


Exploring Cannabidiol’s Therapeutic Role in Colorectal Cancer: Network Pharmacology and Molecular Docking Insights

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Acquisition
2.2. Enrichment of Gene Ontology and Metabolic Pathways
2.3. Construction of Protein–Protein Interaction Network
2.4. Molecular Docking Analysis Between Cannabidiol and Hub Genes
2.5. Analysis of Expression and Infiltration of Immune Cells
3. Results
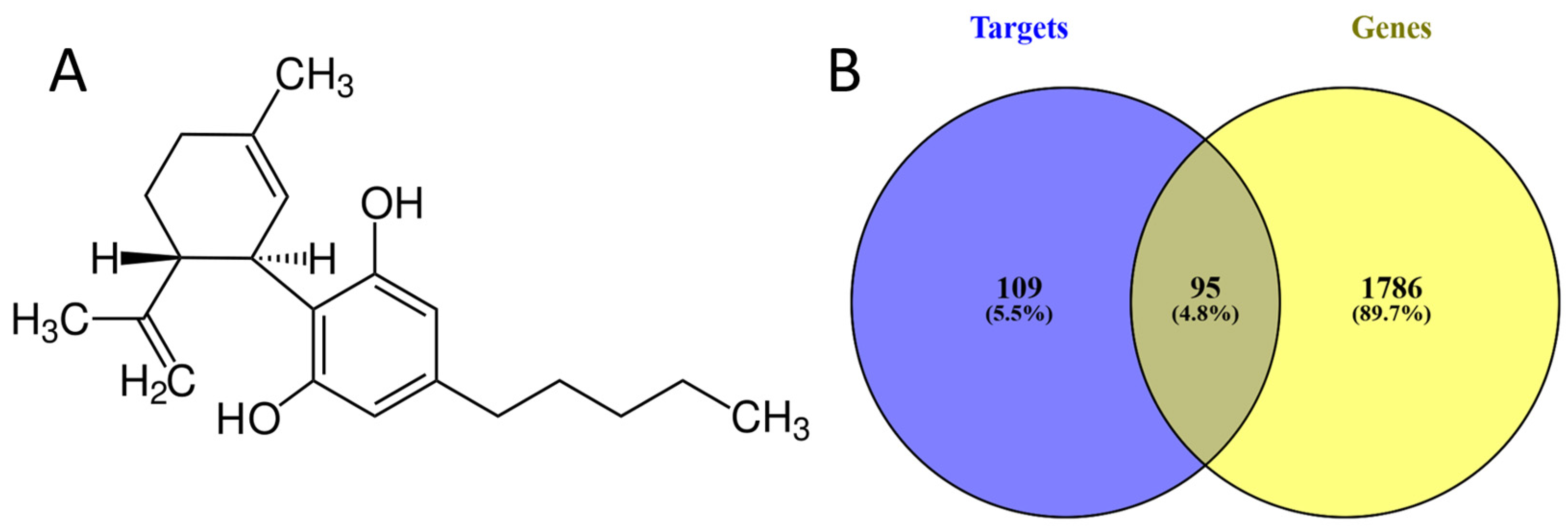
3.1. Analysis of Cannabidiol Targets and CRC-Associated Genes
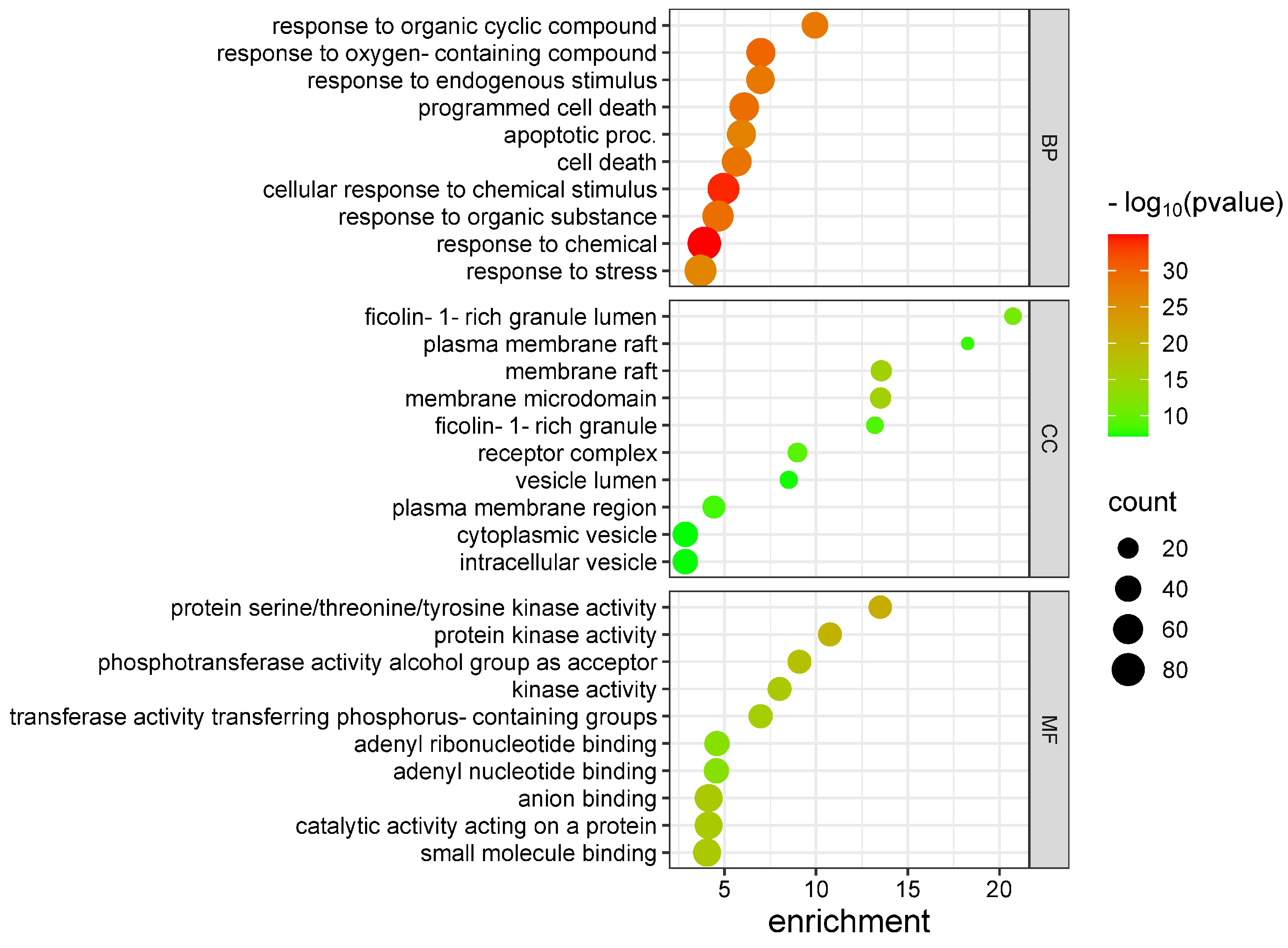
3.2. Gene Ontology Enrichment Analysis and KEEG Pathways
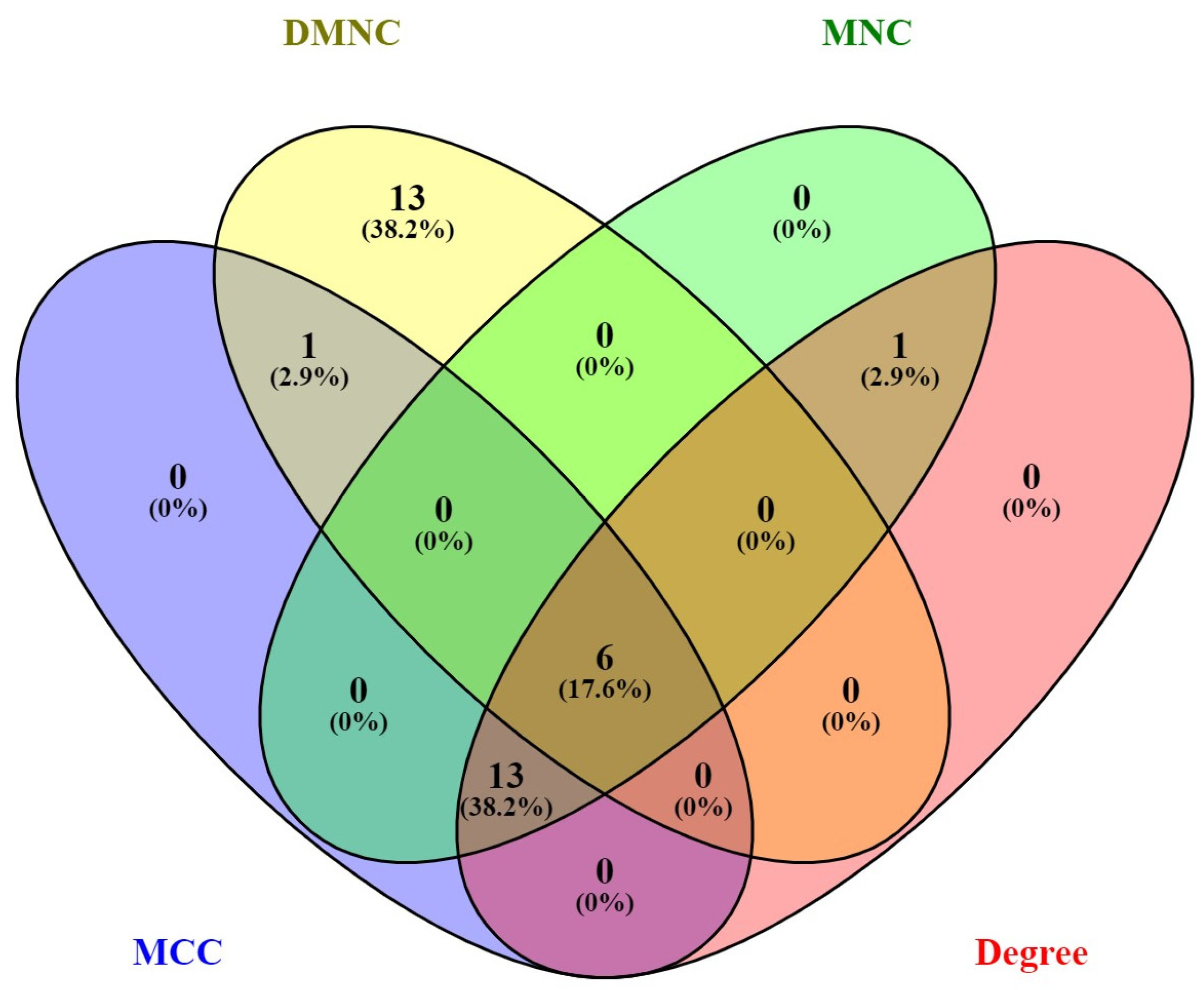
3.3. Construction of the Protein–Protein Interaction Network and Prediction of Hub Genes
3.4. Molecular Docking Between Cannabidiol and Hub Genes
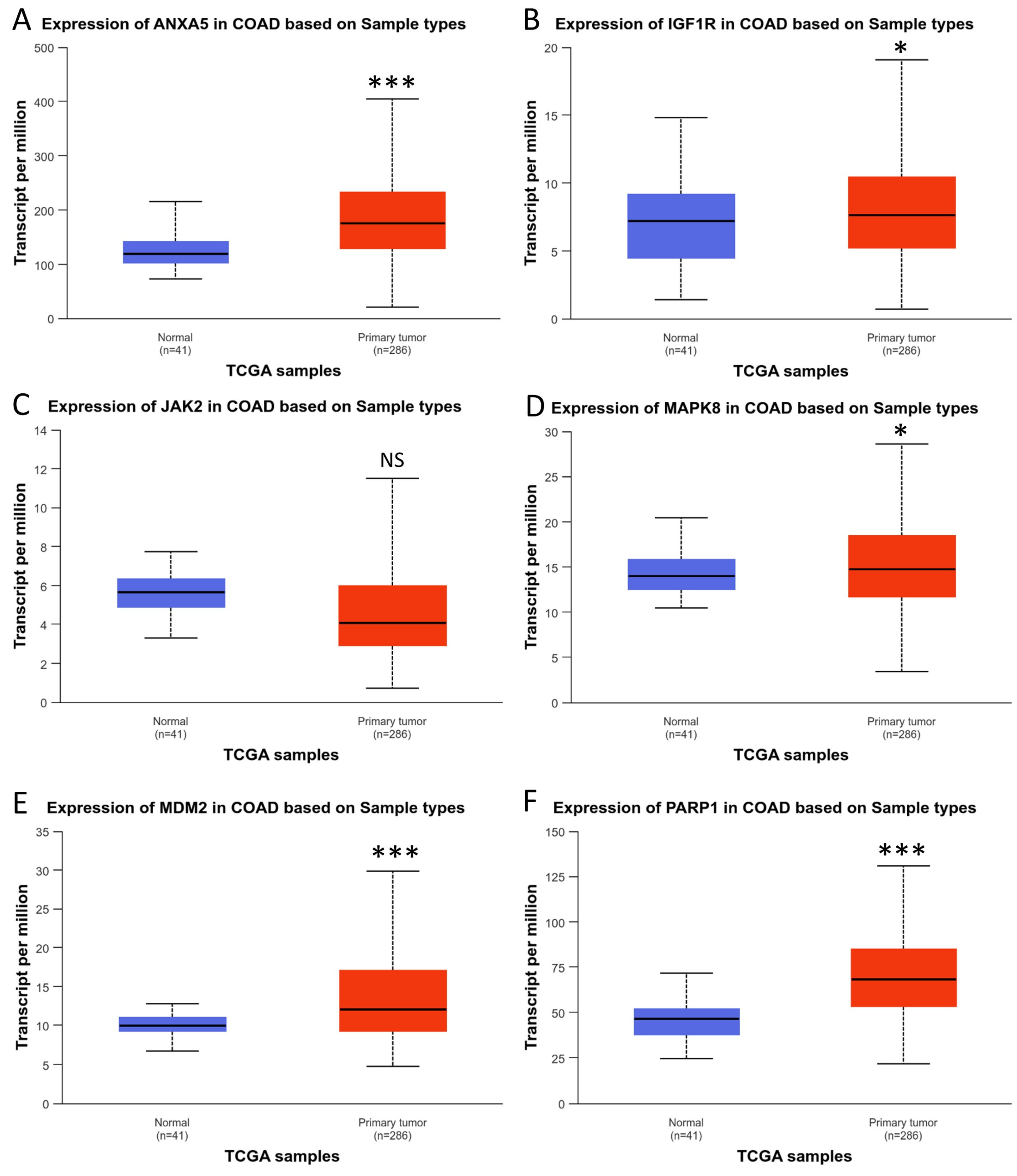
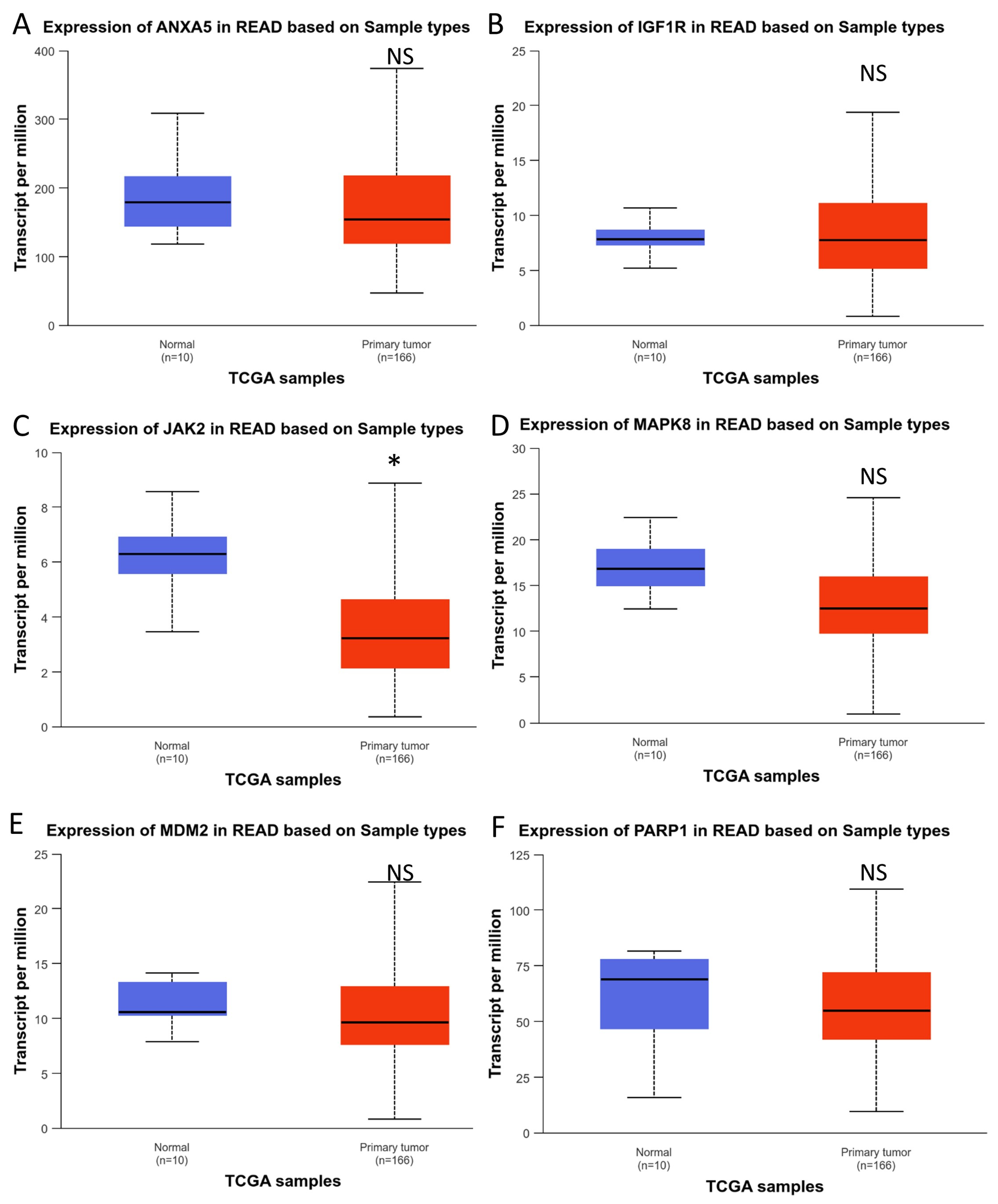
3.5. Analysis of Hub Gene Expression in CRC
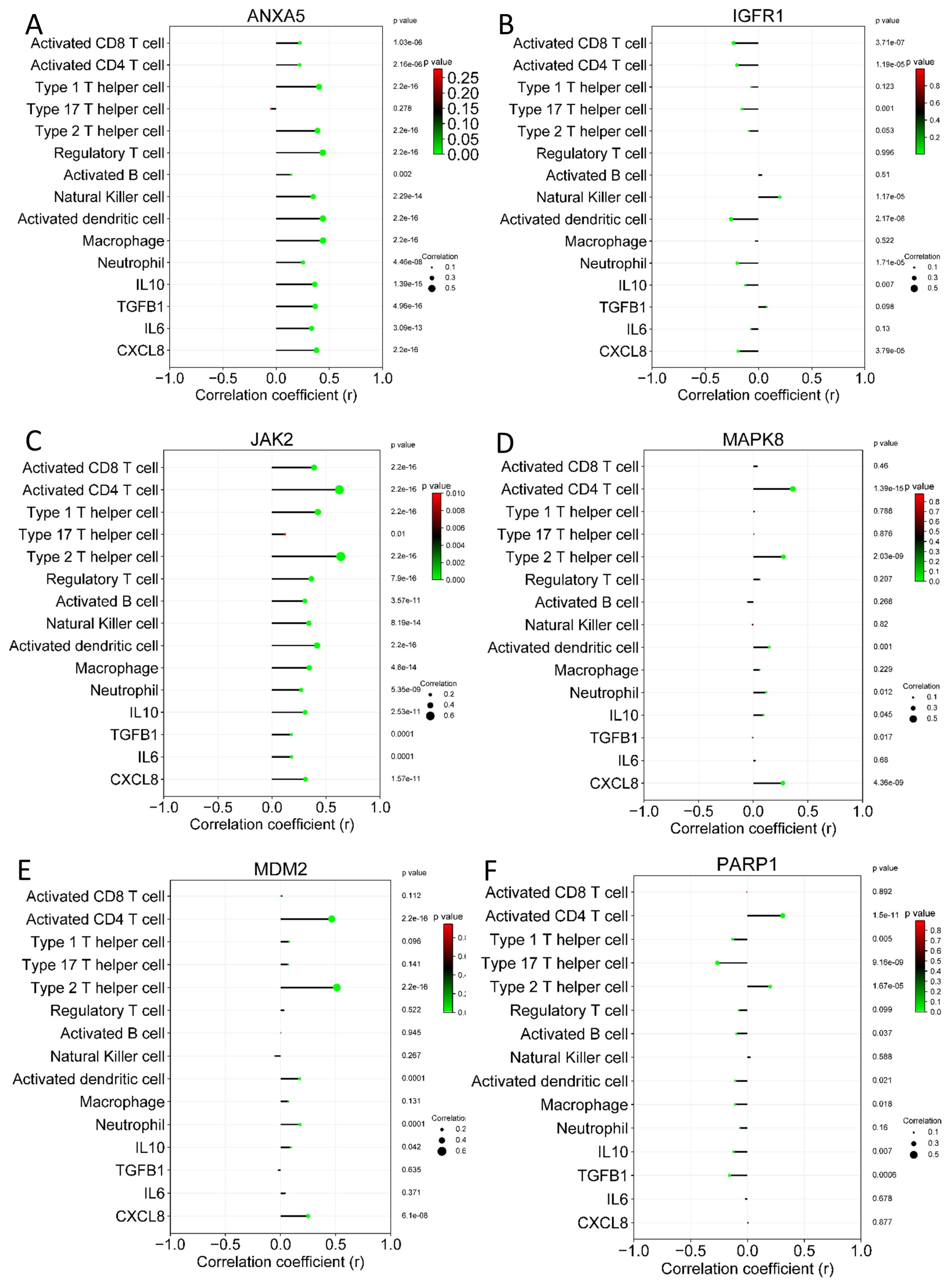
3.6. Immune Cell Infiltration Analysis
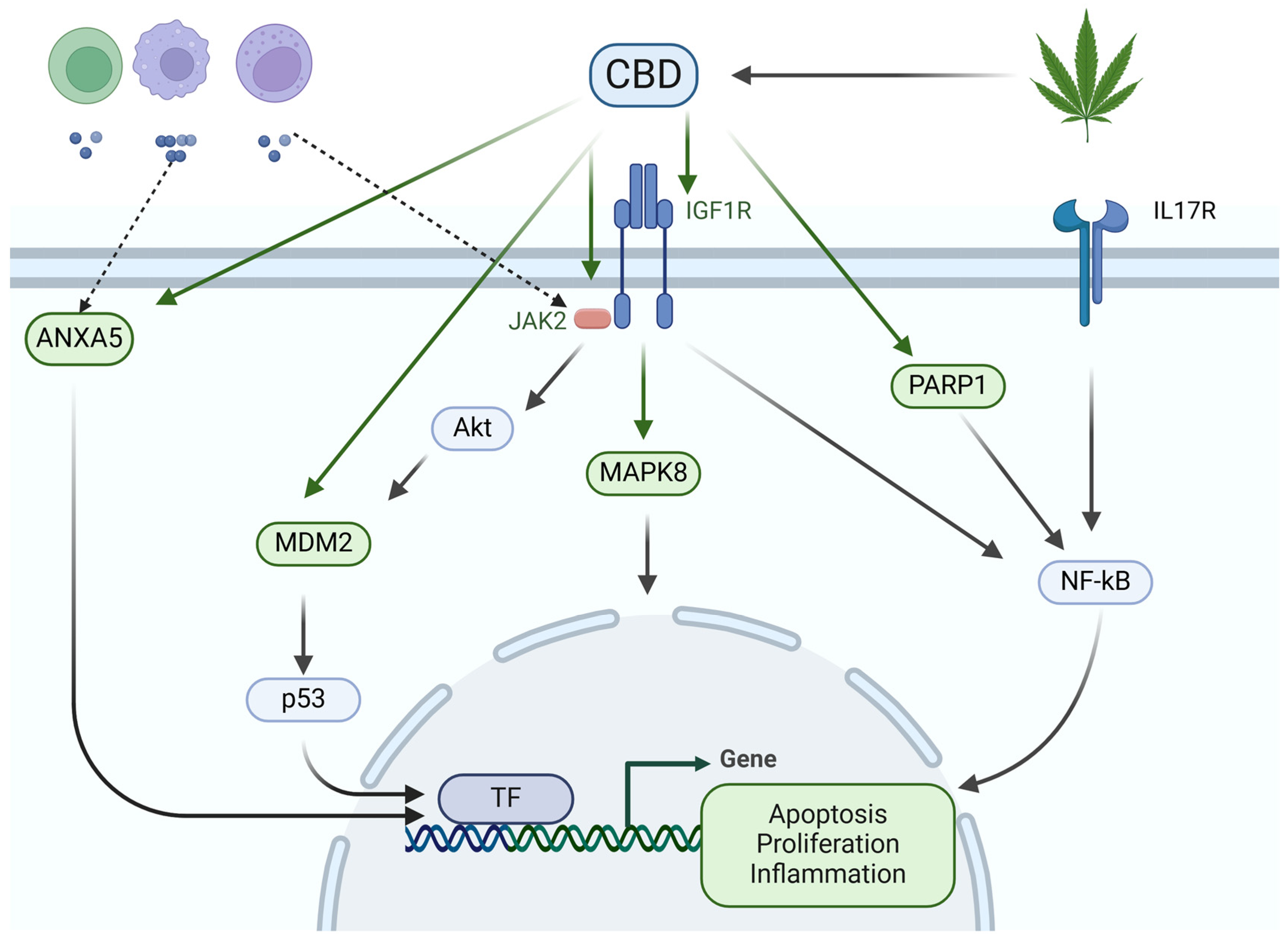
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, Z.; Wang, X.; Zhou, H.; Jiang, M.; Wang, J.; Sui, B. Molecular Complexity of Colorectal Cancer: Pathways, Biomarkers, and Therapeutic Strategies. Cancer Manag. Res. 2024, 16, 1389–1403. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef]
- Espinosa-Tamez, P.; Suazo-Zepeda, E.; Sanchez-Blas, H.; Meneses-Medina, M.; Huitzil-Melendez, F.D.; Van Loon, K.; Potter, M.; Lajous, M. National and state-level colorectal cancer mortality trends in Mexico, 1998–2018. Salud Publica Mex. 2021, 64, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, A.; Religioni, U.; Czerw, A.; Deptala, A.; Karakiewicz, B.; Partyka, O.; Pajewska, M.; Sygit, K.; Cipora, E.; Kmiec, K.; et al. Nutritional Treatment of Patients with Colorectal Cancer. Int. J. Environ. Res. Public Health 2022, 19, 6881. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Esquivias, F.; Gutierrez-Angulo, M.; Becerra-Ruiz, J.S.; Martinez-Perez, L.A.; de la Cruz-Ahumada, C.J.; Guzman-Flores, J.M. Bioinformatic Analysis of the Effect of Silver Nanoparticles on Colorectal Cancer Cell Line. Biomed. Res. Int. 2022, 2022, 6828837. