
Apoptosis Induction Associated with Enhanced ER Stress Response and Up-Regulation of c-Jun/p38 MAPK Proteins in Human Cervical Cancer Cells by Colocasia esculenta var. aquatilis Hassk Extract



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Plant Extraction and Identification
2.3. Cell Lines and Cell Culture
2.4. Cytotoxicity Assay
2.5. Detection of Nuclear Morphological Changes
2.6. Mitochondrial Membrane Potential (ΔΨm) Detection
2.7. Cell Cycle Analysis
2.8. Detection of Intracellular ROS
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
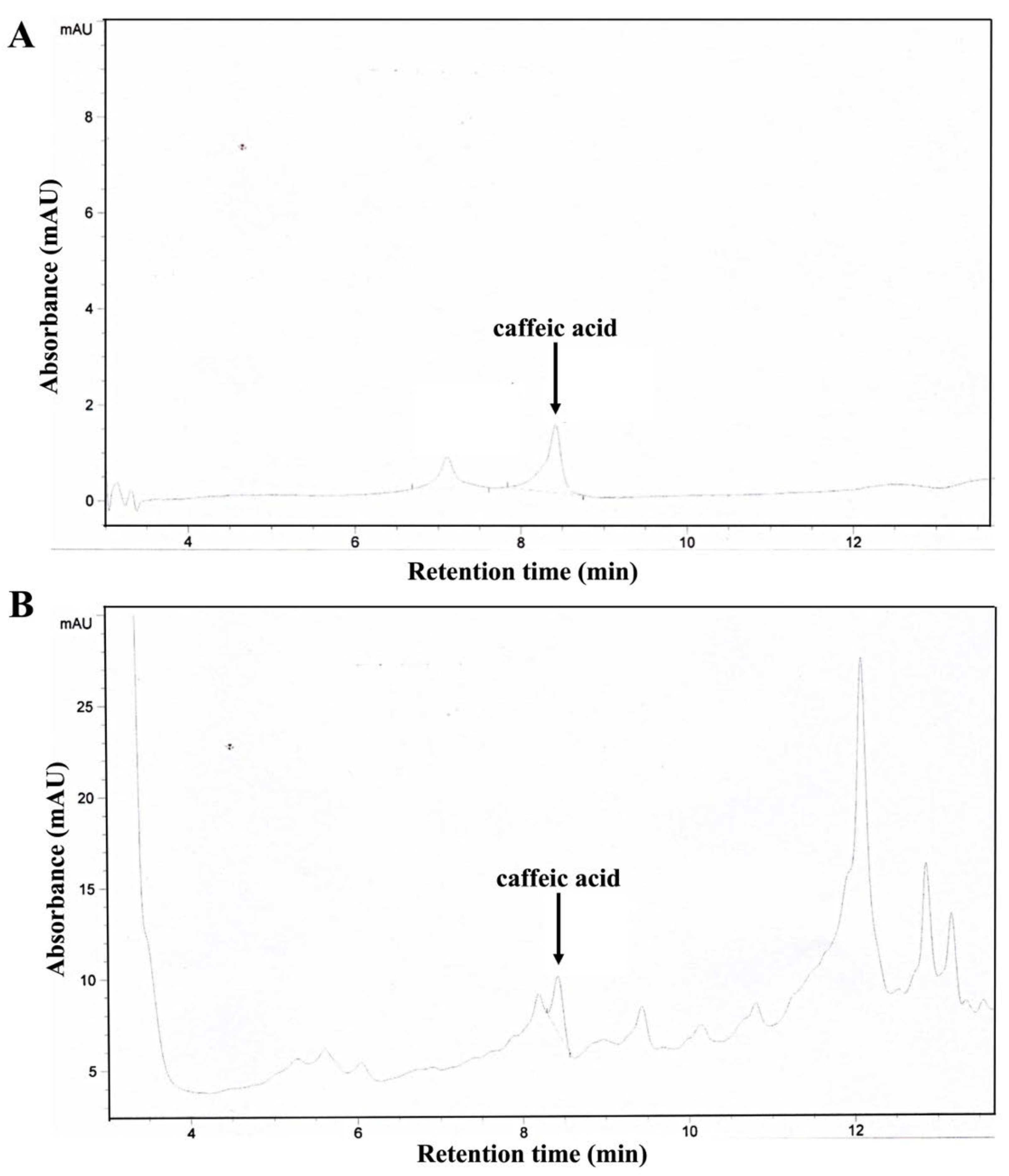
3.1. HPLC Analysis of Caffeic Acid
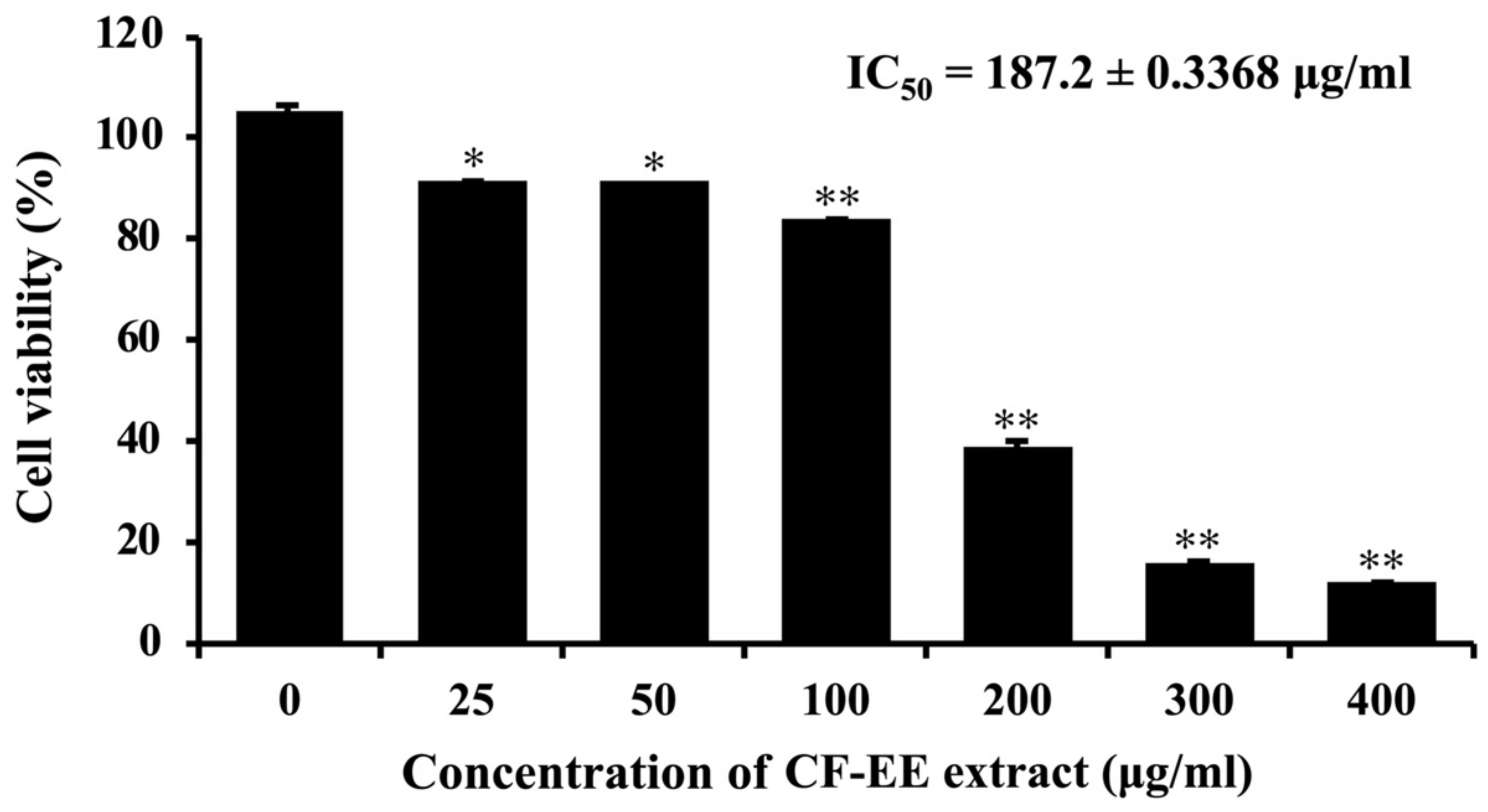
3.2. CF-EE Suppresses Cell Proliferation of HeLa Cells
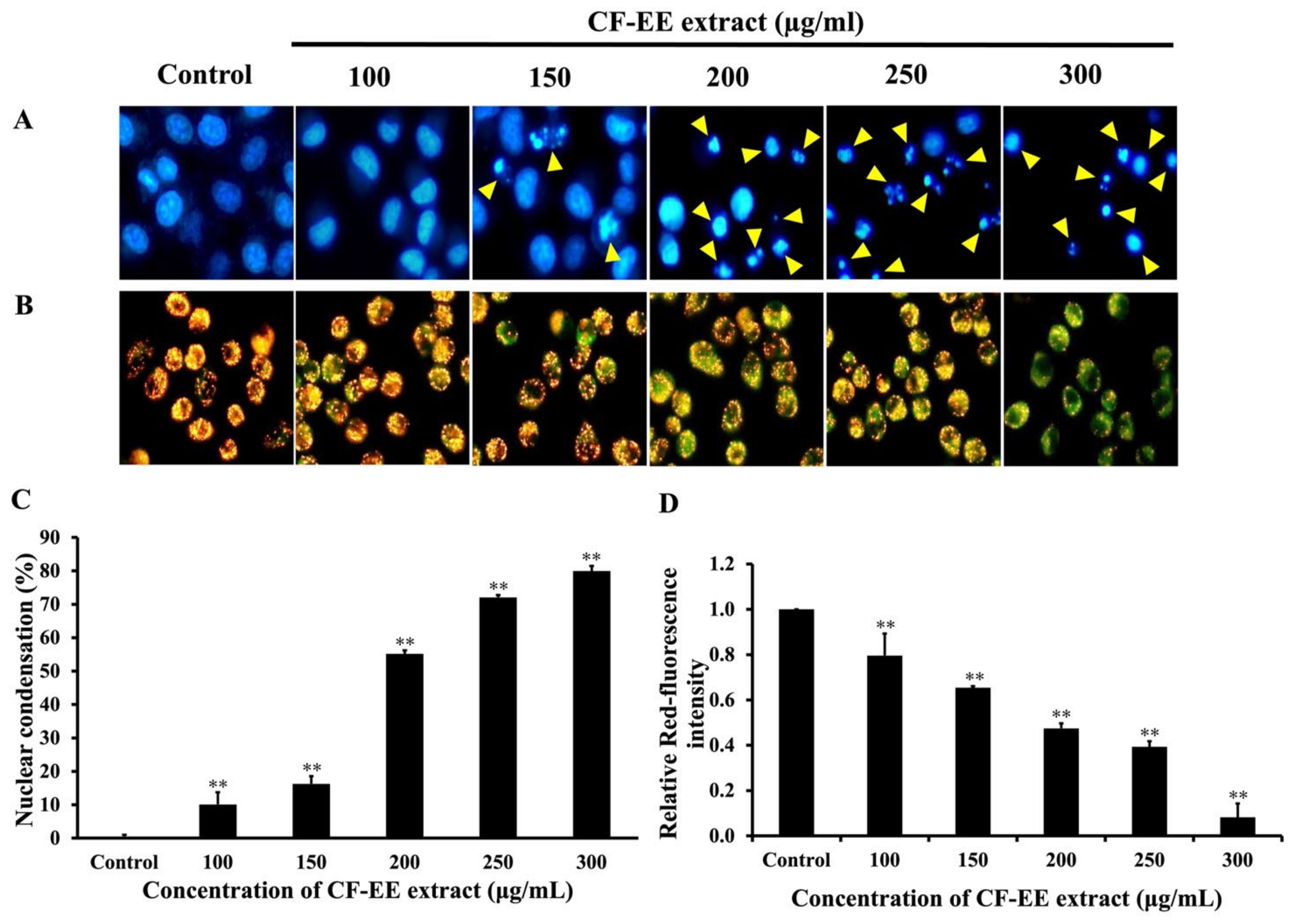
3.3. Effect of CF-EE Extract on Apoptosis Induction in HeLa Cells
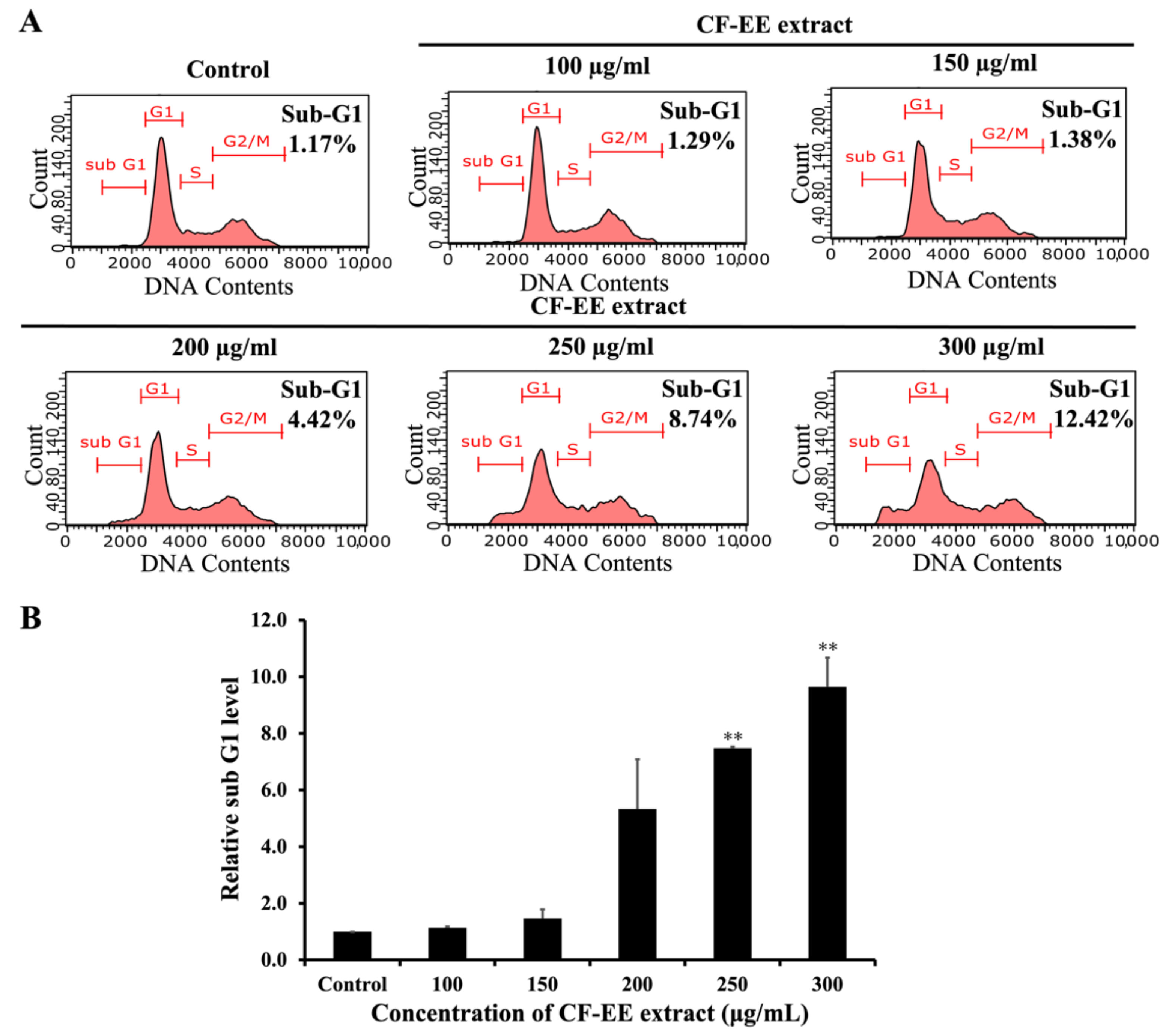
3.4. CF-EE Extract Induces Sub-G1 Population in HeLa Cells
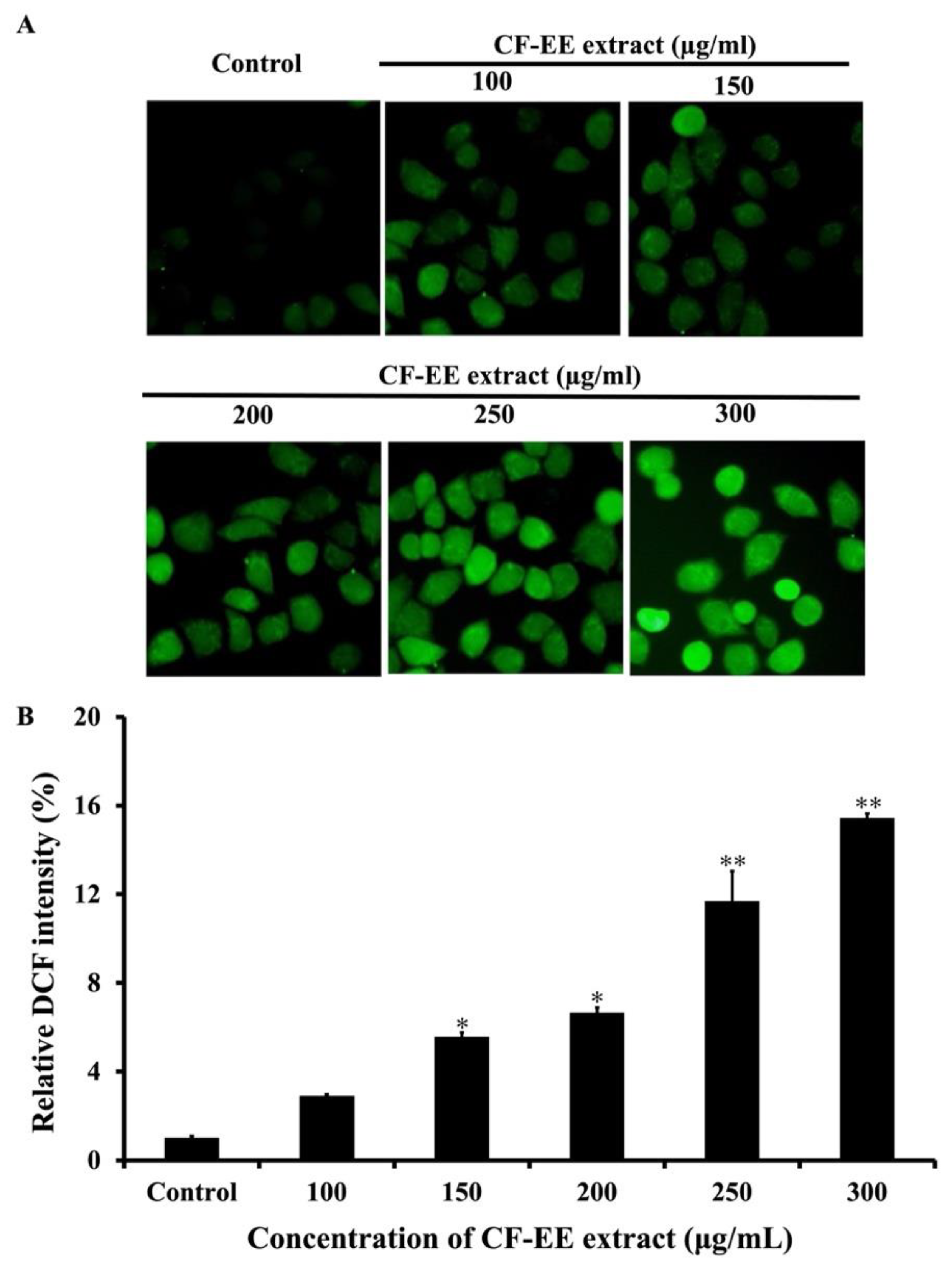
3.5. CF-EE Extract Induces Oxidative Stress in HeLa Cells
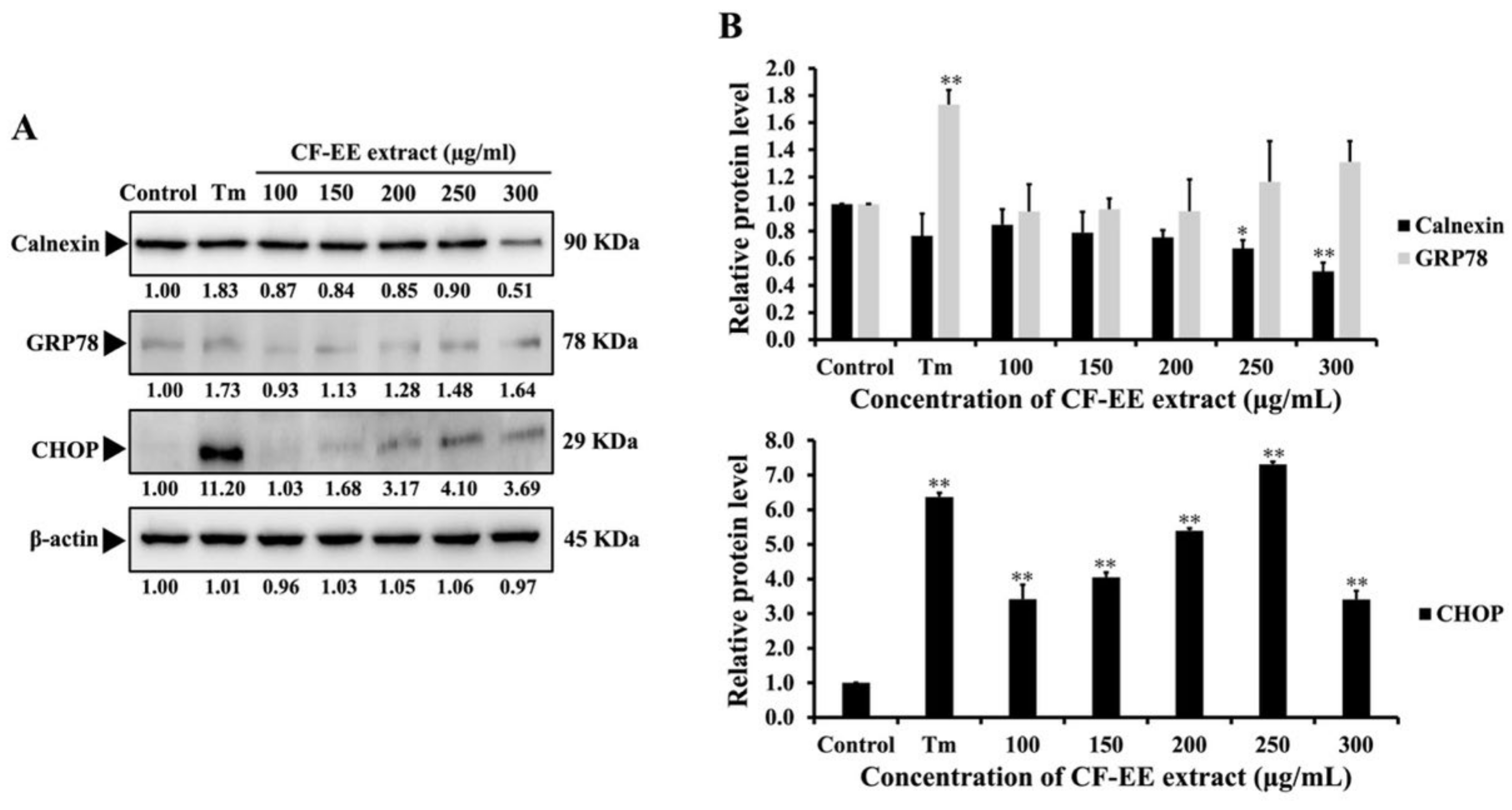
3.6. CF-EE Extract Induces Endoplasmic Reticulum Stress in HeLa Cells
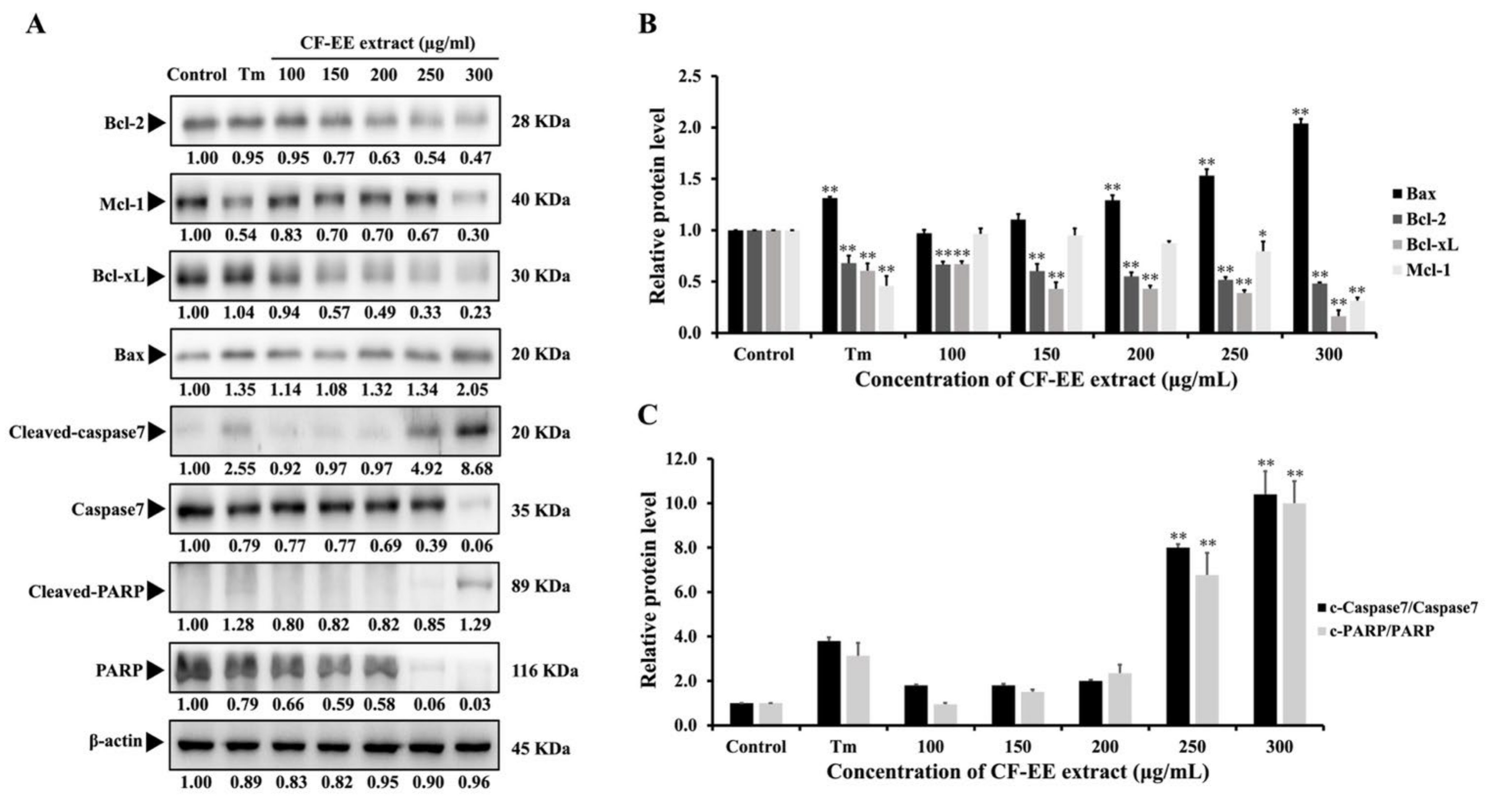
3.7. CF-EE Extract Activates Intrinsic Apoptotic Pathway in HeLa Cells
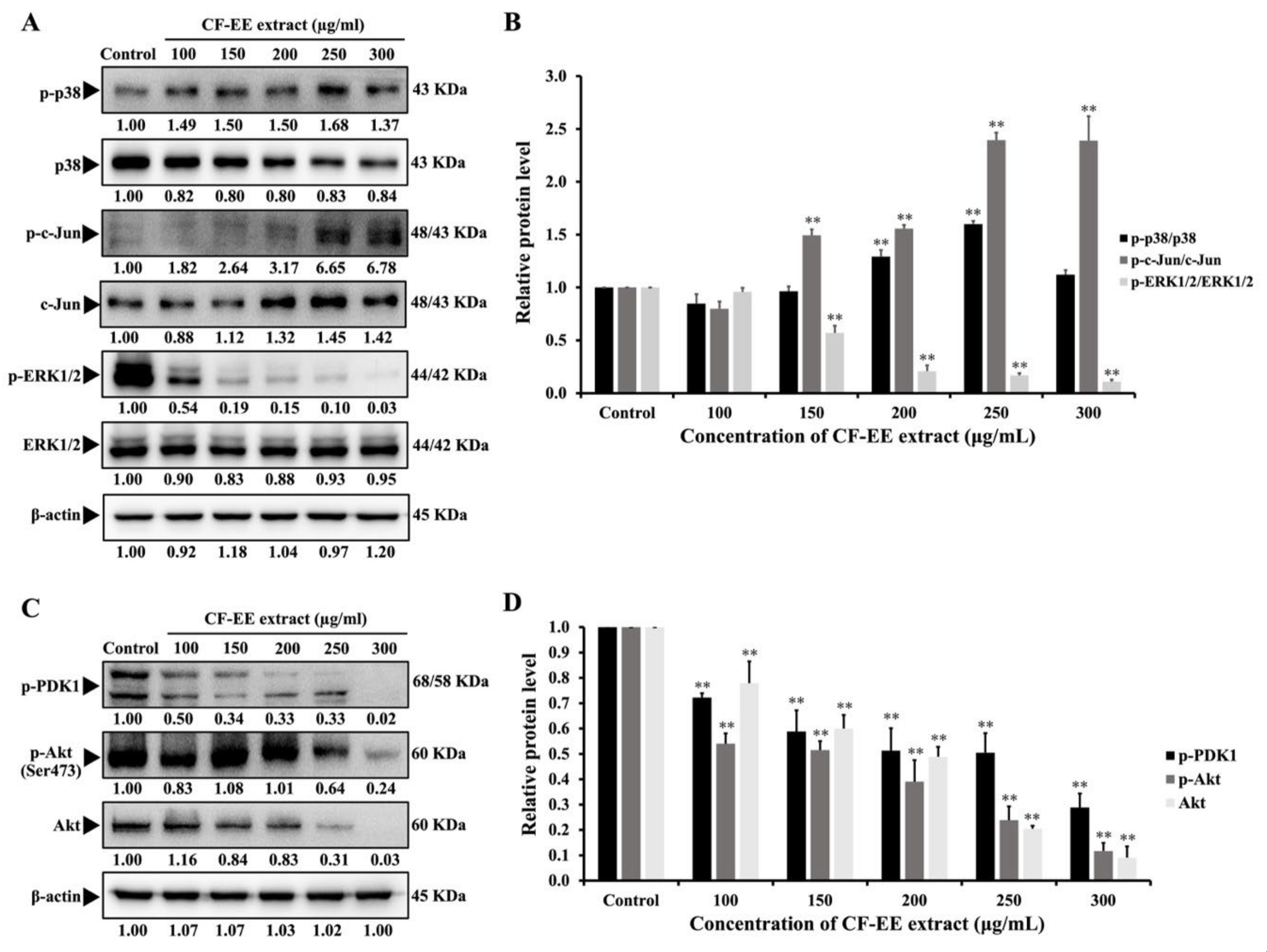
3.8. Effect of CF-EE Extract on MAPK and Akt Signaling Pathway in HeLa Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A. Cancer statistics for the year 2020: An overview. Int. J. Cancer. 2021, 149, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–2049. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Chen, H.D.; Yu, Y.; Li, N.; Chen, W. Changing profiles of cancer burden worldwide and in China: A secondary analysis of the global cancer statistics 2020. Chin. Med. J. 2021, 137, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Hrgovic, Z.; Fures, R.; Stanic, Z. The diagnostics and treatment of cervical cancer. Acta. Med. Croatica 2021, 75, 53–68. [Google Scholar]
