Metabolomics Analysis Reveals Global Metabolic Changes in the Evolved E. coli Strain with Improved Growth and 1-Butanol Production in Minimal Medium


 and
and 
Abstract
1. Introduction
2. Results and Discussion
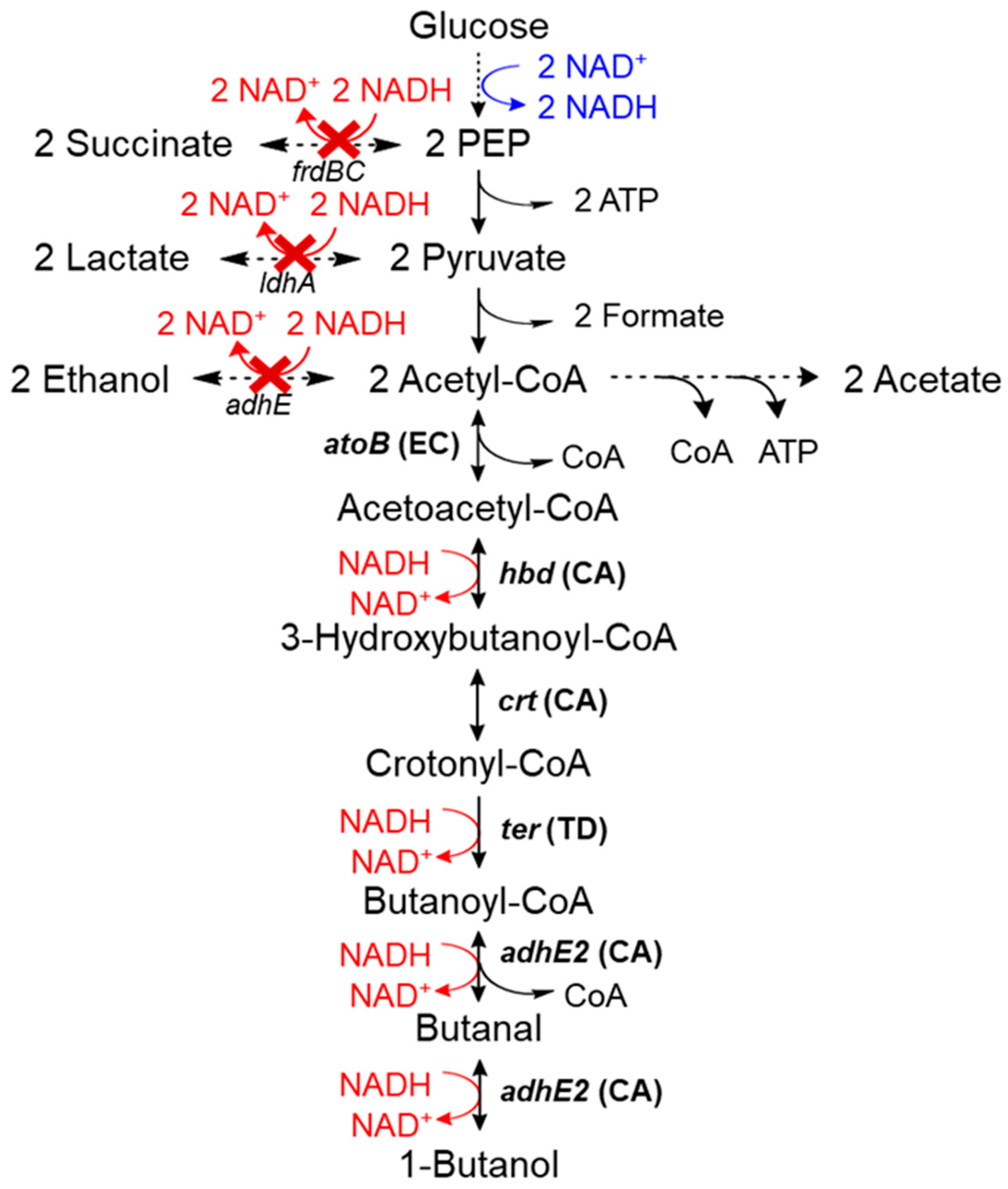
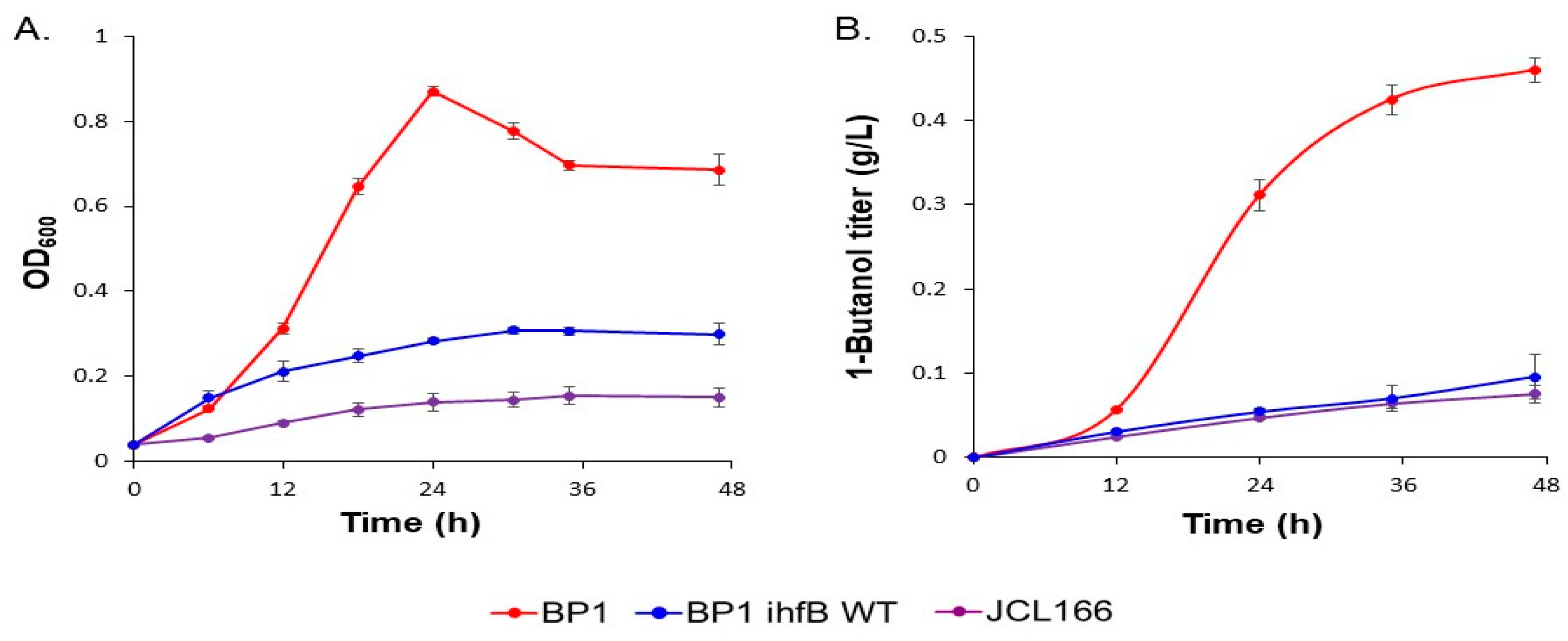
2.1. ALE Resulted in Global Metabolic Changes That Enhanced Growth and 1-Butanol Production in E. coli Strain BP1
2.2. Comparative Metabolic Profiling of Parental Strain (JC166) and Evolved Strain (BP1) Revealed Differences in Intracellular Levels of Several Metabolite Groups
2.3. Comparative Metabolic Profiling of BP1 and Wild-Type Reversion Strain BP1 ihfB WT
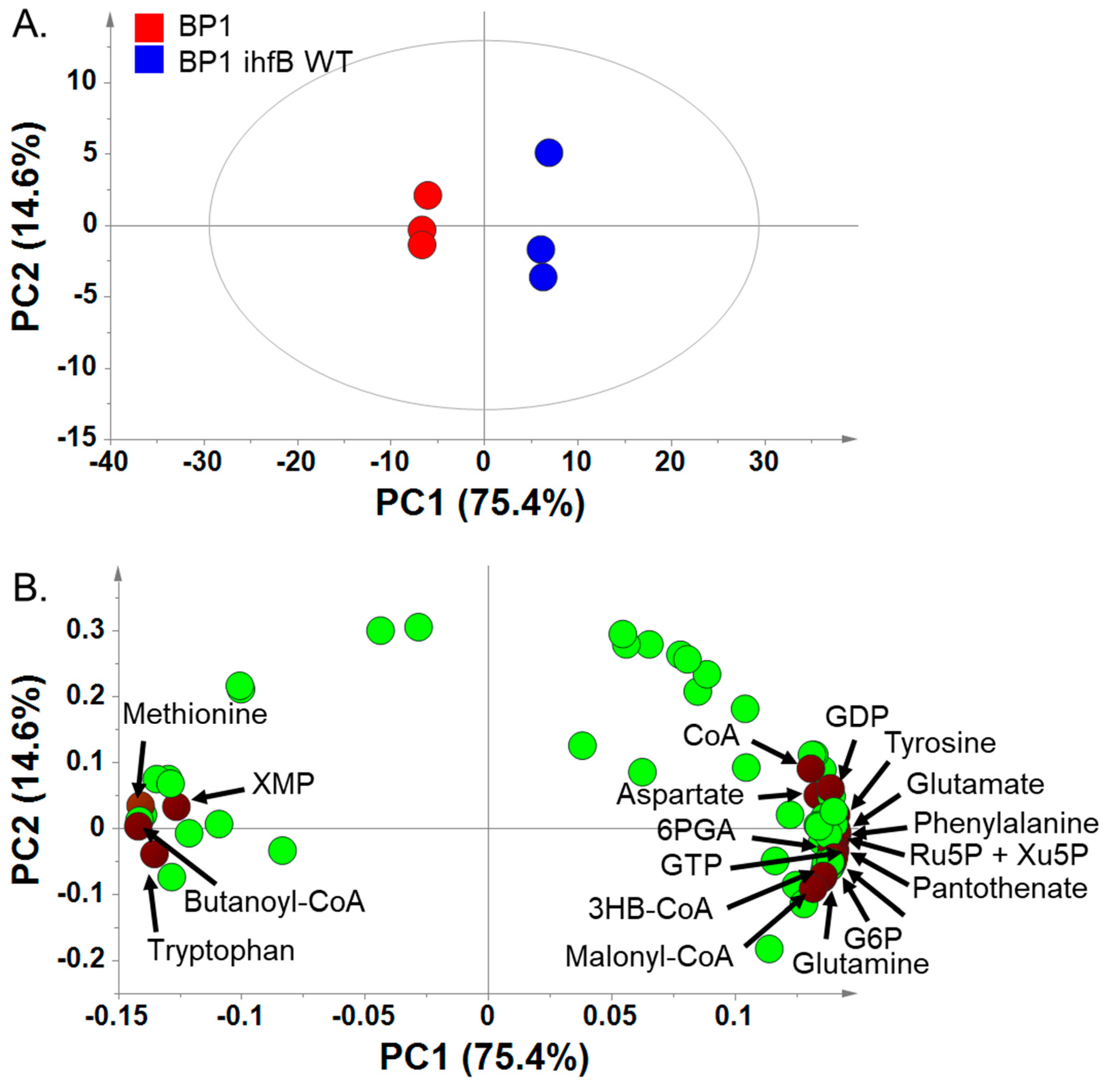
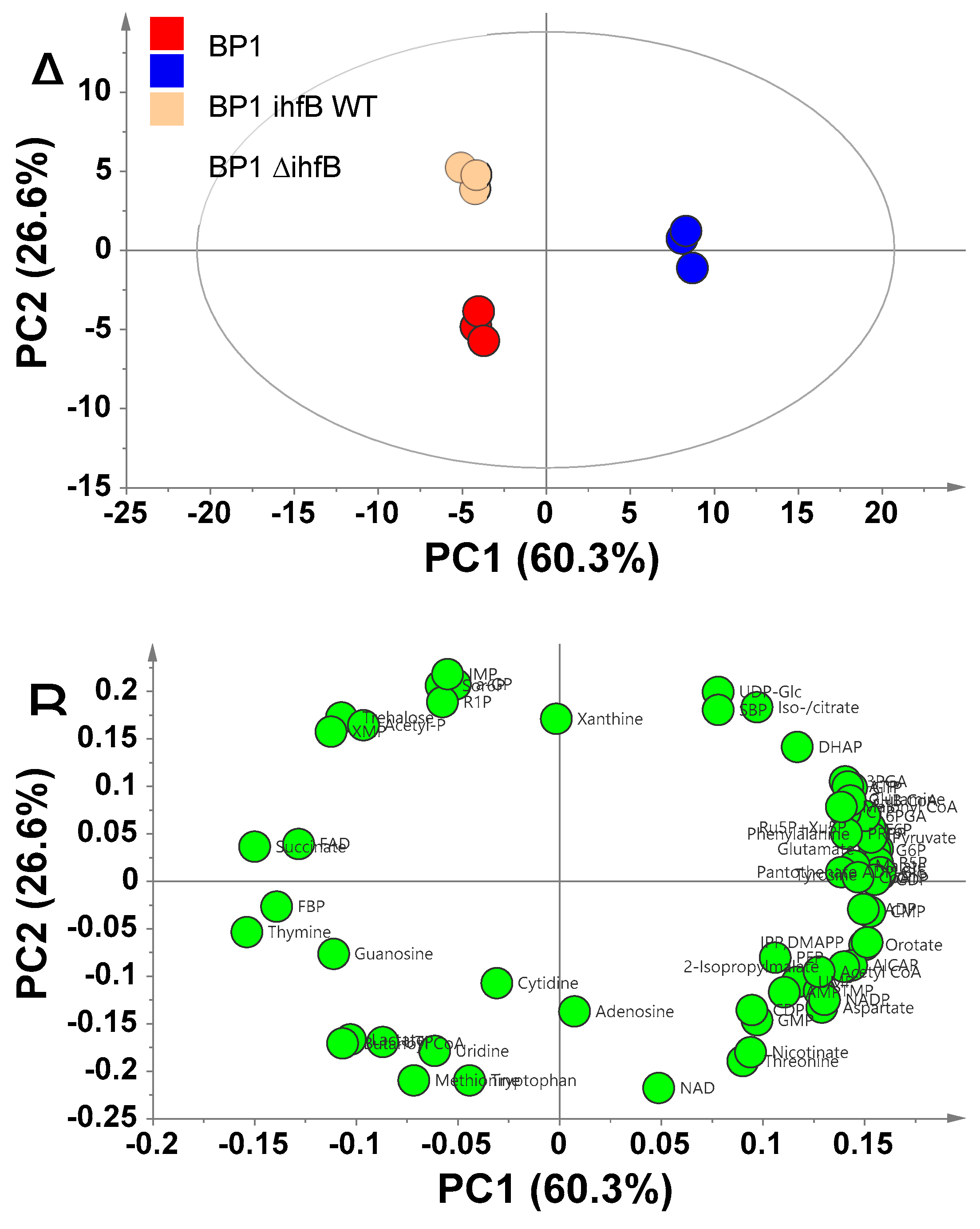
2.3.1. Principal Component Analysis of BP1 and BP1 ihfB WT Reveals That Mutation in ihfB Has a Large Effect on the Metabolome Profile
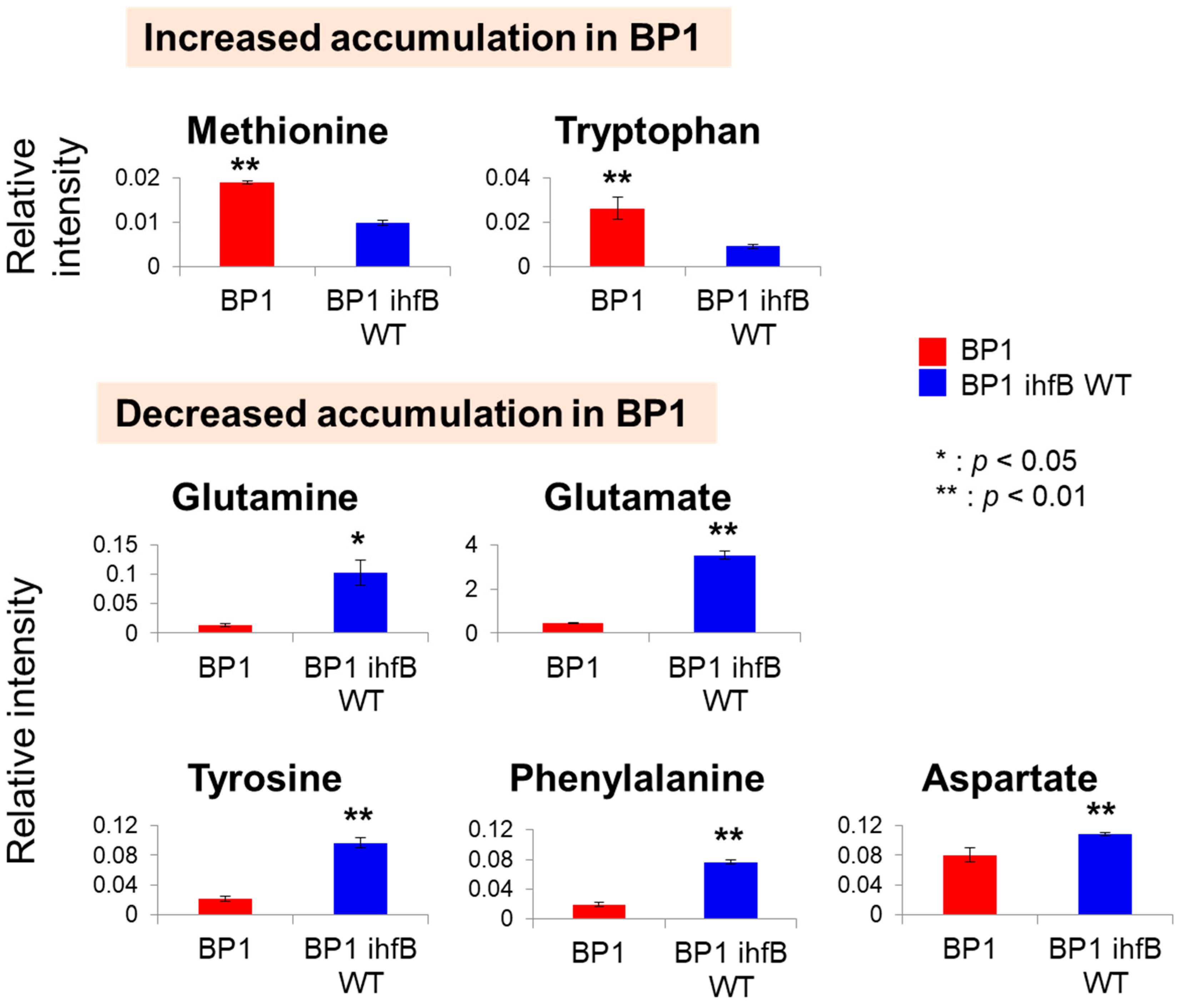
2.3.2. Mutation of ihfB in BP1 Affects Intracellular Amino Acid Levels
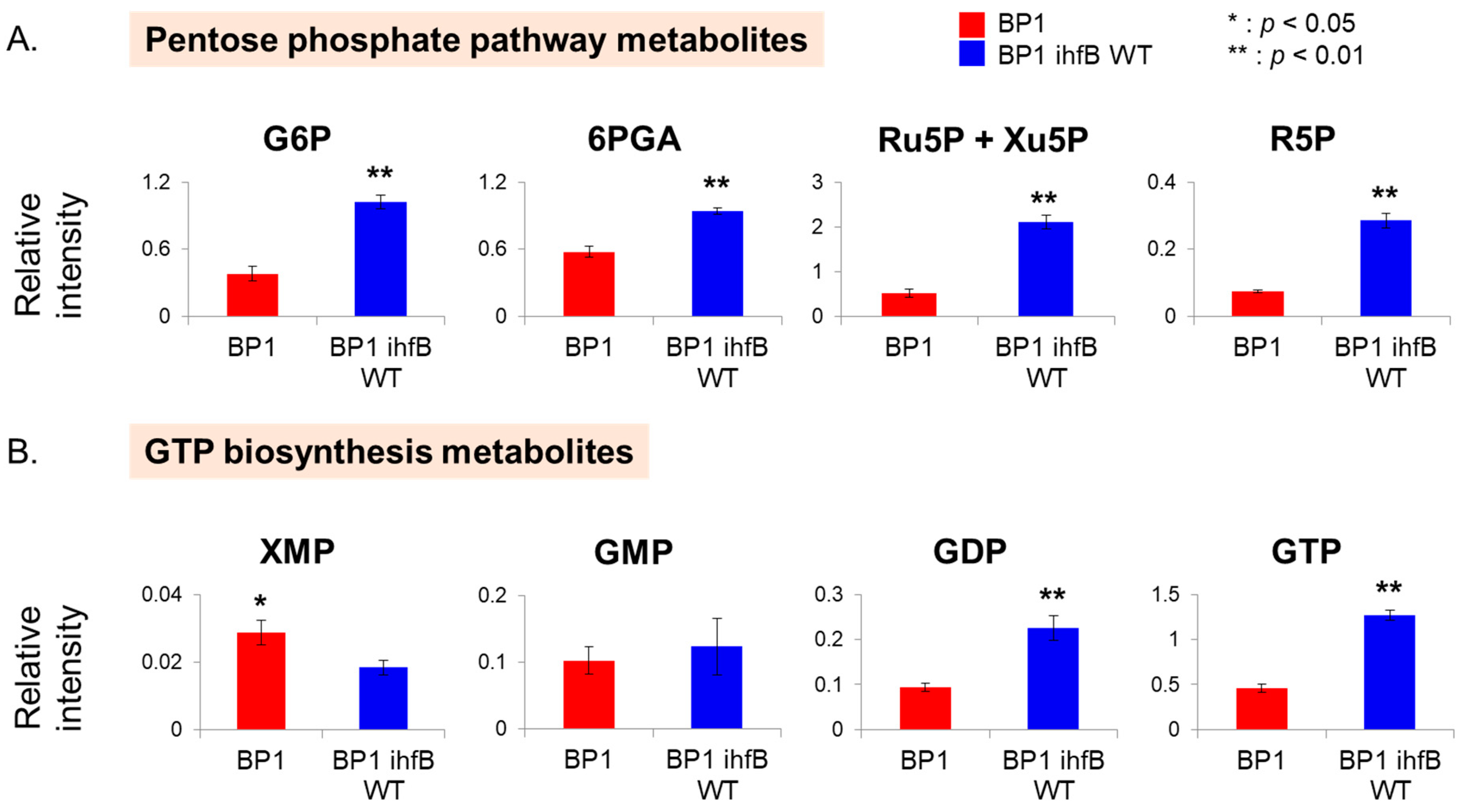
2.3.3. Mutation of ihfB in BP1 Affects Intracellular Levels of the Pentose Phosphate Pathway and GTP Biosynthesis Metabolites