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Esquivias, F.; Gutierrez-Angulo, M.; Perez-Larios, A.; Sanchez-Burgos, J.A.; Becerra-Ruiz, J.S.; Guzman-Flores, J.M. Anticancer Activity of Selenium Nanoparticles In Vitro Studies. Anticancer Agents Med. Chem. 2022, 22, 1658–1673. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Akash, S.; Rahman, M.M.; Nowrin, F.T.; Akter, T.; Shohag, S.; Rauf, A.; Aljohani, A.S.M.; Simal-Gandara, J. Colon cancer and colorectal cancer: Prevention and treatment by potential natural products. Chem. Biol. Interact. 2022, 368, 110170. [Google Scholar] [CrossRef] [PubMed]
- Silva-Reis, R.; Silva, A.M.S.; Oliveira, P.A.; Cardoso, S.M. Antitumor Effects of Cannabis sativa Bioactive Compounds on Colorectal Carcinogenesis. Biomolecules 2023, 13, 764. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Tamia, G.; Song, H.J.; Amarakoon, D.; Wei, C.I.; Lee, S.H. Cannabidiol exerts anti-proliferative activity via a cannabinoid receptor 2-dependent mechanism in human colorectal cancer cells. Int. Immunopharmacol. 2022, 108, 108865. [Google Scholar] [CrossRef]
- Kim, N.Y.; Mohan, C.D.; Sethi, G.; Ahn, K.S. Cannabidiol activates MAPK pathway to induce apoptosis, paraptosis, and autophagy in colorectal cancer cells. J. Cell Biochem. 2024, 125, e30537. [Google Scholar] [CrossRef]
- Orrego-Gonzalez, E.; Londono-Tobon, L.; Ardila-Gonzalez, J.; Polania-Tovar, D.; Valencia-Cardenas, A.; Velez-Van Meerbeke, A. Cannabinoid Effects on Experimental Colorectal Cancer Models Reduce Aberrant Crypt Foci (ACF) and Tumor Volume: A Systematic Review. Evid. Based Complement. Alternat. Med. 2020, 2020, 2371527. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Bashiri Dezfouli, A.; Multhoff, G. The immunomodulatory effects of cannabidiol on Hsp70-activated NK cells and tumor target cells. Mol. Immunol. 2024, 174, 1–10. [Google Scholar] [CrossRef]