- Park, S.; Kim, M.; Lee, S.; Jung, W.; Kim, B. Therapeutic Potential of Natural Products in Treatment of Cervical Cancer: A Review. Nutrients 2021, 13, 154. [Google Scholar] [CrossRef]
- Ahmed, I.; Lockhart, P.J.; Agoo, E.; Naing, K.W.; Nguyen, D.V.; Medhi, D.K. Evolutionary origins of taro (Colocasia esculenta) in Southeast Asia icle. Ecol. Evol. 2020, 10, 13530–13543. [Google Scholar] [CrossRef]
- Prajapati, R.; Kalariya, M.; Umbarkar, R.; Parmar, S.; Sheth, N. Colocasia esculenta: A potent indigenous plant. Int. J. Nutr. Pharmacol Nryrol Dis. 2011, 1, 90–96. [Google Scholar] [CrossRef]
- El-Mesallamya, A.M.; El-Tawilb, N.A.; Ibrahimb, S.A.; Husseinc, S.A. Phenolic Profile: Antimicrobial Activity and Antioxidant Capacity of Colocasia esculenta (L.) Schott. Egypt. J. Chem. 2021, 64, 2165–2172. [Google Scholar]
- Eleazu, C.O. Characterization of the natural products in cocoyam (Colocasia esculenta) using GC–MS. Pharm Biol. 2016, 54, 2880–2885. [Google Scholar] [CrossRef] [Green Version]
- Pawar, H.A.; Choudhary, P.D.; Kamat, S.R. An Overview of Traditionally Used Herb, Colocasia esculenta, as a Phytomedicine. Med. Aromat. Plants 2018, 7, 1000317. [Google Scholar] [CrossRef]
- Li, H.M.; Hwang, S.H.; Kang, B.G.; Hong, J.S.; Lim, S.S. Inhibitory Effects of Colocasia esculenta (L.) Schott Constituents on Aldose Reductase. Molecules 2014, 19, 13212–13224. [Google Scholar] [CrossRef] [Green Version]
- Lebot, V.; Lawac, F.; Michalet, S.; Legendre, L. Characterization of taro [Colocasia esculenta (L.) Schott] germplasm for improved flavonoid composition and content. Plant Genet. Resour. 2015, 15, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Ferreres, F.; Goncalves, R.F.; Gil-Izquierdo, A.; Valentao, V.; Silva, A.M.; Silva, J.B. Further knowledge on the phenolic profile of Colocasia esculenta (L.) Shott. J. Agric Food Chem. 2012, 60, 7005–7015. [Google Scholar] [CrossRef]