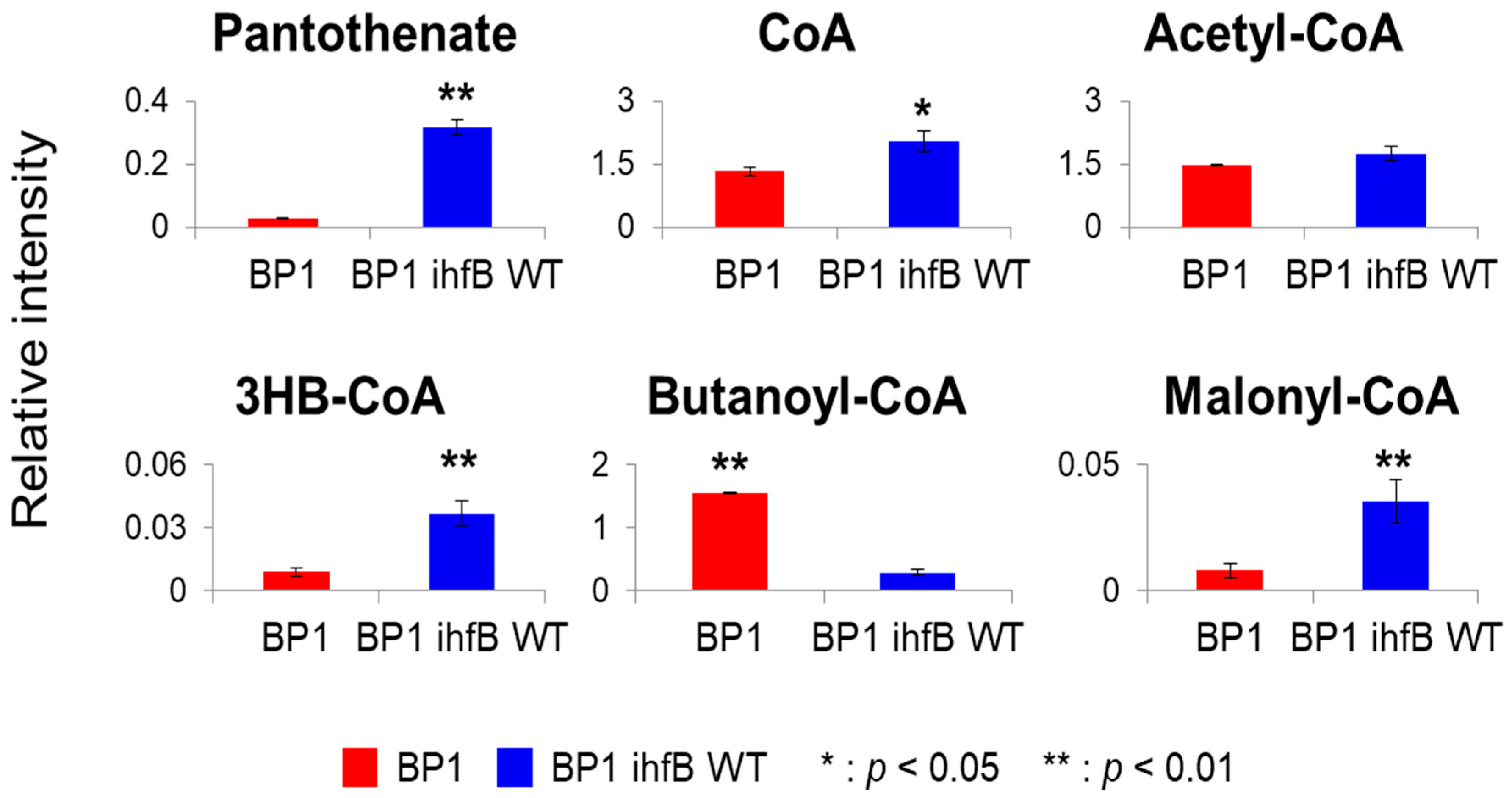
2.3.4. Mutation of ihfB in BP1 Affects Intracellular Levels of CoA-Related Metabolites
3. Materials and Methods
3.1. E. coli Strains and Plasmids
3.2. Medium and Growth Conditions
3.3. Metabolome Sampling and Extraction
3.4. Analysis of Intracellular Metabolites
3.5. Multivariate Analysis
3.6. Analysis of Extracellular Metabolites
3.7. Construction of Wild-Type Reversion Strains
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, D.T.; Woods, D.R. Acetone-Butanol Fermentation Revisited. Microbiol. Rev. 1986, 50, 484–524. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, S.; Cann, A.F.; Connor, M.R.; Shen, C.R.; Smith, K.M.; Brynildsen, M.P.; Chou, K.J.Y.; Hanai, T.; Liao, J.C. Metabolic engineering of Escherichia coli for 1-butanol production. Metab. Eng. 2008, 10, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.R.; Lan, E.I.; Dekishima, Y.; Baez, A.; Cho, K.M.; Liao, J.C. Driving forces enable high-titer anaerobic 1-butanol synthesis in Escherichia coli. Appl. Environ. Microbiol. 2011, 77, 2905–2915. [Google Scholar] [CrossRef] [PubMed]