- Nogales, C.; Mamdouh, Z.M.; List, M.; Kiel, C.; Casas, A.I.; Schmidt, H. Network pharmacology: Curing causal mechanisms instead of treating symptoms. Trends Pharmacol. Sci. 2022, 43, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Noor, F.; Tahir Ul Qamar, M.; Ashfaq, U.A.; Albutti, A.; Alwashmi, A.S.S.; Aljasir, M.A. Network Pharmacology Approach for Medicinal Plants: Review and Assessment. Pharmaceuticals 2022, 15, 572. [Google Scholar] [CrossRef] [PubMed]
- Agu, P.C.; Afiukwa, C.A.; Orji, O.U.; Ezeh, E.M.; Ofoke, I.H.; Ogbu, C.O.; Ugwuja, E.I.; Aja, P.M. Molecular docking as a tool for the discovery of molecular targets of nutraceuticals in diseases management. Sci. Rep. 2023, 13, 13398. [Google Scholar] [CrossRef]
- Paggi, J.M.; Pandit, A.; Dror, R.O. The Art and Science of Molecular Docking. Annu. Rev. Biochem. 2024, 93, 389–410. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New data content and improved web interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shen, Y.; Wang, S.; Li, S.; Zhang, W.; Liu, X.; Lai, L.; Pei, J.; Li, H. PharmMapper 2017 update: A web server for potential drug target identification with a comprehensive target pharmacophore database. Nucleic Acids Res. 2017, 45, W356–W360. [Google Scholar] [CrossRef] [PubMed]
- Rappaport, N.; Twik, M.; Plaschkes, I.; Nudel, R.; Iny Stein, T.; Levitt, J.; Gershoni, M.; Morrey, C.P.; Safran, M.; Lancet, D. MalaCards: An amalgamated human disease compendium with diverse clinical and genetic annotation and structured search. Nucleic Acids Res. 2017, 45, D877–D887. [Google Scholar] [CrossRef] [PubMed]
- Pinero, J.; Ramirez-Anguita, J.M.; Sauch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.P.; Wiegers, T.C.; Sciaky, D.; Barkalow, F.; Strong, M.; Wyatt, B.; Wiegers, J.; McMorran, R.; Abrar, S.; Mattingly, C.J. Comparative toxicogenomics database’s 20th anniversary: Update 2025. Nucleic Acids Res. 2024, 53, D1724. [Google Scholar] [CrossRef]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Chen, M.; Huang, X.; Zhang, G.; Zeng, L.; Zhang, G.; Wu, S.; Wang, Y. SRplot: A free online platform for data visualization and graphing. PLoS ONE 2023, 18, e0294236. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.H.; Chen, S.H.; Wu, H.H.; Ho, C.W.; Ko, M.T.; Lin, C.Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8 (Suppl. 4), S11. [Google Scholar] [CrossRef] [PubMed]
