Cell-Based Methods for Determination of Efficacy for Candidate Therapeutics in the Clinical Management of Cancer



Abstract
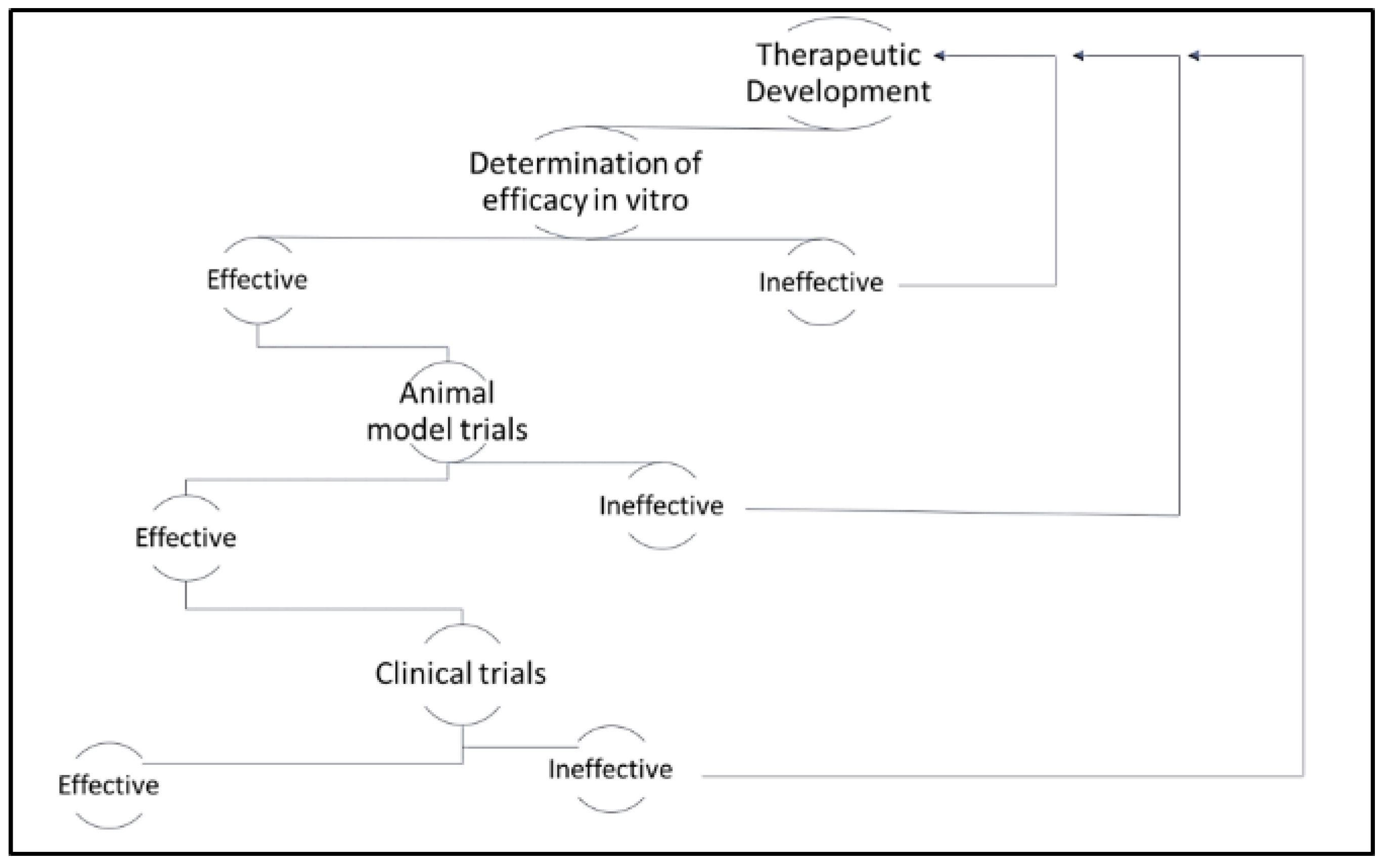
1. Introduction
2. Cell-Based Assays for Determination of Therapeutic Efficacy
2.1. Cell Viability and Proliferation Assays
2.2. Colorimetric Assays
2.3. Binding Assays
Neutral Red Uptake
2.4. ATP Production
2.5. Colony Formation Assays
2.6. Cytotoxicity Assays
2.6.1. Staining and Imaging
2.6.2. LDH Cytotoxicity Assay
2.7. Cell Apoptosis Assays
2.7.1. DNA Fragmentation
2.7.2. Caspase Activation
2.7.3. Flow Cytometry
2.8. Cell Cycle Arrest Assays
2.9. 3D Cell Culture Systems
2.9.1. Multicellular Tumor Spheroids
2.9.2. Hollow Fibre Assays
2.10. Cell-Based Systems for Evaluating Combinatorial Efficacy
3. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bradbury, R.H. Overview. Cancer 2007, 1, 1–17. [Google Scholar]
- World Health Organisation Global Health Observatory (GHO) Data: Top 10 Causes of Death. 2017, pp. 1–3. Available online: http://www.who.int/gho/mortality_burden_disease/causes_death/top_10/en/ (accessed on 18 December 2017).
