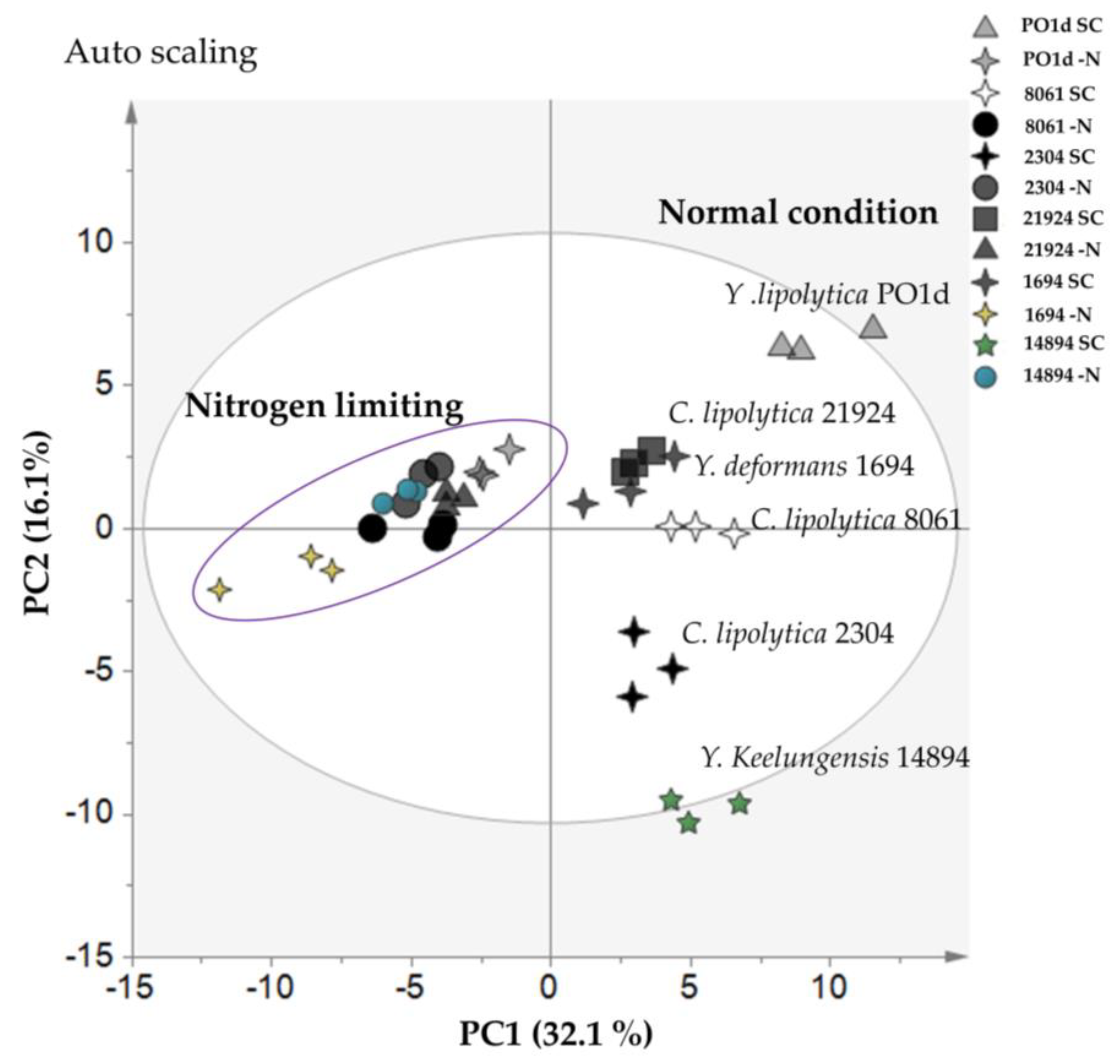
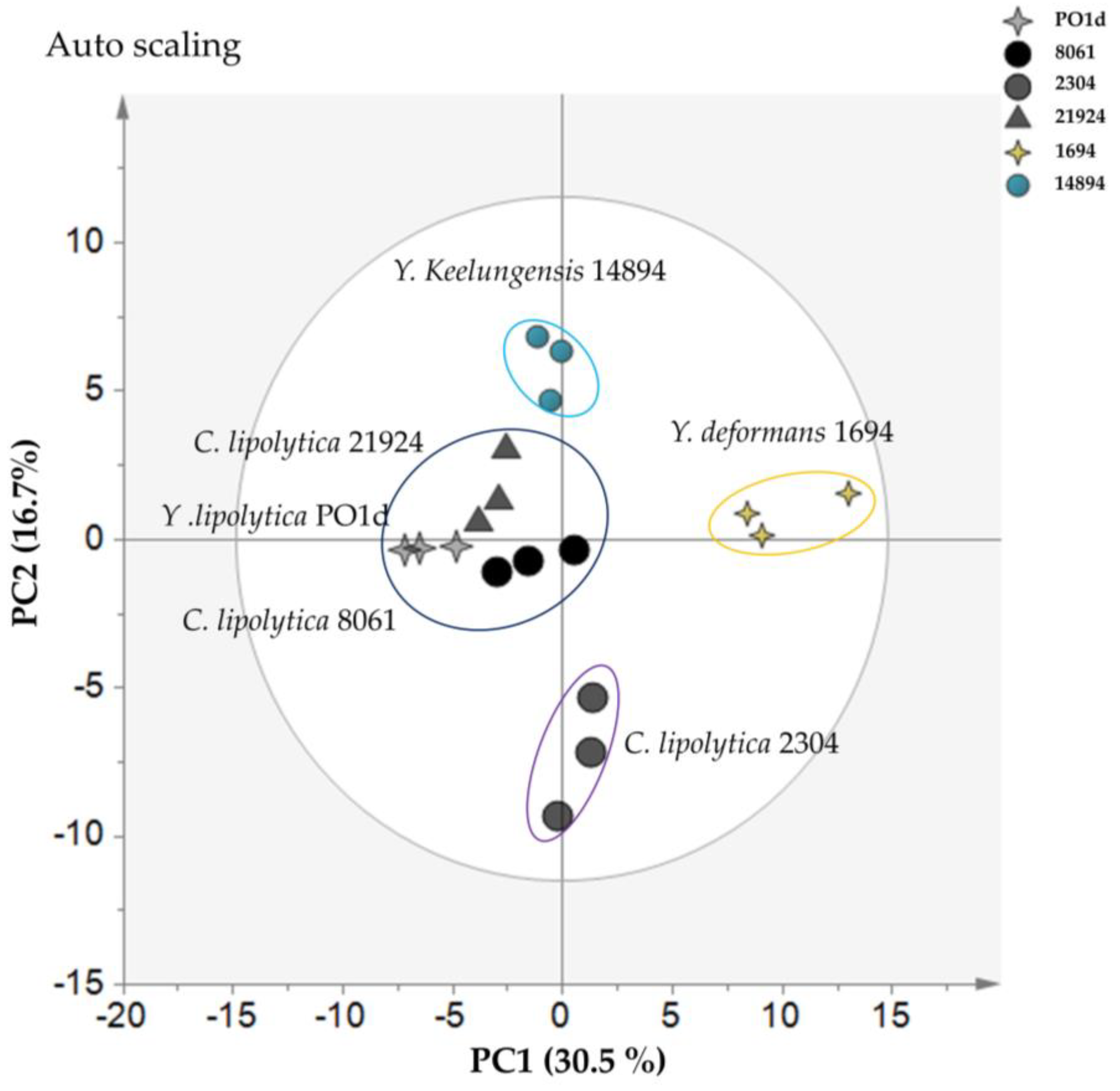
2.3. Intracellular Metabolome Analysis of Different Yarrowia spp. in Normal and Nitrogen-Limiting Conditions
3. Materials and Methods
3.1. Yeast Strains, Cultivation, and Sample Collection
3.2. Fast Filtration and Extraction of Intracellular Metabolites
3.3. Analysis of Intracellular Metabolites by LC-MS/MS
3.4. Multivariate Analysis
3.5. Glucose Measurement
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Papanikolaou, S.; Aggelis, G. Yarrowia lipolytica: A model microorganism used for the production of tailor-made lipids. Eur. J. Lipid Sci. Technol. 2010, 112, 639–654. [Google Scholar] [CrossRef]
- Fickers, P.; Benetti, P.H.; Waché, Y.; Marty, A.; Mauersberger, S.; Smit, M.S.; Nicaud, J.M. Hydrophobic substrate utilisation by the yeast Yarrowia lipolytica, and its potential applications. FEMS Yeast Res. 2005, 5, 527–543. [Google Scholar] [CrossRef]
- Ledesma-Amaro, R.; Nicaud, J.-M. Metabolic Engineering for Expanding the Substrate Range of Yarrowia lipolytica. Trends Biotechnol. 2016, 34, 798–809. [Google Scholar] [CrossRef]
- Zhu, Q.; Jackson, E.N. Metabolic engineering of Yarrowia lipolytica for industrial applications. Curr. Opin. Biotechnol. 2015, 36, 65–72. [Google Scholar] [CrossRef]
- Michely, S.; Gaillardin, C.; Nicaud, J.M.; Neuvéglise, C. Comparative Physiology of Oleaginous Species from the Yarrowia Clade. PLoS ONE 2013, 8, 1–10. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Carnegie, A.J.; Hardy, G.E.S.J.; Smith, D.; Summerell, B.A.; Guarro, J.; Houbraken, J.; Lombard, L.; et al. Fungal Planet Description Sheets: 625–715. In Persoonia—Molecular Phylogeny and Evolution of Fungi; Naturalis Biodiversity Center: Leiden, The Netherlands, 2017; Volume 39, pp. 270–467. [Google Scholar]
- Nagy, E.; Dlauchy, D.; Medeiros, A.O.; Péter, G.; Rosa, C.A. Yarrowia porcina sp. nov. and Yarrowia bubula f.a. sp. nov., two yeast species from meat and river sediment. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2014, 105, 697–707. [Google Scholar] [CrossRef]