- Gupta, K.; Kumar, A.; Tomer, V.; Kumar, V.; Saini, M. Potential of Colocasia leaves in human nutrition: Review on nutritional and phytochemical properties. J. Food Biochem. 2019, 43, e12878. [Google Scholar] [CrossRef]
- Akuz, M. Determination of Antioxidant Activity of Ethanol Extract of Gölevez [(Colocasia esculenta (L.)] Tubers. KSU J. Agric Nat. 2019, 22, 388–394. [Google Scholar]
- Liua, H.; Laia, W.; Liua, X.; Yanga, H.; Fanga, Y.; Tiana, L. Exposure to copper oxide nanoparticles triggers oxidative stress and endoplasmic reticulum (ER)-stress induced toxicology and apoptosis in male rat liver and BRL-3A cell. J. Hazard Mater. 2021, 401, 123349. [Google Scholar] [CrossRef]
- Chea, J.; Lva, H.; Yang, J.; Zhaoa, B.; Zhoue, S.; Yua, T. Iron overload induces apoptosis of osteoblast cells via eliciting ER stress-mediated mitochondrial dysfunction and p-eIF2α/ATF4/CHOP pathway in vitro. Cell Signal. 2021, 84, 110024. [Google Scholar] [CrossRef]
- Lin, C.; Lee, C.; Chen, C.; Cheng, C.; Chen, P.; Ying, T. Protodioscin Induces Apoptosis Through ROS-Mediated Endoplasmic Reticulum Stress via the JNK/p38 Activation Pathways in Human Cervical Cancer Cells. Cell Physiol Biochem. 2018, 46, 322–334. [Google Scholar] [CrossRef]
- Martucciello, S.; Masullo, M.; Cerulli, A.; Piacente, S. Natural Products Targeting ER Stress, and the Functional Link to Mitochondria. Int. J. Mol Sci. 2020, 21, 1905. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Xu, S.; Gao, W.; Feng, J.; Zhao, G. Honokiol induces endoplasmic reticulum stress-mediated apoptosis in T human lung cancer cells. Life Sci. 2019, 221, 204–211. [Google Scholar] [CrossRef]