- Joudeh, J.; Allen, J.; Das, A.; Prabhu, V. Impact of Genetic Targets on Cancer Therapy; El-Deiry, W.S., Ed.; Springer: New York, NY, USA, 2013; ISBN 9781461461753. [Google Scholar]
- NCI SEER data. Five-Year Relative Survival. Available online: https://seer.cancer.gov/seerstat/WebHelp/Relative_Survival.htm (accessed on 12 December 2017).
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2017. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Mocellin, S.; Shrager, J.; Scolyer, R.; Pasquali, S.; Verdi, D.; Marincola, F.M.; Briarava, M.; Gobbel, R.; Rossi, C.; Nitti, D. Targeted Therapy Database (TTD): A Model to Match Patient’s Molecular Profile with Current Knowledge on Cancer Biology. PLoS ONE 2010, 5, e11965. [Google Scholar] [CrossRef] [PubMed]
- American Cancer Society. Signs and Symptoms of Cancer. Available online: https://www.cancer.org/cancer/cancer-basics/signs-and-symptoms-of-cancer.html (accessed on 5 January 2018).
- Goodspeed, A.; Heiser, L.M.; Gray, J.W.; Costello, J.C. Tumor-Derived Cell Lines as Molecular Models of Cancer Pharmacogenomics. Mol. Cancer Res. 2016, 14, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Gillet, J.-P.; Varma, S.; Gottesman, M.M. The Clinical Relevance of Cancer Cell Lines. JNCI J. Natl. Cancer Inst. 2013, 105, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.V.; Haber, D.A.; Settleman, J. Cell line-based platforms to evaluate the therapeutic efficacy of candidate anticancer agents. Nat. Rev. Cancer 2010, 10, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Tanimoto, A.; Yamada, T.; Nanjo, S.; Takeuchi, S. Receptor ligand-triggered resistance to alectinib and its circumvention by Hsp90 inhibition in EML4-ALK lung cancer cells. Oncotarget 2014, 5, 4920–4928. [Google Scholar] [CrossRef] [PubMed]
- Miyawaki, M.; Yasuda, H.; Tani, T.; Hamamoto, J.; Arai, D.; Ishioka, K.; Ohgino, K.; Nukaga, S.; Hirano, T.; Kawada, I.; et al. Overcoming EGFR Bypass Signal-Induced Acquired Resistance to ALK Tyrosine Kinase Inhibitors in ALK-Translocated Lung Cancer. Mol. Cancer Res. 2017, 15, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Guan, S.; Cao, W.; Wang, H.; Chen, Z.; Zhao, Y.; Yu, Y.; Zhang, H.; Pang, J.C.; et al. Novel ALK inhibitor AZD3463 inhibits neuroblastoma growth by overcoming crizotinib resistance and inducing apoptosis. Sci. Rep. 2016, 6, 19423. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cheng, B.F.; Yang, H.J.; Wang, M.; Feng, Z.W. NF-κB protects human neuroblastoma cells from nitric oxide-induced apoptosis through upregulating biglycan. Am. J. Transl. Res. 2015, 7, 1541–1552. [Google Scholar] [PubMed]
- Xu, F.; Li, H.; Sun, Y. Inhibition of Axl improves the targeted therapy against ALK-mutated neuroblastoma. Biochem. Biophys. Res. Commun. 2014, 454, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Guan, S.; Zhao, Y.; Yu, Y.; Woodfield, S.E.; Zhang, H.; Yang, K.L.; Bieerkehazhi, S.; Qi, L.; Li, X.; et al. The second-generation ALK inhibitor alectinib effectively induces apoptosis in human neuroblastoma cells and inhibits tumor growth in a TH-MYCN transgenic neuroblastoma mouse model. Cancer Lett. 2017, 400, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Borowicz, S.; Van Scoyk, M.; Avasarala, S.; Karuppusamy Rathinam, M.K.; Tauler, J.; Bikkavilli, R.K.; Winn, R.A. The Soft Agar Colony Formation Assay. J. Vis. Exp. 2014, 92, 51998. [Google Scholar] [CrossRef] [PubMed]
- McGowan, E.M.; Alling, N.; Jackson, E.A.; Yagoub, D.; Haass, N.K.; Allen, J.D.; Martinello-Wilks, R. Evaluation of cell cycle arrest in estrogen responsive MCF-7 breast cancer cells: Pitfalls of the MTS assay. PLoS ONE 2011, 6, e20623. [Google Scholar] [CrossRef] [PubMed]
- Chapter 4: Cell viability. Promega Protocols and Applications Guide; Promega: Madison, WI, USA, 2011; Revision 3/11; pp. 1–23. Available online: www.promega.com (accessed on 22 January 2018).
