In Vitro Comparison of the Anti-Proliferative Effects of Galenia africana on Human Skin Cell Lines


Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Galenia africana Plant Extract
2.2. Cell Lines and Cell Culture
2.3. Determination of Cell Proliferation and Viability
2.4. Trypan Blue Exclusion Assay
2.5. Determination of Cytotoxicity
2.6. Nuclear Morphological Changes
2.7. Statistical Analysis
3. Results
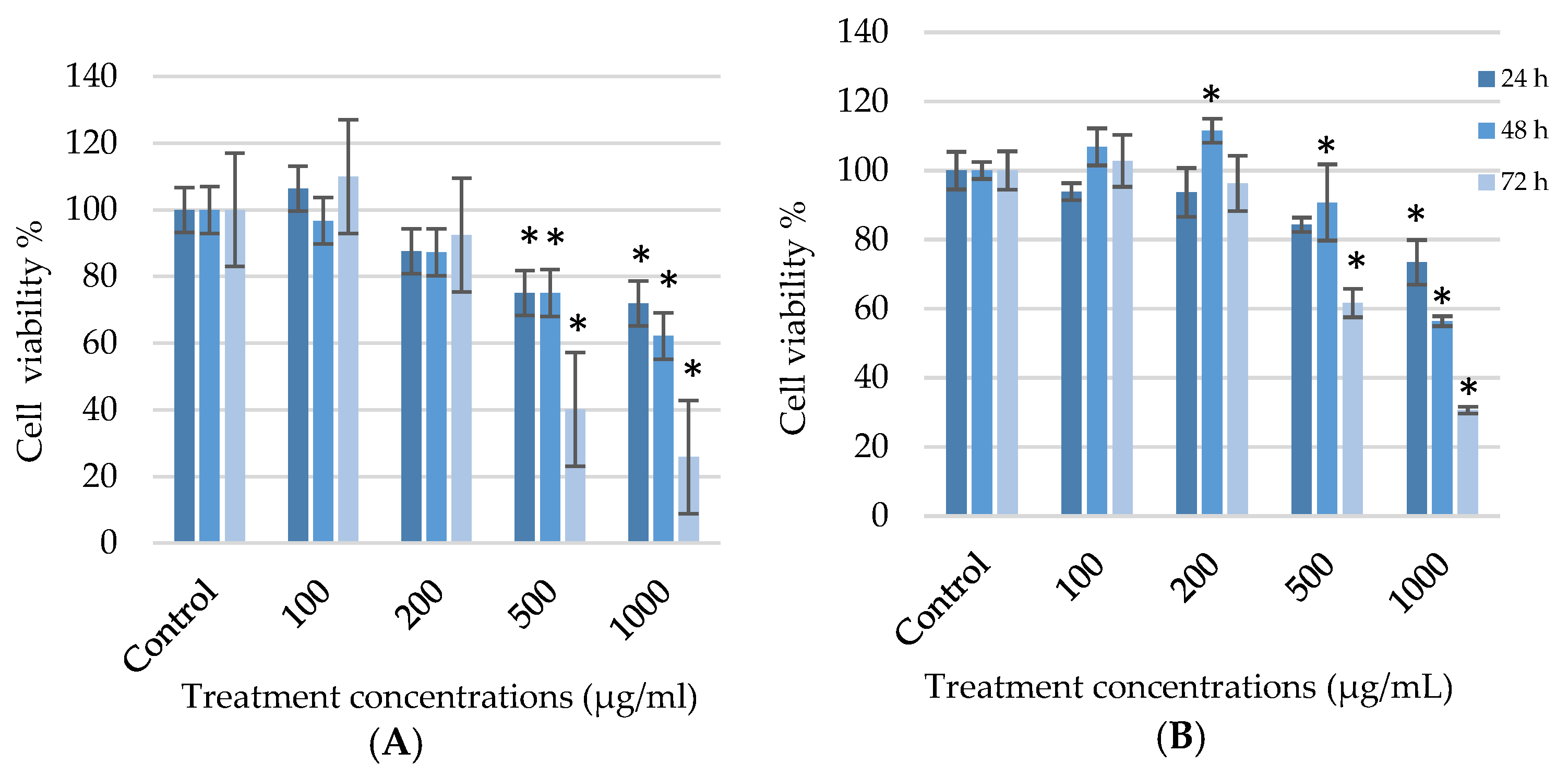
3.1. Effect of Galenia africana on Cell Viability
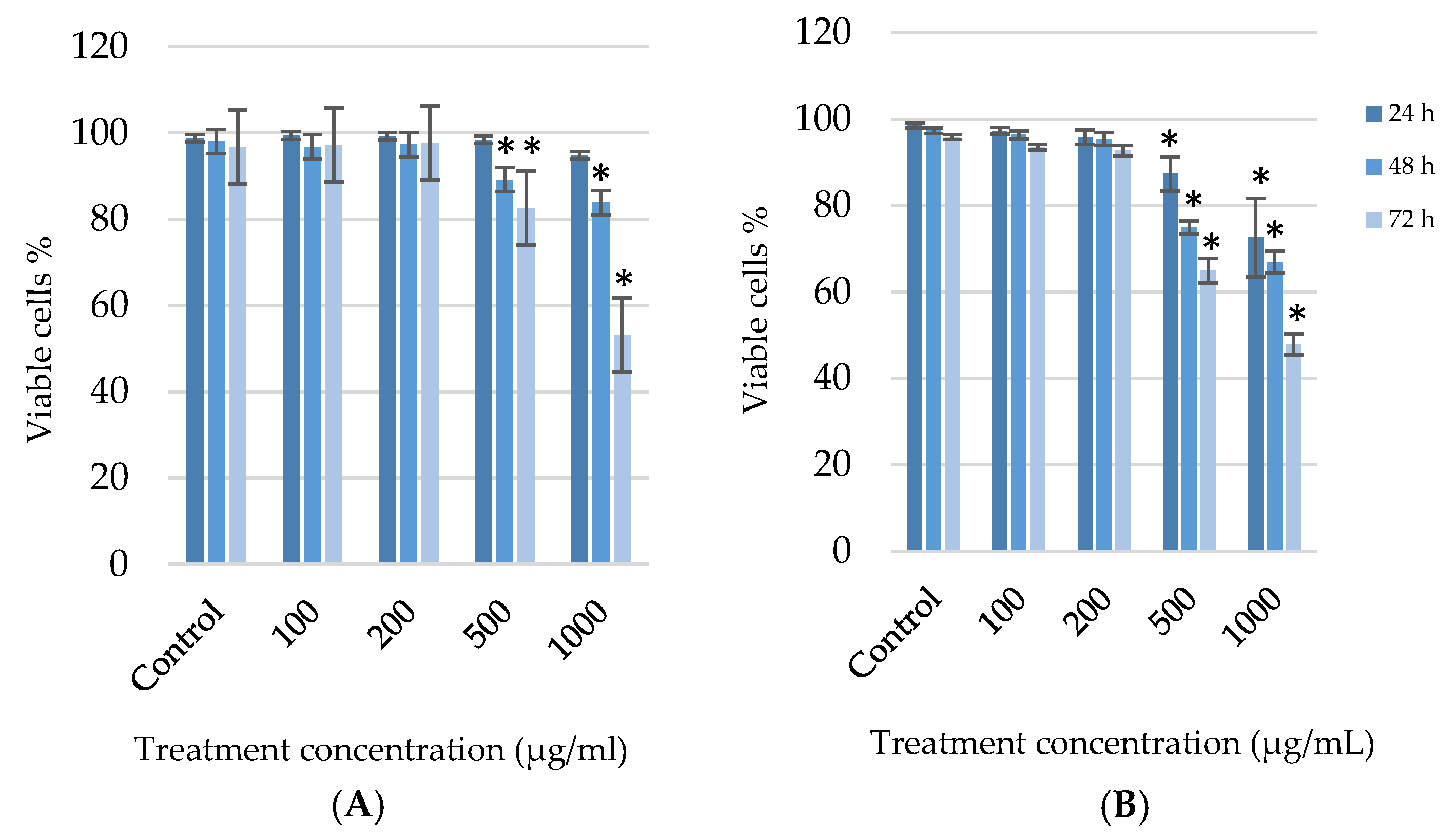
3.2. Potential Cytotoxic Effect of G. africana
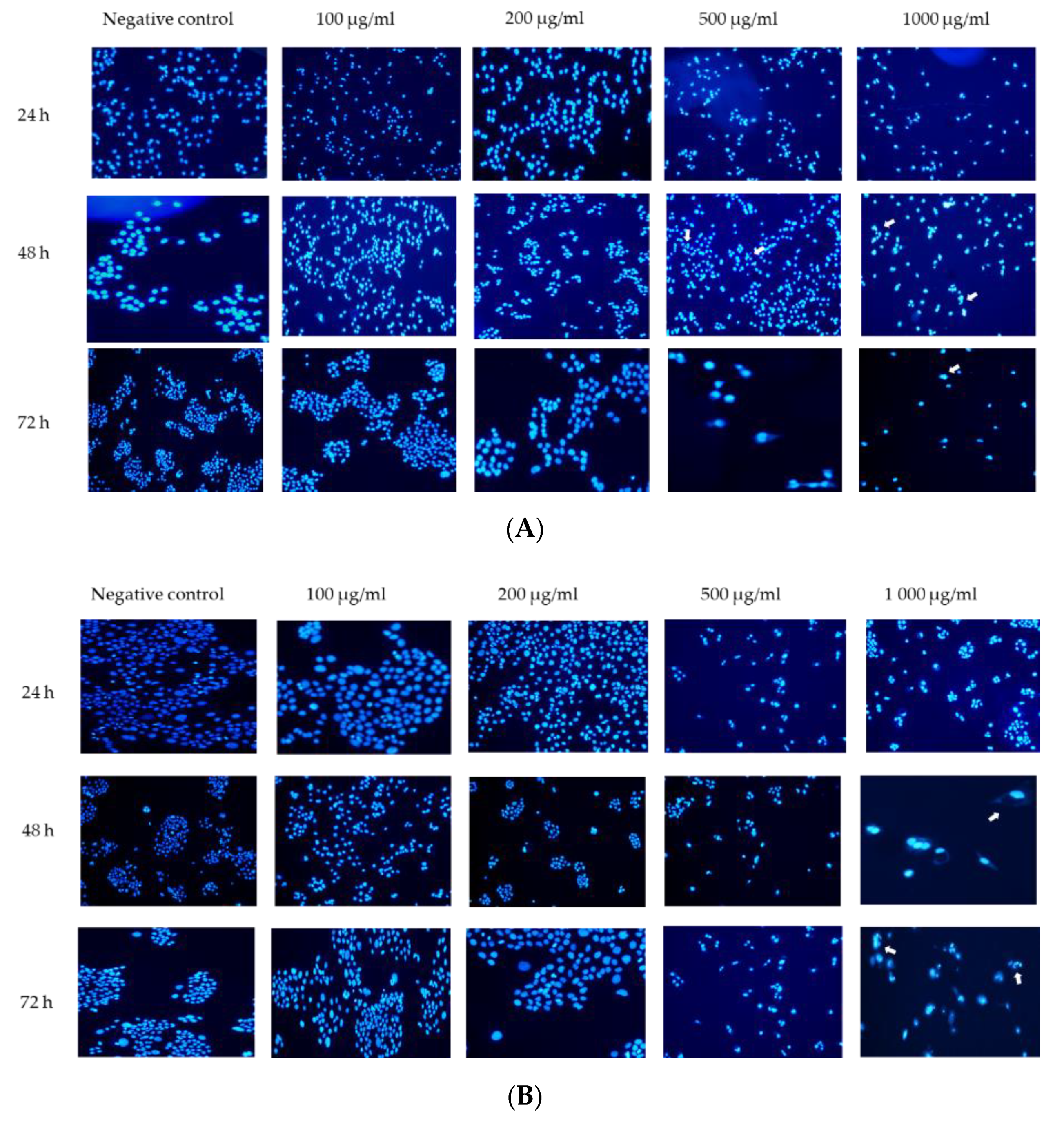
3.3. Effects of G. africana on Nuclei Morphology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gelzo, M.; Granato, G.; Albano, F.; Arcucci, A.; Russo, A.D.; De Vendittis, E.; Ruocco, M.R.; Corso, G. Evaluation of cytotoxic effects of 7-dehydrocholesterol on melanoma cells. Free Radic. Biol. Med. 2014, 70, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Carr, S.; Smith, C.; Wernberg, J. Epidemiology and Risk Factors of Melanoma. Surg. Clin. 2020, 100, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Wang, Z.; Liu, Y.; Peng, X.; Liu, Y.; Zhu, S.; Zhang, Z.; Qiu, Y.; Jin, M.; Wang, R. Oxyfadichalcone C inhibits melanoma A375 cell proliferation and metastasis via suppressing PI3K/Akt and MAPK/ERK pathways. Life Sci. 2018, 206, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Abildgaard, C.; Guldberg, P. Molecular drivers of cellular metabolic reprogramming in melanoma. Trends. Mol. Med. 2015, 21, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Rawson, R.V.; Scolyer, R.A. From Breslow to BRAF and immunotherapy: Evolving concepts in melanoma pathogenesis and disease progression and their implications for changing management over the last 50 years. Hum. Pathol. 2020, 95, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Agar, N.; Young, A.R. Melanogenesis: A photoprotective response to DNA damage? Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2005, 571, 121–132. [Google Scholar] [CrossRef]
