
A Narrative Review on the Bioactivity and Health Benefits of Alpha-Phellandrene


 ,
,  ,
, 
Abstract
1. Introduction
2. Properties of Phellandrene
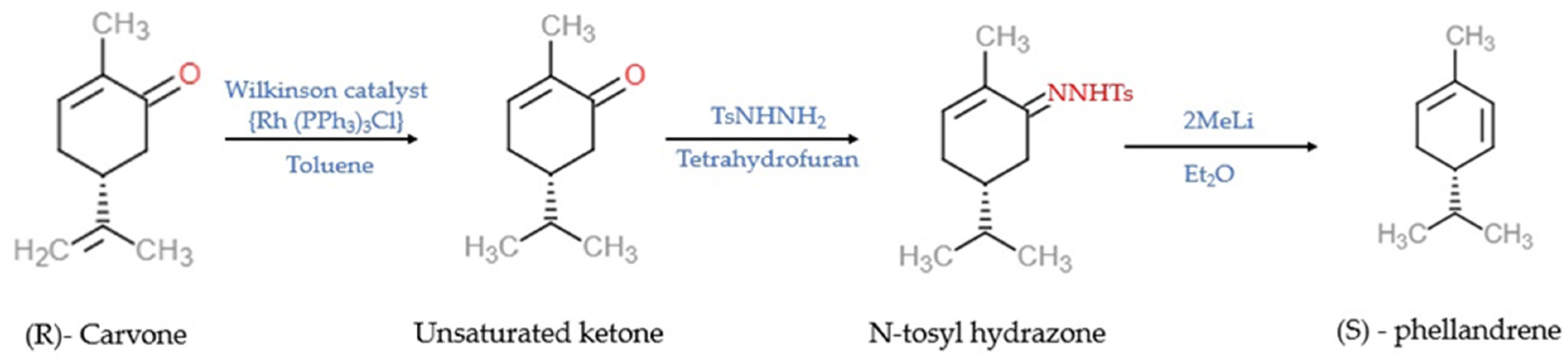
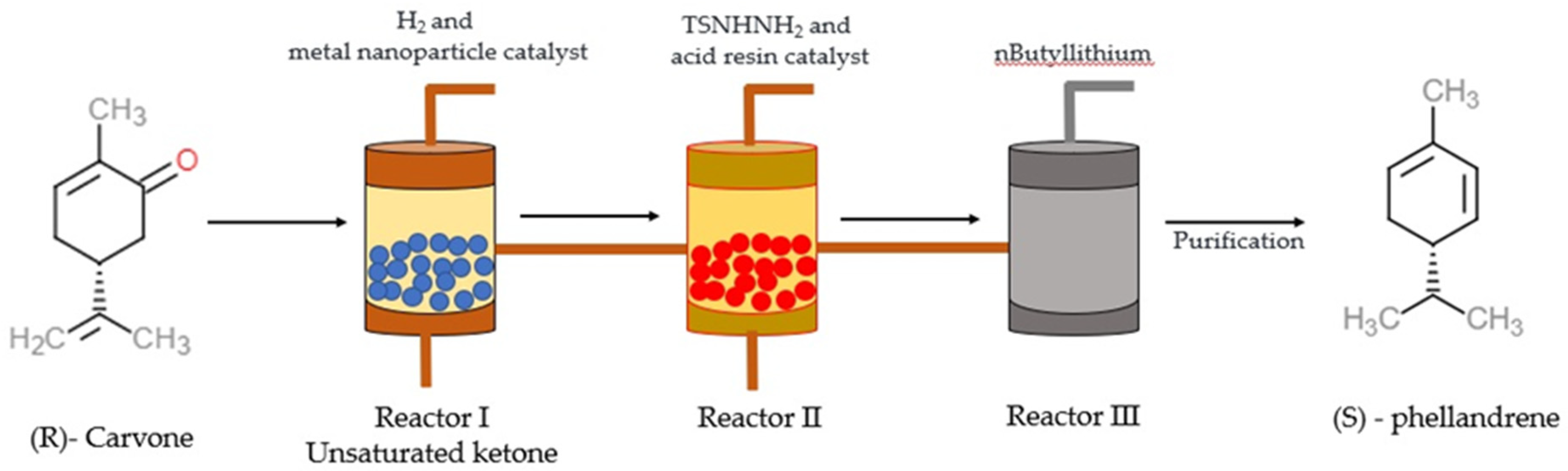
3. Chemistry and Synthesis of α-PHE
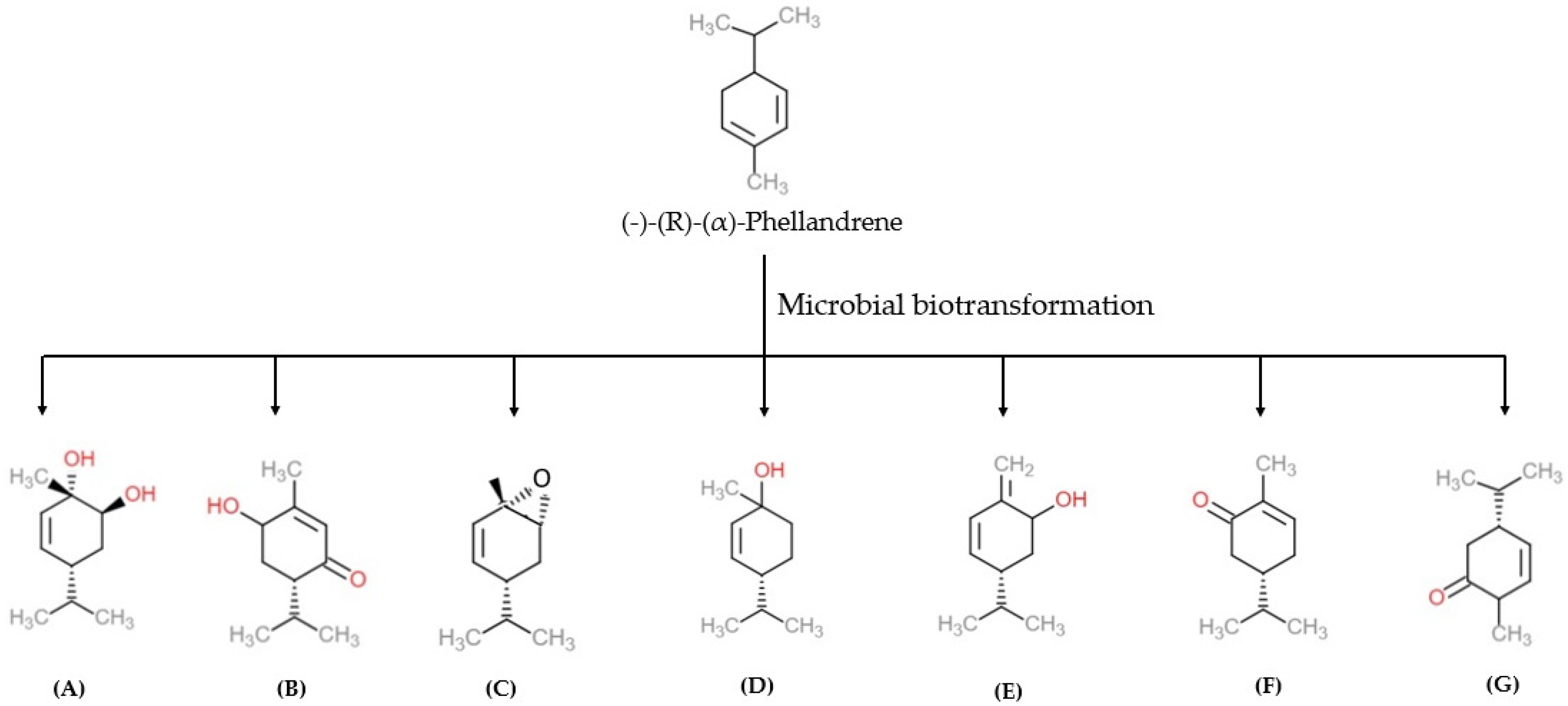
4. Biotransformation of α-PHE
5. α-PHE in Atmospheric Emission
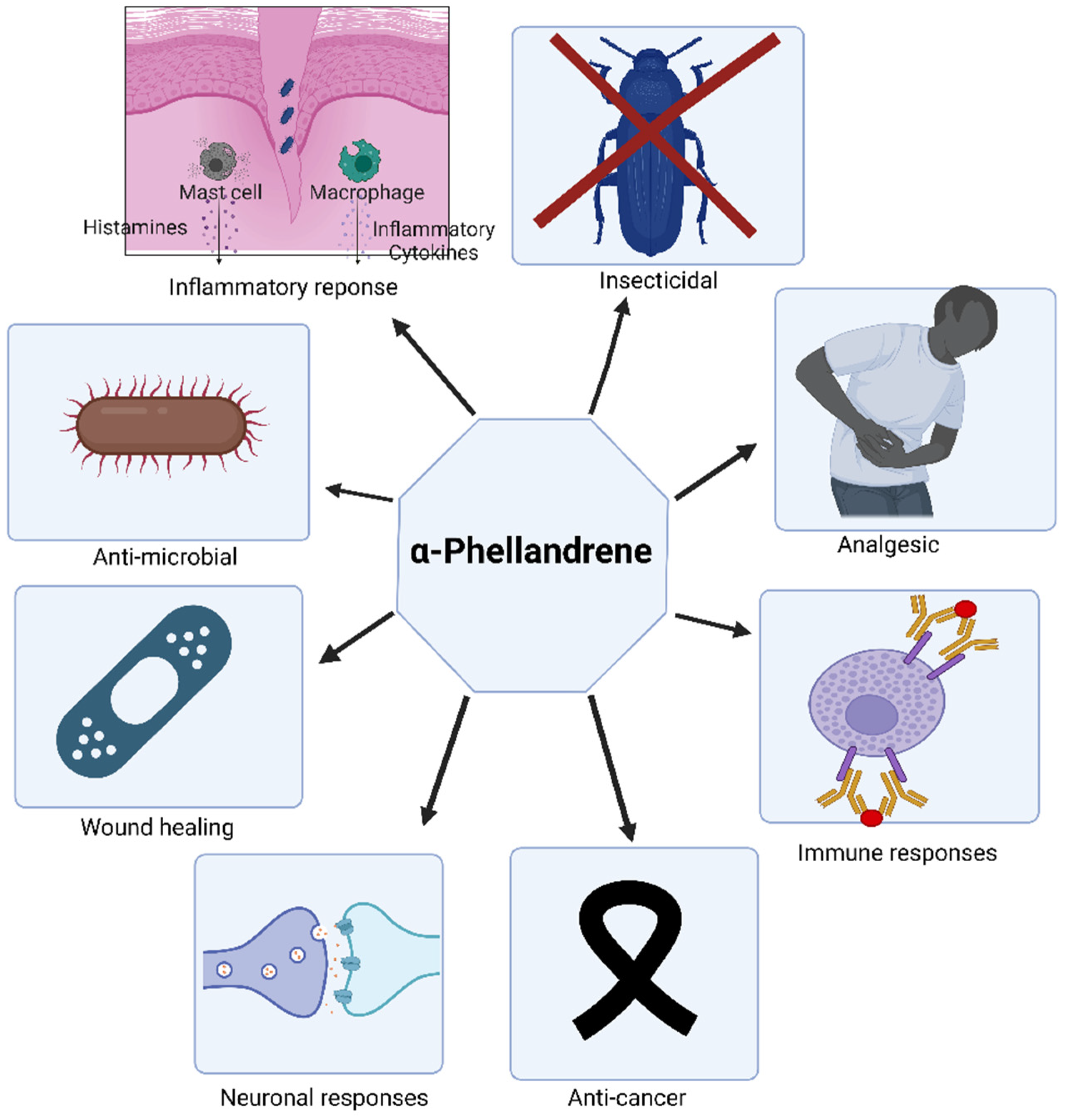
6. Bioactivities of α-PHE
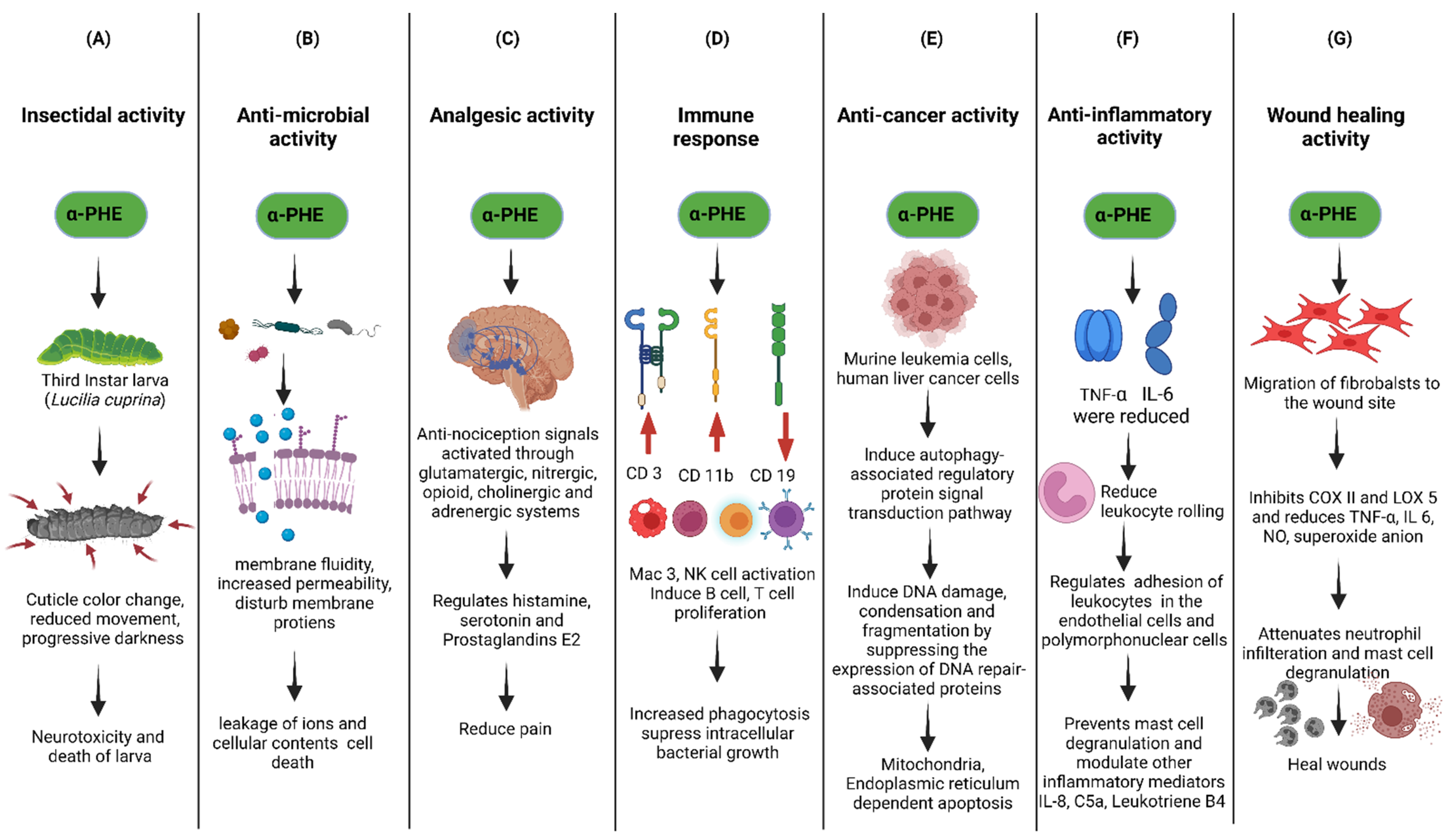
6.1. Insecticidal Property of α-PHE
6.2. Anti-Microbial Properties of α-PHE
6.3. Analgesic Property of α-PHE
6.4. Immune Responses of α-PHE
6.5. Anti-Cancer Property of α-PHE
6.6. Anti-Inflammatory Properties of α-PHE
6.7. Wound-Healing Activities of α-PHE
6.8. Neuronal Properties of Terpenes
7. Possible Mechanism of Action of α-PHE
8. Conclusions and Future Prospectus
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baser, K.H.C. Analysis, and Assessment of Essential Oils. In A Manual on the Essential Oil Industry; De Silva, K.T., Ed.; United Nations Industrial Development Organization: Vienna, Austria, 1995; pp. 155–178. [Google Scholar]