- Nitta, K.; Laviña, W.A.; Pontrelli, S.; Liao, J.C.; Putri, S.P.; Fukusaki, E. Orthogonal partial least squares / projections to latent structures regression-based metabolomics approach for identification of gene targets for improvement of 1-butanol production in Escherichia coli. J. Biosci. Bioeng. 2017, 124, 498–505. [Google Scholar] [CrossRef]
- Nitta, K.; Laviña, W.A.; Pontrelli, S.; Liao, J.C.; Putri, S.P.; Fukusaki, E. Metabolome analysis revealed the knockout of glyoxylate shunt as an effective strategy for improvement of 1-butanol production in transgenic Escherichia coli. J. Biosci Bioeng. 2018, 127, 301–308. [Google Scholar] [CrossRef]
- Ohtake, T.; Pontrelli, S.; Laviña, W.A.; Liao, J.C.; Putri, S.P.; Fukusaki, E. Metabolomics-driven approach to solving a CoA imbalance for improved 1-butanol production in Escherichia coli. Metab. Eng. 2017, 41, 135–143. [Google Scholar] [CrossRef]
- Pontrelli, S.; Fricke, R.C.B.; Sakurai, S.S.M.; Putri, S.P.; Fitz-Gibbon, S.; Chung, M.; Wu, H.Y.; Chen, Y.J.; Pellegrini, M.; Fukusaki, E.; et al. Directed strain evolution restructures metabolism for 1-butanol production in minimal media. Metab Eng. 2018, 49, 153–163. [Google Scholar] [CrossRef]