- Prakash, O.; Kumar, A.; Kumar, P. Anticancer potential of plants and natural products. Am. J. Pharmacol. Sci. 2013, 1, 104–115. [Google Scholar] [CrossRef]
- Force, J.; Salama, A.K. First-line treatment of metastatic melanoma: Role of nivolumab. Immuno Targets Ther. 2017, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Niezgoda, A.; Niezgoda, P.; Czajkowski, R. Novel approaches to treatment of advanced melanoma: A review on targeted therapy and immunotherapy. BioMed. Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Shilkrut, M.; Zhao, Z.; Li, M.; Batty, N.; Barber, B. Autoimmune comorbidities in patients with metastatic melanoma: A retrospective analysis of us claims data. BMC Cancer 2018, 18, 145. [Google Scholar] [CrossRef] [PubMed]
- Linardou, H.; Gogas, H. Toxicity management of immunotherapy for patients with metastatic melanoma. Ann. Transl. Med. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Olver, I.; Carey, M.; Boyes, A.; Hall, A.; Noble, N.; Bryant, J.; Walsh, J.; Sanson-Fisher, R. The timeliness of patients reporting the side effects of chemotherapy. Support. Care Cancer 2018, 26, 3579–3586. [Google Scholar] [CrossRef]
- Oun, R.; Moussa, Y.E.; Wheate, N.J. The side effects of platinum-based chemotherapy drugs: A review for chemists. Dalton Trans. 2018, 47, 6645–6653. [Google Scholar] [CrossRef]
- Gaweł-Bęben, K.; Osika, P.; Asakawa, Y.; Antosiewicz, B.; Głowniak, K.; Ludwiczuk, A. Evaluation of anti-melanoma and tyrosinase inhibitory properties of marchantin A, a natural macrocyclic bisbibenzyl isolated from Marchantia species. Phytochem. Lett. 2019, 31, 192–195. [Google Scholar] [CrossRef]