- Guan, X.; Ge, D.; Li, S.; Huang, K.; Liu, J.; Li, F. Chemical composition and antimicrobial activities of Artemisia argyi Levl. et Vant essential oils extracted by simultaneous distillation-extraction, subcritical extraction and hydrodistillation. Molecules 2019, 24, 483. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Park, J. A density functional theory study on the reaction mechanism of α-Phellandrene with NO3. Bull. Korean Chem. Soc. 2015, 36, 2139–2142. [Google Scholar] [CrossRef]
- Bizzo, R.H.; Hovell, A.M.C.; Rezende, C.M. Brazilian essential oil: General view, developments, and perspectives. Quim. Nova 2009, 3, 588–594. [Google Scholar] [CrossRef]
- İşcan, G.; Kirimer, N.; Demirci, F.; Demirci, B.; Noma, Y.; Başer, K.H. Biotransformation of (−)-(R)-α-phellandrene: Antimicrobial activity of its major metabolite. Chem. Biodivers. 2012, 9, 1525–1532. [Google Scholar] [CrossRef]
- Gaich, T.; Mulzer, J. Chiral Pool Synthesis: Starting from Terpenes. In Comprehensive Chirality, 1st ed.; Carreira, E.M., Yamamoto, H., Eds.; Elsevier: Amsterdam, The Netherland, 2012; Volume 2, pp. 163–206. [Google Scholar]
- New Directions Aromatics. Available online: https://www.newdirectionsaromatics.com/tags/phellandrene/essential-oils/ (accessed on 13 June 2022).