- Stoddart, M.J.; Louis, K.S.; Siegel, A.C.; Kupcsik, L.; Czekanska, E.M. Chapters 1-5: Mammalian Cell Viability; Stoddart, M.J., Ed.; Humana Press: Totowa, NJ, USA, 2011; Volume 740, pp. 1–27. ISBN 978-1-61779-107-9. [Google Scholar]
- Yang, N.; Ray, S.D.; Krafts, K. Cell Proliferation. Encycl. Toxicol. 2014, 1, 761–765. [Google Scholar] [CrossRef]
- Prabst, K.; Engelhardt, H.; Ringgeler, S.; Hubner, H.; Ates, G.; Vanhaeke, T.; Rogiers, V.; Rodrigues, R.; Chan, L.L.-Y.; McCulley, K.J.; et al. Chapters 1-4, 8: Cell Viability Assays; Gilbert, D., Friedrich, O., Eds.; Humana Press: Totowa, NJ, USA, 2017; Volume 1601, pp. 1–43, 89–97. ISBN 978-1-4939-6959-3. [Google Scholar]
- van Meerlo, J.; Kaspers, G.J.L.; Cloos, J.; Glaysher, S.; Cree, I.A.; Hartley, J.M.; Spanswick, V.J.; Hartley, J.A. Chapters 20-22, 25: Cancer Cell Culture Methods and Protocols, 2nd ed.; Cree, I.A., Ed.; Humana Press: Totowa, NJ, USA, 2013; Volume 731, pp. 237–259, 309–321. ISBN 978-1-84973-379-3. [Google Scholar]
- Blumenthal, R.D. Chapter 1: Chemosensitivity; Blumenthal, R.D., Ed.; Humana Press: Totowa, NJ, USA, 2005; Volume 1, pp. 3–21. ISBN 9780874216561. [Google Scholar]
- Pozarowski, P.; Huang, X.; Gong, R.W.; Priebe, W.; Darzynkiewicz, Z. Simple, semiautomatic assay of cytostatic and cytotoxic effects of antitumor drugs by laser scanning cytometry: Effects of the bis-intercalator WP631 on growth and cell cycle of T-24 cells. Cytometry 2004, 57A, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Kunze, D.; Wuttig, D.; Füssel, S.; Meye, A.; Wirth, M.P. Sirna-Mediated Inhibition of Antiapoptotic Genes in Human Bladder Cancer Cells. Eur. Urol. Suppl. 2006, 5, 800. [Google Scholar] [CrossRef]
- Bhola, N.E.; Balko, J.M.; Duggar, T.C.; Kuba, M.G.; Sanchez, V.; Sanders, V.; Sanders, M.; Stanford, J.; Cook, R.S.; Arteaga, C.L. TGF-β Inhibition enhances chemotherapy action against triple-negative breast cancer. J. Clin. Investig. 2013, 123, 1348–1358. [Google Scholar] [CrossRef] [PubMed]
- Pagliacci, M.C.; Spinozzi, F.; Migliorati, G.; Fumi, G.; Smacchia, M.; Grignani, F.; Riccardi, C.; Nicoletti, I. Genistein Inhibits Tumour Cell Growth in vitro but Enhances Mitochondrial Reduction of Tetrazolium Salts: A Further Pitfall in the Use of the MTT Assay for Evaluating Cell Growth and Survival. Eur. J. Cancer 1993, 3030A, 1573–1577. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Brady, A.J.; Kearney, P.; Tunney, M.M. Comparative evaluation of 2,3-bis [2-methyloxy-4-nitro-5-sulfophenyl]-2H-tetrazolium-5-carboxanilide (XTT) and 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2, 4-disulfophenyl)-2H-tetrazolium, monosodium salt (WST-8) rapid colorimetric assays for antim. J. Microbiol. Methods 2007, 71, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Uzunoglu, S.; Karaca, B.; Atmaca, H.; Kisim, A.; Sezgin, C.; Karabulut, B.; Uslu, R. Comparison of XTT and Alamar blue assays in the assessment of the viability of various human cancer cell lines by AT-101 (-/-gossypol). Toxicol. Mech. Methods 2010, 20, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Riss, T.; Moravec, R.; Niles, A.; Duellman, S. Cell Viability Assays. In Assay Guidance Manual; Springer: Berlin, Germany, 2013; pp. 1–55. ISBN 978-1-4939-6959-3. [Google Scholar]
- Cell Counting Kit-8 Technical Manual; Dojindo Molecules Technology Inc.: Rockville, MD, USA, 2009; Volume 8, pp. 1–7. Available online: www.dojindo.com (accessed on 12 February 2018).