- Burley, S.K.; Bhatt, R.; Bhikadiya, C.; Bi, C.; Biester, A.; Biswas, P.; Bittrich, S.; Blaumann, S.; Brown, R.; Chao, H.; et al. Updated resources for exploring experimentally-determined PDB structures and Computed Structure Models at the RCSB Protein Data Bank. Nucleic Acids Res. 2025, 53, D564–D574. [Google Scholar] [CrossRef]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with PyRx. Methods Mol. Biol. 2015, 1263, 243–250. [Google Scholar] [CrossRef]
- Varadi, M.; Bertoni, D.; Magana, P.; Paramval, U.; Pidruchna, I.; Radhakrishnan, M.; Tsenkov, M.; Nair, S.; Mirdita, M.; Yeo, J.; et al. AlphaFold Protein Structure Database in 2024: Providing structure coverage for over 214 million protein sequences. Nucleic Acids Res. 2024, 52, D368–D375. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, X.; Gan, J.; Chen, S.; Xiao, Z.X.; Cao, Y. CB-Dock2: Improved protein-ligand blind docking by integrating cavity detection, docking and homologous template fitting. Nucleic Acids Res. 2022, 50, W159–W164. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Karthikeyan, S.K.; Korla, P.K.; Patel, H.; Shovon, A.R.; Athar, M.; Netto, G.J.; Qin, Z.S.; Kumar, S.; Manne, U.; et al. UALCAN: An update to the integrated cancer data analysis platform. Neoplasia 2022, 25, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Ru, B.; Wong, C.N.; Tong, Y.; Zhong, J.Y.; Zhong, S.S.W.; Wu, W.C.; Chu, K.C.; Wong, C.Y.; Lau, C.Y.; Chen, I.; et al. TISIDB: An integrated repository portal for tumor-immune system interactions. Bioinformatics 2019, 35, 4200–4202. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Madhukar Kudke, A.; Joseph Nepveux V, F.; Xu, Y. Network-Based Pharmacology Study Reveals Protein Targets for Medical Benefits and Harms of Cannabinoids in Humans. Appl. Sci. 2022, 12, 2205. [Google Scholar] [CrossRef]