- Lee, S.H.; Young, L.H.; Benson, D.R.; Suni, T.; Kulmala, M.; Junninen, H.; Campos, T.L.; Rogers, D.C.; Jensen, J. Observations of nighttime new particle formation in the troposphere. J. Geophys. Res. 2008, 113, D10210. [Google Scholar] [CrossRef]
- Suni, T.; Kulmala, M.; Hirsikko, A.; Bergman, T.; Laakso, L.; Aalto, P.P.; Leuning, R.; Cleugh, H.; Zegelin, S.; Hughes, D.; et al. Formation and characteristics of ions and charged aerosol particles in a native Australian Eucalypt Forest. Atmos. Chem. Phys. 2008, 8, 129–139. [Google Scholar] [CrossRef]
- Clarke, S. Essential Chemistry for Aromatherapy, 2nd ed.; Churchill Livingstone: London, UK, 2009; pp. 123–229. [Google Scholar]
- Mackenzie-Rae, F.A.; Liu, T.; Deng, W.; Saunders, S.M.; Fang, Z.; Zhang, Y.; Wang, X. Ozonolysis of α-phellandrene—Part 1: Gas- and particle-phase characterisation. Atmos. Chem. Phys 2017, 17, 6583–6609. [Google Scholar] [CrossRef]
- Valsalan Soba, S.; Babu, M.; Panonnummal, R. Ethosomal gel formulation of alpha phellandrene for the transdermal delivery in gout. Adv. Pharm. Bull. 2021, 11, 137–149. [Google Scholar] [CrossRef]
- Trulieve. Available online: https://www.trulieve.com/discover/blog/everything-you-need-to-know-about-a-phellandrene-and-its-benefits (accessed on 20 July 2022).
- Zhang, J.H.; Sun, H.L.; Chen, S.Y.; Wang, T.T. Anti-fungal activity, mechanism studies on α-Phellandrene and nonanal against Penicillium cyclopium. Bot. Stud. 2017, 58, 13. [Google Scholar] [CrossRef]
- Piccinelli, A.C.; Santos, J.A.; Konkiewitz, E.C.; Oesterreich, S.A.; Formagio, A.S.; Croda, J.H.; Ziff, E.B.; Kassuya, C.A. Antihyperalgesic and antidepressive actions of (R)-(+)-limonene, α-phellandrene, and essential oil from Schinus terebinthifolius fruits in a neuropathic pain model. Nutr. Neurosci. 2015, 18, 217–224. [Google Scholar] [CrossRef]
- Lima, D.F.; Brandão, M.S.; Moura, J.B.; Leitão, J.M.; Carvalho, F.A.; Miúra, L.M.; Leite, J.R.; Sousa, D.P.; Almeida, F.R. Antinociceptive activity of the monoterpene α-phellandrene in rodents: Possible mechanisms of action. J. Pharm. Pharmacol. 2012, 64, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.L.; Li, Y.C.; Chang, W.C.; Chung, J.G.; Hsieh, L.C.; Wu, C.C. Induction of necrosis in human liver tumor cells by α-phellandrene. Nutr. Cancer 2014, 66, 970–979. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.J.; Hsu, S.C.; Lu, K.W.; Ma, Y.S.; Wu, C.C.; Lu, H.F.; Chen, J.C.; Lin, J.G.; Wu, P.P.; Chung, J.G. Alpha-phellandrene-induced apoptosis in mice leukemia WEHI-3 cells in vitro. Environ. Toxicol. 2016, 31, 1640–1651. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.O.; Wightman, E.L. Herbal Extracts and Phytochemicals: Plant Secondary Metabolites and the Enhancement of Human Brain function. Adv. Nutr. 2011, 2, 32–50. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Singh, O.P.; Maurya, S. Chemical and biocidal investigations on essential oils of some Indian Curcuma species. Prog. Cryst. Growth Charact. Mater. 2002, 45, 75–81. [Google Scholar] [CrossRef]
- Stashenko, E.E.; Jaramillo, B.E.; Martínez, J.R. Analysis of volatile secondary metabolites from Colombian Xylopia aromatica (Lamarck) by different extraction and headspace methods and gas chromatography. J. Chromatogr. A 2004, 1025, 105–113. [Google Scholar] [CrossRef]
- Napoli, E.M.; Curcuruto, G.; Ruberto, G. Screening of the essential oil composition of wild Sicilian rosemary. Biochem. Syst. Ecol. 2010, 38, 659–670. [Google Scholar] [CrossRef]
- Gilles, M.; Zhao, J.; An, M.; Agboola, S. Chemical composition and antimicrobial properties of essential oil of three Australian Eucalyptus species. Food Chem. 2010, 119, 731–737. [Google Scholar] [CrossRef]
- Nascimento, A.F.; Camara, C.A.; Moraes, M.M.; Ramos, C.S. Essential oil composition and acaricidal activity of Schinus terebinthifolius from Atlantic Forest of Pernambuco, Brazil against Tetranychusurticae. Nat. Prod. Commun. 2012, 7, 129–132. [Google Scholar]
- Bendaoud, H.; Romdhane, M.; Souchard, J.P.; Cazaux, S.; Bouajila, J. Chemical composition and anticancer and antioxidant activities of Schinus molle L. and Schinus terebinthifolius Raddi berries essential oils. J. Food Sci. 2010, 75, C466–C472. [Google Scholar] [CrossRef]
- Essien, E.E.; Ogunwande, I.A.; Setzer, W.N.; Ekundayo, O. Chemical composition, antimicrobial, and cytotoxicity studies on S. erianthum and S. macranthum essential oils. Pharm. Biol. 2012, 50, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Asbaghian, S.; Shafaghat, A.; Zarea, K.; Kasimov, F.; Salimi, F. Comparison of volatile constituents, and antioxidant and antibacterial activities of the essential oils of Thymus caucasicus, T. kotschyanus and T. vulgaris. Nat. Prod. Commun. 2011, 6, 137–140. [Google Scholar] [CrossRef]
- Arjouni, M.Y.; Bahri, F.; Romane, A.; El Fels, M.A. Chemical composition and antimicrobial activity of essential oil of Cupressus atlantica. Nat. Prod. Commun. 2011, 6, 1519–1522. [Google Scholar] [CrossRef]
- Hajhashemi, V.; Abbasi, N. Hypolipidemic activity of Anethum graveolens in rats. Phytother. Res. 2008, 22, 372–375. [Google Scholar] [CrossRef] [PubMed]
- Popovici, J.; Bertrand, C.; Bagnarol, E.; Fernandez, M.P.; Comte, G. Chemical composition of essential oil and headspace-solid micro extracts from fruits of Myrica gale L. and antifungal activity. Nat. Prod. Res. 2008, 22, 1024–1032. [Google Scholar] [CrossRef]
- Brandt, J.J.; Wulf, S. Composition of the Essential Oils of Ligusticum mutellina (L.) Crantz (Apiaceae). J. Essen. Oil. Res. 1995, 7, 231–235. [Google Scholar] [CrossRef]
- Thakuri, B.C.; Padalia, R.C.; Tiwari, A.; Chanotiya, C.S.; Mathela, C.S. α-Phellandrene rich essential oil from Ligusticum marginatum C.B. Clarke. J. Essent. Oil Bear. Plants 2007, 10, 1–4. [Google Scholar] [CrossRef]
- Cha, J.D.; Jung, E.K.; Kil, B.S.; Lee, K.Y. Chemical composition, and antibacterial activity of essential oil from Artemisia feddei. J. Microbiol. Biotechnol. 2007, 17, 2061–2065. [Google Scholar]