- De Silva, M.B.; Tencomnao, T. The protective effect of some Thai plants and their bioactive compounds in UV light-induced skin carcinogenesis. J. Photochem. Photobiol. B 2018, 185, 80–89. [Google Scholar] [CrossRef]
- Khosravi, R.A.; Shokri, H.; Farahnejat, Z.; Chalangari, R.; Katalin, M. Antimycotic efficacy of Iranian medicinal plants towards dermatophytes obtained from patients with dermatophytosis. Chin. J. Nat. Med. 2013, 11, 43–48. [Google Scholar] [CrossRef]
- George, V.C.; Kumar, D.N.; Suresh, P.; Kumar, R.A. Oleanolic acid inhibits cell growth and induces apoptosis in A375 melanoma cells. Biomed. Prev. Nutr. 2014, 4, 95–99. [Google Scholar] [CrossRef]
- Watt, J.M.; Breyer-Brandwijk, M.G. The Medicinal and Poisonous Plants of Southern and Eastern Africa being an Account of their Medicinal and other Uses, Chemical Composition, Pharmacological Effects and Toxicology in Man and Animal. In The Medicinal and Poisonous Plants of Southern and Eastern Africa Being an Account of their Medicinal and Other Uses, Chemical Composition, Pharmacological Effects and Toxicology in Man and Animal; E. & S. Livingstone Ltd.: Edinburgh, Scotland, 1962. [Google Scholar]
- De Beer, J.J.; Van Wyk, B.-E. An ethnobotanical survey of the Agter–Hantam, Northern Cape Province, South Africa. S. Afr. J. Bot. 2011, 77, 741–754. [Google Scholar] [CrossRef]
- Mativandlela, S.P.; Muthivhi, T.; Kikuchi, H.; Oshima, Y.; Hamilton, C.; Hussein, A.A.; van der Walt, M.L.; Houghton, P.J.; Lall, N. Antimycobacterial flavonoids from the leaf extract of Galenia africana. J. Nat. Prod. 2009, 72, 2169–2171. [Google Scholar] [CrossRef] [PubMed]