- Zhang, J.; Greasham, R. Chemically defined media for commercial fermentations. Appl. Microbiol. Biotechnol. 1999, 51, 407–421. [Google Scholar] [CrossRef]
- Müller, J.; Beckers, M.; Mußmann, N.; Bongaerts, J.; Büchs, J. Elucidation of auxotrophic deficiencies of Bacillus pumilus DSM 18097 to develop a defined minimal medium. Microb. Cell Fact. 2018, 17, 106. [Google Scholar] [CrossRef]
- Blaby, I.K.; Lyons, B.J.; Wroclawska-Hughes, E.; Phillips, G.C.F.; Pyle, T.P.; Chamberlin, S.G.; Benner, S.A.; Lyons, T.J.; Crecy-Lagard, V.; Crécy, E. Experimental evolution of a facultative thermophile from a mesophilic ancestor. Appl. Environ. Microbiol. 2012, 78, 144–155. [Google Scholar] [CrossRef]
- Horinouchi, T.; Sakai, A.; Kotani, H.; Tanabe, K.; Furusawa, C. Improvement of isopropanol tolerance of Escherichia coli using adaptive laboratory evolution and omics technologies. J. Biotech. 2017, 255, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Matsusako, T.; Toya, Y.; Yoshikawa, K.; Shimizu, H. Identification of alcohol stress tolerance genes of Synechocystis sp. PCC 6803 using adaptive laboratory evolution. Biotechnol. Biofuels 2017, 10, 307. [Google Scholar] [CrossRef] [PubMed]