- Hengstermann, A.; Müller, T. Endoplasmic reticulum stress induced by aqueous extracts of cigarette smoke in 3T3 cells activates the unfolded-protein-response-dependent PERK pathway of cell survival. Free Radic Biol Med. 2018, 44, 1097–1107. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Wei, J.; Liu, R.; Hu, X.; Liang, T.; Zhou, Z.; Huang, Z. MAPK signaling pathway-targeted marine compounds in cancer therapy. J. Cancer Res. Clin. Oncol. 2021, 147, 3–22. [Google Scholar] [CrossRef]
- Cao, X.; Fu, M.; Bi, R.; Zheng, X.; Fu, B.; Tian, S. Cadmium induced BEAS-2B cells apoptosis and mitochondria damage via MAPK signaling pathway. Chemosphere 2021, 263, 128346. [Google Scholar] [CrossRef] [PubMed]
- Darling, N.J.; Cook, S.J. The role of MAPK signalling pathways in the response to endoplasmic reticulum stress. Biochim. Biophys. Acta 2014, 1843, 2150–2163. [Google Scholar] [CrossRef] [Green Version]
- Yua, J.; Lopez, J. Understanding MAPK Signaling Pathways in Apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef] [Green Version]
- Yangnok, K.; Innajak, S.; Sawasjirakij, R.; Mahabusarakam, W.; Watanapokasin, R. Effects of Artonin E on Cell Growth Inhibition and Apoptosis Induction in Colon Cancer LoVo and HCT116 Cells. Molecules 2022, 27, 2095. [Google Scholar] [CrossRef]
- Nur-Hadirah, K.; Arifullah, M.; Amaludin, N.A.; Klaiklay, S.; Chumkaew, P.; Zain, N.M. Total phenolic content and antioxidant activity of an edible Aroid, Colocasia esculenta (L.) Schott. IOP Conf Ser Earth Environ Sci. 2021, 756, 012044. [Google Scholar] [CrossRef]
- Gao, X.; Yanan, J.; Santhanam, R.K.; Wang, Y.; Lu, Y.; Zhang, M. Garlic flavonoids alleviate H2O2 induced oxidative damage in L02 cells and induced apoptosis in HepG2 cells by Bcl-2/Caspase pathway. J. Food Sci. 2021, 86, 366–375. [Google Scholar] [CrossRef]