- Ticha, L.A.; Klaasen, J.A.; Green, I.R.; Naidoo, S.; Baker, B.; Pietersen, R.-D. Phytochemical and Antimicrobial Screening of Flavanones and Chalcones from Galenia africana and Dicerothamnus rhinocerotis. Nat. Prod. Commun. 2015, 10, 1934578X1501000713. [Google Scholar] [CrossRef]
- Panche, A.; Diwan, A.; Chandra, S. Flavonoids: An overview. J. Nutr. Sci. 2016, 5. [Google Scholar] [CrossRef]
- Raffa, D.; Maggio, B.; Raimondi, M.V.; Plescia, F.; Daidone, G. Recent discoveries of anticancer flavonoids. Eur. J. Med. Chem. 2017, 142, 213–228. [Google Scholar] [CrossRef]
- Vries, F.; El Bitar, H.; Green, I.; Klaasen, J.; Mabulesa, W.; Bodo, B.; Johnson, Q. An antifungal active extract from the aerial parts of Galenia africana. In Proceedings of the 11th NAPRECA, Antananarivo, Madagascar, 9–12 August 2005; pp. 123–131. [Google Scholar]
- Li, J.; Li, X.; Chen, Z.; Rasul, A.; Zhao, C.; Millimouno, F.M.; Tsuji, I.; Yamamura, T.; Iqbal, R.; Malhi, M. Antiproliferative and apoptotic effects of pinocembrin in human prostate cancer cells. Bangladesh J. Pharmacol. 2013, 8, 255–262. [Google Scholar]
- Rasul, A.; Millimouno, F.M.; Ali Eltayb, W.; Ali, M.; Li, J.; Li, X. Pinocembrin: A novel natural compound with versatile pharmacological and biological activities. Biomed. Res. Int. 2013, 2013. [Google Scholar] [CrossRef]
- Ng’uni, T. Medicinal Uses of Galenia africana: A Study of the Antimicrobial, Antifungal and Anticancer Properties. University of Western Cape: Bellville, South Africa, Unpublished work. 2017. [Google Scholar]
- Alley, M.C.; Scudiero, D.A.; Monks, A.; Hursey, M.L.; Czerwinski, M.J.; Fine, D.L.; Abbott, B.J.; Mayo, J.G.; Shoemaker, R.H.; Boyd, M.R. Feasibility of drug screening with panels of human tumor cell lines using a microculture tetrazolium assay. Cancer Res. 1988, 48, 589–601. [Google Scholar] [PubMed]