- Bian, Y.M.; He, X.B.; Jing, Y.K.; Wang, L.R.; Wang, J.M.; Xie, X.Q. Computational systems pharmacology analysis of cannabidiol: A combination of chemogenomics-knowledgebase network analysis and integrated in silico modeling and simulation. Acta Pharmacol. Sin. 2019, 40, 374–386. [Google Scholar] [CrossRef]
- He, M.; Shi, J.; Wu, C.; Xu, Y.J.; Liu, Y. Integrating Lipidomics, Metabolomics, and Network Pharmacology to Reveal the Mechanism of Cannabidiol against Inflammation in High-Fat, High-Cholesterol Diet-Induced Mice. J. Agric. Food Chem. 2024, 72, 19246–19256. [Google Scholar] [CrossRef]
- Ma, H.; Xu, F.; Liu, C.; Seeram, N.P. A Network Pharmacology Approach to Identify Potential Molecular Targets for Cannabidiol’s Anti-Inflammatory Activity. Cannabis Cannabinoid Res. 2021, 6, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Lin, Y.; Lin, C.; Xu, M.; Wang, J.; Zeng, Z.; Wang, P.; Li, Q.; Wang, X.; Wang, W.; et al. Cannabidiol Inhibits Epithelial Ovarian Cancer: Role of Gut Microbiome. J. Nat. Prod. 2024, 87, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Yun, H.K.; Jeong, Y.A.; Jo, M.J.; Kang, S.H.; Kim, J.L.; Kim, D.Y.; Park, S.H.; Kim, B.R.; Na, Y.J.; et al. Cannabidiol-induced apoptosis is mediated by activation of Noxa in human colorectal cancer cells. Cancer Lett. 2019, 447, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.L.; Kim, B.R.; Kim, D.Y.; Jeong, Y.A.; Jeong, S.; Na, Y.J.; Park, S.H.; Yun, H.K.; Jo, M.J.; Kim, B.G.; et al. Cannabidiol Enhances the Therapeutic Effects of TRAIL by Upregulating DR5 in Colorectal Cancer. Cancers 2019, 11, 642. [Google Scholar] [CrossRef] [PubMed]
- du Plessis, J.; Deroubaix, A.; Omar, A.; Penny, C. A Bioinformatic Analysis Predicts That Cannabidiol Could Function as a Potential Inhibitor of the MAPK Pathway in Colorectal Cancer. Curr. Issues Mol. Biol. 2024, 46, 8600–8610. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Li, X.; Nie, S.; Liu, J.; Wang, S. Disorders of cancer metabolism: The therapeutic potential of cannabinoids. Biomed. Pharmacother. 2023, 157, 113993. [Google Scholar] [CrossRef]