- Kadar, N.N.; Ahmad, F.; Teoh, S.L.; Yahaya, M.F. Caffeic Acid on Metabolic Syndrome: A Review. Molecules 2021, 26, 5490. [Google Scholar] [CrossRef]
- Kępa, M.; Miklasińska-Majdanik, M.; Wojtyczka, R.D.; Idzik, D.; Korzeniowski, K.; Smoleń-Dzirba, J. Antimicrobial Potential of Caffeic Acid against Staphylococcus aureus Clinical Strains. BioMed Res. Int. 2018, 2018, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espindola, K.M.; Ferreira, R.G.; Mosquera, L.E.; Rosario, A.R.; Silva, A.M.; Silva, A.B. Chemical and Pharmacological Aspects of Caffeic Acid and Its Activity in Hepatocarcinoma. Front. Oncol. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilani-Jaziri, S.; Mokdad-Bzeouich, I.; Krifa, M.; Nasr, N.; Ghedira, K.; Chekir-Ghedira, L. Immunomodulatory and cellular anti-oxidant activities of caffeic, ferulic, and p-coumaric phenolic acids: A structure–activity relationship study. Drug Chem. Toxicol. 2017, 40, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Prasad, N.R.; Karthikeyan, A.; Karthikeyan, S.; Reddy, B.V. Inhibitory effect of caffeic acid on cancer cell proliferation by oxidative mechanism in human HT-1080 fibrosarcoma cell line. Mol. Cell Biochem. 2011, 439, 11–19. [Google Scholar] [CrossRef]
- Magnani, C.; Corrêa, M.A.; Isaac, V.; Salgado, H. Caffeic acid: A review of its potential use in medications and cosmetics. Anal. Methods 2014, 6, 3203. [Google Scholar] [CrossRef]
- Kanimozhi, G.; Prasad, N.R. Anticancer Effect of Caffeic Acid on Human Cervical Cancer Cells. Coffee Health Dis. Prev. 2015, 2015, 655–661. [Google Scholar]
- Smets, L.A. Programmed cell death (apoptosis) and response to anti-cancer drug. Anticancer Drugs 1994, 5, 3–9. [Google Scholar] [CrossRef]