- Griffiths, G.D.; Lindsay, C.D.; Upshall, D.G. Examination of the toxicity of several protein toxins of plant origin using bovine pulmonary endothelial cells. Toxicology 1994, 90, 11–27. [Google Scholar] [CrossRef]
- Fotakis, G.; Timbrell, J.A. In vitro cytotoxicity assays: Comparison of LDH, neutral red, MTT and protein assay in hepatoma cell lines following exposure to cadmium chloride. Toxicol. Lett. 2006, 160, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Perez, M.G.; Fourcade, L.; Mateescu, M.A.; Paquin, J. Neutral Red versus MTT assay of cell viability in the presence of copper compounds. Anal. Biochem. 2017, 535, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Mothana, R.A.A.; Kriegisch, S.; Harms, M.; Wende, K.; Lindequist, U. Assessment of selected Yemeni medicinal plants for their in vitro antimicrobial, anticancer, and antioxidant activities. Pharm. Biol. 2011, 49, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, S.; Wahab, N.; Zainal Abidin, N.; Manickam, S.; Zakaria, Z. Growth inhibition of human gynecologic and colon cancer cells by Phyllanthus watsonii through apoptosis induction. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Borenfreund, E.; Puerner, J.A.; York, N. A Simple Quantitative Procedure Using Monolayer Cultures for Cytotoxicity Assays (HTD/NR-90). J. Tissue Cult. Methods 1984, 9, 7–9. [Google Scholar] [CrossRef]
- Chiba, K.; Kawakami, K.; Tohyama, K. Simultaneous Evaluation of Cell Viability by Neutral Red, MTT and Crystal Violet Staining Assays of the Same Cells. Toxicol. Vitr. 1998, 12, 251–258. [Google Scholar] [CrossRef]
- Nemes, Z.; Dietz, R.; Luth, J.B. The Pharmacological Relevance of Vital Staining with Neutral Red. Experientia 1979, 35, 1475–1476. [Google Scholar] [CrossRef] [PubMed]
- Soo, J.S.S.; Ng, C.H.; Tan, S.H.; Malik, R.A.; Teh, Y.C.; Tan, B.S.; Ho, G.F.; See, M.H.; Taib, N.A.M.; Yip, C.H.; et al. Metformin synergizes 5-fluorouracil, epirubicin, and cyclophosphamide (FEC) combination therapy through impairing intracellular ATP production and DNA repair in breast cancer stem cells. Apoptosis 2015, 20, 1373–1387. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Chan, D.K.; Miskimins, W. Preventing feedback activation of glycolytic ATP production enhances metformin cytotoxicity in breast cancer cells when oxidative phosphorylation is inhibited. Cancer Metab. 2014, 2, 89. [Google Scholar] [CrossRef]
- Distelmaier, F.; Valsecchi, F.; Liemburg-Apers, D.C.; Lebiedzinska, M.; Rodenburg, R.J.; Heil, S.; Keijer, J.; Fransen, J.; Imamura, H.; Danhauser, K.; et al. Mitochondrial dysfunction in primary human fibroblasts triggers an adaptive cell survival program that requires AMPK-α. Biochim. Biophys. Acta-Mol. Basis Dis. 2015, 1852, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, P.; Ruh, N.; Harnoss, J.M.; Kiss, J.; Mollenhauer, M.; Scherr, A.L.; Platzer, L.K.; Schmidt, T.; Podar, K.; Opferman, J.T.; et al. Prolyl hydroxylase 3 attenuates MCL-1-mediated ATP production to suppress the metastatic potential of colorectal cancer cells. Cancer Res. 2016, 76, 2219–2230. [Google Scholar] [CrossRef] [PubMed]
- ATP Determination Kit (A22066); Invitrogen Molecular Probes Inc.: Eugene, OR, USA, 2005; pp. 1–3. Available online: http://www.thermofisher.com/order/catalog/product/A22066?CID=AFLBC-A22066 (accessed on 26 February 2018).