- Fotakis, G.; Timbrell, J.A. In vitro cytotoxicity assays: Comparison of LDH, neutral red, MTT and protein assay in hepatoma cell lines following exposure to cadmium chloride. Toxicol. Lett. 2006, 160, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Ng’uni, T.; Klaasen, J.A.; Fielding, B.C. Acute toxicity studies of the South African medicinal plant Galenia africana. Toxicol. Rep. 2018, 5, 813–818. [Google Scholar] [CrossRef]
- Van Wyk, B.-E.; De Wet, H.; Van Heerden, F. An ethnobotanical survey of medicinal plants in the Southeastern Karoo, South Africa. S. Afr. J. Bot. 2008, 74, 696–704. [Google Scholar] [CrossRef]
- Ghosh, S.; Bishayee, K.; Khuda-Bukhsh, A.R. Oleanolic acid isolated from ethanolic extract of Phytolacca decandra induces apoptosis in A375 skin melanoma cells: Drug-DNA interaction and signaling cascade. J. Integr. Med. 2014, 12, 102–114. [Google Scholar] [CrossRef]
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Flavonoids in cancer and apoptosis. Cancers 2019, 11, 28. [Google Scholar] [CrossRef]
- Mohamed, L.; Chakraborty, S.; ArulJothi, K.N.; Mabasa, L.; Sayah, K.; Costa-Lotufo, L.V.; Jardine, A.; Prince, S. Galenia africana plant extract exhibits cytotoxicity in breast cancer cells by inducing multiple programmed cell death pathways. Saudi Pharm. J. 2020, 28, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Mativandlela, S.P.N.; Meyer, J.J.M.; Hussein, A.A.; Houghton, P.J.; Hamilton, C.J.; Lall, N. Activity against Mycobacterium smegmatis and M. tuberculosis by extract of South African medicinal plants. Phytother. Res. 2008, 22, 841–845. [Google Scholar] [CrossRef]
- Powis, G. Dose-dependent metabolism, therapeutic effect, and toxicity of anticancer drugs in man. Drug Metab. Rev. 1983, 14, 1145–1163. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, A.-S.H.; Mohammed, A.S.; Abdullah, R.; Mirghani, M.E.S.; Al-Qubaisi, M. Cytotoxic effects of Mangifera indica L. kernel extract on human breast cancer (MCF-7 and MDA-MB-231 cell lines) and bioactive constituents in the crude extract. BMC Compl. Altern. Med. 2014, 14, 199. [Google Scholar] [CrossRef]