- Salas-Oropeza, J.; Jimenez-Estrada, M.; Perez-Torres, A.; Castell-Rodriguez, A.E.; Becerril-Millan, R.; Rodriguez-Monroy, M.A.; Jarquin-Yañez, K.; Canales-Martinez, M.M. Wound healing activity of α-Pinene and α-Phellandrene. Molecules 2021, 26, 2488. [Google Scholar] [CrossRef]
- Koudou, J.; Etou, A.W.; Ossibi, K.; Abena, A.A.; Gbeassor, M.; Bessiere, J.M. Chemical composition and hypotensive effects of essential oil Monodora myristica Gaertn. J. Biol. Sci. 2007, 7, 937–942. [Google Scholar] [CrossRef]
- Onyenekwe, P.C.; Stahl, M.; Adejo, G. Post-irradiation changes of the volatile oil constituents of Monodora myristica (Gaertn) Dunal. Nat. Prod. Res. 2012, 26, 2030–2034. [Google Scholar] [CrossRef]
- Jirovetz, L.; Buchbauer, G.; Ngassoum, M.B.; Geissler, M. Aroma compound analysis of Piper nigrum and Piper guineense essential oils from Cameroon using solid-phase microextraction-gas chromatography, solid-phase microextraction-gas chromatography-mass spectrometry and olfactometry. J. Chromatogr. A 2002, 976, 265–275. [Google Scholar] [CrossRef]
- POWO Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. 2022. Available online: http://www.plantsoftheworldonline.org/ (accessed on 17 September 2022).
- Hausen, B.M.; Reichling, J.; Harkenthal, M. Degradation products of monoterpenes are the sensitizing agents in tea tree oil. Am. J. Contact Dermat. 1999, 10, 68–77. [Google Scholar] [CrossRef]
- Iwanami, Y.; Tateba, H.; Kodama, N.; Kishino, K. Changes of lemon flavor components in an aqueous solution during UV irradiation. J. Agric. Food Chem. 1997, 45, 463–466. [Google Scholar] [CrossRef]
- Marzec, K.M.; Reva, I.; Fausto, R.; Proniewicz, L.M. Comparative matrix isolation infrared spectroscopy study of 1,3- and 1,4-diene monoterpenes (α-phellandrene and γ-terpinene). J. Phys. Chem. A 2011, 115, 4342–4353. [Google Scholar] [CrossRef]
- Burgstahler, A.W.; Ziffer, H.; Weiss, U. The configurations of levopimaric acid and α-phellandrene; Interpretation of their rotatory dispersions. J. Am. Chem. Soc. 1961, 83, 4660–4661. [Google Scholar] [CrossRef]
- Horsman, G.A.; Emeis, C.A. Low-temperature optical rotatory dispersion. Tetrahedron 1965, 22, 167–173. [Google Scholar] [CrossRef]
- Takahashi, O.; Yamasaki, K.; Kohno, Y.; Ueda, K.; Suezawa, H.; Nishio, M. Origin of the axial-alkyl preference of (R)-α-phellandrene and related compounds investigated by high-level ab initio MO calculations. Importance of the CH/π hydrogen bond. Tetrahedron 2008, 64, 5773–5778. [Google Scholar] [CrossRef]
- Sevov, C.S.; Wiest, O. Selectivity in the electron transfer catalyzed Diels-Alder reaction of (R)-alpha-phellandrene and 4-methoxystyrene. J. Org. Chem. 2008, 73, 7909–7915. [Google Scholar] [CrossRef]
- Arruda, B.C.; Smith, B.; Spears, K.G.; Sension, R.J. Ultrafast ring-opening reactions: A comparison of alpha-terpinene, alpha-phellandrene, and 7-dehydrocholesterol with 1,3-cyclohexadiene. Faraday Discuss. 2013, 163, 159–275. [Google Scholar] [CrossRef]
- Reid, P.J.; Doig, S.J.; Wickham, S.D.; Mathies, R.A. Photochemical ring-opening reactions are complete in picoseconds: A time resolved UV resonance Raman study of 1,3 cyclohexadiene. J. Am. Chem. Soc. 1993, 115, 4754–4763. [Google Scholar] [CrossRef]
- Sen, A.; Grosch, W. The synthesis of enantiomerically pure (S)(+)-a-phellandrene from (R)(−)-carvone. Flavour Fragr. J. 1990, 5, 233–234. [Google Scholar] [CrossRef]
- Miller, S.J.; Ishitani, H.; Furiya, Y.; Shu, K. High-Throughput Synthesis of (S)-α-Phellandrene through Three-Step Sequential Continuous-Flow Reactions. Org. Process Res. Dev. 2021, 25, 192–198. [Google Scholar] [CrossRef]
- Straathof, A.J.; Panke, S.; Schmid, A. The production of fine chemicals by biotransformations. Curr. Opin. Biotechnol. 2002, 13, 548–556. [Google Scholar] [CrossRef]
- Abraham, W.R.; Stumpf, B.; Kieslich, K. Microbial Transformations of Terpenoids with 1-p-Menthene Skeleton. Appl. Microbiol. Biotechnol. 1986, 24, 24–30. [Google Scholar] [CrossRef]
- Lai-King, S.Y.; Saunders, R.M.K.; Brown, G.D. Phytochemistry of Illicium dunnianum and the systematic position of the illiciaceae. Phytochemistry 1997, 44, 1099–1108. [Google Scholar]
- Montiel, V.; Lopez-Segura, M.; Aldaz, A.; Grande, M.; Barba, F. Electrooxidation of terpenes-I. Synthesis of dihydrocarvone and 1-hydroxyneodihydrocarveol by anodic oxidation of limonene. Electrochim. Acta 1984, 29, 1123–1126. [Google Scholar] [CrossRef]
- Hoffmann, T.; Odum, J.R.; Bowman, F.; Collins, D.; Klockow, D.; Flagan, R.C.; Seinfeld, J.H. Formation of organic aerosols from the oxidation of biogenic hydrocarbons. J. Atmos. Chem. 1997, 26, 189–222. [Google Scholar] [CrossRef]
- Griffin, R.J.; Cocker, D.R.; Flagan, R.C.; Seinfeld, J.H. Organic aerosol formation from the oxidation of biogenic hydrocarbons. J. Geophys. Res. 1999, 104, 3555–3567. [Google Scholar] [CrossRef]
- Schurgers, G.; Arneth, A.; Holzinger, R.; Goldstein, A.H. Process-based modelling of biogenic monoterpene emissions combining production and release from storage. Atmos. Chem. Phys. 2009, 9, 3409–3423. [Google Scholar] [CrossRef]
- Mackenzie-Rae, F.A.; Wallis, H.J.; Rickard, A.R.; Pereira, K.L.; Saunders, S.M.; Wang, X.; Hamilton, J.F. Ozonolysis of α-phellandrene—Part 2: Compositional analysis of secondary organic aerosol highlights the role of stabilised Criegee intermediates. Atmos. Chem. Phys. 2017, 18, 4673–4693. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Chaaban, A.; Richardi, V.S.; Carrer, A.R.; Brum, J.S.; Cipriano, R.R.; Martins, C.; Navarro-Silva, M.A.; Deschamps, C.; Molento, M.B. Cuticular damage of Lucilia cuprina larvae exposed to Curcuma longa leaves essential oil and its major compound α-phellandrene. Data Brief 2020, 21, 1776–1778. [Google Scholar] [CrossRef]