- Paciello, L.; Falco, F.C.; Parascandola, P. Determination of yeast cell viability: Viable count vs ATP-based bioluminescence assay. J. Biotechnol. 2010, 150, 386–387. [Google Scholar] [CrossRef]
- Wannlund, J.C. Analytical applications of firefly luciferase. TrAC Trends Anal. Chem. 1983, 2, 7–9. [Google Scholar] [CrossRef]
- Saotome, K.; Morita, H.; Umeda, M. Cytotoxicity test with simplified crystal violet staining method using microtitre plates and its application to injection drugs. Toxicol. Vitr. 1989, 3, 317–321. [Google Scholar] [CrossRef]
- Horibata, S.; Vo, T.V.; Subramanian, V.; Thompson, P.R.; Coonrod, S.A. Utilization of the Soft Agar Colony Formation Assay to Identify Inhibitors of Tumorigenicity in Breast Cancer Cells. J. Vis. Exp. 2015, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Liao, T.T.; Jia, R.W.; Shi, Y.L.; Jia, J.W.; Wang, L.; Chua, H. Propidium iodide staining method for testing the cytotoxicity of 2,4,6-trichlorophenol and perfluorooctane sulfonate at low concentrations with Vero cells. J. Environ. Sci. Heal.-Part A Toxic/Hazardous Subst. Environ. Eng. 2011, 46, 1769–1775. [Google Scholar] [CrossRef] [PubMed]
- Cincin, Z.B.; Unlu, M.; Kiran, B.; Bireller, E.S.; Cakmakoglu, B. Apoptotic Effects of Quercitrin on DLD-1 Colon Cancer Cell Line. Pathol. Oncol. Res. 2015, 21, 333–338. [Google Scholar] [CrossRef] [PubMed]
- De Nicola, M.; Gattia, D.M.; Bellucci, S.; De Bellis, G.; Micciulla, F.; Pastore, R.; Tiberia, A.; Cerella, C.; D’Alessio, M.; Antisari, M.V.; et al. Effect of different carbon nanotubes on cell viability and proliferation. J. Phys. Condens. Matter 2007, 19. [Google Scholar] [CrossRef]
- Loo, D.T.; Darzynkiewicz, Z.; Zhao, H. Chapter 1 & 8: DNA Damage Detection In Situ, Ex Vivo, and In Vivo; Humana Press: Totowa, NJ, USA, 2011; Volume 682, ISBN 978-1-60327-408-1. [Google Scholar]
- Matsuzaki, T.; Takeshi, S.; Fujikura, K.; Takata, K. Nuclear Staining for Laser Confocal Microscopy. Acta Histochem. Cytochem. 1997, 30, 309–314. [Google Scholar] [CrossRef]
- Boyd, V.; Cholewa, O.; Papas, K. Limitations in the use of fluorescein diacetate/propidium iodide (FDA/PI) and cell permeable nucleic acid stains for viability measurements of isolated islets of Langerhans. Curr. Trends Biotechnol. Pharm. 2008, 2, 66–84. [Google Scholar] [PubMed]
- Smith, S.M.; Wunder, M.B.; Norris, D.A.; Shellman, Y.G. A simple protocol for using a LDH-Based cytotoxicity assay to assess the effects of death and growth inhibition at the same time. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Dorstyn, L.; Wlodkowic, D.; Skommer, J.; Darzynkiewicz, Z.; Tait, S.W.G.; Bouchier-Hayes, L.; Oberst, A.; Connell, S.; Green, D. Chapters 1-3: Apoptosis Methods and Protocols, 2nd ed.; Erhardt, P., Toth, A., Eds.; Humana Press: Totowa, NJ, USA, 2009; Volume 559, pp. 3–33. ISBN 978-1-60327-016-8. [Google Scholar]
- Ribeiro, T.M.; Bertolla, R.P.; Spaine, D.M.; Fraietta, R.; Ortiz, V.; Cedenho, A.P. Sperm nuclear apoptotic DNA fragmentation in men with testicular cancer. Fertil. Steril. 2008, 90, 1782–1786. [Google Scholar] [CrossRef] [PubMed]