- Hacker, G. The morphology of apoptosis. Cell Tissue Res. 2000, 301, 5–17. [Google Scholar] [CrossRef]
- Kim, J.K.; Kang, K.A.; Ryu, Y.S.; Piao, M.J.; Han, X.; Oh, M.C. Induction of Endoplasmic Reticulum Stress via Reactive Oxygen Species Mediated by Luteolin in Melanoma. Anticancer Res. 2016, 36, 2281–2289. [Google Scholar]
- Chiun, T.; Su, C.C. Tanshinone IIA increases protein expression levels of PERK, ATF6, IRE1α, CHOP, caspase-3 and caspase-12 in pancreatic cancer BxPC-3 cell-derived xenograft tumors. Mol. Med. Rep. 2017, 15, 3259–3263. [Google Scholar]
- Wang, X.; Zhuang, Y.; Fang, Y.; Cao, H.; Zhang, C.; Xing, C. Endoplasmic reticulum stress aggravates copper-induced apoptosis via the PERK/ATF4/CHOP signaling pathway in duck renal tubular epithelial cells. Environ. Pollut. 2020, 272, 115981. [Google Scholar] [CrossRef]
- Li, X.; Wang, D.; Sui, C.; Meng, F.; Sun, S.; Zheng, J. Oleandrin induces apoptosis via activating endoplasmic reticulum stress in breast cancer cells. Biomed. Pharmacother. 2020, 124, 109852. [Google Scholar] [CrossRef]
- Kamiya, T.; Nishihara, H.; Hara, H.; Adachi, T. Ethanol Extract of Brazilian Red Propolis Induces Apoptosis in Human Breast Cancer MCF-7 Cells through Endoplasmic Reticulum Stress. J. Agric. Food Chem. 2012, 60, 11065–11070. [Google Scholar] [CrossRef]
- Kadam, C.Y.; Abhang, S.A. Apoptosis Markers in Breast Cancer Therapy. Adv. Clin. Chem. 2016, 74, 143–193. [Google Scholar]
- Lamkanfi, M.; Kanneganti, T. Caspase-7: A protease involved in apoptosis and inflammation. Int. J. Biochem Cell Biol. 2012, 42, 21–24. [Google Scholar] [CrossRef] [Green Version]
- Gee, M.M.; Hyland, E.; Campiani, G.; Ramunno, A.; Nacci, V.; Zisterer, D.M. Caspase-3 is not essential for DNA fragmentation in MCF-7 cells during apoptosis induced by the pyrrolo-1,5-benzoxazepine, PBOX-6. FEBS Lett. 2002, 515, 66–70. [Google Scholar]