- Reyes, L.H.; Almario, M.P.; Winkler, J.; Orozco, M.M.; Kao, K.C. Visualizing evolution in real time to determine the molecular mechanisms of n-butanol tolerance in Escherichia coli. Metab. Eng. 2012, 14, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Conrad, T.M.; Frazier, M.; Joyce, A.R.; Cho, B.-K.; Knight, E.M.; Lewis, N.E.; Landick, R.; Palsson, B.Ø. RNA polymerase mutants found through adaptive evolution reprogram Escherichia coli for optimal growth in minimal media. Proc. Natl. Acad. Sci. USA 2010, 107, 20500–20505. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Palsson, B.O. Adaptive evolution of Escherichia coli K-12 MG1655 during growth on a nonnative carbon source, L-l,2-propanediol. Appl. Environ. Microbiol. 2010, 76, 4158–4168. [Google Scholar] [CrossRef] [PubMed]
- Sonderegger, M.; Sauer, U. Evolutionary Engineering of Saccharomyces cerevisiae for Anaerobic Growth on Xylose. Appl Environ. Microbiol. 2003, 69, 1990–1998. [Google Scholar] [CrossRef]
- Tang, J. Microbial Metabolomics. Curr Genomics. 2011, 12, 391–403. [Google Scholar] [CrossRef]