- Nath, M.; Vats, M.; Roy, P. Tri- and diorganotin(IV) complexes of biologically important orotic acid: Synthesis, spectroscopic studies, in vitro anti-cancer, DNA fragmentation, enzyme assays and in vivo anti-inflammatory activities. Eur. J. Med. Chem. 2013, 59, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Ragheb, A.; Mahfouz, R.; Ghoneim, I.; Sharma, R.; Agarwal, A.; Sabanegh, E. Pretreatment Sperm DNA Fragmentation Index (DFI) in Cancer Patients and Its Relationship To Postcryopreservation Motile Sperm Concentration (PCMSC) and Sperm Motility (PCSM). J. Urol. 2010, 183, e750. [Google Scholar] [CrossRef]
- Matassov, D.; Kagan, T.; Leblanc, J.; Sikorska, M.; Zakeri, Z.; Kumar, S.; Williams, O. Chapters 1-3: Apoptosis Methods and Protocols; Brady, H., Ed.; Humana Press: Totowa, NJ, USA, 2004; Volume 282, ISBN 1617794899. [Google Scholar]
- Ostling, O.; Johansen, K.L. Microelectrophoretic study of radiation-induced DNA damages in individual mammalian cells. Biochem. Biophys. Res. Commun. 1984, 123, 291–298. [Google Scholar] [CrossRef]
- Olive, P.L.; Banáth, J.P.; Durand, R.E.; Banath, J.P. Heterogeneity in Radiation-Induced DNA Damage and Repair in Tumor and Normal Cells Measured Using the “Comet” Assay. Radiat. Res. 1990, 122, 86. [Google Scholar] [CrossRef] [PubMed]
- Olive, P.L.; Banáth, J.P. The comet assay: A method to measure DNA damage in individual cells. Nat. Protoc. 2006, 1, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Gorczyca, W.; Gong, J.; Darzynkiewicz, Z. Detection of DNA strand breaks in individual apoptic cells by the in situ terminal deoxynucleotidyltransferse and nick translation assays. Cancer Res. 1993, 53, 1945–1951. [Google Scholar] [PubMed]
- Gavrieli, Y.; Sherman, Y.; Ben-Sasson, S.A. Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell Biol. 1992, 119, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Negoescu, A.; Lorimier, P.; Labat-Moleur, F.; Drouet, C.; Robert, C.; Guillermet, C.; Brambilla, C.; Brambilla, E. In Situ Apoptotic Cell Labeling by the TUNEL Method: Improvement and Evaluation on Cell Preparations. J. Histochem. Cytochem. 1996, 4, 959–968. [Google Scholar] [CrossRef]
- O’Brien, I.E.W.; Ferguson, I.B.; Holdaway, K.M. Annexin-V and TUNEL Use in Monitoring the Progression of Apoptosis in Plants. Cytometry 1997, 29, 28–33. [Google Scholar] [CrossRef]
- Jeong, M.; Paek, A.R.; Seung, J.; Youp, C.; Chae, K.; Hyang, J.; Hyun, S.; Jin, H. Regulation of cancer cell death by a novel compound, C604, in a c-Myc-overexpressing cellular environment. Eur. J. Pharmacol. 2015, 769, 257–265. [Google Scholar] [CrossRef]
- Katsumi, Y.; Iehara, T.; Miyachi, M.; Yagyu, S.; Tsubai-shimizu, S.; Kikuchi, K.; Tamura, S.; Kuwahara, Y.; Tsuchiya, K.; Kuroda, H.; et al. Sensitivity of malignant rhabdoid tumor cell lines to PD 0332991 is inversely correlated with p16 expression. Biochem. Biophys. Res. Commun. 2011, 413, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, T.; Yang, P.C. Western Blot: Technique, Theory, and Trouble Shooting. N. Am. J. Med. Sci. 2012, 4, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, E.; Atmaca, H.; Kisim, A.; Uzunoglu, S. Effects of Thymus serpyllum Extract on Cell Proliferation, Apoptosis and Epigenetic Events in Human Breast Cancer Cells. Nutr. Cancer 2012, 64, 1245–1250. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Sakaguchi, M.; Yoneyama, H.; Usami, Y.; Harusawa, S. Biochemical and Biophysical Research Communications Histamine H 3 receptor antagonist OUP-186 attenuates the proliferation of cultured human breast cancer cell lines. Biochem. Biophys. Res. Commun. 2016, 480, 479–485. [Google Scholar] [CrossRef] [PubMed]