- Manikandan, K.; Viruthagiri, T. Optimization of C/N ratio of the medium and fermentation conditions of ethanol production from tapioca starch using co-Culture of Aspergillus niger and Sachormyces cerevisiae. Int. J. ChemTech Res. 2010, 2, 947–955. [Google Scholar]
- Egli, T.; Quayle, J.R. Influence of the carbon: Nitrogen ratio of the growth medium on the cellular composition and the ability of the methylotrophic yeast Hansenula polymorpha to utilize mixed carbon sources. J. Gen. Microbiol. 1986, 132, 1779–1788. [Google Scholar] [CrossRef][Green Version]
- Gao, L.; Sun, M.H.; Liu, X.Z.; Che, Y.S. Effects of carbon concentration and carbon to nitrogen ratio on the growth and sporulation of several biocontrol fungi. Mycol. Res. 2007, 111, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.A.; Bothast, R.J. Carbon concentration and carbon-to-nitrogen ratio influence submerged-culture conidiation by the potential bioherbicide Colletotrichum truncatum NRRL 13737. Appl. Environ. Microbiol. 1990, 56, 3435–3438. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Feng, X. Metabolic engineering of yeast to produce fatty acid-derived biofuels: Bottlenecks and solutions. Front. Microbiol. 2015, 6, 554. [Google Scholar] [CrossRef] [PubMed]
- Ryan, D.; Robards, K. Metabolomics: The greatest omics of them all? Anal. Chem. 2006, 78, 7954–7958. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, A.K.; Robert, V.; Poot, G.A.; Epping, W.; Figge, M.; Holst-Jensen, A.; Skaar, I.; Smith, M.T. Polyphasic re-examination of Yarrowia lipolytica strains and the description of three novel Candida species: Candida oslonensis sp. nov., Candida alimentaria sp. nov. and Candida hollandica sp. nov. Int. J. Syst. Evol. Microbiol. 2007, 57, 2426–2435. [Google Scholar] [CrossRef]
- Barth, G.; Gaillardin, C. Yarrowia lipolytica. In Nonconventional Yeasts in Biotechnology; Springer: Berlin/Heidelberg, Gemany, 1996; pp. 313–388. [Google Scholar]
- Nakase, T.; Goto, S.; Koyama, Y.; Komagata, K.; Iizuka, H. Microbiological Studies on Cheese (II). Food Hyg. Saf. Sci. (Shokuhin Eiseigaku Zasshi) 1977, 18, 353–361. [Google Scholar] [CrossRef]