- Chaaban, A.; Richardi, V.S.; Carrer, A.R.; Brum, J.S.; Cipriano, R.R.; Martins, C.E.; Silva, M.A.; Deschamps, C.; Molento, M.B. Insecticide activity of Curcuma longa (leaves) essential oil and its major compound a-phellandrene against Lucilia cuprina larvae (Diptera: Calliphoridae): Histological and ultrastructural biomarkers assessment. Pestic. Biochem. Physiol. 2019, 153, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro-Neto, F.R.; Lopes, E.M.; Acha, B.T.; Gomes, L.; Dias, W.A.; Reis Filho, A.; Leal, B.S.; Rodrigues, D.; Silva, J.; Dittz, D.; et al. α-Phellandrene exhibits antinociceptive and tumor-reducing effects in a mouse model of oncologic pain. Toxicol. Appl. Pharmacol. 2021, 418, 115497. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.J.; Lin, J.H.; Hsu, S.C.; Weng, S.W.; Huang, Y.P.; Tang, N.Y.; Lin, J.G.; Chung, J.G. Alpha-phellandrene promotes immune responses in normal mice through enhancing macrophage phagocytosis and natural killer cell activities. Vivo 2013, 27, 809–814. [Google Scholar]
- Wu, C.C.; Lin, C.L.; Huang, C.Y.; Hsieh, S.; Liu, C.H.; Hsieh, S.L. α-Phellandrene enhances the immune response and resistance against Vibrio alginolyticus in white shrimp (Litopenaeus vannamei). Fish Shellfish Immunol. 2019, 84, 1108–1114. [Google Scholar] [CrossRef]
- Hsieh, L.C.; Hsieh, S.L.; Chen, C.T.; Chung, J.G.; Wang, J.J.; Wu, C.C. Induction of α-phellandrene on autophagy in human liver tumor cells. Am. J. Chin. Med. 2015, 43, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.J.; Wu, C.C.; Hsu, S.C.; Weng, S.W.; Ma, Y.S.; Huang, Y.P.; Lin, J.G.; Chung, J.G. Alpha-phellandrene-induced DNA damage and affect DNA repair protein expression in WEHI-3 murine leukemia cells in vitro. Environ. Toxicol. 2015, 30, 1322–1330. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.J.; Lu, K.W.; Ma, Y.S.; Tang, N.Y.; Wu, P.P.; Wu, C.C.; Lu, H.F.; Lin, J.G.; Chung, J.G. Alpha-phellandrene, a natural active monoterpene, influences a murine WEHI-3 leukemia model in vivo by enhancing macrophage phagocytosis and natural killer cell activity. Vivo 2014, 28, 583–588. [Google Scholar]
- Lin, J.J.; Yu, C.C.; Lu, K.W.; Chang, S.J.; Yu, F.S.; Liao, C.L.; Lin, J.G.; Chung, J.G. α-Phellandrene alters expression of genes associated with DNA damage, cell cycle, and apoptosis in murine leukemia WEHI-3 cells. Anticancer Res. 2014, 34, 4161–4180. [Google Scholar] [PubMed]
- Siqueira, H.D.S.; Neto, B.S.; Sousa, D.P.; Gomes, B.S.; da Silva, F.V.; Cunha, F.V.M.; Wanderley, C.; Pinheiro, G.; Cândido, A.; Wong, D.; et al. α-Phellandrene, a cyclic monoterpene, attenuates inflammatory response through neutrophil migration inhibition and mast cell degranulation. Life Sci. 2016, 160, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, H.D.S.; Neto, B.S.; Sousa, D.P.; Gomes, B.S.; Cunha, F.V.M.; Oliveira, F. Antiedematogenic effect of alpha-phellandrene in animal models. Curr. Bioact. Compd. 2016, 12, 107–113. [Google Scholar] [CrossRef]
- Gonçalves, R.; Cunha, F.; Sousa-Neto, B.; Oliveira, L.; Lopes, M.E.; Rezende, D.C.; Sousa, I.; Nogueira, K.M.; Souza, L.; Medeiros, J.; et al. α-Phellandrene attenuates tissular damage, oxidative stress, and TNF-α levels on acute model ifosfamide-induced hemorrhagic cystitis in mice. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 1835–1848. [Google Scholar] [CrossRef]
- De Christo Scherer, M.M.; Marques, F.M.; Figueira, M.M.; Peisino, M.; Schmitt, E.; Kondratyuk, T.P.; Endringer, D.C.; Scherer, R.; Fronza, M. Wound healing activity of terpinolene and α-phellandrene by attenuating inflammation and oxidative stress in vitro. J. Tissue. Viability 2019, 28, 94–99. [Google Scholar] [CrossRef]
- Al-Burtamani, S.K.S.; Fatope, M.O.; Marwah, R.G.; Onifade, A.K.; Al-Saidi, S.H. Chemical composition, antibacterial and antifungal activities of the essential oil of Haplophyllum tuberculatum from Oman. J. Ethnopharmacol. 2005, 96, 107–112. [Google Scholar] [CrossRef]
- Inouye, S.; Takizawa, T.; Yamaguchi, H. Antibacterial activity of essential oils and their major constituents against respiratory tract pathogens by gaseous contact. J. Antimicrob. Chemother. 2001, 47, 565–573. [Google Scholar] [CrossRef]
- Hernández, V.; Mora, F.; Araque, M.; Montijo, S.D.; Rojas, L.; Meléndez, P.; Tommasi, N.D. Chemical composition and antibacterial activity of Astronium graveolens JACQ essential oil. Rev. Latinoamer. Quím. 2013, 41, 89–94. [Google Scholar]
- Nurettin, Y.; Canan, G.; Osman, Ü.; Ahmet, Y.; Serdar, Ü.; Kamil, C.; Salih, T. Composition and antimicrobial activities of volatile components of Minuartia meyeri. Turk. J. Chem. 2006, 30, 71–76. [Google Scholar]
- Sharma, P.; Shah, G.C.; Dhami, D.S.; Chauhan, P.K.; Singh, V. Chemical composition, antibacterial and antioxidant activities of senecio laetus edgew. From cold desert of western Himalaya. Int. J. Pharm. Res. Bio. Sci. 2014, 3, 188–199. [Google Scholar]
- Costa, T.R.; Fernandes, O.F.L.; Santos, S.C.; Oliveira, C.M.A.; Liao, L.M.; Ferri, P.H.; Paula, J.R.; Ferreira, H.D.; Sales, B.H.N.; Silva, M.R.R. Antifungal activity of volatile constituents of Eugenia dysenterica leaf oil. J. Ethnopharmacol. 2000, 72, 111–117. [Google Scholar] [CrossRef]
- Perez, C.; Agnese, A.M.; Cabrera, J.L. The essential oil of Scenecio graveolans (compositae)-chemical composition and antimicrobial activity tests. J. Ethnopharmacol. 1999, 66, 91–96. [Google Scholar] [CrossRef]
- Kurade, N.P.; Jaitak, V.; Kaul, V.K.; Sharma, O.P. Chemical composition and antibacterial activity of essential oils of Lantana camara, Ageratum houstonianum and Eupatorium adenophorum. Pharm. Biol. 2010, 48, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, S.; Chempakam, B.; Leela, N.K.; Suseela Bhai, R. Chemoprevention by essential oil of turmeric leaves (Curcuma longa L.) on the growth of Aspergillus flavus and aflatoxin production. Food Chem. Toxicol. 2011, 49, 1188–1192. [Google Scholar] [CrossRef]
- Smith-Palmer, A.; Stewart, J.; Fyfe, L. Antimicrobial properties of plant essential oils and essences against five important food-borne pathogens. Lett. Appl. Microbiol. 1998, 26, 118–122. [Google Scholar] [CrossRef]
- Tassou, C.C.; Nychas, G.J.E. Antimicrobial activity of the essential oil of Mastic gum (Pistacia lentiscus var chia) on gram-positive and gram-negative bacteria in broth and model food systems. Int. Biodeterior. Biodegrad. 1995, 36, 411–420. [Google Scholar] [CrossRef]
- Mann, C.M.; Cox, S.D.; Markham, J.L. The outer membrane of Pseudomonas aeruginosa NCTC6749 contributes to its tolerance to the essential oil of Melaleuca alternifolia (tea tree oil). Lett. Appl. Microbiol. 2000, 30, 294–297. [Google Scholar] [CrossRef]