- Fang, X.; Sastry, A.; Mih, N.; Kim, D.; Tan, J.; Yurkovich, J.T.; Colton, J.L.; Gao, Y.; Yang, L.; Palsson, B.O. Global transcriptional regulatory network for Escherichia coli robustly connects gene expression to transcription factor activities. Proc. Natl. Acad. Sci. USA 2017, 114, 201702581. [Google Scholar] [CrossRef]
- Martínez-Antonio, A.; Collado-Vides, J. Identifying global regulators in transcriptional regulatory networks in bacteria. Curr. Opin. Microbiol. 2003, 6, 482–489. [Google Scholar] [CrossRef]
- Arfin, S.M.; Long, A.D.; Ito, E.T.; Tolleri, L.; Riehle, M.M.; Paegle, E.S.; Hatfield, W.G. Global Gene Expression Profiling in Escherichia coli K12: The Effects of Integration Host Factor. J. Biol. Chem. 2005, 275, 29672–29684. [Google Scholar] [CrossRef]
- Paul, L.; Blumenthal, R.M. Activation from a Distance: Roles of Lrp and Integration Host Factor in Transcriptional Activation of gltBDF. J. Bacteriol. 2001, 183, 3910–3918. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Alonso, A.P.; Allen, D.K.; Reed, J.L.; Shachar-hill, Y. Synergy between 13 C-metabolic flux analysis and flux balance analysis for understanding metabolic adaption to anaerobiosis in E. coli. Metab. Eng. 2011, 13, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Kargeti, M.; Venkatesh, K.V. The effect of global transcriptional regulators on the anaerobic fermentative metabolism of Escherichia coli. Mol. Biosyst. 