- Chang, W.; Hsieh, C.; Hsieo, M.; Li, W.; Hung, Y.; Ye, J. Caffeic acid induces apoptosis in human cervical cancer cells through the mitochondrial pathway. Taiwan J. Obstet. Gynecol. 2010, 49, 419–424. [Google Scholar] [CrossRef] [Green Version]
- Tseng, T.; Shen, C.; Huang, W.; Chen, C.; Liang, W.; Lin, T.; Kuo, H. Activation of neutral-sphingomyelinase, MAPKs, and p75 NTR-mediating caffeic acid phenethyl ester–induced apoptosis in C6 glioma cells. J. Biomed. Sci. 2014, 21, 61. [Google Scholar] [CrossRef] [Green Version]
- Inoshita, S.; Takeda, K.; Hatai, T.; Terada, Y.; Sano, M.; Hata, J. Phosphorylation and inactivation of myeloid cell leukemia 1 by JNK in response to oxidative stress. J. Biol. Chem. 2002, 277, 43730–43734. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Ichijo, H.; Korsmeyer, S.J. BCL-2 is phosphorylated and inactivated by an ASK1/Jun N-terminal protein kinase pathway normally activated at G(2)/M. Mol. Cell Biol. 1999, 19, 8469–8478. [Google Scholar] [CrossRef] [Green Version]
- Cai, B.; Chang, S.H.; Becker, E.B.; Bonni, A.; Xia, Z. p38 MAP Kinase Mediates Apoptosis through Phosphorylation of BimEL at Ser-65. J. Biol. Chem. 2006, 281, 25215–25222. [Google Scholar] [CrossRef] [Green Version]
- Hiraishi, N.; Kanmura, S.; Oda, K.; Arima, S.; Kumagai, K.; Mawatari, S. Extract of Lactobacillus plantarum strain 06CC2 induces JNK/p38 MAPK T pathway-mediated apoptosis through endoplasmic reticulum stress in Caco2 colorectal cancer cells. Biochem. Biophys. Rep. 2019, 20, 100691. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Mi, Q.; Hardwick, J.M.; Longo, D.L. Deletion of the loop region of Bcl-2 completely blocks paclitaxel-induced apoptosis. Proc. Natl. Acad. Sci. USA 1999, 96, 3775–3780. [Google Scholar] [CrossRef] [Green Version]