- Oonmetta-aree, J.; Suzuki, T.; Gasaluck, P.; Eumkeb, G. Antimicrobial properties and action of galangal (Alpinia galangal Linn.) on Staphylococcus aureus. LWT-Food Sci. Technol. 2006, 39, 1214–1220. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods-A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Fadli, M.; Saad, A.; Sayadi, S.; Chevalier, J.; Mezriouia, N.E.; Pagèsc, J.M.; Hassani, L. Antibacterial activity of Thymus maroccanus and Thymus broussonetii essential oils against nosocomial infection-bacteria and their synergistic potential with antibiotics. Phytomedicine 2012, 19, 464–471. [Google Scholar] [CrossRef]
- Khan, A.; Ahmad, A.; Akhtar, F.; Yousuf, S.; Xess, I.; Khan, L.A.; Manzoor, N. Ocimum sanctum essential oil and its active principles exert their antifungal activity by disrupting ergosterol biosynthesis and membrane integrity. Res. Microbiol. 2010, 161, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, A.K.; Malik, A. Antimicrobial potential and chemical composition of Mentha piperita oil in liquid and vapour phase against food spoiling microorganisms. Food Control 2011, 22, 1707–1714. [Google Scholar] [CrossRef]
- Yahyazadeh, M.; Omidbaigi, R.; Zare, R.; Taheri, H. Effect of some essential oils on mycelial growth of Penicillium digitatum Sacc. World J. Microbiol. Biotechnol. 2008, 24, 1445–1450. [Google Scholar] [CrossRef]
- Silva, L.J.; Mendanha, D.M.; Gomes, P.P. The use of opioids in the treatment of oncologic pain in the elderly. Braz. J. Pain 2020, 3, 63–72. [Google Scholar] [CrossRef]
- Gouveia, D.N.; Costa, J.S.; Oliveira, M.A.; Rabelo, T.K.; Silva, A.M.O.; Carvalho, A.A.; Miguel-Dos-Santos, R.; Lauton-Santos, S.; Scotti, L.; Scotti, M.T.; et al. α-Terpineol reduces cancer pain via modulation of oxidative stress and inhibition of iNOS. Biomed. Pharmacother. 2018, 105, 652–661. [Google Scholar] [CrossRef]
- Sylvestre, M.; Legault, J.; Dufour, D.; Pichette, A. Chemical composition and anticancer activity of leaf essential oil of Myrica gale L. Phytomedicine 2005, 12, 299–304. [Google Scholar] [CrossRef]
- Cobos Jimenez, V.; Bradley, E.J.; Willemsen, A.M.; van Kampen, A.H.; Baas, F.; Kootstra, N.A. Next-generation sequencing of microRNAs uncovers expression signatures in polarized macrophages. Physiol. Genom. 2014, 46, 91–103. [Google Scholar] [CrossRef]
- Adib-Conquy, M.; Scott-Algara, D.; Cavaillon, J.M.; Souza Fonseca-Guimaraes, F. TLR-mediated activation of NK cells and their role in bacterial/viral immune responses in mammals. Immunol. Cell Biol. 2014, 92, 256–262. [Google Scholar] [CrossRef]
- Wright, H.L.; Moots, R.J.; Bucknall, R.C.; Edwards, S.W. Neutrophil function in inflammation and inflammatory diseases. Rheumatology 2010, 49, 1618–1631. [Google Scholar] [CrossRef]
- Waldleigh, D.J.; Reddy, S.T.; Kopp, E.; Ghosh, S.; Herschman, H.R. Transcriptional activation of the cyclooxygenase-2 gene in endotoxin-treated RAW 264.7 macrophages. J. Biol. Chem. 2000, 275, 6259–6266. [Google Scholar] [CrossRef]
- Romano, M.; Faggioni, R.; Sironi, M.; Sacco, S.; Echtenacher, B.; Di Santo, E.; Salmona, M.; Ghezzi, P. Carrageenan-induced acute inflammation in the mouse air pouch synovival model. Role of tumour necrosis factor. Mediat. Inflamm. 1997, 6, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Leahy, K.M.; Koki, A.T.; Masferrer, J.L. Role of cyclooxygenases in angiogenesis. Curr. Med. Chem. 2000, 7, 1163–1170. [Google Scholar] [CrossRef] [PubMed]
- Damas, J.; Bourdon, V.; Remacle-Volon, G.; Adam, A. Kinins and peritoneal exudates induced by carrageenin and zymosan in rats. Br. J. Pharmacol. 1990, 101, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.A.; Souza-Filho, M.V.; Souza, M.H.; Oliveira, S.H.; Costa, C.H.; Cunha, F.Q.; Ferreira, H.S. Role of resident mast cells and macrophages in the neutrophil migration induced by LTB4, fMLP and C5a des arg. Int. Arch. Allergy Immunol. 1997, 112, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.H.; Canetti, C.; Ribeiro, R.A.; Cunha, F.Q. Neutrophil migration induced by IL1beta depends upon LTB4 released by macrophages and upon TNF-alpha and IL1beta released by mast cells. Inflammation 2008, 31, 36–46. [Google Scholar] [CrossRef]
- Perretti, M. Endogenous mediators that inhibit the leukocyte–endothelium interaction. Trends Pharmacol. Sci. 1997, 18, 418–425. [Google Scholar]
- Carvalho, A.M.R.; Rocha, N.F.M.; Vasconcelos, L.F.; RiosERV, D.L.M.; Silva, M.I.G.; Fonteles, M.M.F.; Barbosa Filho, J.M.; Gutierrez, S.J.C.; Sousa, F.C.F. Evaluation of the anti-inflammatory activity of riparin II (O-methil-N-2-hidroxi-benzoyl tyramine) in animal models. Chem. Biol. Interact. 2013, 205, 165–172. [Google Scholar] [CrossRef]
- Sencer, S.F.; Haake, R.J.; Weisdorf, D.J. Hemorrhagic cystitis after bone marrow transplantation. Transplantation 1993, 56, 875–879. [Google Scholar] [CrossRef]
- Dobrek, Ł.; Thor, P.J. Bladder urotoxicity pathophysiology induced by the oxazaphosphorine alkylating agents and its chemoprevention. Postepy Hig. Med. Dosw. (Online) 2012, 66, 592–602. [Google Scholar] [CrossRef]
- Gurtner, G.; Werner, S.; Barrandon, Y.; Longaker, M. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef]
- Barrientos, S.; Stojadinovic, O.; Golinko, M.S.; Brem, H.; Tomic-Canic, M. Growth factors and cytokines in wound healing. Wound Repair Regen. 2008, 16, 585–601. [Google Scholar] [CrossRef] [PubMed]
- Sawatdee, S.; Choochuay, K.; Chanthorn, W.; Srichana, T. Evaluation of the topical spray containing Centella asiatica extract and its efficacy on excision wounds in rats. Acta Pharm. 2016, 66, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Tracy, L.E.; Minasian, R.A.; Caterson, E.J. Extracellular Matrix and Dermal Fibroblast Function in the Healing Wound. Adv. Wound Care 2016, 5, 119–136. [Google Scholar] [CrossRef]
- Salas-Oropeza, J.; Jimenez-Estrada, M.; Perez-Torres, A.; Castell-Rodriguez, A.E.; Becerril-Millan, R.; Rodriguez-Monroy, M.A.; Canales-Martinez, M.M. Wound healing activity of the essential oil of Bursera morelensis, in mice. Molecules 2020, 25, 1795. [Google Scholar] [CrossRef] [PubMed]