2017, 13, 1388–1398. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Arita, M.; Kanazawa, M.; Ogiwara, A.; Bamba, T.; Fukusaki, E. MRMPROBS: A data assessment and metabolite identification tool for large-scale multiple reaction monitoring based widely targeted metabolomics. Anal. Chem. 2013, 85, 5191–5199. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K- 12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Description or Genotype | Source |
|---|---|---|
| JCL166 | BW25113/F’ [traD36 proAB+ lacIqZΔM15 (Tetr)] ΔldhA ΔadhE ΔfrdBC | [2] |
| BP1 | Evolved JCL166 strain functioning in minimal media | [7] |
| BP1 ihfB WT | BP1 with wild-type ihfB sequence | [7] |
| BP1 ∆ihfB | BP1 with ihfB deleted | [7] |
| Plasmids | Description or genotype | Source |
| pEL11 | PLlacO1::atoBEC-adhE2CA-crtCA-hbdCA ColE1 ori Ampr | [3] |
| pIM8 | PLlacO1::terTD ColA ori Kanr | [3] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laviña, W.A.; Sakurai, S.S.M.; Pontrelli, S.; Putri, S.P.; Fukusaki, E. Metabolomics Analysis Reveals Global Metabolic Changes in the Evolved E. coli Strain with Improved Growth and 1-Butanol Production in Minimal Medium. Metabolites 2020, 10, 192. https://doi.org/10.3390/metabo10050192
Laviña WA, Sakurai SSM, Pontrelli S, Putri SP, Fukusaki E. Metabolomics Analysis Reveals Global Metabolic Changes in the Evolved E. coli Strain with Improved Growth and 1-Butanol Production in Minimal Medium. Metabolites. 2020; 10(5):192. https://doi.org/10.3390/metabo10050192
Chicago/Turabian StyleLaviña, Walter A., Sana Subhan Memon Sakurai, Sammy Pontrelli, Sastia Prama Putri, and Eiichiro Fukusaki. 2020. "Metabolomics Analysis Reveals Global Metabolic Changes in the Evolved E. coli Strain with Improved Growth and 1-Butanol Production in Minimal Medium" Metabolites 10, no. 5: 192. https://doi.org/10.3390/metabo10050192
APA StyleLaviña, W. A., Sakurai, S. S. M., Pontrelli, S., Putri, S. P., & Fukusaki, E. (2020). Metabolomics Analysis Reveals Global Metabolic Changes in the Evolved E. coli Strain with Improved Growth and 1-Butanol Production in Minimal Medium. Metabolites, 10(5), 192. https://doi.org/10.3390/metabo10050192