- Razi, S.; Baradaran Noveiry, B.; Keshavarz-Fathi, M.; Rezaei, N. IL-17 and colorectal cancer: From carcinogenesis to treatment. Cytokine 2019, 116, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Harvey, B.S.; Sia, T.C.; Wattchow, D.A.; Smid, S.D. Interleukin 17A evoked mucosal damage is attenuated by cannabidiol and anandamide in a human colonic explant model. Cytokine 2014, 65, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.; Hao, L.Q.; Ding, F.X.; Mei, Q.; Huang, J.J.; Fu, C.G.; Yan, H.L.; Sun, S.H. Expression of annexin a5 is associated with higher tumor stage and poor prognosis in colorectal adenocarcinomas. J. Clin. Gastroenterol. 2009, 43, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Shali, H.; Ahmadi, M.; Kafil, H.S.; Dorosti, A.; Yousefi, M. IGF1R and c-met as therapeutic targets for colorectal cancer. Biomed. Pharmacother. 2016, 82, 528–536. [Google Scholar] [CrossRef] [PubMed]
- D’Aloia, A.; Ceriani, M.; Tisi, R.; Stucchi, S.; Sacco, E.; Costa, B. Cannabidiol Antiproliferative Effect in Triple-Negative Breast Cancer MDA-MB-231 Cells Is Modulated by Its Physical State and by IGF-1. Int. J. Mol. Sci. 2022, 23, 7145. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Lang, X.; Li, X. The role of IL-6/JAK2/STAT3 signaling pathway in cancers. Front. Oncol. 2022, 12, 1023177. [Google Scholar] [CrossRef]
- Kim, M.S.; Lee, J.H.; Kim, S.W.; Bang, C.H. Cannabidiol Alleviates Imiquimod-Induced Psoriasis by Inhibiting JAK2-STAT3 in a Mouse Model. Biomedicines 2024, 12, 2084. [Google Scholar] [CrossRef]
- Alibakhshi, A.; Malekzadeh, R.; Hosseini, S.A.; Yaghoobi, H. Investigation of the therapeutic role of native plant compounds against colorectal cancer based on system biology and virtual screening. Sci. Rep. 2023, 13, 11451. [Google Scholar] [CrossRef]
- Shi, H.; Tian, S.; Tian, H. Network Pharmacology Interpretation of Fuzheng-Jiedu Decoction against Colorectal Cancer. Evid. Based Complement. Alternat. Med. 2021, 2021, 4652492. [Google Scholar] [CrossRef]
- Guzman-Flores, J.M.; Perez-Vazquez, V.; Martinez-Esquivias, F.; Isiordia-Espinoza, M.A.; Viveros-Paredes, J.M. Molecular Docking Integrated with Network Pharmacology Explores the Therapeutic Mechanism of Cannabis sativa against Type 2 Diabetes. Curr. Issues Mol. Biol. 2023, 45, 7228–7241. [Google Scholar] [CrossRef] [PubMed]