- Babawale, O.P.; Taiwo, F.; Adetunj, O.S. Ethnobotanical survey of plants used as memory enhancers in three states of southwestern Nigeria. J. Appl. Pharm. Sci. 2016, 6, 209–214. [Google Scholar] [CrossRef]
- Kennedy, D.; Okello, E.; Chazot, P.; Howes, M.J.; Ohiomokhare, S.; Jackson, P.; Haskell-Ramsay, C.; Khan, J.; Forster, J.; Wightman, E. Volatile Terpenes and Brain Function: Investigation of the Cognitive and Mood Effects of Mentha × Piperita L. Essential Oil with In Vitro Properties Relevant to Central Nervous System Function. Nutrients 2018, 10, 1029. [Google Scholar] [CrossRef]
- Mkaddem, M.; Bouajila, J.; Ennajar, M.; Lebrihi, A.; Mathieu, F.; Romdhane, M. Chemical composition and antimicrobial and antioxidant activities of Mentha (Longifolia L. and viridis) essential oils. J. Food Sci. 2009, 74, M358–M363. [Google Scholar] [CrossRef]
- Hong, Y.K.; Park, S.H.; Lee, S.; Hwang, S.; Lee, M.J.; Kim, D.; Lee, J.H.; Han, S.Y.; Kim, S.T.; Kim, Y.K.; et al. Neuroprotective effect of SuHeXiang Wan in Drosophila models of Alzheimer’s disease. J. Ethnopharmacol. 2011, 134, 1028–1032. [Google Scholar] [CrossRef]
- Gonçalves, J.C.; Oliveira, F.; Benedito, R.B.; de Sousa, D.P.; de Almeida, R.N.; de Araújo, D.A. Antinociceptive activity of (−)-carvone: Evidence of association with decreased peripheral nerve excitability. Biol. Pharm. Bull. 2008, 31, 1017–1020. [Google Scholar] [CrossRef]
- França, D.S.; Souza, A.L.; Almeida, K.R.; Dolabella, S.S.; Martinelli, C.; Coelho, M.M. B vitamins induce an antinociceptive effect in the acetic acid and formaldehyde models of nociception in mice. Eur. J. Pharmacol. 2001, 421, 157–164. [Google Scholar] [CrossRef]
- Millan, M.J. The induction of pain: An integrative review. Prog. Neurobiol. 1999, 57, 161–164. [Google Scholar]
- Beirith, A.; Santos, A.R.; Calixto, J.B. Mechanisms underlying the nociception and paw oedema caused by injection of glutamate into the mouse paw. Brain Res. 2002, 924, 219–228. [Google Scholar] [CrossRef]
- Oliveira, M.G.; Marques, R.B.; Santana, M.F.; Santos, A.B.; Brito, F.A.; Barreto, E.O.; Sousa, D.P.; Almeida, F.R.; Badauê-Passos, D.; Antoniolli, A.R.; et al. α-Terpineol reduces mechanical hypernociception and inflammatory response. Basic Clin. Pharmacol. Toxicol. 2012, 111, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Kawata, J.; Kameda, M.; Miyazawa, M. Cyclooxygenase-2 inhibitory effects of monoterpenoids with a p-methane skeleton. Int. J. Essent. Oil Ther. 2008, 2, 145–148. [Google Scholar]
- Yamamoto, S.; Ohsawa, M.; Ono, H. Contribution of TRPV1 receptor-expressing fibers to spinal ventral root after-discharges and mechanical hyperalgesia in a spared nerve injury (SNI) rat model. J. Pharmacol. Sci. 2013, 121, 9–16. [Google Scholar] [CrossRef]
- Dhaka, A.; Murray, A.N.; Mathur, J.; Earley, T.J.; Petrus, M.J.; Patapoutian, A. TRPM8 is required for cold sensation in mice. Neuron 2007, 54, 371–378. [Google Scholar] [CrossRef]
- Thiaucourt, M.; Shabes, P.; Schloss, N.; Sack, M.; Baumgartner, U.; Schmahl, C.; Ende, G. Posterior insular GABA levels inversely correlate with the intensity of experimental mechanical pain in healthy subjects. Neuroscience 2018, 387, 116–122. [Google Scholar] [CrossRef]
- Sigel, E.; Steinmann, M.E. Structure, function, and modulation of GABAA receptors. J. Biol. Chem. 2012, 287, 40224–40231. [Google Scholar] [CrossRef]
- Ren, K.; Torres, R. Role of interleukin-1beta during pain and inflammation. Brain Res. Rev. 2009, 60, 57–64. [Google Scholar] [CrossRef]
- Heal, C.F.; Banks, J.L.; Lepper, P.D.; Kontopantelis, E.; van Driel, M.L. Topical antibiotics for preventing surgical site infection in wounds healing by primary intention. Cochrane Database Syst. Rev. 2016, 11, Cd011426. [Google Scholar] [CrossRef]
- Nedelec, B.; Ghahary, A.; Scott, P.G.; Tredget, E.E. Control of wound contraction. Basic and clinical features. Hand Clin. 2000, 16, 289–302. [Google Scholar] [CrossRef]
- Held, S.; Schieberle, P.; Somoza, V. Characterization of alpha terpineol as an anti-inflammatory component of orange juice by in vitro studies using oral buccal cells. J. Agric. Food Chem. 2007, 55, 8040–8046. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Plant Species | α-Phellandrene Content (%) # | References |
|---|---|---|---|
| 1 | Curcuma zedoaria (Christm) | 14.93 | [20] |
| 2 | Xylopia aromatica L. | 2.2–6.4 | [21] |
| 3 | Rosmarinus officinalis L. | 0.1–0.4 | [22] |
| 4 | Eucalyptus dives | 17.4 | [23] |
| 5 | Eucalyptus staigeriana | 8.8 | [23] |
| 7 | Schinus terebinthifolius Raddi | 15.7 | [24] |
| 34.38 | [25] | ||
| 6 | Schinus molle L. | 46.52 | [25] |
| 8 | Solanum erianthum D. Don. | 17.5 | [26] |
| 9 | Thymus kotschyanus Boiss. and Hohen. | 10.8 | [27] |
| 10 | Cupressus atlantica Gaussen | 5.5 | [28] |
| 11 | Anethum graveolens L. | 32 | [29] |
| 12 | Myrica gale L. | 8 | [30] |
| 13 | Ligusticum mutellina L. Crantz | 23.4 | [31] |
| 14 | Ligusticum marginatum | 50.2 | [32] |
| 15 | Artemisia feddei | 5.78 | [33] |
| 16 | Bursera morelensis | 1 | [34] |
| 17 | Monodora myristica (Gaertn.) | 34.4 | [35] |
| Monodora myristica (Gaertn.) Dunal | 53 | [36] | |
| 18 | Piper nigrum | 8.56 | [37] |
| Study Model | Details of α-PHE and EO Used | Experimental Condition | Study Results | References |
|---|---|---|---|---|
| Insecticidal activity | ||||
| Third instar stage larvae (L3) of the Australian blowfly Lucilia cuprina | Commercial α-PHE and Curcuma longa leaves EO (CLLEO). | L3 larvae were kept on tissue paper infused with 0.15–2.86 mL/cm2 of CLLEO and 0.29–1.47 mL/cm2 of α-PHE dissolved in ethanol. Toxicity was observed at 6, 24 and 48 h. |
| [59] |
| Third instar stage larvae (L3) of the Australian blowfly Lucilia cuprina | Commercial α-PHE and Curcuma longa leaves EO (CLLEO). | L3 larvae were kept on tissue paper infused with 0.15–2.86 mL/cm2 of CLLEO and 0.29–1.47 mL/cm2 of α-PHE s dissolved in ethanol. Toxicity was observed at 6, 24 and 48 h. |
| [60] |
| Anti-microbial activity | ||||