- Yao, Z.; Sun, B.; Hong, Q.; Yan, J.; Mu, D.; Li, J.; Sheng, H.; Guo, H. Pace4 regulates apoptosis in human prostate cancer cells via endoplasmic reticulum stress and mitochondrial signaling pathways. Drug Des. Devel. Ther. 2015, 9, 5911–5923. [Google Scholar]
- Lei, K.; Davis, R.J. JNK phosphorylation of Bim-related members of the Bcl2 family induces Bax-dependent apoptosis. Proc Natl. Acad Sci. USA 2003, 100, 2432–2437. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chomlamay, N.; Poorahong, W.; Innajak, S.; Watanapokasin, R. Apoptosis Induction Associated with Enhanced ER Stress Response and Up-Regulation of c-Jun/p38 MAPK Proteins in Human Cervical Cancer Cells by Colocasia esculenta var. aquatilis Hassk Extract. Sci. Pharm. 2022, 90, 45. https://doi.org/10.3390/scipharm90030045
Chomlamay N, Poorahong W, Innajak S, Watanapokasin R. Apoptosis Induction Associated with Enhanced ER Stress Response and Up-Regulation of c-Jun/p38 MAPK Proteins in Human Cervical Cancer Cells by Colocasia esculenta var. aquatilis Hassk Extract. Scientia Pharmaceutica. 2022; 90(3):45. https://doi.org/10.3390/scipharm90030045
Chicago/Turabian StyleChomlamay, Natharika, Watcharaporn Poorahong, Sukanda Innajak, and Ramida Watanapokasin. 2022. "Apoptosis Induction Associated with Enhanced ER Stress Response and Up-Regulation of c-Jun/p38 MAPK Proteins in Human Cervical Cancer Cells by Colocasia esculenta var. aquatilis Hassk Extract" Scientia Pharmaceutica 90, no. 3: 45. https://doi.org/10.3390/scipharm90030045
APA StyleChomlamay, N., Poorahong, W., Innajak, S., & Watanapokasin, R. (2022). Apoptosis Induction Associated with Enhanced ER Stress Response and Up-Regulation of c-Jun/p38 MAPK Proteins in Human Cervical Cancer Cells by Colocasia esculenta var. aquatilis Hassk Extract. Scientia Pharmaceutica, 90(3), 45. https://doi.org/10.3390/scipharm90030045