- Kadan, S.; Saad, B.; Sasson, Y.; Zaid, H. In vitro evaluations of cytotoxicity of eight antidiabetic medicinal plants and their effect on GLUT4 translocation. Evid. Based Complement. Altern. Med. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Specian, A.F.L.; Serpeloni, J.M.; Tuttis, K.; Ribeiro, D.L.; Cilião, H.L.; Varanda, E.A.; Sannomiya, M.; Martinez-Lopez, W.; Vilegas, W.; Cólus, I.M. LDH, proliferation curves and cell cycle analysis are the most suitable assays to identify and characterize new phytotherapeutic compounds. Cytotechnology 2016, 68, 2729–2744. [Google Scholar] [CrossRef]
- Barry, M.-J.J. Compositions Derived from Galenia Africana and Methods of Use for Cancer Treatment. U.S. Patents 20190365840A1, 12 December 2019. [Google Scholar]
- Pang, W.; Liu, S.; He, F.; Li, X.; Saira, B.; Zheng, T.; Chen, J.; Dong, K.; Pei, X.-F. Anticancer activities of Zanthoxylum bungeanum seed oil on malignant melanoma. J. Ethnopharmacol. 2019, 229, 180–189. [Google Scholar] [CrossRef]
- Ravishankar, D.; Rajora, A.K.; Greco, F.; Osborn, H.M. Flavonoids as prospective compounds for anti-cancer therapy. Int. J. Biochem. Cell Biol. 2013, 45, 2821–2831. [Google Scholar] [CrossRef]
- Drira, R.; Sakamoto, K. Sakuranetin induces melanogenesis in B16BL6 melanoma cells through inhibition of ERK and PI3K/AKT signaling pathways. Phytother. Res. 2016, 30, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, K.; Wu, Y.; Chen, Y.; Chen, X.; Hu, C.W.; Hu, F. Pinocembrin induces ER stress mediated apoptosis and suppresses autophagy in melanoma cells. Cancer Lett. 2018, 431, 31–42. [Google Scholar] [CrossRef]