| Penicillium cyclopium | Commercial α-PHE and Nonanal | 6mm diameter discs of P. cyclopium inoculum were taken out from culture and placed at the center of Petri dishes containing α-PHE (0, 0.25, 0.50, 0.75, 1.00, 1.25, 1.50, 1.75 and 2.00 mL/L) and Nonanal (0, 50, 100, 150, 200, 250, 300, 350 and 400 μL/L) |
| [14] |
| Swiss male mice and Wistar male rats. Macrophages were isolated from the peritoneal cavity of the mice. | Commercial α-PHE | Mice were treated with α-PHE and saline before (1 h) the treatment with 0.75% acetic acid, 2% formalin, capsaicin (2 mg/paw), glutamate (20 mmol/paw), 1% carrageenan. The anti-nociceptive effect of α-PHE and mechanical hypernociception were analyzed. |
| [16] |
| Spared Nerve Injury (SNI) induced hyperalgesia in Wistar rats. | Commercial (R)-(+)-phellandrene, α-limonene and Schinus terebinthifolius fruits EO | Commercial (R)-(+)-phellandrene (10 mg/kg) and α-limonene (10 mg/kg) and S. terebinthifolius EO (3, 30 and 100 mg/kg) were treated with SNI-induced hyperalgesia rats. |
| [15] |
| S-180 inoculated Swiss mice female. Melanoma B-16/F-10 and S-180 murine cells. | Commercial α-PHE | Animals were separated into seven groups as follows: Sham treated, vehicle-treated (pregabalin 10 mg/kg), positive control (25 mg/kg of 5-fluorouracil), and α-PHE in doses of 6.25, 12.5, 25 and 50 mg/kg orally by gavage once a day. |
| [61] |
| Immune responses | ||||
| BALB/c mice | Commercial α-PHE | BALB/c mice were divided into five groups; I: normal diet; II: olive oil vehicle as positive control; other groups III, IV and V: α-PHE at 1, 5 and 25 mg/kg dissolved in olive oil for 27 days. |
| [62] |
| Litopenaeus vannamei White shrimp | Commercial α-PHE | L. vannamei was administered with α-PHE (4,8 and 12 μg/g) and challenged against V. alginolyticus (2 × 105 CFU/shrimp). Non-pre-treated shrimps as controls. Shrimp survival was observed. The immune responses, gene expression and hemocytes morphology, were studied. |
| [63] |
| Anti-cancer activity | ||||
| Human liver cancer (J5) cells | Commercial α-PHE | J5 cells were treated with 0, 10, 30, or 50 mM of α-PHE for 0, 6, 12, 24, or 48 h. The cytotoxic effects and expressions of Bax, Bcl-2, PARP, and caspase-3 proteins were studied. The necrotic cells level, ROS, MMP, ATP and LDH levels in J5 cells were investigated. |
| [17] |
| Human liver cancer (J5) cells | Commercial α-PHE | J5 cells were treated with 0, 10, 30, or 50 mM of α-PHE. The expression levels of cytoplasmic proteins PI3K-I, PI3K-III, Akt, mTOR, phosphorylated Bcl-2 (p-Bcl-2), Beclin-1, LC3-II, p53, TIGAR, phosphorylated IκB (p-IκB), IκB, and NF-κB (p65) and nuclear p53, DARM, and NF-B were evaluated. |
| [64] |
| WEHI-3 murine leukemia cells | Commercial α-PHE | WEHI-3 cells were treated with α-PHE (0, 10, 20, 30, and 50 µM) for 48 h. DNA damage and condensation and DNA repair associated-proteins expression were analyzed. |
| [65] |
| Male BALB/c mice injected with WEHI-3 murine leukemia cells | Commercial α-PHE | WEHI-3 injected mice were treated with 25 and 50 mg/kg of α-PHE. After 2 weeks, immune responses and macrophage phagocytic and NK cell activities were studied. |
| [66] |
| WEHI-3 murine leukemia cell line | Commercial α-PHE | WEHI-3 cells were treated with 10 μM of α-PHE or vehicle for 48 h. The cells were analyzed for expression of DNA damage-related genes, cell cycle and apoptotic cell death. |
| [67] |
| WEHI-3 murine leukemia cell line | Commercial α-PHE | WEHI-3 cells were treated with 0, 5, 10, 30, 40, and 50 µM of α-PHE. The cytotoxicity effect of α-PHE was assessed. |
| [18] |
| Anti-inflammatory activity | ||||
| Wistar male rats and Swiss mice | Commercial α-PHE | Before carrageenan injection, rats or mice were treated with vehicle or α-PHE (50, 100, or 200 mg/kg) or dexamethasone (0.5 mg/kg). |
| [68] |
| Wistar male rats and Swiss mice | Commercial α-PHE | Before carrageenan injection, rats or mice were treated with vehicle or α-PHE (50, 100, or 200 mg/kg) or dexamethasone (0.5 mg/kg). |
| [69] |
| Male Mus musculus mice of Swiss albino lineage. | Commercial α-PHE | Mice were treated with tween (vehicle control), ifosfamide (IFOS; negative control), Mesna (positive control), and α-PHE. |
| [70] |
| Wound healing activities | ||||
| Fibroblasts (L929), macrophages and human embryonic renal cells (HEK 293) transfected with the luciferase-expressing gene | Commercial α-PHE and Terpinolene | For the cytotoxicity study: Fibroblasts and macrophages were treated with terpinolene and α-PHE. For wound healing study: Fibroblasts were exposed to 10, 100 and 200 μM of terpinolene and α-PHE. |
| [71] |
| Male CD-1 mice and fibroblasts | Commercial α-PHE and α-Pinene | The mice’s back skin was shaved, and an incision was made. Mice were divided into groups I: untreated skin without wounds, group II: untreated wounds (negative control), group III: wounds treated with Recoverón NC® (positive control), group IV: cosmetic grade mineral oil (vehicle), and the next four groups treated with α-pinene and α-PHE at 0.1 and 0.01 mg/mL concentrations for 10 days. |
| [34] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thangaleela, S.; Sivamaruthi, B.S.; Kesika, P.; Tiyajamorn, T.; Bharathi, M.; Chaiyasut, C. A Narrative Review on the Bioactivity and Health Benefits of Alpha-Phellandrene. Sci. Pharm. 2022, 90, 57. https://doi.org/10.3390/scipharm90040057
Thangaleela S, Sivamaruthi BS, Kesika P, Tiyajamorn T, Bharathi M, Chaiyasut C. A Narrative Review on the Bioactivity and Health Benefits of Alpha-Phellandrene. Scientia Pharmaceutica. 2022; 90(4):57. https://doi.org/10.3390/scipharm90040057
Chicago/Turabian StyleThangaleela, Subramanian, Bhagavathi Sundaram Sivamaruthi, Periyanaina Kesika, Tanawat Tiyajamorn, Muruganantham Bharathi, and Chaiyavat Chaiyasut. 2022. "A Narrative Review on the Bioactivity and Health Benefits of Alpha-Phellandrene" Scientia Pharmaceutica 90, no. 4: 57. https://doi.org/10.3390/scipharm90040057
APA StyleThangaleela, S., Sivamaruthi, B. S., Kesika, P., Tiyajamorn, T., Bharathi, M., & Chaiyasut, C. (2022). A Narrative Review on the Bioactivity and Health Benefits of Alpha-Phellandrene. Scientia Pharmaceutica, 90(4), 57. https://doi.org/10.3390/scipharm90040057