- McStay, G.P.; Green, D.R. Measuring apoptosis: Caspase inhibitors and activity assays. Cold Spring Harb. Protoc. 2014, 2014, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Zhang, Q.-J.; Liu, Y.-Y. Chapter 11: Drug Design and Discovery; Satyanarayanajois, S.D., Ed.; Humana Press: Totowa, NJ, USA, 2011; Volume 716, pp. 179–193. ISBN 978-1-61779-011-9. [Google Scholar]
- Gottliner, C.; Mechtold, B.; Radbruch, A. Chapter 1: Flow Cytometry and Cell Sorting, 2nd ed.; Radbruch, A., Ed.; Springer: Berlin, Germany, 2000; pp. 1–26. ISBN 9783642084928. [Google Scholar]
- Sigma Aldrich. Antibodies for Apoptosis. Available online: https://www.sigmaaldrich.com/life-science/cell-biology/antibodies/antibody-products.html?TablePage=13832047 (accessed on 10 September 2018).
- Wlodkowic, D.; Skommer, J.; Darzynkiewicz, Z. Flow cytometry-based apoptosi detection. Methods Mol. Biol. 2009, 559, 19–32. [Google Scholar] [CrossRef]
- Dai, Y.; Grant, S. Methods to Study Cancer Therapeutic Drugs That Target Cell Cycle Checkpoints. In Cell Cycle Checkpoints; Springer: Berlin, Germany, 2011; Volume 782, pp. 257–304. ISBN 978-1-61779-272-4. [Google Scholar]
- Mayhew, C.; Bosco, E.; Solomon, D.; Knudsen, E.; Angus, S.; Berndt, N. Checkpoint Controls and Cancer, 2nd ed.; Schonthal, A.H., Ed.; Humana Press: Totowa, NJ, USA, 2004; Volume 280, pp. 301–311. ISBN 1-59259-788-2. [Google Scholar]
- Huang, B.W.; Gao, J.Q. Application of 3D cultured multicellular spheroid tumor models in tumor-targeted drug delivery system research. J. Control. Release 2018, 270, 246–259. [Google Scholar] [CrossRef] [PubMed]
- Griffith, L.G.; Swartz, M.A. Capturing complex 3D tissue physiology in vitro. Nat. Rev. Mol. Cell Biol. 2006, 7, 211. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.M.; Cukierman, E. Modeling Tissue Morphogenesis and Cancer in 3D. Cell 2007, 130, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Cozzolino, A.M.; Noce, V.; Battistelli, C.; Marchetti, A.; Grassi, G.; Cicchini, C.; Tripodi, M.; Amicone, L. Modulating the Substrate Stiffness to Manipulate Differentiation of Resident Liver Stem Cells and to Improve the Differentiation State of Hepatocytes. Stem Cells Int. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.Y.; Kenny, P.A.; Lee, E.H.; Bissell, M.J. Three-dimensional culture models of normal and malignant breast epithelial cells. Nat. Methods 2007, 4, 359. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Tang, T. Engineering 3D approaches to model the dynamic microenvironments of cancer bone metastasis. Bone Res. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Loessner, D.; Holzapfel, B.M.; Clements, J.A. Engineered microenvironments provide new insights into ovarian and prostate cancer progression and drug responses. Adv. Drug Deliv. Rev. 2014, 79, 193–213. [Google Scholar] [CrossRef] [PubMed]
- Mehta, G.; Hsiao, A.Y.; Ingram, M.; Luker, G.D.; Takayama, S. Opportunities and challenges for use of tumor spheroids as models to test drug delivery and efficacy. J. Control. Release 2012, 164, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Suggitt, M.; Cooper, P.A.; Shnyder, S.D.; Bibby, M.C. The hollow fibre model - Facilitating anti-cancer pre-clinical pharmacodynamics and improving animal welfare. Int. J. Oncol. 2006, 29, 1493–1499. [Google Scholar] [CrossRef] [PubMed]