- Elshazli, R.M.; Toraih, E.A.; Elgaml, A.; Kandil, E.; Fawzy, M.S. Genetic polymorphisms of TP53 (rs1042522) and MDM2 (rs2279744) and colorectal cancer risk: An updated meta-analysis based on 59 case-control studies. Gene 2020, 734, 144391. [Google Scholar] [CrossRef] [PubMed]
- Libro, R.; Scionti, D.; Diomede, F.; Marchisio, M.; Grassi, G.; Pollastro, F.; Piattelli, A.; Bramanti, P.; Mazzon, E.; Trubiani, O. Cannabidiol Modulates the Immunophenotype and Inhibits the Activation of the Inflammasome in Human Gingival Mesenchymal Stem Cells. Front. Physiol. 2016, 7, 559. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Dezfouli, A.B.; Khosravi, M.; Sievert, W.; Stangl, S.; Schwab, M.; Wu, Z.; Steiger, K.; Ma, H.; Multhoff, G. Cannabidiol-induced crosstalk of apoptosis and macroautophagy in colorectal cancer cells involves p53 and Hsp70. Cell Death Discov. 2023, 9, 286. [Google Scholar] [CrossRef]
- Wang, W.; Liu, D.; Yao, J.; Yuan, Z.; Yan, L.; Cao, B. ANXA5: A Key Regulator of Immune Cell Infiltration in Hepatocellular Carcinoma. Med. Sci. Monit. 2024, 30, e943523. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Chen, H.; Zhou, Z.; Guo, J. ANXA5 predicts prognosis and immune response and mediates proliferation and migration in head and neck squamous cell carcinoma. Gene 2024, 931, 148867. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.M.; Kaplan, B.L.F. Immune Responses Regulated by Cannabidiol. Cannabis Cannabinoid Res. 2020, 5, 12–31. [Google Scholar] [CrossRef] [PubMed]
- Peyravian, N.; Deo, S.; Daunert, S.; Jimenez, J.J. Cannabidiol as a Novel Therapeutic for Immune Modulation. Immunotargets Ther. 2020, 9, 131–140. [Google Scholar] [CrossRef]
- O’Sullivan, S.E.; Jensen, S.S.; Kolli, A.R.; Nikolajsen, G.N.; Bruun, H.Z.; Hoeng, J. Strategies to Improve Cannabidiol Bioavailability and Drug Delivery. Pharmaceuticals 2024, 17, 244. [Google Scholar] [CrossRef]
- Mozaffari, K.; Willette, S.; Lucker, B.F.; Kovar, S.E.; Holguin, F.O.; Guzman, I. The Effects of Food on Cannabidiol Bioaccessibility. Molecules 2021, 26, 3573. [Google Scholar] [CrossRef] [PubMed]