- Groenewald, M.; Smith, M.T. The teleomorph state of Candida deformans Langeron & Guerra and description of Yarrowia yakushimensis comb. nov. Antonie Van Leeuwenhoek 2013, 103, 1023–1028. [Google Scholar]
- Chang, C.-F.; Chen, C.-C.; Lee, C.-F.; Liu, S.-M. Identifying and characterizing Yarrowia keelungensis sp. nov., an oil-degrading yeast isolated from the sea surface microlayer. Antonie Van Leeuwenhoek 2013, 104, 1117–1123. [Google Scholar] [CrossRef]
- Trygg, J.; Holmes, E.; Lundstedt, T. Chemometrics in metabonomics. J. Proteome Res. 2007, 6, 469–479. [Google Scholar] [CrossRef]
- Vichi, M.; Saporta, G. Clustering and disjoint principal component analysis. Comput. Stat. Data Anal. 2009, 53, 3194–3208. [Google Scholar] [CrossRef]
- Bylesjö, M.; Eriksson, D.; Sjödin, A.; Jansson, S.; Moritz, T.; Trygg, J. Orthogonal projections to latent structures as a strategy for microarray data normalization. BMC Bioinform. 2007, 8, 207. [Google Scholar] [CrossRef]
- Soutourina, J.; Plateau, P.; Blanquet, S. Metabolism of d-Aminoacyl-tRNAs in Escherichia coli and Saccharomyces cerevisiae Cells. J. Biol. Chem. 2000, 275, 32535–32542. [Google Scholar] [CrossRef]
- Ibba, M.; Söll, D. Aminoacyl-tRNAs: Setting the limits of the genetic code. Genes Dev. 2004, 18, 731–738. [Google Scholar] [CrossRef] [PubMed]
- O’donovan, G.A.; Neuhard, J. Pyrimidine Metabolism in Microorganisms. Bacteriol. Rev. 1970, 34, 278. [Google Scholar] [CrossRef] [PubMed]
- Vogels, G.D.; van der Drift, C. Degradation of Purines and Pyrimidines by Microorganisms. Bacteriol. Rev. 1976, 40, 403. [Google Scholar] [CrossRef] [PubMed]
- Henderson, J.F.; Paterson, R.P. Catabolism of purine nucleotides. In Nucleotide Metabolism: An Introduction; Academic Press: New York, NY, USA, 1973; pp. 3–20. [Google Scholar]
- De Virgilio, C.; Hottiger, T.; Dominguez, J.; Boller, T.; Wiemken, A. The role of trehalose synthesis for the acquisition of thermotolerance in yeast I. Genetic evidence that trehalose is a thermoprotectant. Eur. J. Biochem. 1994, 219, 179–186. [Google Scholar] [CrossRef]