- Decker, S.; Hollingshead, M.; Bonomi, C.A.; Carter, J.P.; Sausville, E.A. The hollow fibre model in cancer drug screening: The NCI experience. Eur. J. Cancer 2004, 40, 821–826. [Google Scholar] [CrossRef] [PubMed]
- Hall, L.; Krauthauser, C.; Wexler, R.; Hollingshead, M.; Slee, A.; Kerr, J. The hollow fiber assay: continued characterization with novel approaches. Anticancer Res. 2000, 20, 903–911. [Google Scholar] [PubMed]
- Lehár, J.; Krueger, A.S.; Avery, W.; Heilbut, A.M.; Johansen, L.M.; Price, E.R.; Rickles, R.J.; Short, G.F.; Staunton, J.E.; Jin, X.; et al. Synergistic drug combinations tend to improve therapeutically relevant selectivity. Nat. Biotechnol. 2009, 27, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Neve, R.M.; Chin, K.; Fridlyand, J.; Yeh, J.; Baehner, F.L.; Fevr, T.; Clark, L.; Bayani, N.; Coppe, J.P.; Tong, F.; et al. A collection of breast cancer cell lines for the study of functionally distinct cancer subtypes. Cancer Cell 2006, 10, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Sos, M.L.; Michel, K.; Zander, T.; Weiss, J.; Frommolt, P.; Peifer, M.; Li, D.; Ullrich, R.; Koker, M.; Fischer, F.; et al. Predicting drug susceptibility of non-small cell lung cancers based on genetic lesions. J. Clin. Invest. 2009, 119, 1727–1740. [Google Scholar] [CrossRef] [PubMed]
- McDermott, U.; Sharma, S.V.; Dowell, L.; Greninger, P.; Montagut, C.; Lamb, J.; Archibald, H.; Raudales, R.; Tam, A.; Lee, D.; et al. Identification of genotype-correlated sensitivity to selective kinase inhibitors by using high-throughput tumor cell line profiling. Proc. Natl. Acad. Sci. USA 2007, 104, 19936–19941. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Assay | Real-Time Measurements | Sensitivity | Multiplexing | Water-Soluble | Additional Intermediates Not Required | Ease-of-Use | Cost |
|---|---|---|---|---|---|---|---|
| MTT | χ | √ | χ | χ | √ | √ | √ |
| MTS | χ | √ | χ | √ | χ | √√ | √ |
| XTT | χ | √ | χ | √ | χ | √√ | √ |
| WST-1 | χ | √ | χ | √ | χ | √√ | √ |
| WST-8 | √ | √√ | √ | √ | √ (Already incorporated) | √√√ | √ |
| Resazurin | √ | √√ | √ | √ | √ | √√ | √√ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gordon, J.L.; Brown, M.A.; Reynolds, M.M. Cell-Based Methods for Determination of Efficacy for Candidate Therapeutics in the Clinical Management of Cancer. Diseases 2018, 6, 85. https://doi.org/10.3390/diseases6040085
Gordon JL, Brown MA, Reynolds MM. Cell-Based Methods for Determination of Efficacy for Candidate Therapeutics in the Clinical Management of Cancer. Diseases. 2018; 6(4):85. https://doi.org/10.3390/diseases6040085
Chicago/Turabian StyleGordon, Jenna L., Mark A. Brown, and Melissa M. Reynolds. 2018. "Cell-Based Methods for Determination of Efficacy for Candidate Therapeutics in the Clinical Management of Cancer" Diseases 6, no. 4: 85. https://doi.org/10.3390/diseases6040085
APA StyleGordon, J. L., Brown, M. A., & Reynolds, M. M. (2018). Cell-Based Methods for Determination of Efficacy for Candidate Therapeutics in the Clinical Management of Cancer. Diseases, 6(4), 85. https://doi.org/10.3390/diseases6040085