- Abbotts, K.S.S.; Ewell, T.R.; Butterklee, H.M.; Bomar, M.C.; Akagi, N.; Dooley, G.P.; Bell, C. Cannabidiol and Cannabidiol Metabolites: Pharmacokinetics, Interaction with Food, and Influence on Liver Function. Nutrients 2022, 14, 2152. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | AutodockVina RCSB | AutodockVina AlphaFold | CB-Dock2 AlphaFold | CB-Dock2 RSCB |
|---|---|---|---|---|
| ANXA5 | −6.6 | −7.3 | −7.1 | −7.0 |
| IGF1R | −7.3 | −6.4 | −6.8 | −7.6 |
| JAK2 | −7.3 | −6.9 | −7.8 | −7.6 |
| MAPK8 | −6.7 | −7.7 | −7.6 | −7.2 |
| MDM2 | −7.2 | −6.7 | −6.3 | −8.2 |
| PARP1 | −8.0 | −7.2 | −8.1 | −8.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Published by MDPI on behalf of the Österreichische Pharmazeutische Gesellschaft. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guzmán-Flores, J.M.; Martínez-Esquivias, F.; Alviz-Amador, A.; Avilés-Rodríguez, G.T.; García-Azuela, M.F. Exploring Cannabidiol’s Therapeutic Role in Colorectal Cancer: Network Pharmacology and Molecular Docking Insights. Sci. Pharm. 2025, 93, 12. https://doi.org/10.3390/scipharm93010012
Guzmán-Flores JM, Martínez-Esquivias F, Alviz-Amador A, Avilés-Rodríguez GT, García-Azuela MF. Exploring Cannabidiol’s Therapeutic Role in Colorectal Cancer: Network Pharmacology and Molecular Docking Insights. Scientia Pharmaceutica. 2025; 93(1):12. https://doi.org/10.3390/scipharm93010012
Chicago/Turabian StyleGuzmán-Flores, Juan Manuel, Fernando Martínez-Esquivias, Antistio Alviz-Amador, Guadalupe Thonanzyn Avilés-Rodríguez, and Michel Fabricio García-Azuela. 2025. "Exploring Cannabidiol’s Therapeutic Role in Colorectal Cancer: Network Pharmacology and Molecular Docking Insights" Scientia Pharmaceutica 93, no. 1: 12. https://doi.org/10.3390/scipharm93010012
APA StyleGuzmán-Flores, J. M., Martínez-Esquivias, F., Alviz-Amador, A., Avilés-Rodríguez, G. T., & García-Azuela, M. F. (2025). Exploring Cannabidiol’s Therapeutic Role in Colorectal Cancer: Network Pharmacology and Molecular Docking Insights. Scientia Pharmaceutica, 93(1), 12. https://doi.org/10.3390/scipharm93010012