- Hottiger, T.; de Virgilio, C.; Hall, M.N.; Boller, T.; Wiemken, A. The role of trehalose synthesis for the acquisition of thermotolerance in yeast II. Physiological concentrations of trehalose increase the thermal stability of proteins in vitro. Eur. J. Biochem. 1994, 219, 187–193. [Google Scholar] [CrossRef]
- Leslie, B.; Teter, S.A.; Crowe, L.M.; Crowe, J.H. Trehalose lowers membrane phase transitions in dry yeast cells. Biochim. Biophys. Acta BBA Biomembr. 1994, 1192, 7–13. [Google Scholar] [CrossRef]
- Soliman, S.S.M.; Tsao, R.; Raizada, M.N. Chemical inhibitors suggest endophytic fungal paclitaxel is derived from both mevalonate and non-mevalonate-like pathways. J. Nat. Prod. 2011, 74, 2497–2504. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Dempo, Y.; Ohta, E.; Nakayama, Y.; Bamba, T.; Fukusaki, E. Molar-based targeted metabolic profiling of cyanobacterial strains with potential for biological production. Metabolites 2014, 4, 499–516. [Google Scholar] [CrossRef]
- Tsugawa, H.; Arita, M.; Kanazawa, M.; Ogiwara, A.; Bamba, T.; Fukusaki, E. MRMPROBS: A data assessment and metabolite identification tool for large-scale multiple reaction monitoring based widely targeted metabolomics. Anal. Chem. 2013, 85, 5191–5199. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dissook, S.; Putri, S.P.; Fukusaki, E. Metabolomic Analysis of Response to Nitrogen-Limiting Conditions in Yarrowia spp. Metabolites 2021, 11, 16. https://doi.org/10.3390/metabo11010016
Dissook S, Putri SP, Fukusaki E. Metabolomic Analysis of Response to Nitrogen-Limiting Conditions in Yarrowia spp. Metabolites. 2021; 11(1):16. https://doi.org/10.3390/metabo11010016
Chicago/Turabian StyleDissook, Sivamoke, Sastia Prama Putri, and Eiichiro Fukusaki. 2021. "Metabolomic Analysis of Response to Nitrogen-Limiting Conditions in Yarrowia spp." Metabolites 11, no. 1: 16. https://doi.org/10.3390/metabo11010016
APA StyleDissook, S., Putri, S. P., & Fukusaki, E. (2021). Metabolomic Analysis of Response to Nitrogen-Limiting Conditions in Yarrowia spp. Metabolites, 11(1), 16. https://doi.org/10.3390/metabo11010016