- Shi, H.; Wu, Y.; Wang, Y.; Zhou, M.; Yan, S.; Chen, Z.; Gu, D.; Cai, Y. Liquiritigenin potentiates the inhibitory effects of cisplatin on invasion and metastasis via downregulation MMP-2/9 and PI3 K/AKT signaling pathway in B16F10 melanoma cells and mice model. Nutr. Cancer 2015, 67, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Lu, J.; Liu, Y.; Meng, Q.; Xie, J.; Wang, Z.; Teng, L. Liquiritigenin induces tumor cell death through mitogen-activated protein kinase-(MPAKs-) mediated pathway in hepatocellular carcinoma cells. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reduction in Cell Viability (IC50 Value ± SD, µg/mL) | ||
|---|---|---|
| Exposure Times (h) | A375 | HaCaT |
| 24 | 1438.68 ± 0.09 | 1893.89 ± 0.02 |
| 48 | 1258.98 ± 0.077 | 1102.21 ± 0.006 |
| 72 | 572.70 ± 0.06 | 711.04 ± 0.055 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndlovu, B.; De Kock, M.; Klaasen, J.; Rahiman, F. In Vitro Comparison of the Anti-Proliferative Effects of Galenia africana on Human Skin Cell Lines. Sci. Pharm. 2021, 89, 12. https://doi.org/10.3390/scipharm89010012
Ndlovu B, De Kock M, Klaasen J, Rahiman F. In Vitro Comparison of the Anti-Proliferative Effects of Galenia africana on Human Skin Cell Lines. Scientia Pharmaceutica. 2021; 89(1):12. https://doi.org/10.3390/scipharm89010012
Chicago/Turabian StyleNdlovu, Banele, Maryna De Kock, Jeremy Klaasen, and Farzana Rahiman. 2021. "In Vitro Comparison of the Anti-Proliferative Effects of Galenia africana on Human Skin Cell Lines" Scientia Pharmaceutica 89, no. 1: 12. https://doi.org/10.3390/scipharm89010012
APA StyleNdlovu, B., De Kock, M., Klaasen, J., & Rahiman, F. (2021). In Vitro Comparison of the Anti-Proliferative Effects of Galenia africana on Human Skin Cell Lines. Scientia Pharmaceutica, 89(1), 12. https://doi.org/10.3390/scipharm89010012